
Nutritional Composition, Antioxidant Activity, Cytotoxicity, and Enzymatic Potential of Ficus nitida-Associated Tomophagus colossus
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Isolation of Fungal Material
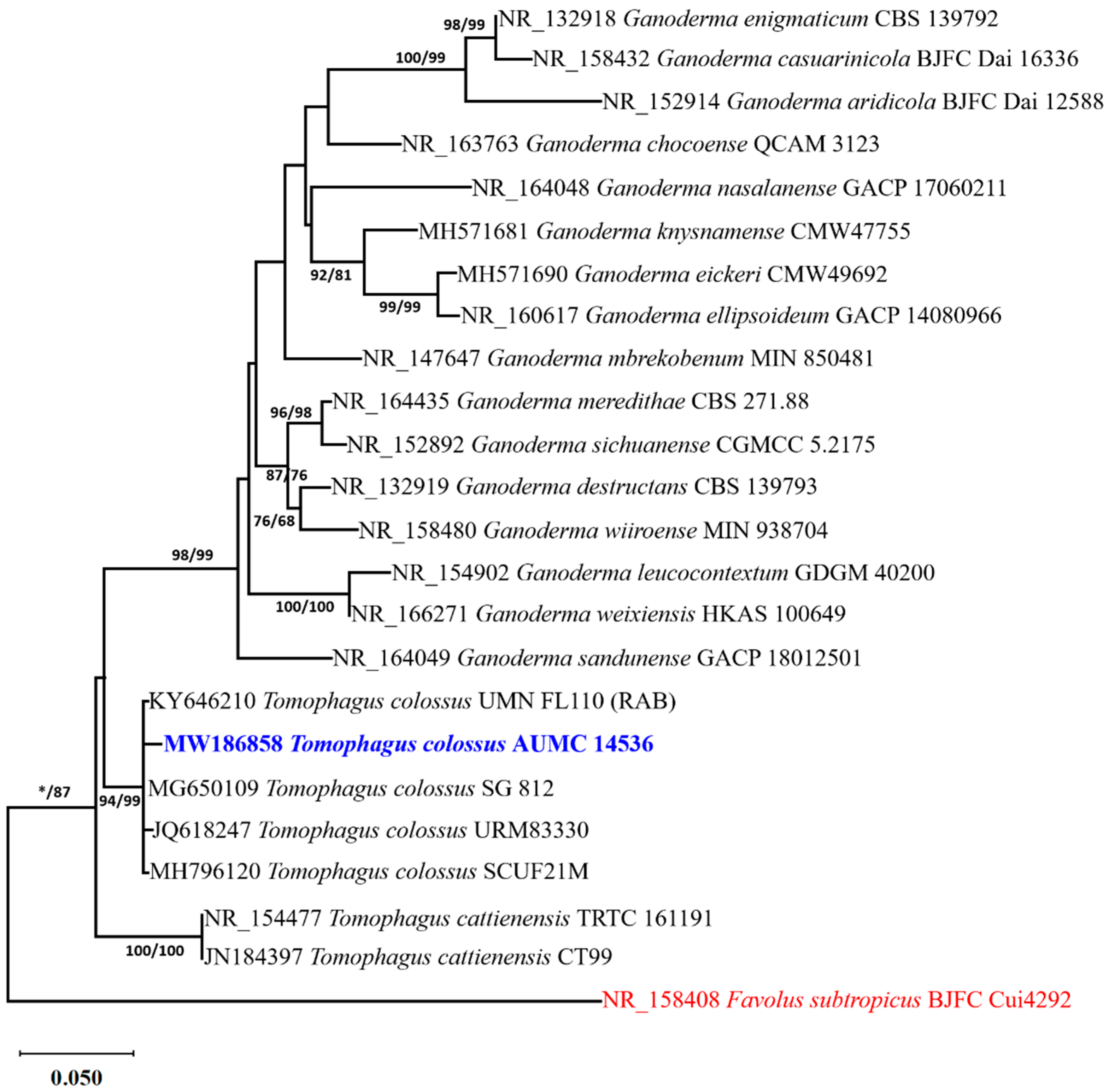
2.2. Molecular Identification of the Fungal Sample
2.3. Alignments and Phylogenetic Analyses
2.4. Macro- and Micronutrient Analyses of Tomophagus colossus Basidioma
2.5. Antioxidant Activity of Tomophagus colossus’s Basidioma
2.6. Amino Acids Content of T. colossus Basidioma
2.6.1. Preparation of Standard Stock Solution
2.6.2. Sample Hydrolysis
2.6.3. Analysis of Amino Acids Content
2.7. Cytotoxicity Test
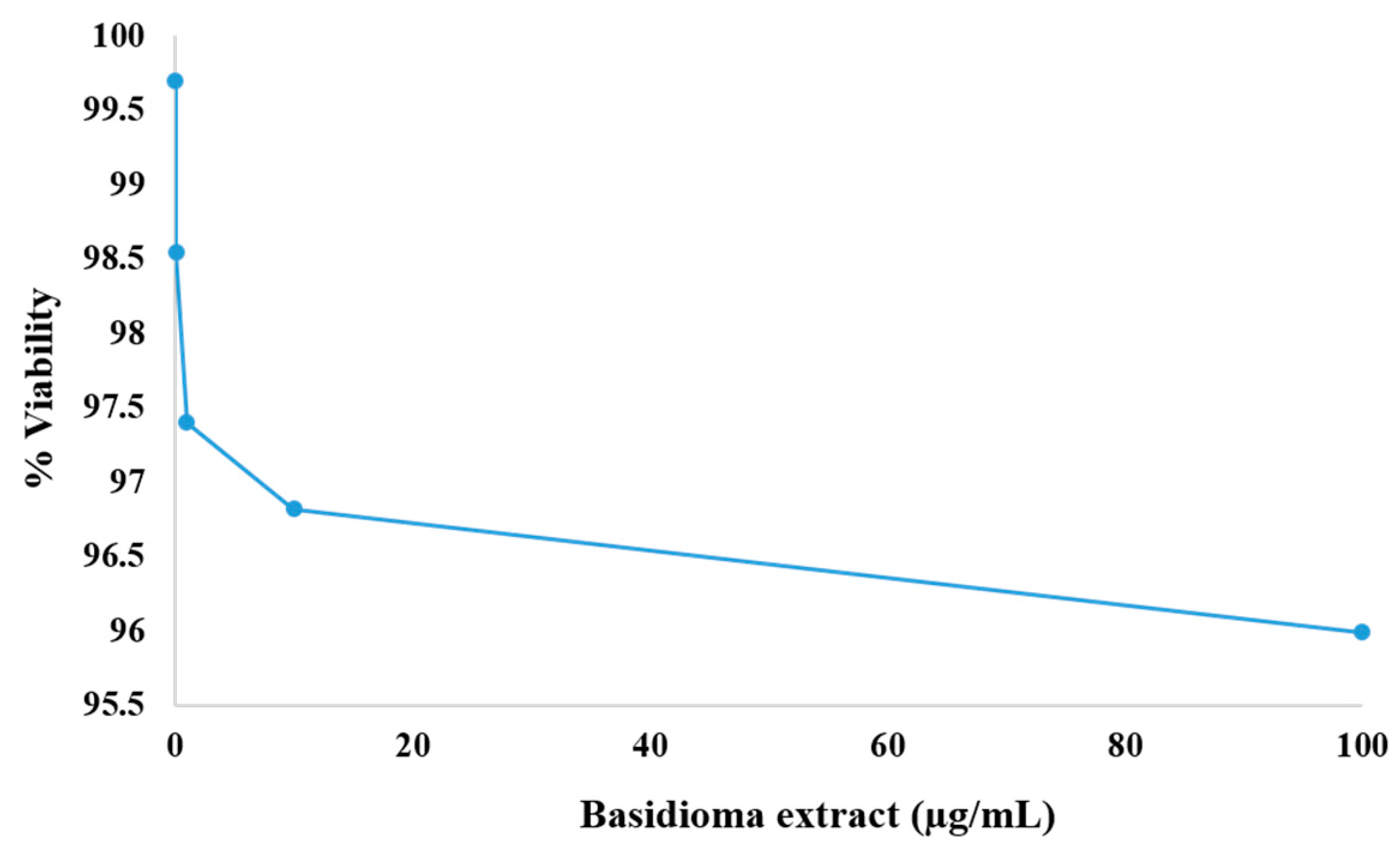
2.8. Cell Viability
2.9. Solid-State Fermentation of Some Lignocellulosic Wastes by T. colossus AUMC 14536
2.9.1. Substrate Pretreatment
2.9.2. Enzyme Production in SSF
2.9.3. Enzyme Assay and Protein Estimation
3. Results
3.1. Molecular Identification of the Tomophagus Isolate
3.2. Brief Description of T. colossus AUMC 14536
3.3. Chemical Analyses of T. colossus’s Basidioma
3.4. Antioxidant Activity of T. colossus AUMC 14536 Biomass
3.5. Analysis of Amino Acids Content
3.6. Cytotoxicity
3.7. Cell Wall Hydrolyzing Enzymatic Potential of T. colossus AUMC 14536 in SSF
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Flood, J.; Bridge, P.; Holderness, M. Ganoderma Diseases of Perennial Crops; CABI: Wallingford, UK, 2000. [Google Scholar]
- Jong, S.; Birmingham, J. Medicinal benefits of the mushroom Ganoderma. Adv. Appl. Microbiol. 1992, 37, 101–134. [Google Scholar]
- Lin, Z.B.; Zhang, H.N. Anti-tumor and immunoregulatory activities of Ganoderma lucidum and its possible mechanisms. Acta Pharmacol. Sin. 2004, 25, 1387–1395. [Google Scholar] [PubMed]
- Le, X.T.; Nguyen Le, Q.H.; Pham, N.D.; Duong, V.H.; Dentinger, B.T.; Moncalvo, J.-M. Tomophagus cattienensis sp. nov., a new Ganodermataceae species from Vietnam: Evidence from morphology and ITS DNA barcodes. Mycol. Prog. 2012, 11, 775–780. [Google Scholar]
- Fries, E.M. Novae symbolae mycologicae, in peregrinis terris a botanicis danicis collectae. In Nova Acta Regia Societis Scientiarum Upsaliensis, Ser. 3; 1851; pp. 17–136. Available online: https://cir.nii.ac.jp/crid/1130000795112975744 (accessed on 16 October 2023).
- Furtado, J.S. Ganoderma colossum and the status of Tomophagus. Mycologia 1965, 57, 979–984. [Google Scholar] [CrossRef]
- Steyaert, R. Species of Ganoderma and related genera mainly of the Bogor and Leiden Herbaria. Persoonia-Mol. Phylogeny Evol. Fungi 1972, 7, 55–118. [Google Scholar]
- Ryvarden, L. Synopsis Fungi, Genera of Polypores: Nomenclature and Taxonomy; Fungiflora A/S: Oslo, Norway, 1991; Volume 5, pp. 1–363. [Google Scholar]
- Corner, E.J.H. Amauroderma and Ganoderma; Cramer: Berlin, Germany, 1983. [Google Scholar]
- Wu, S.-H.; Zhang, X. The finding of three Ganodermataceae species in Taiwan. Collect. Res. 2003, 16, 61–66. [Google Scholar]
- Moncalvo, J.; Ryvarden, L. A nomenclatural study of the Ganodermataceae Donk; Fungiflora: Oslo, Norway, 1997; pp. 107–114. [Google Scholar]
- Hong, S.G.; Jung, H.S. Phylogenetic analysis of Ganoderma based on nearly complete mitochondrial small-subunit ribosomal DNA sequences. Mycologia 2004, 96, 742–755. [Google Scholar] [CrossRef]
- Ofodile, L.N.; Uma, N.; Kokubun, T.; Grayer, R.J.; Ogundipe, O.; Simmons, M.S. Antimicrobial colossolactones from a Nigerian polypore Ganoderma colossum (Fr.) CF Baker. Int. J. Med. Mushrooms 2005, 7, 437–438. [Google Scholar] [CrossRef]
- Patoullard, N. Contributions à la flore mycologique du Tonkin. J. Bot. 1897, 11, 367–370. [Google Scholar]
- Ryvarden, L. Neotropical polypores: Part 1: Introduction, Ganodermataceae & Hymenochaetaceae; Fungiflora: Oslo, Norway, 2004; p. 277. [Google Scholar]
- Abdel-Azeem, A.; Nafady, N. New records on the genus Tomophagus and Battarrea for mycobiota of Egypt. Curr. Res. Environ. Appl. Mycol. 2019, 9, 77–84. [Google Scholar] [CrossRef]
- Dzubak, P.; Hajduch, M.; Vydra, D.; Hustova, A.; Kvasnica, M.; Biedermann, D.; Markova, L.; Urban, M.; Sarek, J. Pharmacological activities of natural triterpenoids and their therapeutic implications. Nat. Prod. Rep. 2006, 23, 394–411. [Google Scholar] [CrossRef]
- Liby, K.T.; Yore, M.M.; Sporn, M.B. Triterpenoids and rexinoids as multifunctional agents for the prevention and treatment of cancer. Nat. Rev. Cancer 2007, 7, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Kleinwächter, P.; Anh, N.; Kiet, T.T.; Schlegel, B.; Dahse, H.-M.; Härtl, A.; Gräfe, U. Colossolactones, new triterpenoid metabolites from a Vietnamese mushroom Ganoderma Colossum. J. Nat. Prod. 2001, 64, 236–239. [Google Scholar]
- El Dine, R.S.; El Halawany, A.M.; Ma, C.-M.; Hattori, M. Anti-HIV-1 protease activity of lanostane triterpenes from the vietnamese mushroom Ganoderma Colossum. J. Nat. Prod. 2008, 71, 1022–1026. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Onions, A.H. The Preservation and Maintenance of Living Fungi; CAB International: Egham, UK, 1994; p. 122. [Google Scholar]
- Moubasher, A.H.; Ismail, M.A.; Al-Bedak, O.A.; Mohamed, R.A. Ramophialophora chlamydospora, a new species from an alkaline lake ofWadi-El-Natron, Egypt. Asian J. Mycol. 2019, 2, 110–117. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Al-Bedak, O.A.; Moubasher, A.H. Aspergillus gaarensis, a new addition to section Circumdati from soil of Lake El-Gaar in Wadi-El-Natron, Egypt. Stud. Fungi 2020, 5, 59–65. [Google Scholar] [CrossRef]
- Al-Bedak, O.A.; Teama, E.; Ali, E.; Said, M.; Shalaby, E.; Moharram, Z. Impact of fumigation with phosphine on viability of wheat grains stored for six months at two levels of moisture content, in addition to description of four new records of associated fungi and assessment of their potential for enzymatic production. J. Basic Appl. Mycol. 2020, 11, 77–97. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Criscuolo, A.; Gribaldo, S. BMGE (Block Mapping and Gathering with Entropy): A new software for selection of phylogenetic informative regions from multiple sequence alignments. BMC Evol. Biol. 2010, 10, 210. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.Y.; Lim, T.T.; Tee, J.J. Antioxidant properties of guava fruit: Comparison with some local fruits. Sunway Acad. J. 2006, 3, 9–20. [Google Scholar]
- Maurya, S.; Singh, D. Quantitative analysis of total phenolic content in Adhatoda vasica Nees extracts. Int. J. PharmTech Res. 2010, 2, 2403–2406. [Google Scholar]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.-E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Ramadan, A.M.; Shehata, R.M.; El-Sheikh, H.H.; Ameen, F.; Stephenson, S.L.; Zidan, S.A.; Al-Bedak, O.A. Exploitation of Sugarcane Bagasse and Environmentally Sustainable Production, Purification, Characterization, and Application of Lovastatin by Aspergillus terreus AUMC 15760 under Solid-State Conditions. Molecules 2023, 28, 4048. [Google Scholar] [CrossRef]
- Yen, G.C.; Duh, P.D. Scavenging effect of methanolic extracts of peanut hulls on free-radical and active-oxygen species. J. Agric. Food Chem. 1994, 42, 629–632. [Google Scholar] [CrossRef]
- Skehan, P.; Storeng, R.; Scudiero, D.; Monks, A.; McMahon, J.; Vistica, D.; Warren, J.T.; Bokesch, H.; Kenney, S.; Boyd, M.R. New colorimetric cytotoxicity assay for anticancer-drug screening. JNCI J. Natl. Cancer Inst. 1990, 82, 1107–1112. [Google Scholar] [CrossRef]
- Allam, R.M.; Al-Abd, A.M.; Khedr, A.; Sharaf, O.A.; Nofal, S.M.; Khalifa, A.E.; Mosli, H.A.; Abdel-Naim, A.B. Fingolimod interrupts the cross talk between estrogen metabolism and sphingolipid metabolism within prostate cancer cells. Toxicol. Lett. 2018, 291, 77–85. [Google Scholar] [CrossRef]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef]
- Bland, J.M.; Altman, D. Statistical methods for assessing agreement between two methods of clinical measurement. Lancet 1986, 327, 307–310. [Google Scholar] [CrossRef]
- Deyo, R.A.; Diehr, P.; Patrick, D.L. Reproducibility and responsiveness of health status measures statistics and strategies for evaluation. Control. Clin. Trials 1991, 12, S142–S158. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.B.; Nielsen, S.E.; Berg, K. Re-examination and further development of a precise and rapid dye method for measuring cell growth/cell kill. J. Immunol. Methods 1989, 119, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Kalra, K.; Chauhan, R.; Shavez, M.; Sachdeva, S. Isolation of Laccase Producing Trichoderma Spp. and Effect. Int. J. ChemTech Res. 2013, 5, 2229–2235. [Google Scholar]
- Abd El Monssef, R.A.; Hassan, E.A.; Ramadan, E.M. Production of laccase enzyme for their potential application to decolorize fungal pigments on aging paper and parchment. Ann. Agric. Sci. 2016, 61, 145–154. [Google Scholar] [CrossRef]
- Lowry, O.; Rosebrough, N.; Farr, A.; Randall, R. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- AL-Kolaibe, A.M.; Moharram, A.M.; Al-Bedak, O.A. Worthwhile enzyme production and eco-friendly bioconversion of three agricultural residues by Aspergillus curvatus and Aspergillus gaarensis, promising enzyme-producers isolated from extreme environment. J. Basic Appl. Mycol. 2021, 12, 1–14. [Google Scholar]
- Douanla-Meli, C.; Langer, E. Ganoderma carocalcareus sp. nov., with crumbly-friable context parasite to saprobe on Anthocleista nobilis and its phylogenetic relationship in G. resinaceum group. Mycol. Prog. 2009, 8, 145–155. [Google Scholar] [CrossRef]
- Ryvarden, L. Type studies in the Polyporaceae 17. Species described by WA Murrill. Mycotaxon 1995, 23, 169–198. [Google Scholar]
- Hapuarachchi, K.; Cheng, C.R.; Wen, T.C.; Jeewon, R.; Kakumyan, P. Mycosphere Essays 20: Therapeutic potential of Ganoderma species: Insights into its use as traditional medicine. Mycosphere 2017, 8, 1653–1694. [Google Scholar] [CrossRef]
- El Dine, R.S.; El Halawany, A.M.; Ma, C.M.; Hattori, M. Inhibition of the dimerization and active site of HIV-1 protease by secondary metabolites from the Vietnamese mushroom Ganoderma Colossum. J. Nat. Prod. 2009, 72, 2019–2023. [Google Scholar] [CrossRef] [PubMed]
- Isaka, M.; Chinthanom, P.; Kongthong, S.; Srichomthong, K.; Choeyklin, R. Lanostane triterpenes from cultures of the Basidiomycete Ganoderma orbiforme BCC 22324. Phytochemistry 2013, 87, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Paterson, R.R.M. Ganoderma–a therapeutic fungal biofactory. Phytochemistry 2006, 67, 1985–2001. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.-C.; Yang, Z.-L.; Cui, B.-K.; Yu, C.-J.; Zhou, L.-W. Species diversity and utilization of medicinal mushrooms and fungi in China. Int. J. Med. Mushrooms 2009, 11, 287–302. [Google Scholar] [CrossRef]
- Cheng, C.R.; Yang, M.; Wu, Z.Y.; Wang, Y.; Zeng, F.; Wu, W.Y.; Guan, S.H.; Guo, D.A. Fragmentation pathways of oxygenated tetracyclic triterpenoids and their application in the qualitative analysis of Ganoderma lucidum by multistage tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2011, 25, 1323–1335. [Google Scholar] [CrossRef]
- Cao, Y.; Yuan, H.-S. Ganoderma mutabile sp. nov. from southwestern China based on morphological and molecular data. Mycol. Prog. 2013, 12, 121–126. [Google Scholar] [CrossRef]
- De Silva, D.D.; Rapior, S.; Fons, F.; Bahkali, A.H.; Hyde, K.D. Medicinal mushrooms in supportive cancer therapies: An approach to anti-cancer effects and putative mechanisms of action. Fungal Divers. 2012, 55, 1–35. [Google Scholar] [CrossRef]
- De Silva, D.D.; Rapior, S.; Hyde, K.D.; Bahkali, A.H. Medicinal mushrooms in prevention and control of diabetes mellitus. Fungal Divers. 2012, 56, 1–29. [Google Scholar] [CrossRef]
- De Silva, D.D.; Rapior, S.; Sudarman, E.; Stadler, M.; Xu, J.; Aisyah Alias, S.; Hyde, K.D. Bioactive metabolites from macrofungi: Ethnopharmacology, biological activities and chemistry. Fungal Divers. 2013, 62, 1–40. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, Z.; Cheng, C.; Qing, F. Synthesis and Antitumor Activity Evaluation of γ-Monofluorinated and γ, γ-Difluorinated Goniothalamin Analogues. Chin. J. Chem. 2013, 31, 805–812. [Google Scholar] [CrossRef]
- Hapuarachchi, K.; Wen, T.; Jeewon, R.; Wu, X.; Kang, J.; Hyde, K. Mycosphere Essays 7. Ganoderma lucidum-are the beneficial anti-cancer properties substantiated? Mycosphere 2016, 7, 305–332. [Google Scholar]
- Baby, S.; Johnson, A.J.; Govindan, B. Secondary metabolites from Ganoderma. Phytochemistry 2015, 114, 66–101. [Google Scholar] [CrossRef]
- Bishop, K.S.; Kao, C.H.; Xu, Y.; Glucina, M.P.; Paterson, R.R.M.; Ferguson, L.R. From 2000 years of Ganoderma lucidum to recent developments in nutraceuticals. Phytochemistry 2015, 114, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-H.; Lin, Y.-S.; Lin, K.-L.; Lu, Y.-L.; Chen, C.-H.; Chien, M.-Y.; Shang, H.-F.; Lin, S.-Y.; Hou, W.-C. Effects of hot-water extracts from Ganoderma lucidum residues and solid-state fermentation residues on prebiotic and immune-stimulatory activities in vitro and the powdered residues used as broiler feed additives in vivo. Bot. Stud. 2015, 56, 17. [Google Scholar] [CrossRef]
- Gąsecka, M.; Mleczek, M.; Siwulski, M.; Niedzielski, P.; Kozak, L. Phenolic and flavonoid content in Hericium erinaceus, Ganoderma lucidum, and Agrocybe aegerita under selenium addition. Acta Aliment. 2016, 45, 300–308. [Google Scholar] [CrossRef]
- Yalcin, O.U.; Sarikurkcu, C.; Cengiz, M.; Gungor, H.; Gungor, H.; Ćavar Zeljković, S. Ganoderma carnosum and Ganoderma pfeifferi: Metal concentration, phenolic content, and biological activity. Mycologia 2020, 112, 1–8. [Google Scholar] [CrossRef]
- Gil-Ramírez, A.; Pavo-Caballero, C.; Baeza, E.; Baenas, N.; Garcia-Viguera, C.; Marín, F.R.; Soler-Rivas, C. Mushrooms do not contain flavonoids. J. Funct. Foods 2016, 25, 1–13. [Google Scholar] [CrossRef]
- Ferreira, I.C.; Barros, L.; Abreu, R. Antioxidants in wild mushrooms. Curr. Med. Chem. 2009, 16, 1543–1560. [Google Scholar] [CrossRef]
- Palacios, I.; Lozano, M.; Moro, C.; D’Arrigo, M.; Rostagno, M.A.; Martínez, J.A.; García-Lafuente, A.; Guillamón, E.; Villares, A. Antioxidant properties of phenolic compounds occurring in edible mushrooms. Food Chem. 2011, 128, 674–678. [Google Scholar] [CrossRef]
- Stanikunaite, R.; Khan, S.I.; Trappe, J.M.; Ross, S.A. Cyclooxygenase-2 inhibitory and antioxidant compounds from the truffle Elaphomyces granulatus. Phytother. Res. 2009, 23, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Muszyńska, B.; Grzywacz-Kisielewska, A.; Kala, K.; Gdula-Argasińska, J. Anti-inflammatory properties of edible mushrooms: A review. Food Chem. 2018, 243, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Tapwal, A.; Pandey, S.; Borah, R.K.; Borah, D.; Borgohain, J. Macro-fungal diversity and nutrient content of some edible mushrooms of Nagaland, India. Nusant. Biosci. 2013, 5, 1–7. [Google Scholar] [CrossRef]
- Ghazala, I.; Haddar, A.; Romdhane, M.B.; Ellouz-Chaanouni, S. Screening and molecular identification of new microbial strains for production of enzymes of biotechnological interest. Braz. Arch. Biol. Technol. 2016, 59, 1–12. [Google Scholar] [CrossRef]
- Karthikeyan, V.; Ragunathan, R.; Johney, J.; Kabesh, K. Production, optimization and purification of laccase produced by Pleurotus ostreatus MH591763. Res. Rev. A J. Microbiol. Virol. 2019, 9, 56–64. [Google Scholar]
- Torres-Farradá, G.; Manzano Leon, A.M.; Rineau, F.; Ledo Alonso, L.L.; Sánchez-López, M.I.; Thijs, S.; Colpaert, J.; Ramos-Leal, M.; Guerra, G.; Vangronsveld, J. Diversity of ligninolytic enzymes and their genes in strains of the genus Ganoderma: Applicable for biodegradation of xenobiotic compounds? Front. Microbiol. 2017, 8, 898. [Google Scholar] [CrossRef]
- Aftab, A.; Siddique, I.; Aftab, M.; Yousaf, Z.; Chaudhry, F.A. Wood degrading mushrooms potentially strong towards laccase biosynthesis in pakistan. Wood Res. 2020, 65, 809–818. [Google Scholar] [CrossRef]
- Elissetche, J.-P.; Ferraz, A.; Freer, J.; Rodriguez, J. Enzymes produced by Ganoderma australe growing on wood and in submerged cultures. World J. Microbiol. Biotechnol. 2007, 23, 429–434. [Google Scholar] [CrossRef]
- Alshiekheid, M.A.; Umar, A.; Ameen, F.; Alyahya, S.A.; Dufosse, L. Biodegradation of chromium by laccase action of Ganoderma Multipileum. J. King Saud Univ. Sci. 2023, 35, 102948. [Google Scholar] [CrossRef]
- de Souza Silva, C.M.M.; De Melo, I.S.; De Oliveira, P.R. Ligninolytic enzyme production by Ganoderma spp. Enzym. Microb. Technol. 2005, 37, 324–329. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutritional Component | Value (%) | Nutritional Component | Value (mg/100 g) |
|---|---|---|---|
| Carbohydrates | 28.81 | K | 133.59 |
| Crude Fibers | 23.44 | P | 114.46 |
| Crude Proteins | 20.64 | Ca | 6.27 |
| Crude Fats | 25.34 | Se | 3.08 |
| Ash | 3.02 | Fe | 1.28 |
| Zn | 0.73 | ||
| Total phenolic compounds | 3926 | ||
| Total flavonoids | 562 |
| Retention Time (min) | Area (mV.s) | Compound Name | Amount (mg/100 g Dry wt.) |
|---|---|---|---|
| 9.061 | 13,936.801 | Aspartic acid | 2.727 |
| 11.171 | 5248.426 | Threonine | 1.371 |
| 11.933 | 6686.706 | Serine | 1.390 |
| 14.325 | 31,964.8 | Glutamic acid | 6.801 |
| 16.272 | 76.739 | Proline | 0.691 |
| 20.347 | 10,087.029 | Glycine | 1.256 |
| 22.013 | 12,613.589 | Alanine | 5.556 |
| 23.376 | 7872.286 | Valine | 1.388 |
| 25.669 | 1234.684 | Methionine | 0.282 |
| 28.229 | 3966.965 | Isoleucine | 0.932 |
| 29.651 | 8677.939 | Leucine | 1.945 |
| 31.560 | 2391.457 | Tyrosine | 0.732 |
| 32.808 | 4518.379 | Phenylalanine | 1.241 |
| 35.373 | 23,137.48 | Histidine | 4.558 |
| 39.749 | 8056.973 | Lysine | 1.714 |
| 43.747 | 5677.821 | Arginine | 14.396 |
| Total | 174,933.787 | 16 amino acids | 46.98 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Bedak, O.A.-H.M.; Moharram, A.M.; Abdel-Raheam, H.E.-D.F.; Stephenson, S.L.; Ameen, F. Nutritional Composition, Antioxidant Activity, Cytotoxicity, and Enzymatic Potential of Ficus nitida-Associated Tomophagus colossus. Agronomy 2023, 13, 2850. https://doi.org/10.3390/agronomy13112850
Al-Bedak OA-HM, Moharram AM, Abdel-Raheam HE-DF, Stephenson SL, Ameen F. Nutritional Composition, Antioxidant Activity, Cytotoxicity, and Enzymatic Potential of Ficus nitida-Associated Tomophagus colossus. Agronomy. 2023; 13(11):2850. https://doi.org/10.3390/agronomy13112850
Chicago/Turabian StyleAl-Bedak, Osama Abdel-Hafeez Mohamed, Ahmed Mohamed Moharram, Hossam El-Dean Farghaly Abdel-Raheam, Steven L. Stephenson, and Fuad Ameen. 2023. "Nutritional Composition, Antioxidant Activity, Cytotoxicity, and Enzymatic Potential of Ficus nitida-Associated Tomophagus colossus" Agronomy 13, no. 11: 2850. https://doi.org/10.3390/agronomy13112850
APA StyleAl-Bedak, O. A.-H. M., Moharram, A. M., Abdel-Raheam, H. E.-D. F., Stephenson, S. L., & Ameen, F. (2023). Nutritional Composition, Antioxidant Activity, Cytotoxicity, and Enzymatic Potential of Ficus nitida-Associated Tomophagus colossus. Agronomy, 13(11), 2850. https://doi.org/10.3390/agronomy13112850