Abstract
Glutaredoxins (GRXs) are small-molecular-weight proteins present in a wide range of organisms, and they play a key role in maintaining the redox homeostasis of cells. Most studies on GRXs are conducted in animals and humans, and those conducted on plants are scarce. The number and types of GRX genes vary in different plants. According to the active sites, the GRX family can be further divided into the CPYC, CGFS, and CC subfamilies. The CPYC and CGFS subfamilies are present in eukaryotes. The CC subfamily is exclusively present in higher plants and has the highest number of genes. In this study, 85 GRX genes were identified in common wheat (Triticum aestivum L.) using a bioinformatic method, wherein 12, 9, and 64 belonged to the CPYC, CGFS, and CC subfamilies, respectively. All TaGRX genes were homogeneously distributed in the three subgenomes of wheat. The gene structure analysis revealed that TaGRX members had 1–7 introns. The conserved motif analysis revealed that members of the same TaGRX subfamily had similar motifs. An analysis of cis-regulatory elements of promoters demonstrated that most TaGRX members had auxin-responsive elements; cis-regulatory elements, such as methyl jasmonate (MeJA), MYB, and abscisic acid (ABA), were distributed in all subfamilies, and the cell-cycle regulation element was only observed in the CC and CPYC subfamily members. Additionally, the synteny of the GRX genes in wheat, in wheat and Arabidopsis, and in wheat and barley was analyzed to clarify the evolutionary correlation of TaGRXs. The expression characteristics of TaGRXs were investigated, and TaGRX expression in various tissues and its responses to different abiotic stresses were preliminarily determined. This study provides a reference for the functional analysis of TaGRXs and understanding their role in molecular breeding of wheat.
1. Introduction
Reactive oxygen species (ROS) are one of the byproducts of aerobic metabolism in plant cells and mainly consist of a superoxide anion (O2−), hydrogen peroxide (H2O2), and hydroxyl radical (-OH). Under normal growth conditions, the generation and removal of ROS are in dynamic balance in plant cells and do not harm the plant itself. ROS at low concentrations are important signaling molecules in the normal growth process of plants [1]. However, when plants are in an adverse state, their redox homeostasis is unbalanced, and ROS accumulation increases. This leads to oxidative stress and membrane peroxidation, breaking the integrity of the cell membrane and disrupting the physiological and biochemical metabolism of plant cells, which ultimately causes the growth inhibition and even death of the plant [2]. Hence, it is very important to maintain redox homeostasis in cells and reduce the oxidative damage caused by ROS accumulation. Both enzymatic and nonenzymatic mechanisms play roles in maintaining redox homeostasis in plants. Among them, the enzymatic system mainly includes superoxide dismutase (SOD), peroxidase (POD), catalase (CAT), and ascorbate peroxidase (APX). The nonenzymatic system is mainly ascorbic acid–glutathione system (AsA–GSH system). In addition, plants have evolved more complex thioredoxin and glutaredoxin systems, which regulate the redox state of proteins and maintain their normal physiological functions [3]. The thioredoxin system comprises triphosphopyridine nucleotide (NADPH), thioredoxin (TRX), and thioredoxin reductase (TR); the glutaredoxin system comprises NADPH, glutathione reductase (GR), glutaredoxins (GRXs), and glutathione (GSH).
GRX is a TRX-like redox enzyme commonly present in organisms and restores growth in TRX-deficient mutants of Escherichia coli [4]. The molecular weight of GRX proteins is generally small. GRX proteins possess a typical active site (CxxC or CxxS) [5], which is exposed on the surface of the protein in 3D space and has a nearby GSH binding site. In the GRX system, electrons are transferred from NADPH to GR, then to GSH, and finally to GRXs, which use electrons to reduce disulfide bonds in the target protein. Therefore, GRXs can mediate the reversible reduction in target proteins in the presence of GSH, thus affecting the stability, correct folding, activity, and function of the protein molecules. This, in turn, maintains and regulates the cellular redox state and redox-dependent signaling pathways [6]. According to the amino acid sequences of the active sites, the GRX genes can be divided into the CPYC, CGFS, and CC types, possessing active sites such as C[P/G/S][Y/F][C/S], strictly conserved CGFS sequences, and CC[M/L][C/S/G/A/T], respectively [7]. The CPYC and CGFS subfamily members are widely present in nature, whereas the CC subfamily members are exclusively present in higher plants.
GRXs regulate plant growth. ROXY1, a CC subfamily gene of Arabidopsis, is an essential gene for petal development. It is involved in the initiation process of flower primordium and petal morphology; therefore, the roxy1 mutant in Arabidopsis exhibits abnormalities in the petal primordium and reduced number of petals [8]. OsROXY1 and OsROXY2, the homologs in rice, exhibit floral expression patterns similar to those of ROXY1 in Arabidopsis, and OsROXY1 and OsROXY2 can complement the floral phenotype of the roxy1 mutant in Arabidopsis [9]. The involvement of GRXs in the regulation of organ development may be related to the activation of specific transcription factors. For example, the interaction between the nucleus-localized ROXY and the TGA transcription factor is required for petal development [10], and plants lacking TGA9 and TGA10 exhibit defects in male gamete development, which is similar to that observed in roxy1 and roxy2 double mutants [11]. Additionally, the interaction between ROXY1 and PETAL LOSS (PTL) in Arabidopsis limits intra- and inter-sepal growth but promotes petal initiation [12]. OsGRXC2.2 is a member of CPYC subfamily. It is related to embryonic development, and its overexpression suppresses the expression of endodermal marker genes, thereby interfering with the normal development of rice embryos [13]. The overexpression of OsGRX_C7 can increase rice grain yield and promote plant growth [14]. PHS9 is a member of CC subfamily. It plays a key role in the regulation of pre-harvest sprouting by integrating ROS and ABA signals [15]. GRXS17 is a member of CGFS subfamily in Arabidopsis. It plays an important role in temperature-dependent postembryonic growth by regulating auxin responses [16]. In the grxs17 mutant of Arabidopsis, auxin sensitivity and polar transport are inhibited; however, its auxin biosynthesis is not affected; ROS accumulates at high levels, and cell membrane is damaged at high temperature. Additionally, the grxs17 mutant is sensitive to high temperature and long sunlight cycles, which results in leaf elongation, smaller stem tip meristem, altered photoperiod, and delayed sedge [17].
GRXs are involved in the responses of plants to biotic and abiotic stresses. Compared with wild-type plants, rice with OsGRXS15 overexpression exhibited increased resistance to Xanthomonas oryzae pv. Oryzae and Fusarium, whereas the resistance of the OsGRXS15-knockout mutant decreased [18]. OsGRX20 overexpression enhanced resistance of rice to X. oryzae pv.oryzae and Fusarium, whereas plants with the RNAi of this gene exhibited reduced resistance [19]. These results suggest that OsGRXS15 and OsGRX20 positively regulate disease resistance in rice. Silencing of SlGRX1, a CGFS subfamily gene in tomato, resulted in increased sensitivity of plants to salt stress. However, the overexpression of this gene in Arabidopsis significantly promoted the expression of oxidative-stress-related genes (Apx2 and Apx6) and a salt-stress-response-related gene (AtRD22), thereby enhancing salt resistance in plants [20]. An analysis with gene silencing technology revealed that OsGRXS17 may play an important role in drought tolerance in rice through two methods: by regulating ABA-dependent expression of drought-responsive gene, participating in the regulation of the downstream signaling pathway of ABA, and by acting as a ROS scavenger to regulate H2O2 homeostasis, participating in H2O2-mediated stomatal closure in response to drought [21]. The ectopic overexpression of AtGRXS17 in Arabidopsis can improve the response of Chrysanthemum to thermal stress, as characterized by the reduction in H2O2 accumulation, reduction in oxidative damage to the cell membrane, enhancement of photosynthetic capacity, and effective prevention of floret shape abnormality and flower wilting caused by thermal stress [22]. Calcium-dependent kinase OsCPK24 in rice reduces its sulfotransferase activity by phosphorylating OsGRX10 to inhibit the oxidative damage caused by cold stress, and high GSH levels and high GSH/GSSG ratio are maintained, thus improving the cold tolerance of rice [23]. Rice can enhance the expression of GRX genes and GSH-dependent antioxidant enzyme activities in plants through the GRXs/GSH/GR system, thereby resisting arsenic (As) toxicity [16]. Compared with wild-type Arabidopsis, under As stress, Arabidopsis with overexpression of chickpea CaGRX exhibited higher seed germination and survival rates, longer root system, and lower As accumulation; this demonstrated that CaGRX can improve the As resistance of plants [24]. Compared with wild-type plants, Arabidopsis with overexpression of chickpea CaGRX exhibited enhanced resistance to drought and salt stresses [25].
Wheat (Triticum aestivum L.) is one of the most widespread grain crops worldwide. However, during its growth, wheat is often subjected to various biotic and abiotic stresses such as insect or pathogen infestations, high or low temperatures, and drought and salt stresses, causing serious decreases in yield and quality. GRXs are involved in many cellular processes in plants, including growth regulation, signal transduction, and response to stress. GRXs are studied more extensively and in detail in humans and animals compared with plants. Among the GRX family members reported to date, 31, 29, and 25 are present in Arabidopsis [26], rice [27], and barley [28], respectively. In this study, we systematically analyzed GRX family members in wheat, classified TaGRXs, and analyzed their evolutionary relationships and expressions. The results of this study can provide useful information for the subsequent functional analysis of TaGRXs and provide candidate genes for breeding adversity-resistant wheat varieties.
2. Materials and Methods
2.1. Bioinformatics Analysis
2.1.1. Identification of GRX Family Members in Wheat
The genome sequences and genome annotation files for wheat, Arabidopsis, and barley were downloaded from the Ensembl Plants database (http://plants.ensembl.org) (accessed on 2 February 2023). The Hidden Markov Model (HMM) file for the typical structural domain of the GRX protein (PF00462) was downloaded from the Pfam database (http://pfam.xfam.org/) (accessed on 3 February 2023). The hmm-search function of HMMER3.0 was used to search for protein sequence files of the wheat genome, with the E-value set to 1 × 10−5, and the protein sequences meeting the requirements were retained as candidate wheat GRX family members. The candidate protein sequences were submitted to SMART (http://smart.embl.de/) (accessed on 15 February 2023) and NCBI-CDD (https://www.ncbi.nlm.nih.gov/cdd/) (accessed on 16 February 2023) databases for further screening, and the sequences that did not match with the sequences of core structural domains or were incomplete were removed. Finally, TaGRX family members were obtained. Additionally, the amino acid length, molecular weight (MW), and isoelectric point (pI) of TaGRXs were calculated using the ExPASy tool (http://web.expasy.org/protparam/) (accessed on 22 February 2023). The subcellular localization was predicted by Cell-PLoc 2.0 (http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/) (accessed on 25 February 2023).
2.1.2. Phylogeny of Wheat GRX Family
Phylogenetic analysis of 85 TaGRX proteins and GRX protein of barley and Arabidopsis was performed using ClustalW, and the phylogenetic tree was constructed using the neighbor-joining method with MEGA11.0. The bootstrap value was set to 1000 replicates. The iTOL2.0 website (https://itol.embl.de/) was used to edit the evolutionary tree (15 March 2023).
2.1.3. Gene Structure and Conserved Motif of Protein
Based on the GFF3 file of wheat genome, the exon and intron numbers and length of TaGRX genes were analyzed using GSDS tool (http://gsds.cbi.pku.edu.cn/) (accessed on 25 March 2023). The conserved motif of TaGRX proteins was analyzed using the MEME tool (http://meme-suite.org/) (accessed on 27 March 2023), with the number of motifs set to 10 and the width of motifs set to 6–50. The gene structure and conserved motif of TaGRXs were visualized using Tbtools [29].
2.1.4. Cis-Regulatory Element of Promoters in TaGRXs
Using TBtools (v2.008), 2000 bp sequences from upstream of the ATG were extracted and submitted to the PlantCARE website (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/) (accessed on 5 April 2023) for the analysis of cis-regulatory elements of promoter. The results were collated and imported into TBtools for visualization.
2.1.5. Chromosomal Localization and Gene Duplication
Based on the GFF3 file of the wheat genome, the chromosome length of the wheat genome and chromosome location files of TaGRX genes were obtained. The chromosomal localization of TaGRX genes was visualized using MapChart tools (v2.3.2), and the genes were named according to their location information. TBtools was used to extract wheat chromosome length, gene density, chromosome location of the TaGRX genes, and synteny information between genes from wheat genome database to generate the synteny of TaGRX genes. The synteny of GRX genes between wheat and other species (including Arabidopsis and barley) was analyzed using MCScanX, and the information was visualized using TBtools.
2.1.6. Expressions of TaGRX Genes
Transcript per kilobase per million mapped reads (TPM) values for different tissues (roots, stems, leaves, flowers, and seeds) of wheat were downloaded from the Wheat Omics 1.0 (http://wheatomics.sdau.edu.cn/) (accessed on 20 April 2023) database, and TBtools was used to draw expression heat maps based on the TPM values of TaGRX genes. The transcriptome data of wheat under different stress levels (salt, high temperature, drought, and low phosphorus (P)) were downloaded from WheatOmics 1.0. The expression data of TaGRX genes were obtained, and the expression heat map was plotted using TBtools.
2.1.7. Protein–Protein Interaction (PPI) Network of TaGRXs
TaGRX protein sequences were submitted to String database (https://cn.string-db.org/) (accessed on 25 April 2023) with the species parameter option set to wheat, and the interactions between TaGRXs proteins were analyzed and visualized using Cytoscape (V3.7.2).
2.2. Gene Expression Validation
2.2.1. Plant Material
Chinese Spring (a wheat variety) with full grain and uniformly sized seeds was used in this study.
2.2.2. Wheat Hydroponics and Stress Treatments
The seeds were disinfected with 75% alcohol for 5 min, rinsed with distilled water 2–3 times, and placed in Petri dishes lined with two layers of moist test paper for germination. After 7 days, the seeds were transferred to a 5 L black plastic bucket containing 1/5 Hoagland nutrient solution (pH 5.8–6.0) for further incubation (16 h/8 h; 23 °C/18 °C; day/night) with continuous aeration. During the growth period, the nutrient solution was changed every 2–3 days. When the seedlings exhibited two leaves and one heart, they were treated with 200 mM NaCl to simulate salt stress and 15% PEG6000 to simulate drought stress, respectively, with three biological replicates. The roots and leaves treated with stress for 3, 6, 9, 12, 24, and 48 h were harvested as stressed samples, and the roots and leaves of 0 h before treating with stress were harvested as control samples. All samples were rapidly cooled in liquid nitrogen and stored at −80 °C for gene expression verification.
2.2.3. Validation of Gene Expression Using qRT-PCR
The total RNA from the root and leaf were extracted using SteadyPure Plant RNA Extraction Kit (AG, Qingdao, China). cDNA was obtained using Evo M-MLV Reverse Transcription Kit for qPCR (AG, Qingdao, China), and PCR was conducted using SYBR Green Pro Taq HS Premixed type Kit (AG, Qingdao, China). The fluorescence quantitative PCR was performed on a CFX96 PCR instrument (Bio-Rad, Hercules, CA, USA). The specific PCR protocol was as follows: 95 °C for 30 s, followed by 40 cycles of 95 °C for 5 s and 60 °C for 30 s. The dissolution curve program was set from 60 °C to 95 °C, with an increment of 0.5 °C and each temperature lasting for 5 s. The specificity of the PCR primers was verified using the dissolution curve. Actin was used as the internal reference to correct the expression value. Finally, the relative expression values of genes were calculated using the 2−ΔΔCt method. The qRT-PCR primers are used are given in Table S1.
3. Results
3.1. The Identification of Wheat GRX Genes and Analysis of Their Physicochemical Characteristics
To identify all GRX genes in wheat genome, the wheat protein database was searched using the HMM profile of structural domains of GRXs as a query file. After removing redundancies, the preliminary screening results of wheat GRXs were obtained, and the structural domain integrity was further confirmed using SMART and NCBI-CDD databases. Finally, 85 TaGRX genes in wheat were identified, which were named TaGRX1–TaGRX85 (Table S2). The physicochemical analysis of TaGRX proteins revealed that the amino acid number ranged from 86 to 363 (average 127.7 aa); the molecular weight ranged from 9.2 to 40.7 kDa (average 13.7 kDa); the isoelectric point ranged from 5.2 to 10.3 (average 8.1) with 80% proteins exhibiting pI > 7, suggesting that the proteins of this family were slightly alkaline (Table S2). The subcellular localization prediction results showed that, except for TaGRX15, TaGRX22, TaGRX29, and TaGRX33, which were located in mitochondria, all other TaGRX genes were located in chloroplasts.
3.2. Phylogenetic Analysis of TaGRX Proteins
To further clarify the evolutionary relationships of the TaGRX family of wheat, sequences of 25 HvGRX proteins of barley and 31 GRX proteins of Arabidopsis were used as references for multiple sequence comparison with the sequences of amino acids encoded by 85 wheat TaGRX genes. A developmental evolution tree was constructed using the neighbor-joining method (Figure 1). As indicated, the 85 wheat TaGRX genes could be divided into the CPYC, CGFS, and CC subfamilies according to the subfamilies of Arabidopsis and barley and conservative structural features. Among them, the CC subfamily had the most members, with 64 in wheat, 18 in barley, and 21 in Arabidopsis. The CPYC subfamily had 12, 3, and 6 members in wheat, barley, and Arabidopsis, respectively. The CGFS subfamily had nine, four, and four members in wheat, barley, and Arabidopsis, respectively. Additionally, the number of hexaploid wheat TaGRX genes was approximately three times the number of diploid barley HvGRX genes. Interestingly, one large branch of the CC subfamily was entirely composed of GRX members of wheat and barley, whereas the other branch was entirely composed of GRX members of Arabidopsis.
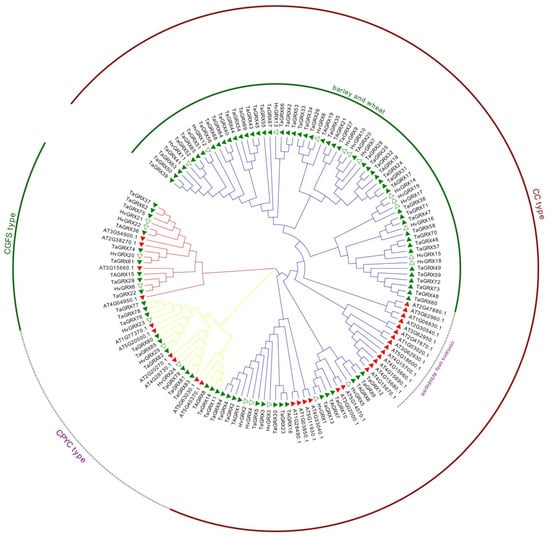
Figure 1.
Analysis of phylogenetic relationships in the GRX family in wheat, barley, and Arabidopsis. The genes in wheat, barley, and Arabidopsis are indicated with a green solid triangle, green hollow triangle, and red solid triangle, respectively. The CC, CPYC, and CGFS subfamilies are indicated with a dark red arc, dashed arc (outer circle), and dark green arc (outer circle), respectively. The dashed arc (inner circle) is the branch containing only GRX genes of Arabidopsis, and the dark green arc (inner circle) is the branch containing only GRX genes of barley and wheat.
3.3. Analysis of the Conserved Motif and Gene Structure of TaGRXs
To better understand the sequence characteristics of TaGRX genes, the conserved motif (Figure 2B), functional domain (Figure 2C), and structural features of the gene (Figure 2D) were analyzed based on the evolutionary analysis (Figure 2A). The functional domain analysis revealed that all TaGRX genes contained the structural domain of GRXs (Figure 2C). The conserved motif analysis of TaGRX proteins using MEME identified 10 motifs, named Motif1–10, with lengths ranging from 6 to 50 aa and members of the same subfamily exhibiting a similar number, type, and distribution of motifs (Figure 2B). For example, all CC subfamily members contained Motif1, Motif2, Motif3, and Motif5; all CPYC subfamily members contained Motif1, Motif2, Motif3, and Motif4; and all CGFS subfamily members contained Motif2 and Motif10. However, Motif6 and Motif8 were only present in the CC subfamily members. Motif9 was only present in the CPYC and CGFS subfamily members, and Motif7 and Motif10 were only present in the CGFS subfamily members. This demonstrated that these motifs had a role in the specific functions of different subfamily members. Additionally, all TaGRX proteins contained Motif2, demonstrating that this motif was highly conservative among all subfamilies and may be important for the function of TaGRX proteins (Figure 2B). Gene structure analysis revealed that 33% TaGRX genes contained introns, and the intron number ranged from 1 to 7 (Figure 2D). The members of the same subfamily had similar exon/intron structures, with most CC subfamily members not containing introns and most CPYC and CGFS subfamily members containing higher number of introns (Figure 2D). Among the 64 CC subfamily members, 57 did not contain introns; 6 contained only 1 intron, and 1 contained 2 introns. Among the 12 CPYC subfamily members, 9 and 3 members contained 3 and 4 introns, respectively. The 9 CGFS subfamily members contained varying number of introns ranging from 1 to 7, and 5 of them contained more than 4 introns. Overall, members of the same subfamily had similar conserved motifs and gene structural features.
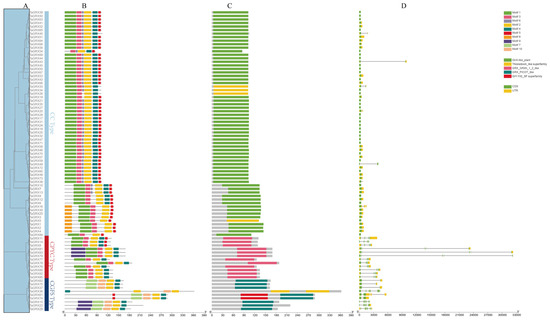
Figure 2.
Analysis of motif and gene structure of TaGRX genes. (A) Phylogenetic evolution tree of wheat TaGRX genes, (B) conservative motif composition of TaGRX genes, (C) functional domains of TaGRX genes, (D) and exon/intron structure of TaGRX genes.
The analysis of active sites of TaGRX proteins revealed that the three subfamilies had different types of active sites, and members of the same subfamily had more conserved active sites (Figure 3). The CC, CPYC, and CGFS subfamily members had conserved CC[M/L][C/S/T], C[P/S][Y/F][C/S], and CGFS sequences at the active sites, respectively. Additionally, all TaGRX proteins contained a more conserved LxxLL motif at the C-terminus, and the CC subfamily members contained more conserved ALWL termini.
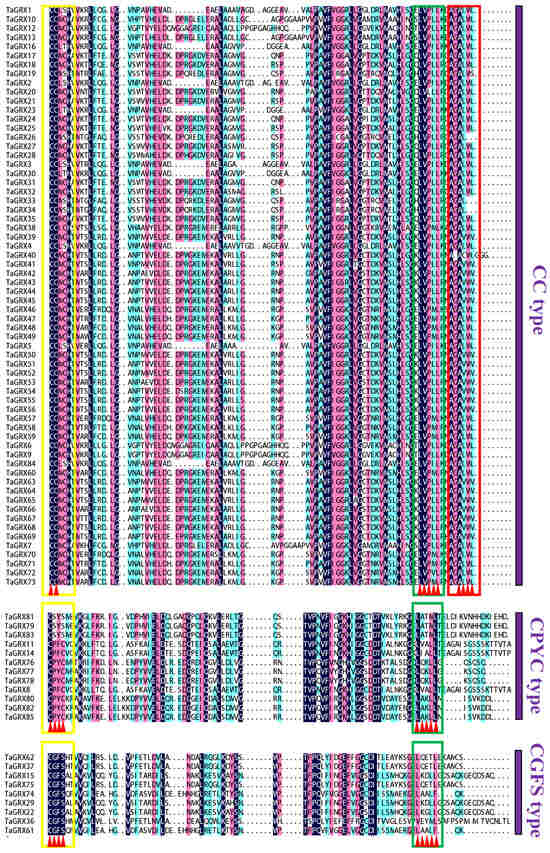
Figure 3.
Comparison of the active sites and C-terminal protein sequences of TaGRX proteins. The yellow, green, and red boxes indicate the Cxx[C/S], C-terminal conserved LxxLL motifs, and conserved ALWL terminus of the CC subfamily, respectively. The background color of dark blue represents 100% identity, pink represents 75% identity, blue-green represents 50% identity, as defined by DNAMAN (v9).
3.4. Chromosomal Localization of TaGRX Genes
Based on the chromosome annotation information of wheat genome, the identified GRX genes in wheat were assessed for chromosomal localization. The results reveal that 85 TaGRX genes were evenly distributed in A, B, and D subgenomes but unevenly distributed on 21 chromosomes. Among them, the TaGRX genes on the 5A, 5B, and 5D chromosomes had the highest incidence (11, 13, and 13, respectively), followed by the 3A, 3B, and 3D chromosomes with seven TaGRX genes each. No GRX gene was present on the 4B chromosome. The number of TaGRX genes on the other chromosomes varied from 1 to 3 (Figure 4).
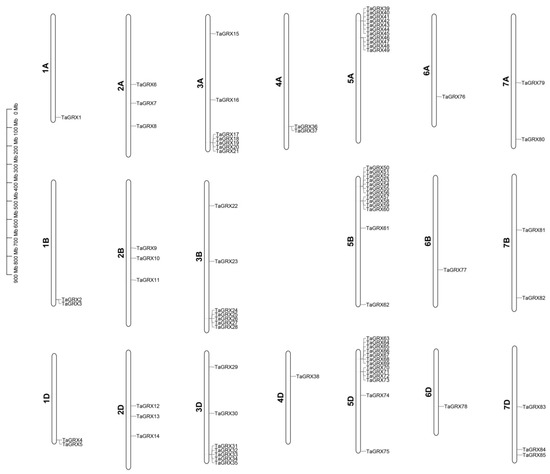
Figure 4.
Chromosomal localization of TaGRX genes. The left scale indicates the length and physical location of the chromosome, and the chromosome number is shown on the left side of the image of each chromosome.
3.5. Cis-Regulatory Element of TaGRX Promoter
To understand the potential functions of TaGRX genes, the cis-regulatory element of the promoter zone of 85 TaGRX genes was analyzed using PlantCARE (Figure 5). The results reveal that 54% of TaGRX members contained auxin-responsive elements; the CC subfamily had the largest proportion with 46 genes. The CC, CPYC, and CGFS subfamily members contained a stress-responsive element; however, the number of genes varied greatly, with 23, 5, and 3 genes, respectively. Similarly, the salicylic-acid-responsive element was present in all subfamilies, with 18, 5, and 1 gene in the CC, CPYC, and CGFS subfamilies, respectively. Additionally, methyl jasmonate (MeJA), MYB, abscisic acid (ABA), and meristem and gibberellin cis-regulatory elements were evenly distributed among all TaGRX genes. However, cell-cycle-regulatory elements were only present in the CC and CPYC subfamily members. These results suggest that TaGRX genes are extensively involved in the growth and response processes of wheat to stress, with some functional differences among subfamily members.

Figure 5.
Cis-regulatory element of the promoter of TaGRX genes in wheat. (A) Phylogenetic evolution tree of TaGRX genes in wheat; (B) schematic diagram of the cis-regulatory element of the promoter of TaGRX genes.
3.6. TaGRX Gene Duplication Event
To further investigate the amplification mechanism of the wheat TaGRX family, the synteny of 85 wheat TaGRX genes was analyzed. The results reveal that 75 TaGRX gene pairs in the TaGRX family existed in synteny, and all gene pairs were located on different chromosomes (Figure 6). Among these, 27, 24, and 22 gene pairs were observed between A and B, A and D, and B and D subgenomes, respectively. Moreover, the B and D subgenomes each had one pair of genes, and no gene pair with collinearity was observed in the A subgenome. These results suggest that the main reason for the expansion of the wheat TaGRX family was fragment duplication, and duplication events mainly occurred among A, B, and D subgenomes but hardly occurred within the same subgenome. Additionally, 43 TaGRX genes had more than two synteny pairs, and three of them (TaGRX2, TaGRX17, and TaGRX30) had four synteny pairs. For example, TaGRX17 on the 3A chromosome had synteny with TaGRX24 and TaGRX25 on the 3B chromosome and TaGRX31 and TaGRX35 on the 3D chromosome, demonstrating that these genes played an important role in the expansion of the GRX family in wheat.
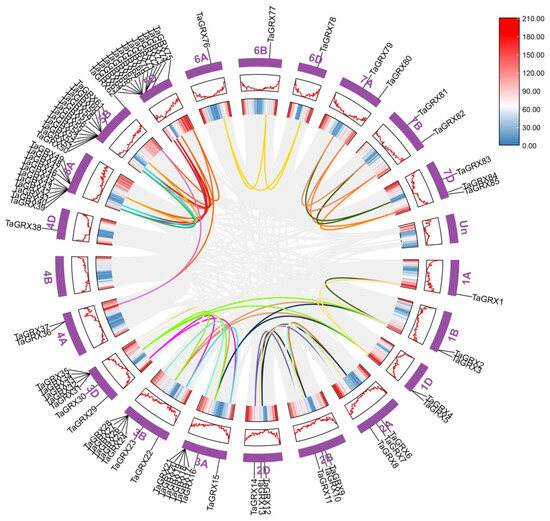
Figure 6.
Synteny analysis of TaGRX family in wheat. Gray lines indicate all synteny gene pairs in the wheat genome; colored lines indicate TaGRX gene pairs with synteny. Chromosome numbers are shown next to each chromosome. The scale represents the gene density; the redder the color, the higher the gene density.
The synteny of GRX genes between wheat and two other species (barley and Arabidopsis) was analyzed. In total, 46 homologous gene pairs existed between wheat and barley, whereas only 7 homologous gene pairs existed between wheat and Arabidopsis (Figure 7). This demonstrated that, compared with the dicotyledonous plant Arabidopsis, the GRX genes between monocotyledonous plants of wheat and barley retained a high degree of homology during the evolution process. Additionally, 14 GRX genes of barley and 3 GRX genes of Arabidopsis had synteny with at least 2 TaGRX genes such as HORVU.MOREX.r3.1HG0087850.1 on 1H chromosome in barley, which had homology with 6 wheat TaGRX genes (TaGRX1, TaGRX2, TaGRX4, TaGRX16, TaGRX23, and TaGRX30). Notably, seven TaGRX genes (TaGRX7, TaGRX10, TaGRX15, TaGRX22, TaGRX29, TaGRX61, and TaGRX74) that were homologous to Arabidopsis also had synteny with GRX genes in barley, demonstrating that these genes were highly conserved in different species.
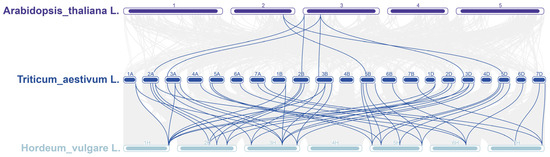
Figure 7.
Synteny analysis of GRX genes in Arabidopsis, wheat, and barley. Gray lines indicate homologous gene pairs in the genome of wheat with those of Arabidopsis and barley. Blue lines indicate GRX gene pairs with homology in the genome of wheat with those of Arabidopsis and barley. The chromosome number is shown at the top of each chromosome.
3.7. PPI Network of TaGRXs
The interaction network of TaGRX proteins was analyzed using the wheat annotation information in the String database. The result revealed complex interactions among some family members (Figure 8). Among these interacting proteins, the seven centrally located genes (TaGRX15, TaGRX22, TaGRX29, TaGRX36, TaGRX37, TaGRX62, and TaGRX75) belong to the CGFS subfamily. These genes interact with both CPYC (for example, TaGRX8, and TaGRX78) and CC (for example, TaGRX10 and TaGRX13) subfamily genes, demonstrating that most CGFS subfamily members perform specific functions through interactions with other genes.
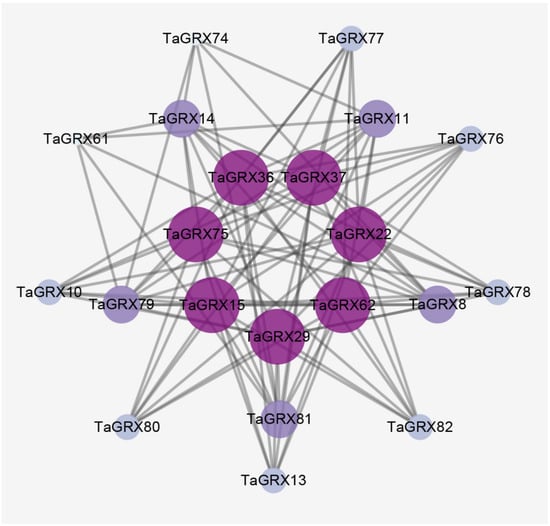
Figure 8.
PPI network prediction model for TaGRXs. A larger circle and darker color indicate more interactions with other proteins.
3.8. Expression Analysis of TaGRX Genes in Different Tissues and Stress Conditions
To understand the expression characteristics of GRX genes in various wheat tissues, a heat map was created by downloading expression levels of 85 TaGRX genes in five tissues (roots, stems, leaves, spikes, and seeds) of the wheat variety China Spring from WheatOmics 1.0. Significant differences were observed in the expression patterns of TaGRX genes in various tissues (Figure 9). These differences were mainly reflected in the different subfamilies, as indicated by the fact that all CPYC and CGFS subfamily members had high expression levels in different tissues, whereas only 11% of the CC subfamily members had high expression levels in different tissues. For example, TaGRX14, TaGRX79, TaGRX82, and TaGRX85 of the CPYC subfamily and TaGRX15, TaGRX22, and TaGRX29 of the CGFS subfamily were highly expressed in all tissues, whereas TaGRX18, TaGRX24, TaGRX31, and other genes of the CC subfamily were hardly expressed in all tissues. Interestingly, the expression of most genes of the CC subfamily was tissue-specific; for example, TaGRX26, TaGRX33, and TaGRX34 were only highly expressed in the roots. TaGRX39 and TaGRX50 were more expressed in the stems; TaGRX46, TaGRX58, and TaGRX71 were more expressed in the stems and leaves, and TaGRX16 and TaGRX30 were more expressed in the spikes and seeds. These results suggested that different GRX subfamily genes play different roles in wheat growth.
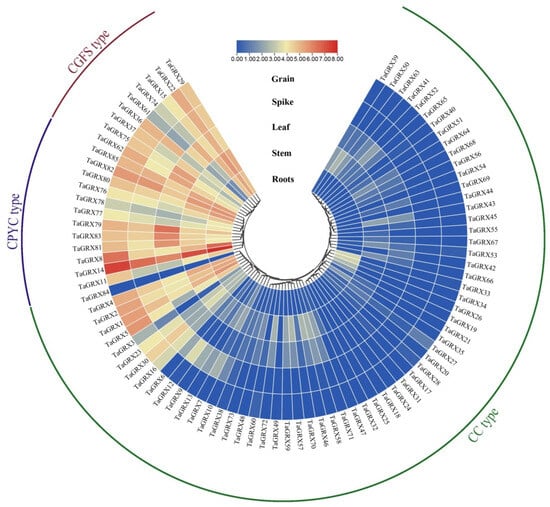
Figure 9.
Expression characteristics of TaGRX genes in different tissues. Expression data are processed using log2. The color scale indicates TPM values from high (red) to low (blue).
Based on the published wheat transcriptome data, the response characteristics of TaGRX genes to abiotic stresses such as drought, high temperatures, low P, and salt were analyzed. The results reveal that the expression levels of some TaGRX genes varied significantly under different stress conditions, and significant differences were observed in the expression characteristics of different subfamilies (Figure 10). Almost all genes from CPYC and CGFS subfamilies mainly responded to high-temperature stress. For example, TaGRX11, TaGRX37, and TaGRX82 were significantly upregulated under high-temperature stress. The CC subfamily genes mainly responded to drought, salt, and low-P stresses. For example, TaGRX53 and TaGRX66 were significantly upregulated in response to salt stress, and TaGRX19 and TaGRX33 were significantly upregulated under low-P stress. Additionally, some CC subfamily genes (such as TaGRX39 and TaGRX50) could respond to both drought and salt stresses, and some (such as TaGRX5 and TaGRX60) could respond to both high-temperature and drought stresses. These results suggested that some TaGRX genes played a certain role in coping with abiotic stress.
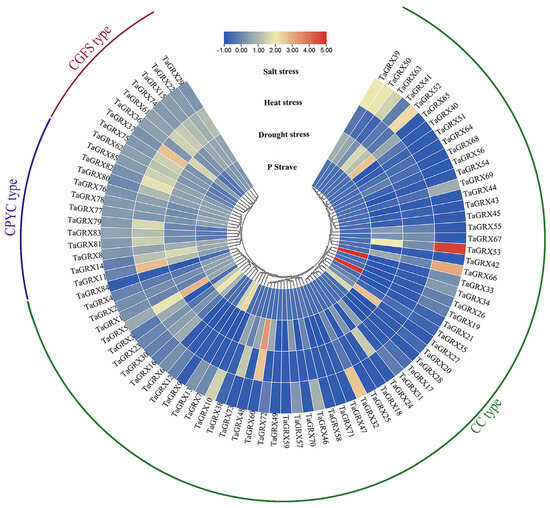
Figure 10.
Expression characteristics of TaGRX genes in response to different abiotic stresses. Expression data are processed using log2. The color scale indicates relative expression values from high (red) to low (blue).
3.9. Verification of the Expression of TaGRX60 and TaGRX66 Genes Using qRT-PCR
To verify the expression of TaGRX60 and TaGRX66 (which significantly responded to drought and salt stresses, respectively), these genes were selected for qRT-PCR validation. Using Chinese Spring as the material, the expression of the two genes in the roots and leaves of plants was further examined via PEG simulation of drought and salt stresses, respectively. The results reveal that the expression of TaGRX60 and TaGRX66 exhibited a trend of first increasing and then decreasing (Figure 11). The expression of TaGRX60 was increased in the roots and leaves after 9 h of PEG treatment; however, the upregulation rate in the roots was slightly higher than that in leaves, and with the prolongation of the treatment time, the expression level was restored to the original level in both roots and leaves. TaGRX66 was upregulated by salt stress in both roots and leaves; however, the response speed was faster in the roots than in the leaves, and the upregulation rate was much higher in the roots than in the leaves. The expression of TaGRX66 in the roots peaked at 3 h after salt treatment, which was 32 times higher than that in the control. The expression of TaGRX66 in the leaves peaked at 12 h after salt treatment, which was five times higher than that in the control. With the prolongation of the treatment time, the expression of TaGRX66 in both roots and leaves began to decrease, and almost recovered to the original level after 48 h of treatment.
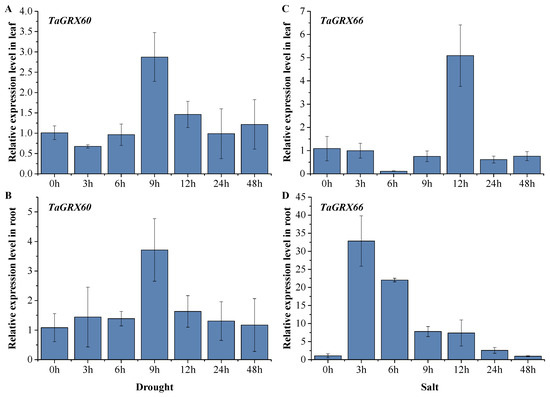
Figure 11.
Expression patterns of TaGRX60 and TaGRX66 genes under drought and salt stresses. Relative expression of TaGRX60 under drought (A,B) stress and of TaGRX66 under salt (C,D) stress. The x-axis indicates treatment duration, and the y-axis indicates relative expression level. Expression data are expressed as the means of three biological replicates.
4. Discussion
Following its rapid development, the whole-genome sequencing of many plants has been completed. GRX genes are found in some plant species [30,31], and the GRX family has been comprehensively analyzed in Arabidopsis, rice, and barley. However, the understanding of GRX genes in wheat is still insufficient. In this study, we identified GRX genes in wheat in the whole genome, and 85 TaGRXs (Table S2) were identified and characterized using bioinformatic and expression analyses. The results of this study are of great significance for the in-depth understanding of the functions of TaGRXs and their mechanisms of action.
4.1. GRX Genes in Plants
GRXs are small-molecular-weight proteins, and they play a key role in maintaining the redox homeostasis of cells. GRXs are widely present in nature. Most studies on GRXs have been conducted in animals and humans, with few focusing on plants. GRXs usually contain a conserved CXXC/S active site. Based on the amino acid sequences of its active sites, GRX genes can be divided into the CPYC, CGFS, and CC subfamilies [32]. The CPYC and CGFS subfamilies are present in eukaryotes, and the CC subfamily is unique to higher plants and has the highest number of genes. GRXs act via both mono- and bisulfhydryl mechanisms. GRXs catalyze the reduction in disulfide bonds linked to GSH through the monosulfhydryl mechanism, which only requires the involvement of cysteine (Cys) at the N-terminal end of the active center of GRXs. In the bisulfhydryl mechanism, the GRX-catalyzed reduction in disulfide bonds of proteins requires the simultaneous involvement of both Cys at the N- and C-terminals [33]. GRXs, GSH, NADPH, and GR together form the glutaredoxin system. Electrons are first transferred from NADPH to GR, then to GSH, and finally to GRXs, which utilize the gained electrons to reduce disulfide bonds in target proteins or between proteins [32]. In plants, the glutaredoxin system, along with the thioredoxin and AsA-GSH systems, is involved in the maintenance of the cellular redox state and regulation of various cellular processes and plays an important role in the resistance of plant cells to biotic and abiotic oxidative stresses [33]. Additionally, GRX genes play important roles in the growth, nitrogen metabolism, and iron–sulfur cluster formation in plants [34].
4.2. Evolution and Expansion of TaGRX Genes
Based on the sequence characterization of 85 TaGRX genes, they could be divided into the CPYC, CGFS, and CC subfamilies (Figure 1 and Figure 3), which is consistent with the reported classification of GRX subfamilies. Members of the same subfamily had similar motif compositions and gene structural features (Figure 2), demonstrating that members of the same subfamily may have similar functions. The phylogenetic tree revealed that the GRX genes of the monocotyledonous plants wheat and barley were more homologous than those of the dicotyledonous plant Arabidopsis. Particularly for the CC subfamily members unique to plants, most of the GRX genes of wheat and barley were in the same evolutionary branch, whereas GRX genes of Arabidopsis were in another evolutionary branch (Figure 1). This demonstrated a certain separation of the GRX genes between monocotyledonous and dicotyledonous plants during the evolutionary process. The main causes of gene family expansion include genome polyploidization, tandem duplication, fragment duplication, reverse transcription transposition, or exon duplication and reorganization [35,36]. Wheat is a hexaploid and barley is a diploid. The wheat genome is approximately three times larger than the barley genome. A total of 25 GRX genes are identified in barley, which indicates that the number of TaGRX genes (85) is approximately three times the number of HvGRX genes. Additionally, although the 85 TaGRX genes were unevenly distributed across 21 chromosomes, they were more evenly distributed in the A, B, and D subgenomes (Figure 4). This demonstrated that the doubling of the number of wheat GRX genes in the hexaploid is due to the doubling of the chromosome compared with the diploid. Additionally, the synteny of TaGRX genes existed among different chromosomes, mainly among different subgenomes; however, it hardly occurred within the same subgenome. Specifically, 97% of the synteny gene pairs existed among A, B, and D subgenomes, and only 3% of the synteny gene pairs existed within the same subgenome (Figure 6). This suggested that the main reason for the expansion of the wheat TaGRX family is a fragment duplication, also indicating that chromosome polyploidization facilitates the expansion of the TaGRX family.
4.3. The CGFS Subfamily Members in Wheat Have a High Conservatism and Perform Specific Functions
The CGFS subfamily members are ubiquitous in nature and have conserved CGFS active sites, which function through a monosulfhydryl mechanism. Studies have reported that yeast mutants lacking the single sulfhydryl GRX protein cannot survive, demonstrating that the single sulfhydryl group has a specific requirement for its substrate, whereas the corresponding double sulfhydryl GRX protein cannot replace this function [37]. In Arabidopsis, the CGFS subfamily includes AtGRXS14, AtGRXS15, AtGRXS16, and AtGRXS17, which are mainly found in the chloroplasts and mitochondria and are involved in the assembly, transport, and insertion of Fe-S clusters [38,39]. The oxidative state reduction in some typical CGFS subfamily members (such as AtGRXS16) depend on ferredoxin–thioredoxin reductase (FTR), rather than GSH [40]. A total of nine CGFS subfamily members in wheat were identified in this study, all containing conserved CGFS active sites (Figure 3). Interestingly, among the seven TaGRX gene synteny that coexisted with barley (a monocotyledonous plant) and Arabidopsis (a dicotyledonous plant), five were CGFS subfamily members (Figure 7). This demonstrated that the CGFS subfamily members retained a high degree of homology during the evolutionary process, and the genes of this subfamily are conserved in both monocotyledonous and dicotyledonous plants. Additionally, all seven genes at the center of the PPI network of TaGRXs belong to the CGFS subfamily (Figure 8), demonstrating that most CGFS subfamily members play an important role in the regulatory functions through interaction with other subfamily members.
4.4. TaGRX Genes Play a Role in Growth and Stress Resistance
Cis-regulatory elements are specific motifs located in the promoter zone domain of genes. Upstream regulatory genes bind to the cis-regulatory element in the promoter zone of target genes to regulate the expression of the target genes and enable the target genes to perform specific functions. Genes containing auxin- and stress-responsive elements in the promoter often play a role in plant growth and stress resistance [41]. To enhance the salt resistance of wheat, TaBZR1 not only directly binds to the G-box motifs of the promoter of TaNCED3 (a key gene for ABA synthesis) to activate its expression and promote ABA synthesis, but also binds to the promoters of ROS-clearance-related genes (TaGPX2 and TaGPX3) and activates their expression [42]. The transcription factor TabHLH49 binds to the promoter of the dehydrin gene WZY2 and regulates the expression of the WZY2 gene, thereby enhancing the drought resistance of wheat [43]. In this study, we analyzed the cis-regulatory element of TaGRXs and reported that their promoter zone contained a large number of auxin- and stress-responsive elements (such as auxin, salicylic acid, MeJA, ABA, and gibberellin, which were hormone-responsive elements) and MYB transcription factor binding sites (Figure 5). Additionally, it contained meristem and cell-cycle-regulation-related elements; however, the cell-cycle-regulation-related element was only present in the promoters of the CC and CPYC subfamily members (Figure 5). These results suggest that TaGRX genes play important roles in hormone regulation, morphogenesis, and stress response in wheat, and the functions of different subfamily members are different.
To date, a number of GRX genes have been reported to be involved in the regulation of growth and stress response in plants. For example, ROXY1 and ROXY2 regulate anther development [44]; the OsGRXC2.2 gene regulates embryonic development [13]; the OsGRXS17 gene regulates drought resistance [21]. The functions of genes are closely related to their expression levels. To understand the function of TaGRX genes, their expressions in different tissues and under different stress conditions were analyzed using the published wheat transcriptome data, and the expression patterns were verified using qRT-PCR (Figure 9, Figure 10 and Figure 11). As indicated, members of the CPYC and CGFS subfamilies were characterized by a constitutive expression, with high background expression levels in different tissues, whereas most CC subfamily members had very low background expression levels (Figure 9). This suggested significant differences in the expression patterns of TaGRX genes in different tissues, and the roles of different GRX subfamily genes were different. Additionally, most of the CPYC and CGFS subfamily genes responded to high-temperature stress, whereas the CC subfamily genes mainly responded to drought, salt, and low-P stresses (Figure 10). This demonstrated that different GRX subfamily genes played different roles in stress response, which may be important for wheat to resist abiotic stresses such as drought and high temperatures.
5. Conclusions
GRX is a class of small-molecular-weight, heat-stable proteins that can regulate the reversible redox response of proteins and play an important role in the growth and resistance to biotic and abiotic stresses in plants. In this study, 85 TaGRX genes were identified in wheat, and their gene sequences, gene structure, evolutionary relationships, chromosomal localization, cis-regulatory element of the promoter, intra- and inter-species synteny, and protein interactions and expression characteristics were analyzed. The 85 TaGRX genes could be further divided into the CPYC, CGFS, and CC subfamilies. Among them, the CC subfamily members were the most numerous, and the genes in the same subfamily had similar gene structure and conserved motif. The TaGRX genes were evenly distributed in the three subgenomes of wheat, and the main reason for their expansion was chromosome doubling and large fragment duplication. The CGFS subfamily retained a high degree of conservatism during the evolution and had an important position in the PPI network. Significant differences in the expression characteristics of different subfamily genes were noted; the genes of CPYC and CGFS subfamilies exhibited constitutive expression characteristics and mainly responded to high-temperature stress, whereas most CC subfamily genes exhibited low background expression levels but were upregulated under drought, salt, and low-P stresses. It is speculated that these genes have a specific function in the response of wheat to abiotic stress. This study lays the foundation for the further investigation of the functions and mechanisms of TaGRX genes in wheat.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy13122985/s1, Table S1: qRT-PCR primers for TaGRX60 and TaGRX66 in this study; Table S2: Basic information of wheat TaGRXs family.
Author Contributions
All authors contributed to the study conception and design. X.H. and W.C. carried out the experimental design, bioinformatic analysis, and writing. W.C. and X.S. analyzed the data and prepared the figures and tables. Y.G. and Y.H. performed the gene expression experiments. X.X., C.S., Y.L., B.R. and H.Y. checked and verified all the data. P.M., J.Z. and W.M. performed the review, supervision, editing and project management. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (32372167), the Shandong Provincial Key Research and Development Plan (Major Science and Technology Innovation Project) (2021LZGC013), the Wheat Innovation Team of Modern Agricultural Production Systems in Shandong Province (SDAIT01-05), the Natural Science Foundation of Shandong Province (ZR2021MC025), the High-Level Talents Project of Qingdao Agricultural University (663/1119013).
Data Availability Statement
Data presented in this paper are contained within the article and Supplementary Materials.
Acknowledgments
The authors would like to thank all the reviewers who participated in the review.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chen, Q.; Yang, G. Signal function studies of ROS, especially RBOH-dependent ROS, in plant growth, development and environmental stress. J. Plant Growth Regul. 2020, 39, 157–171. [Google Scholar] [CrossRef]
- Huang, H.; Ullah, F.; Zhou, D.-X.; Yi, M.; Zhao, Y. Mechanisms of ROS regulation of plant development and stress responses. Front. Plant Sci. 2019, 10, 800. [Google Scholar] [CrossRef] [PubMed]
- Wormuth, D.; Heiber, I.; Shaikali, J.; Kandlbinder, A.; Baier, M.; Dietz, K.-J. Redox regulation and antioxidative defence in Arabidopsis leaves viewed from a systems biology perspective. J. Biotechnol. 2007, 129, 229–248. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, A. Hydrogen donor system for escherichia eoli ribonucleoside-diphosphate reductase dependent upon glutathione. Proc. Natl. Acad. Sci. USA 1976, 73, 2275–2279. [Google Scholar] [CrossRef] [PubMed]
- Rouhier, N. Plant glutaredoxins: Pivotal players in redox biology and iron-sulphur centre assembly. New Phytol. 2010, 186, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Xing, S.; Lauri, A.; Zachgo, S. Redox regulation and flower development: A novel function for glutaredoxins. Plant Biol. 2006, 8, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Rouhier, N.; Gelhaye, E.; Jacquot, J.-P. Plant glutaredoxins: Still mysterious reducing systems. Cell. Mol. Life Sci. 2004, 61, 1266–1277. [Google Scholar] [CrossRef] [PubMed]
- Xing, S.; Rosso, M.G.; Zachgo, S. ROXY1, a member of the plant glutaredoxin family, is required for petal development in Arabidopsis thaliana. Development 2005, 132, 1555–1565. [Google Scholar] [CrossRef]
- Wang, Z.; Xing, S.; Birkenbihl, R.P.; Zachgo, S. Conserved functions of Arabidopsis and Rice CC-Type glutaredoxins in flower development and pathogen response. Mol. Plant 2009, 2, 323–335. [Google Scholar] [CrossRef]
- Li, S.; Lauri, A.; Ziemann, M.; Busch, A.; Bhave, M.; Zachgo, S. Nuclear activity of ROXY1, a glutaredoxin interacting with TGA factors, is required for petal development in Arabidopsis thaliana. Plant Cell. 2009, 21, 429–441. [Google Scholar] [CrossRef]
- Murmu, J.; Bush, M.J.; DeLong, C.; Li, S.; Xu, M.; Khan, M.; Malcolmson, C.; Fobert, P.R.; Zachgo, S.; Hepworth, S.R. Arabidopsis basic leucine-zipper transcription factors TGA9 and TGA10 interact with floral glutaredoxins ROXY1 and ROXY2 and are redundantly required for anther development. Plant Physiol. 2010, 154, 1492–1504. [Google Scholar] [CrossRef]
- Quon, T.; Lampugnani, E.R.; Smyth, D.R. Petal loss and ROXY1 interact to limit growth within and between sepals but to promote petal initiation in Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 152. [Google Scholar] [CrossRef]
- Liu, S.; Fun, H.; Jiang, J.; Chen, Z.; Gao, J.; Shu, H.; Zhang, S.; Yang, C.; Liu, J. Overexpression of a CPYC-Type glutaredoxin, OsGrxC2.2, causes abnormal embryos and an increased grain weight in Rice. Front. Plant Sci. 2019, 10, 848. [Google Scholar] [PubMed]
- Verma, P.K.; Verma, S.; Pande, V.; Mallick, S.; Tripathi, R.D.; Dhankher, O.P.; Chakrabarty, D. Overexpression of Rice glutaredoxin OsGrx_C7 and OsGrx_C2.1 reduces intracellular arsenic accumulation and increases tolerance in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 740. [Google Scholar] [PubMed]
- Xu, F.; Tang, J.; Gao, S.; Chang, X.; Du, L.; Chu, C. Control of Rice pre-harvest sprouting by glutaredoxin-mediated abscisic acid signaling. Plant J. 2019, 100, 1036–1051. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.; Knuesting, J.; Bariat, L.; Dard, A.; Freibert, S.A.; Marchand, C.H.; Young, D.; Nguyen, H.T.D.; Voth, W.; Debures, A.; et al. Redox modification of the iron-sulfur glutaredoxin GRXS17 activates holdase activity and protets plants from heat stress. Plant Physiol. 2020, 184, 676–692. [Google Scholar] [CrossRef] [PubMed]
- Knuesting, J.; Riondet, C.; Maria, C.; Kruse, I.; Becuwe, N.; Koenig, N.; Berndt, C.; Tourrette, S.; Guilleminot-Montoya, J.; Herrero, E.; et al. Arabidopsis glutaredoxin S17 and it’s partner, the nuclear factor Y subunit C11/negative cofactor 2α, contribute to maintenance of the shoot apical meristem under long-day photoperiod. Plant Physiol. 2015, 167, 1643–1658. [Google Scholar] [CrossRef]
- Son, S.; Kim, H.; Lee, K.S.; Kim, S.; Park, S.R. Rice glutaredoxin GRXS15 confers broad-spectrum resistance to Xanthomonas oryzae pv. Oryzae and Fusarium fujikuroi. Biochem. Biophys. Res. Commun. 2020, 533, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Ning, X.; Sun, Y.; Wang, C.; Zhang, W.; Sun, M.; Hu, H.; Liu, J.; Yang, L. A Rice CPYC-Type glutaredoxin OsGRX20 in protection against bacterial blight, methyl viologen and salt stresses. Front. Plant Sci. 2018, 9, 111. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Huang, C.; Xie, Y.; Song, F.; Zhou, X. A Tomato glutaredoxin gene SlGRX1 regulates plant responses to oxidative, drought and salt Stresses. Planta 2010, 232, 1499–1509. [Google Scholar] [CrossRef]
- Hu, Y.; Wu, Q.; Peng, Z.; Sprague, S.A.; Wang, W.; Park, J.; Akhunov, E.; Jagadish, K.S.V.; Nakata, P.A.; Cheng, N.; et al. Silencing of OsGRXS17 in Rice improves drought stress tolerance by modulating ROS accumulation and stomatal closure. Sci. Rep. 2017, 7, 15950. [Google Scholar] [CrossRef]
- Kang, B.-C.; Wu, Q.; Sprague, S.; Park, S.; White, F.F.; Bae, S.-J.; Han, J.-S. Ectopic overexpression of an Arabidopsis monothiol glutaredoxin AtGRXS17 affects floral development and improves response to heat stress in Chrysanthemum (Chrysanthemum morifolium Ramat.). Environ. Exp. Bot. 2019, 167, 103864. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, C.; Zhu, Y.; Zhang, L.; Chen, T.; Zhou, F.; Chen, H.; Lin, Y. The calcium-dependent kinase OsCPK24 functions in cold stress responses in Rice. J. Integr. Plant Biol. 2018, 60, 173–188. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Dubey, A.K.; Kumar, V.; Ansari, M.A.; Narayan, S.; Meenakshi; Kumar, S.; Pandey, V.; Shirke, P.A.; Pande, V.; et al. Over-expression of chickpea glutaredoxin (CaGrx) provides tolerance to heavy metals by reducing metal accumulation and improved physiological and antioxidant defence System. Ecotox. Environ. Saf. 2020, 192, 110252. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, V.; Dubey, A.K.; Ansari, M.A.; Narayan, S.; Meenakshi; Kumar, S.; Pandey, V.; Pande, V.; Sanyal, I. Chickpea glutaredoxin (CaGrx) gene mitigates drought and salinity stress by modulating the physiological performance and antioxidant defense mechanisms. Physiol. Mol. Biol. Plants 2021, 27, 923–944. [Google Scholar] [CrossRef]
- Rouhier, N.; Couturier, J.; Jacquot, J.-P. Genome-wide analysis of plant glutaredoxin systems. J. Exp. Bot. 2006, 57, 1685–1696. [Google Scholar] [CrossRef] [PubMed]
- Garg, R.; Jhanwar, S.; Tyagi, A.K.; Jain, M. Genome-wide survey and expression analysis suggest diverse roles of glutaredoxin gene family members during development and response to various stimuli in Rice. DNA Res. 2010, 17, 353–367. [Google Scholar] [CrossRef]
- Zhang, S.; Yu, J.; Liang, W. Genome-wide identification and bioinformatics analysis of GRX family in barley. Mol. Plant Breed. 2022, 20, 2126–2134. [Google Scholar]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Esposito, S.; Cappetta, E.; Tranchida-Lombardo, V.; Batelli, G.; Ruggiero, A.; Ruocco, M.; Sportelli, G.; Cillo, F.; De Palma, M. Genome-wide survey of glutaredoxin gene family in Four Solanaceae species and exploitation of duplicated CC-Type following different environmental stimuli in Tomato (Solanum lycopersicum). Sci. Hortic. 2023, 319, 112188. [Google Scholar] [CrossRef]
- Malik, W.A.; Wang, X.; Wang, X.; Shu, N.; Cui, R.; Chen, X.; Wang, D.; Lu, X.; Yin, Z.; Wang, J.; et al. Genome-wide expression analysis suggests glutaredoxin genes response to various stresses in cotton. Int. J. Biol. Macromol. 2020, 153, 470–491. [Google Scholar] [CrossRef]
- Meyer, Y.; Belin, C.; Delorme-Hinoux, V.; Reichheld, J.-P.; Riondet, C. Thioredoxin and glutaredoxin systems in plants: Molecular mechanisms, crosstalks, and functional significance. Antioxid. Redox Signal. 2012, 17, 1124–1160. [Google Scholar] [CrossRef] [PubMed]
- Meyer, Y.; Buchanan, B.B.; Vignols, F.; Reichheld, J.-P. Thioredoxins and glutaredoxins: Unifying elements in redox biology. Annu. Rev. Genet. 2009, 43, 335–367. [Google Scholar] [CrossRef]
- Wu, Q.; Yang, J.; Cheng, N.; Hirschi, K.D.; White, F.F.; Park, S. Glutaredoxins in plant development, abiotic stress response, and iron homeostasis: From model organisms to crops. Environ. Exp. Bot. 2017, 139, 91–98. [Google Scholar] [CrossRef]
- Bai, J.; Song, M.J.; Gao, J.; Li, G. Whole genome duplication and dispersed duplication characterize the evolution of the plant PINOID gene family across plant species. Gene 2022, 829, 146494. [Google Scholar] [CrossRef] [PubMed]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Manzaneque, M.T.; Ros, J.; Cabiscol, E.; Sorribas, A.; Herrero, E. Grx5 Glutaredoxin plays a central role in protection against protein oxidative damage in Saccharomyces cerevisiae. Mol. Cell. Biol. 1999, 19, 8180–8190. [Google Scholar] [CrossRef] [PubMed]
- Rey, P.; Taupin-Broggini, M.; Couturier, J.; Vignols, F.; Rouhier, N. Is there a role for glutaredoxins and BOLAs in the perception of the cellular iron status in plants? Front. Plant Sci. 2019, 10, 712. [Google Scholar] [CrossRef] [PubMed]
- Couturier, J.; Przybyla-Toscano, J.; Roret, T.; Didierjean, C.; Rouhier, N. The roles of glutaredoxins ligating Fe-S clusters: Sensing, transfer or repair functions? Biochim. Biophys. Acta-Mol. Cell Res. 2015, 1853, 1513–1527. [Google Scholar] [CrossRef]
- Zannini, F.; Moseler, A.; Bchini, R.; Dhalleine, T.; Meyer, A.J.; Rouhier, N.; Couturier, J. The thioredoxin-mediated recycling of Arabidopsis Thaliana GRXS16 relies on a conserved C-terminal cysteine. Biochim. Biophys. Acta-Gen. Subj. 2019, 1863, 426–436. [Google Scholar] [CrossRef]
- Zhu, S.; Wang, X.; Chen, W.; Yao, J.; Li, Y.; Fang, S.; Lv, Y.; Li, X.; Pan, J.; Liu, C.; et al. Cotton DMP gene family: Characterization, evolution, and expression profiles during development and stress. Int. J. Biol. Macromol. 2021, 183, 1257–1269. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Yang, Z.; Xing, M.; Jing, Y.; Zhang, Y.; Zhang, K.; Zhou, Y.; Zhao, H.; Qiao, W.; Sun, J. TaBZR1 enhances wheat salt tolerance via promoting ABA biosynthesis and ROS scavenging. J. Genet. Genom. 2023, 50, 861–871. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yang, Y.; Liu, D.; Wang, X.; Zhang, L. Transcription factor TabHLH49 positively regulates dehydrin WZY2 gene expression and enhances drought stress tolerance in wheat. BMC Plant Biol. 2020, 20, 259. [Google Scholar] [CrossRef] [PubMed]
- Xing, S.; Zachgo, S. ROXY1 and ROXY2, two Arabidopsis glutaredoxin genes, are required for anther development. Plant J. 2008, 53, 790–801. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).