Identification of Promising Three-Way Hybrids of Pearl Millet for Drought-Prone Environments of North-Western India

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Field Evaluation
2.3. Statistical Analysis
3. Results
3.1. Analysis of Variance and Mean Performance
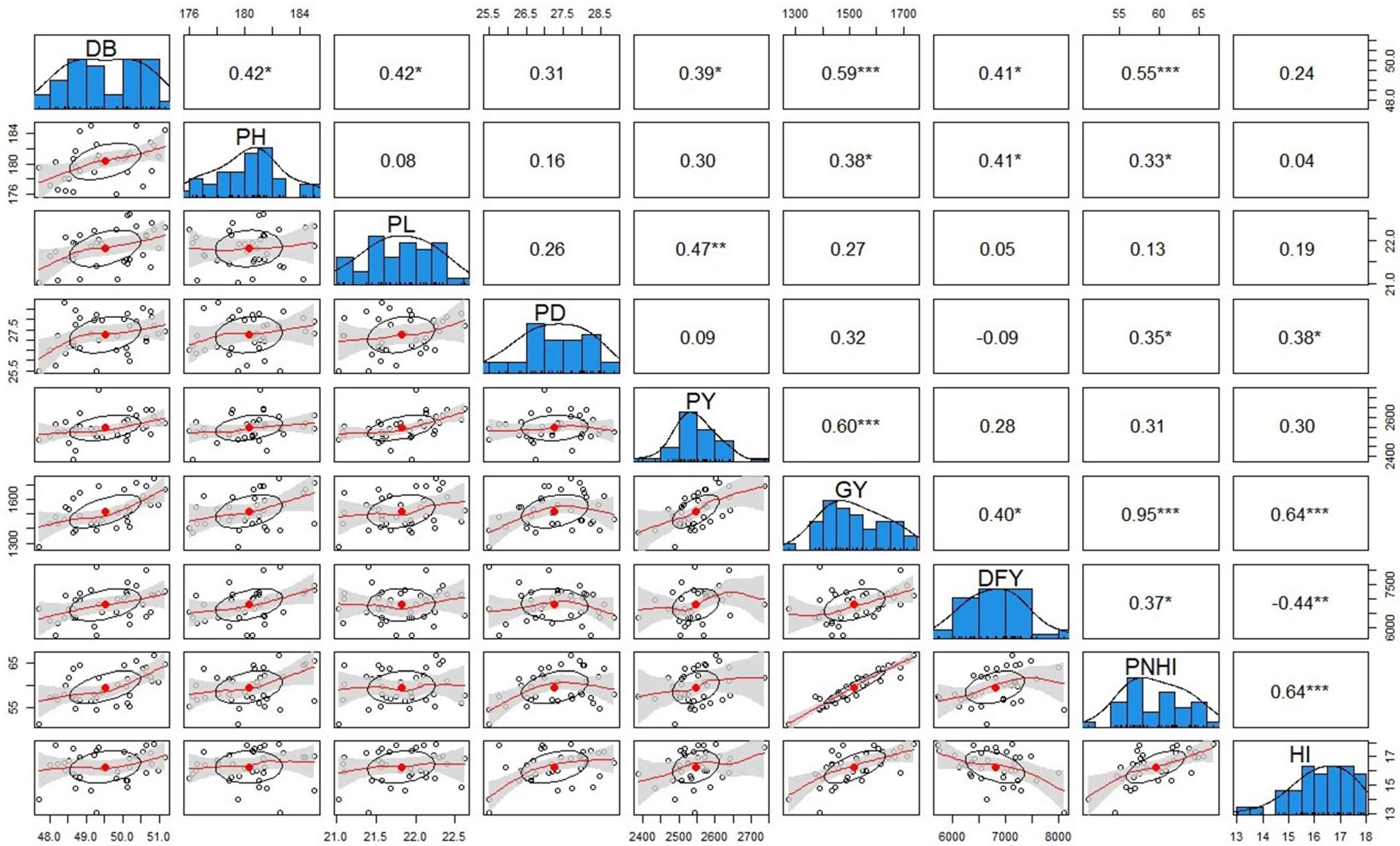
3.2. Trait Association
3.3. Standard Heterosis
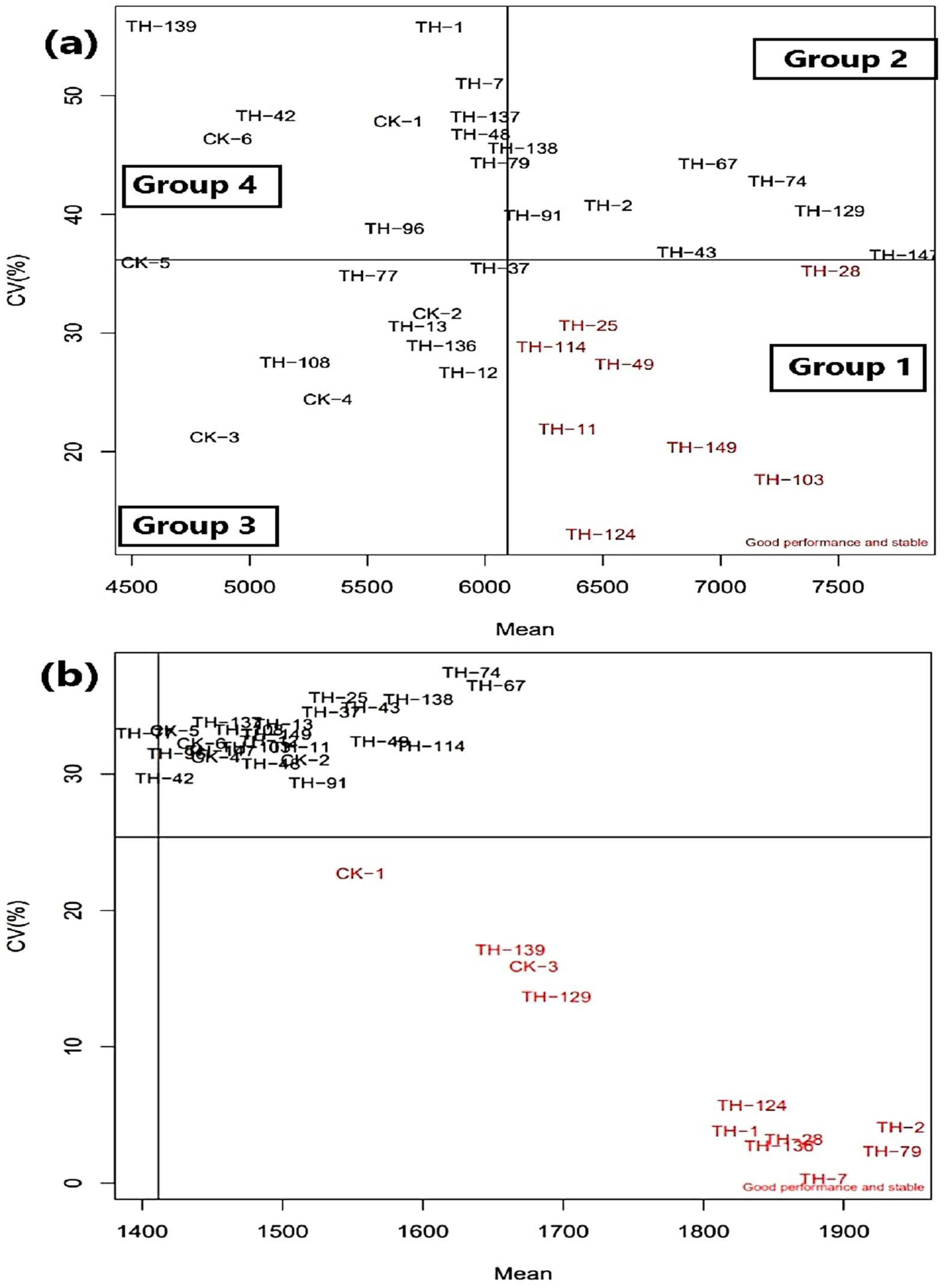
3.4. Stability Performance Visualized through CV vs. Mean Plot
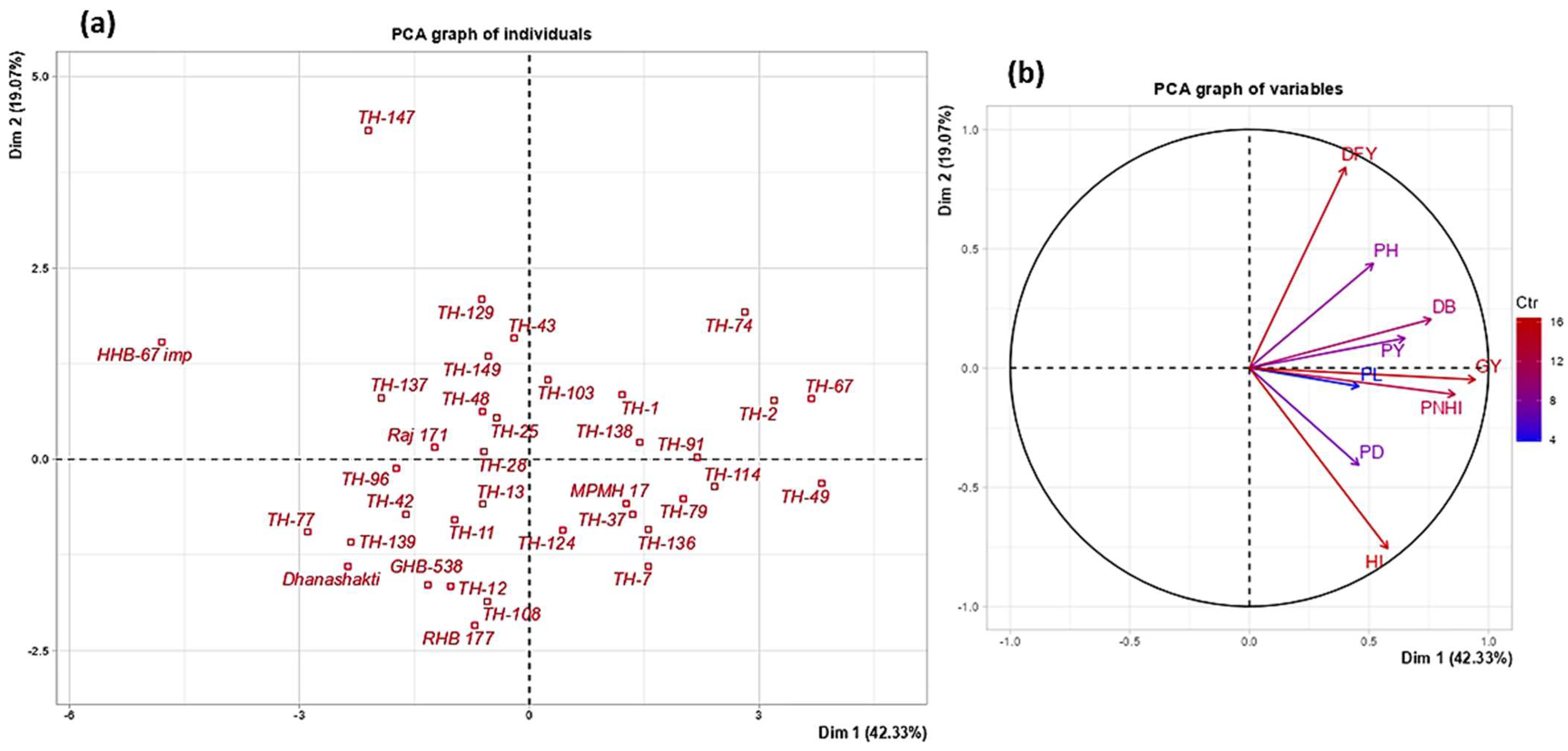
3.5. PCA and Dendrogram
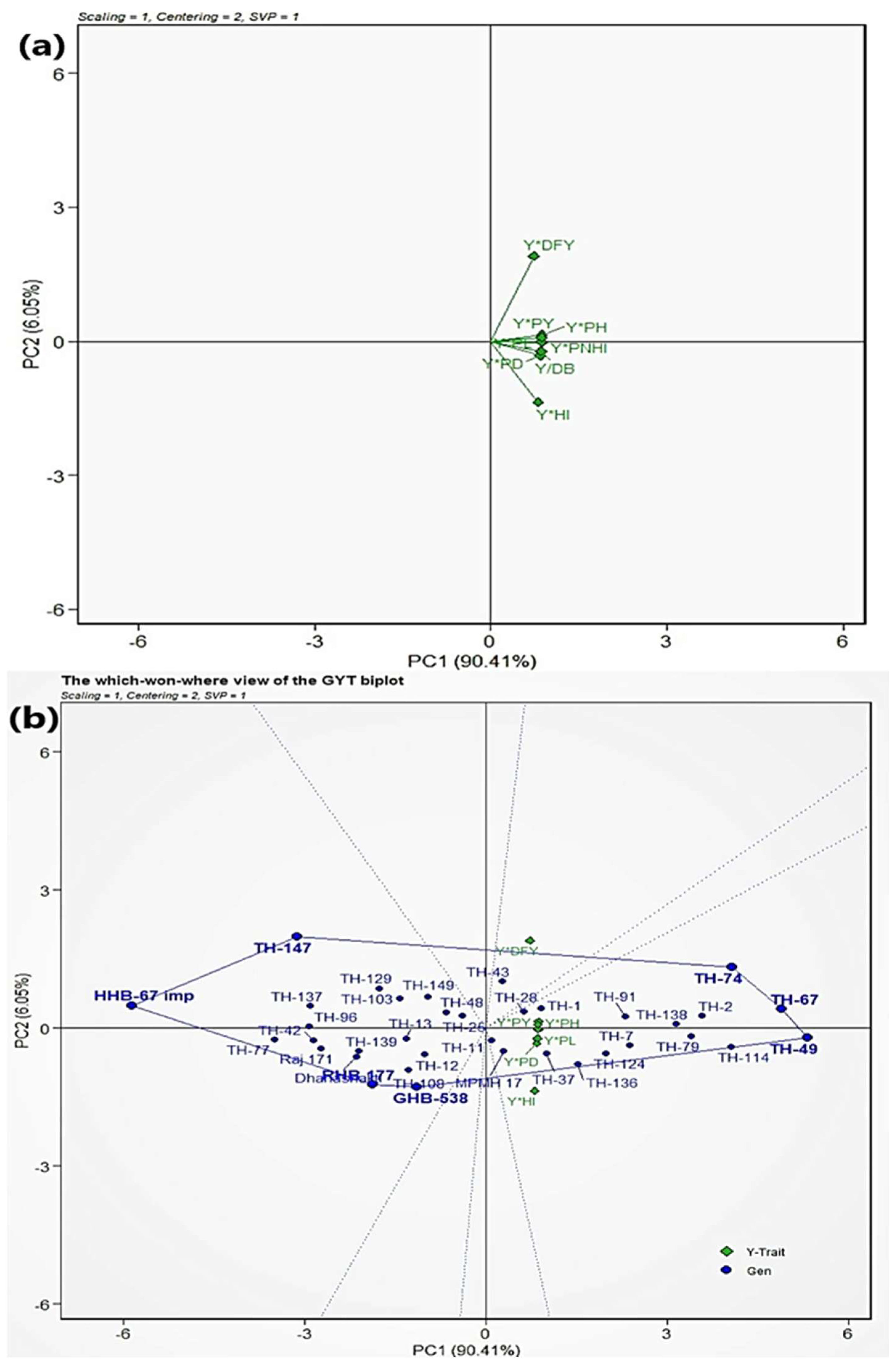
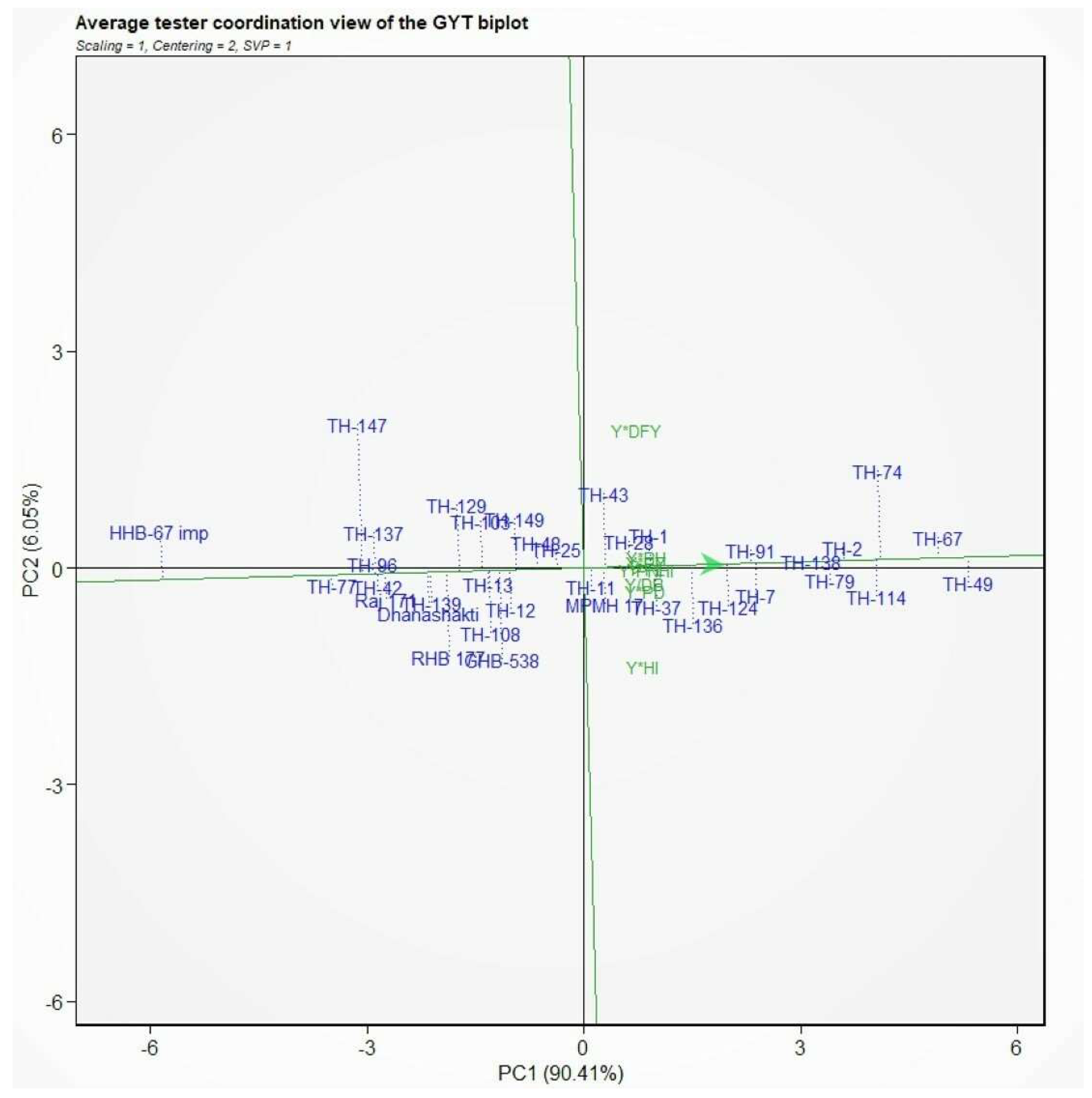
3.6. Genotype by Yield × Trait (GYT) Biplot
3.7. Identification of Most Promising Three-Way Hybrids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jukanti, A.K.; Gowda, C.L.L.; Rai, K.N.; Manga, V.K.; Bhatt, R.K. Crops That Feed the World 11. Pearl Millet (Pennisetum glaucum L.): An Important Source of Food Security, Nutrition and Health in the Arid and Semi-Arid Tropics. Food Secur. 2016, 8, 307–329. [Google Scholar] [CrossRef]
- Sanjana Reddy, P.; Satyavathi, C.T.; Khandelwal, V.; Patil, H.T.; Gupta, P.C.; Sharma, L.D.; Mungra, K.D.; Singh, S.P.; Narasimhulu, R.; Bhadarge, H.H.; et al. Performance and Stability of Pearl Millet Varieties for Grain Yield and Micronutrients in Arid and Semi-Arid Regions of India. Front. Plant Sci. 2021, 12, 670201. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, R.; Gangashetty, P.I.; Karimoune, L.; Ba, N.M. Genetic Variation and Diversity of Pearl Millet [Pennisetum glaucum (L.)] Genotypes Assessed for Millet Head Miner, Heliocheilus Albipunctella Resistance, in West Africa. Euphytica 2020, 216, 158. [Google Scholar] [CrossRef]
- Yadav, O.P.; Rai, K.N.; Khairwal, I.S.; Rajpurohit, B.S.; Mahala, R.S. Breeding Pearl Millet for Arid Zone of North-Western India: Constraints, Opportunities and Approaches; All India Coordinated Pearl Millet Improvement Project (AICPMIP-ICAR): Jodhpur, India, 2011.
- Yadav, O.P.; Rai, K.N. Genetic Improvement of Pearl Millet in India. Agric. Res. 2013, 2, 275–292. [Google Scholar] [CrossRef]
- Yadav, O.P.; Singh, D.V.; Vadez, V.; Gupta, S.K.; Rajpurohit, B.S.; Shekhawat, P.S. Improving Pearl Millet for Drought Tolerance- Retrospect and Prospects. Indian J. Genet. Plant Breed. 2017, 77, 464–474. [Google Scholar] [CrossRef]
- Vadez, V.; Hash, T.; Bidinger, F.R.; Kholova, J., II. 1.5 Phenotyping Pearl Millet for Adaptation to Drought. Front. Physiol. 2012, 3, 1–12. [Google Scholar] [CrossRef]
- Carberry, P.S.; Campbell, L.C.; Bidinger, F.R. The growth and development of pearl millet as affected by plant population. Field Crop. Res. 1985, 11, 193–205. [Google Scholar] [CrossRef]
- Howarth, C.J.; Rattunde, E.W.; Bidinger, F.R.; Harris, D. Seedling Survival of Abiotic Stress: Sorghum and Pearl Millet. In Proceedings of the International Conference on Genetic Enhancement of Sorghum and Pearl Millet, Lubbock, TX, USA, 22–27 September 1996; ICAR: New Delhi, India, 1996; pp. 379–399. [Google Scholar]
- Yadav, O.P.; Rajpurohit, G.R.; Kumar, A. Prospects of Enhancing Pearl Millet (Pennisetum Glaucum)Productivity under Drought Environments of North-Western India through Hybrids. Indian J. Genet. Plant Breed. 2012, 72, 25–30. [Google Scholar]
- Lahiri, A.N.; Kumar, V. Studies on Plant Water Relationship III. Further Studies on the Drought Mediated Alterations in the Performance of Bulrush Millet. Proc. Natl. Acad. Sci. India B 1996, 32, 116–129. [Google Scholar]
- Kholová, J.; Hash, C.T.; Kakkera, A.; Koová, M.; Vadez, V. Constitutive Water-Conserving Mechanisms Are Correlated with the Terminal Drought Tolerance of Pearl Millet [Pennisetum glaucum (L.) R. Br.]. J. Exp. Bot. 2010, 61, 369–377. [Google Scholar] [CrossRef]
- Medina, S.; Gupta, S.K.; Vadez, V. Transpiration Response and Growth in Pearl Millet Parental Lines and Hybrids Bred for Contrasting Rainfall Environments. Front. Plant Sci. 2017, 8, 1846. [Google Scholar] [CrossRef] [PubMed]
- Van Oosterom, E.J.; Weltzien, E.; Yadav, O.P.; Bidinger, F.R. Grain Yield Components of Pearl Millet under Optimum Conditions Can Be Used to Identify Germplasm with Adaptation to Arid Zones. Field Crop. Res. 2006, 96, 407–421. [Google Scholar] [CrossRef]
- Bidinger, F.R.; Mahalakshmi, V.; Rao, G.D.P. Assessment of Drought Resistance in Pearl Millet [Pennisetum americanum (L.) Leeke). I Factors Affecting Yields under Stress. Aust. J. Agric. Res. 1987, 38, 37–48. [Google Scholar] [CrossRef]
- Mahalakshmi, V.; Bidinger, F.R.; Raju, D.S. Effect of Timing of Water Deficit on Pearl Millet (Pennisetum americanum). Field Crop. Res. 1987, 15, 327–339. [Google Scholar] [CrossRef]
- Fussel, L.K.; Bidinger, F.R.; Bieler, P. Crop Physiology and Breeding for Drought Tolerance: Research and Development. Field Crop. Res. 1991, 27, 183–199. [Google Scholar] [CrossRef]
- Srivastava, R.K.; Yadav, O.P.; Kaliamoorthy, S.; Gupta, S.K.; Serba, D.D.; Choudhary, S.; Govindaraj, M.; Kholová, J.; Murugesan, T.; Satyavathi, C.T.; et al. Breeding Drought-Tolerant Pearl Millet Using Conventional and Genomic Approaches: Achievements and Prospects. Front. Plant Sci. 2022, 13, 781524. [Google Scholar] [CrossRef]
- Rai, K.N.; Gupta, S.K.; Govindaraj, M.; Yadav, H.P. Indian Farming; Indian Council of Agricultural Research (ICAR): New Delhi, Indaia, 2015; pp. 19–22. [Google Scholar]
- Tom Hash, C.; Sharma, A.; Kolesnikova-Allen, M.A.; Singh, S.D.; Bhasker Raj, A.G.; Ratnaji Rao, M.N.V.; Nijhawan, D.C.; Beniwal, C.R.; Sagar, P.; Yadav, H.P.; et al. Teamwork Delivers Biotechnology Products to Indian Small-Holder Crop-Livestock Producers: Pearl Millet Hybrid “HHB 67 Improved” Enters Seed Delivery Pipeline. SAT eJ. 2006, 2. Available online: http://oar.icrisat.org/id/eprint/2738 (accessed on 25 June 2023).
- Izge, A.U.; Dugje, I.Y. Performance of Drought Tolerant Three-Way and Top Cross Maize Hybrids in Sudan Savanna of North Eastern Nigeria. J. Plant Breed. Crop Sci. 2011, 3, 269–275. [Google Scholar]
- Meseka, S.; Menkir, A.; Bossey, B.; Mengesha, W. Performance Assessment of Drought Tolerant Maize Hybrids under Combined Drought and Heat Stress. Agronomy 2018, 8, 274. [Google Scholar] [CrossRef]
- Amole, T.; Adekeye, A.; Bakare, B.; Meseka, S.; Mengesha, W.; Menkir, A. Assessing the Fodder Potentials of Drought-Tolerant Maize (Zea mays L.) Hybrids in West Africa. Int. J. Agron. 2022, 2022, 1348711. [Google Scholar] [CrossRef]
- Syngenta Foundation. Affordable, Accessible, Asian (“AAA”) Drought-Tolerant Maize Overview of a Collaborative Program. between CIMMYT and Syngenta, Supported and Coordinated by Syngenta Foundation for Sustainable Agriculture; Syngenta Foundation: Basel, Switzerland, 2021. [Google Scholar]
- Hohls, T. Analysis of Genotype-Environment Interactions. S Afr. J. Sci. 1995, 91, 121–124. [Google Scholar]
- Kandel, M.; Ghimire, S.K.; Ojha, B.R.; Shrestha, J. Cluster Analysis among the Maize Inbred Lines (Zea mays L.) under Heat Stress Condition. Int. J. Glob. Sci. Res. 2018, 5. [Google Scholar] [CrossRef]
- Yan, W.; Frégeau-Reid, J. Genotype by Yield∗Trait (GYT) Biplot: A Novel Approach for Genotype Selection Based on Multiple Traits. Sci. Rep. 2018, 8, 8242. [Google Scholar] [CrossRef]
- Sparks, A. Nasapower: A NASA POWER Global Meteorology, Surface Solar Energy and Climatology Data Client for R. J. Open Source Softw. 2018, 3, 1035. [Google Scholar] [CrossRef]
- Hajjarpoor, A.; Vadez, V.; Soltani, A.; Gaur, P.; Whitbread, A.; Suresh Babu, D.; Gumma, M.K.; Diancoumba, M.; Kholová, J. Characterization of the Main Chickpea Cropping Systems in India Using a Yield Gap Analysis Approach. Field Crop. Res. 2018, 223, 93–104. [Google Scholar] [CrossRef]
- Ronanki, S.; Pavlík, J.; Masner, J.; Jarolímek, J.; Stočes, M.; Subhash, D.; Talwar, H.S.; Tonapi, V.A.; Srikanth, M.; Baddam, R.; et al. An APSIM-Powered Framework for Post-Rainy Sorghum-System Design in India. Field Crop. Res. 2022, 277, 108422. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Revelle, W. Package “Psych” Procedures for Psychological, Psychometric, and Personality Research; R Package Version 2.3.9; Northwestern University: Evanston, IL, USA, 2023; p. 469. Available online: https://CRAN.R-project.org/package=psych (accessed on 25 June 2023).
- Meredith, W.R.; Bridge, R.R. Heterosis and Gene Action in Cotton, Gossypium hirsutum L. Crop Sci. 1972, 12, 304–310. [Google Scholar] [CrossRef]
- Francis, T.R.; Kannenberg, L.W. Yield Stability Studies in Short-Season Maize. I. A Descriptive Method for Grouping Genotypes. Can. J. Plant Sci. 1978, 58, 1029–1034. [Google Scholar] [CrossRef]
- Pacheco, A.; Vargas, M.; Alvarado, G.; Rodríguez, F.; Crossa, J.; Burgueño, J. GEA-R (Genotype × Environment Analysis with R for Windows), Version 4.1. CIMMYT Research Data & Software Repository Network, V16. 2015. Available online: https://hdl.handle.net/11529/10203 (accessed on 25 June 2023).
- Vaissie, P.; Monge, A.; Husson, F. FactoShiny: Perform Factorial Analysis from ’FactoMineR’ with a Shiny. R Package Version 2.4. 2021. Available online: https://cran.r-project.org/web/packages/Factoshiny/Factoshiny.pdf (accessed on 25 June 2023).
- Olivoto, T.; Lúcio, A.D.C. Metan: An R Package for Multi-Environment Trial Analysis. Methods Ecol. Evol. 2020, 11, 783–789. [Google Scholar] [CrossRef]
- Allard, R.W.; Bradshaw, A.D. Implications of Genotype-Environmental Interactions in Applied Plant Breeding. Crop Sci. 1964, 4, 503–508. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Talabi, A.O.; Ifie, B.E.; Chabi, Y.C.; Obeng-Antwi, K.; Haruna, A.; Asiedu, R. Gains in Grain Yield of Extra-Early Maize during Three Breeding Periods under Drought and Rainfed Conditions. Crop Sci. 2018, 58, 2399–2412. [Google Scholar] [CrossRef] [PubMed]
- Khairwal, I.S.; Yadav, O.P. Pearl Millet (Pennisetum glacum) Improvement in India-Retrospect and Prospects. Indian. J. Agric. Sci. 2005, 75, 183–191. [Google Scholar]
- Gupta, S.K.; Rathore, A.; Yadav, O.P.; Rai, K.N.; Khairwal, I.S.; Rajpurohit, B.S.; Das, R.R. Identifying Mega-Environments and Essential Test Locations for Pearl Millet Cultivar Selection in India. Crop Sci. 2013, 53, 2444–2453. [Google Scholar] [CrossRef]
- Austin, R.B.; Ford, M.A.; Morgan, C.L.; Yeoman, D. Old and Modern Wheat Cultivars Compared on the Broadbalk Wheat Experiment. Eur. J. Agron. 1993, 2, 141–147. [Google Scholar] [CrossRef]
- Yadav, O.; Rai, K.; Yadav, H.; Rajpurohit, B.; Gupta, S.; Rathore, A.; Karjagi, C. Assessment of Diversity in Commercial Hybrids of Pearl Millet in India. Indian. J. Plant Genet. Resour. 2016, 29, 130. [Google Scholar] [CrossRef]
- All India Coordinated Research Project on Pearl Millet (Indian Council of Agricultural Research). In 52nd Annual Group Meeting of All India Coordinated Research Project on Pearl Millet; Indian Council of Agricultural Research: Mandor, India, 2017.
- Changizi, M.; Choukan, R.; Heravan, E.M.; Bihamta, M.R.; Darvish, F. Evaluation of Genotype × Environment Interaction and Stability of Corn Hybrids and Relationship among Univariate Parametric Methods. Can. J. Plant Sci. 2014, 94, 1255–1267. [Google Scholar] [CrossRef]
- Bradshaw, A.D. Evolutionary significance of phenotypic plasticity in plants. Adv. Genet. 1965, 13, 115–155. [Google Scholar]
- Schnell, F.W.; Becker, H.C. Yield and Yield Stability in a Balanced System of Widely Differing Population Structures in Zea mays L. Plant Breed. 1986, 97, 30–38. [Google Scholar] [CrossRef]
- Haussmann, B.I.G.; Obilana, A.B.; Ayiecho, P.O.; Blum, A.; Schipprack, W.; Geiger, H.H. Yield and Yield Stability of Four Population Types of Grain Sorghum in a Semi-Arid Area of Kenya. Crop Sci. 2000, 40, 319–329. [Google Scholar] [CrossRef]
- Fayeun, L.; Sesay, S. Evaluation of Drought Tolerant Top Cross and Three-Way Cross Maize Hybrids for Grain Yield and Related Traits in Three Agro-Ecological Zones of Southwest Nigeria. Not. Sci. Biol. 2019, 11, 414–421. [Google Scholar] [CrossRef]
- Singamsetti, A.; Shahi, J.P.; Zaidi, P.H.; Seetharam, K. Study on Applicability of Genotype × Yield × Trait (GYT) Biplots over Genotype × Trait (GT) Biplots in Selection of Maize Hybrids across Soil Moisture Regimes. Indian. J. Agric. Res. 2022, 56, 1–7. [Google Scholar] [CrossRef]
- Sanni, K.A.; Fawole, I.; Ogunbayo, S.A.; Tia, D.D.; Somado, E.A.; Futakuchi, K.; Sié, M.; Nwilene, F.E.; Guei, R.G. Multivariate Analysis of Diversity of Landrace Rice Germplasm. Crop Sci. 2012, 52, 494–504. [Google Scholar] [CrossRef]
- Chaudhary, S.; Sagar, P.; Hooda, B.K.; Arya, R.K. Multivariate Analysis of Pearl Millet Data to Delineate Genetic Variation. Forage Res. 2015, 40, 201–208. [Google Scholar]
- Gupta, D.; Khandelwal, V. Principal Component Analysis for Yield and Its Attributing Characters of Pearl Millet (Pennisetum glaucum [L.] R.Br.). Ann. Plant Soil. Res. 2022, 24, 408–414. [Google Scholar] [CrossRef]
- Shashibhushan, D.; Sameer Kumar, C.V.; Kondi, R.K.R. Principal Component Analysis for Yield and Yield Related Traits in Pearl Millet Cultivars. Ecol. Environ. Conserv. 2022, 28, S104–S107. [Google Scholar] [CrossRef]
- Pujar, M.; Govindaraj, M.; Gangaprasad, S.; Kanatti, A.; Shivade, H. Genetic Variation and Diversity for Grain Iron, Zinc, Protein and Agronomic Traits in Advanced Breeding Lines of Pearl Millet [Pennisetum Glaucum (L.) R. Br.] for Biofortification Breeding. Genet. Resour. Crop Evol. 2020, 67, 2009–2022. [Google Scholar] [CrossRef]
- Bista, P.; Thapa, S.; Rawal, S.; Dhakal, D.; Joshi, D. Agro-Morphological Characterization and Estimation of Genetic Parameters of Spring Maize Hybrids in the Inner Plains of Far-West Nepal. Int. J. Agron. 2022, 2022, 4806266. [Google Scholar] [CrossRef]
- Gholizadeh, A.; Ghaffari, M. Genotype by Yield* Trait (GYT) Biplot Analysis: A Novel Approach for Phenotyping Sunflower Single Cross Hybrids Based on Multiple Traits. Food Sci. Nutr. 2023, 11, 5928–5937. [Google Scholar] [CrossRef]
- Yan, W.; Tinker, N.A. Biplot Analysis of Multi-Environment Trial Data: Principles and Applications. Can. J. Plant Sci. 2006, 86, 623–645. [Google Scholar] [CrossRef]
- Kendal, E. Comparing Durum Wheat Cultivars by Genotype × Yield × Trait and Genotype × Trait Biplot Method. Chil. J. Agric. Res. 2019, 79, 512–522. [Google Scholar] [CrossRef]
- Yan, W.; Fregeau-Reid, J.; Mountain, N.; Kobler, J. Genotype and Management Evaluation Based on Genotype by Yield*Trait (GYT) Analysis. Crop Breed. Genet. Genom. 2019, 1, e190002. [Google Scholar] [CrossRef]
- Sofi, P.A.; Saba, I.; Ara, A.; Rehman, K. Comparative Efficiency of GY*T Approach Over GT Approach in Genotypic Selection in Multiple Trait Evaluations: Case Study of Common Bean (Phaseolus Vulgaris) Grown Under Temperate Himalayan Conditions. Agric. Res. 2022, 11, 373–381. [Google Scholar] [CrossRef]
- Mohammadi, R. Genotype by Yield*trait Biplot for Genotype Evaluation and Trait Profiles in Durum Wheat. Cereal Res. Commun. 2019, 47, 541–551. [Google Scholar] [CrossRef]
- Rai, K.N.; Chandra, S.; Rao, A.S. Potential Advantages of Male-Sterile F1 Hybrids for Use as Seed Parents of Three-Way Hybrids in Pearl Millet. Field Crop Res. 2000, 68, 173–181. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Latitude | Longitude | Elevation | Average Temperature (°C) | Average Relative Humidity (%) | Rainfall (mm) | |
|---|---|---|---|---|---|---|---|
| Maximum | Minimum | ||||||
| SKRAU, Bikaner, Rajasthan | 28°09′ N | 73°35′ E | 230 | 37.35 | 27.06 | 49.74 | 142.67 |
| Hytech Seeds Pvt. Ltd., Malakhera-Alwar, Rajasthan | 27°38′ N | 76°62′ E | 253 | 35.05 | 23.8 | 79.98 | 399 |
| JK Seeds Pvt. Ltd., Chirkhna-Alwar, Rajasthan | 27°67′ N | 76°69′ E | 253 | 32.16 | 24.76 | 81.84 | 519.66 |
| Source | DB | GY | DFY | PH | PL | PD | PY | PNHI | HI |
|---|---|---|---|---|---|---|---|---|---|
| (Days) | (kg/ha) | (kg/ha) | (cm) | (cm) | (mm) | (kg/ha) | (%) | (%) | |
| Location (σ2ε) | 8.96 ** | 224,934.9 * | 4,151,695.74 * | 196.29 * | 3.14 ** | 12.5 *** | 1,361,283.2 * | 17.67 | 0.50 |
| Genotype (σ2G) | 1.57 ** | 22,862.1 ** | 365,319.84 * | 15.83 | 0.48 | 0.60 | 23,238.44 | 5.12 | 0.00 |
| Genotype × Location (σ2G × E) | 1.9 *** | 8198.61 | 88,211.46 | 17.97 | 0.64 | 1.15 * | 104,670.19 * | 29.81 | 0.68 |
| Residual | 1.92 | 64,296.88 | 1,207,954.43 | 110.81 | 3.15 | 3.52 | 103,208.91 | 58.87 | 8.63 |
| Heritability (h2) | 0.62 | 0.63 | 0.61 | 0.39 | 0.39 | 0.38 | 0.23 | 0.14 | 0.00 |
| Grand mean | 49.52 | 1513.17 | 6082.94 | 180.35 | 21.81 | 25.21 | 2532.1 | 53.96 | 13.81 |
| Coefficient of variation (CV) % | 2.80 | 16.76 | 18.07 | 5.84 | 8.14 | 7.44 | 12.69 | 14.22 | 21.28 |
| LSD | 2.27 | 287.47 | 1211.29 | 14.50 | 1.55 | 2.26 | 397.60 | 6.12 | 2.17 |
| Genotype | GY | DFY | PH | PL | PD | PY | PNHI | HI |
|---|---|---|---|---|---|---|---|---|
| TH-1 | 22.90 * | 8.17 | 0.76 | 3.71 | 7.20 | 3.34 | 18.93 * | 15.02 |
| TH-2 | 32.93 * | 9.52 | 2.72 | 6.06 | 7.55 | 3.37 | 26.86 * | 21.60 * |
| TH-7 | 27.57 * | 2.83 | −1.39 | 3.09 | 12.11 | 2.16 | 24.87 * | 24.28 * |
| TH-11 | 20.18 | 0.50 | −0.84 | 2.38 | 4.65 | −0.67 | 20.99 * | 19.96 * |
| TH-12 | 15.34 | −4.30 | −0.80 | 2.57 | 13.17 | 1.51 | 13.63 * | 18.56 * |
| TH-13 | 14.68 | −1.44 | 0.27 | 4.31 | 9.66 | 0.31 | 14.33 * | 15.79 |
| TH-25 | 18.82 | 5.22 | 0.60 | 0.31 | 6.63 | 0.81 | 17.86 * | 14.23 |
| TH-28 | 21.67 | 7.14 | −1.72 | 4.96 | 3.41 | 3.65 | 17.39 * | 14.58 |
| TH-37 | 22.88 * | −1.35 | 1.82 | 1.68 | 11.45 | 1.07 | 21.58 * | 23.74 * |
| TH-43 | 21.30 | 14.20 | −0.37 | 2.03 | 3.16 | 0.84 | 20.29 * | 9.71 |
| TH-48 | 17.10 | 4.74 | −0.26 | 5.23 | 4.39 | 5.40 | 11.10 | 11.61 |
| TH-49 | 33.77 * | 7.32 | 1.65 | 5.86 | 8.60 | 4.59 | 30.77 * | 28.33 * |
| TH-67 | 34.69 * | 12.56 | 3.09 | 6.26 | 9.48 | 4.97 | 28.31 * | 21.90 * |
| TH-74 | 32.84 * | 20.64 * | 1.03 | 5.29 | 6.32 | 6.13 | 25.16 * | 13.84 |
| TH-79 | 30.74 * | 5.75 | 0.60 | 2.37 | 9.83 | 3.53 | 26.28 * | 24.35 * |
| TH-91 | 28.81 * | 7.82 | −0.25 | 6.55 | 10.26 | 6.09 | 19.54 * | 18.13 * |
| TH-96 | 10.07 | −1.54 | −1.99 | 5.81 | 3.99 | 1.84 | 8.08 | 10.76 |
| TH-108 | 14.65 | −9.15 | −0.37 | 5.75 | 9.74 | 1.18 | 13.31 * | 22.40 * |
| TH-124 | 26.17 * | 0.06 | 0.64 | 2.56 | 5.44 | 1.19 | 24.69 * | 25.71 * |
| TH-129 | 12.78 | 9.88 | 3.06 | 3.99 | 7.20 | 2.16 | 10.40 | 4.64 |
| TH-138 | 31.36 * | 8.15 | 2.64 | 0.33 | 10.01 | 2.58 | 26.11 * | 21.32 * |
| TH-136 | 24.91 * | −3.77 | 1.06 | 5.14 | 5.62 | 2.60 | 21.74 * | 27.50 * |
| TH-139 | 12.61 | −5.85 | −1.32 | 2.88 | 4.04 | 0.56 | 11.99 | 17.43 * |
| TH-147 | 9.13 | 22.13 * | 1.69 | 1.76 | 0.00 | 2.28 | 6.69 | -6.51 |
| TH-149 | 16.80 | 8.89 | 0.83 | 2.60 | 4.48 | −0.24 | 17.08 * | 9.77 |
| TH-42 | 8.99 | −4.35 | 0.05 | 4.23 | 11.59 | 1.76 | 7.10 | 12.00 |
| TH-114 | 32.71 * | 2.73 | 0.91 | 4.69 | 5.79 | 10.01 | 20.63 * | 26.73 * |
| TH-103 | 13.70 | 7.70 | 0.84 | 7.39 | 10.88 | 4.02 | 9.30 | 6.56 |
| TH-77 | 7.91 | −4.52 | −1.81 | 4.40 | 4.92 | −4.00 | 12.41 * | 12.86 |
| TH-137 | 9.09 | 4.96 | 1.17 | 3.15 | 8.78 | −2.23 | 11.58 | 5.91 |
| HHB-67-Imp | ||||||||
| MPMH 17 | 17.52 | −3.44 | 1.04 | 7.58 | 8.60 | 6.17 | 12.91 * | 20.87 * |
| RHB 177 | 12.67 | −13.64 | 0.56 | 4.37 | 10.71 | 0.70 | 11.89 | 24.81 * |
| Dhanashakti | 12.66 | −7.15 | −1.64 | 0.24 | 6.67 | 1.55 | 10.94 | 18.31 * |
| GHB-538 | 15.99 | −13.75 | 0.37 | 3.45 | 1.05 | 3.19 | 12.40 * | 27.65 * |
| Raj 171 | 11.18 | −7.65 | 1.08 | 2.55 | 1.76 | 3.13 | 7.81 | 16.69 * |
| Components | PC-I | PC-II | PC-III | PC-IV | PC-V | PC-VI | PC-VII |
|---|---|---|---|---|---|---|---|
| Eigenvalue | 3.80 | 1.72 | 1.15 | 0.89 | 0.65 | 0.47 | 0.32 |
| Variance (%) explained by each axis | 42.23 | 19.07 | 12.80 | 9.83 | 7.26 | 5.22 | 3.55 |
| Cumulative % of variance | 42.23 | 61.30 | 74.10 | 83.93 | 91.19 | 96.41 | 99.96 |
| Parameters (Trait) | Dim.1 | Dim.2 | Dim.3 | ||||
| Days to 50% flowering (DB) | 0.759 | 0.203 | 0.149 | ||||
| Plant height (PH) | 0.518 | 0.437 | −0.126 | ||||
| Panicle length (PL) | 0.453 | −0.078 | 0.811 | ||||
| Panicle diameter (PD) | 0.456 | −0.409 | 0.066 | ||||
| Panicle yield (PY) | 0.648 | 0.124 | 0.424 | ||||
| Grain yield (GY) | 0.94 | −0.047 | −0.228 | ||||
| Dry fodder yield (DFY) | 0.404 | 0.84 | −0.132 | ||||
| Panicle harvest index (PNHI) | 0.862 | −0.113 | −0.423 | ||||
| Grain harvest index (HI) | 0.577 | −0.758 | −0.151 | ||||
| Number of Genotypes | DB | GY | DFY | PH | PL | PD | PY | PNHI | HI | Member Genotypes | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cluster-I (C-I) | 17 | 49.58 | 1491.45 | 6801.21 | 180.08 | 21.88 | 27.20 | 2537.30 | 58.75 | 15.99 | TH-1, TH-2, TH-43, TH-48, TH-25, TH-79, TH-96, TH-108, TH-129, TH-74, TH-136, TH-139, TH-42, TH-103, TH-77, HHB-67-Imp and RHB 177 |
| Cluster-II (C-II) | 4 | 49.19 | 1504.79 | 6642.45 | 180.58 | 21.69 | 26.84 | 2550.86 | 58.95 | 16.36 | TH-11, TH-114, TH-137 and Raj 171 |
| Cluster-III (C-III) | 15 | 49.53 | 1550.31 | 6834.48 | 180.52 | 21.78 | 27.44 | 2555.31 | 60.7 | 16.57 | TH-7, TH-12, TH-13, TH-49, TH-28, TH-37, TH-67, TH-138, TH-91, TH-124, TH-147, TH-149, MPMH 17, Dhanashakti and GHB-538 |
| Overall Mean | 49.51 | 1517.46 | 6797.43 | 180.32 | 21.82 | 27.26 | 2546.31 | 59.60 | 16.28 |
| GEN | Y*DFY | Y*PNHI | Y/DB | Y*PH | Y*PL | Y*PD | Y*HI | Y*PY | SI |
|---|---|---|---|---|---|---|---|---|---|
| TH-49 | 1.50 | 2.07 | 1.79 | 1.97 | 2.06 | 1.82 | 1.99 | 1.84 | 15.02 |
| TH-67 | 1.79 | 1.75 | 1.70 | 1.95 | 1.89 | 1.72 | 1.36 | 1.69 | 13.85 |
| TH-74 | 2.33 | 1.41 | 1.37 | 1.47 | 1.57 | 1.18 | 0.64 | 1.65 | 11.62 |
| TH-114 | 0.85 | 1.08 | 1.79 | 1.44 | 1.48 | 1.11 | 1.58 | 2.10 | 11.42 |
| TH-2 | 1.30 | 1.41 | 0.99 | 1.52 | 1.50 | 1.16 | 1.09 | 1.17 | 10.14 |
| TH-79 | 0.97 | 1.35 | 1.31 | 1.20 | 0.98 | 1.39 | 1.27 | 1.15 | 9.60 |
| TH-138 | 1.07 | 1.24 | 1.54 | 1.33 | 0.58 | 1.27 | 0.95 | 0.92 | 8.90 |
| TH-7 | 0.53 | 1.04 | 0.73 | 0.61 | 0.75 | 1.33 | 1.04 | 0.71 | 6.74 |
| TH-91 | 0.87 | 0.63 | 0.56 | 0.68 | 1.11 | 1.06 | 0.56 | 1.08 | 6.55 |
| TH-124 | 0.23 | 0.93 | 1.18 | 0.73 | 0.54 | 0.46 | 1.05 | 0.47 | 5.60 |
| TH-136 | −0.15 | 0.66 | 0.36 | 0.65 | 0.73 | 0.37 | 1.08 | 0.52 | 4.22 |
| TH-37 | −0.08 | 0.52 | 0.07 | 0.54 | 0.10 | 0.81 | 0.68 | 0.17 | 2.80 |
| TH-1 | 0.64 | 0.34 | 0.02 | 0.40 | 0.35 | 0.35 | 0.09 | 0.41 | 2.61 |
| TH-28 | 0.48 | 0.17 | 0.66 | −0.04 | 0.37 | −0.17 | −0.02 | 0.34 | 1.80 |
| TH-43 | 0.99 | 0.33 | 0.06 | 0.10 | −0.01 | −0.22 | −0.37 | 0.00 | 0.87 |
| MPMH 17 | −0.42 | −0.23 | −0.13 | 0.12 | 0.50 | 0.22 | 0.29 | 0.44 | 0.78 |
| TH-11 | −0.11 | 0.30 | 0.43 | −0.08 | −0.08 | −0.17 | 0.25 | −0.26 | 0.28 |
| TH-25 | 0.15 | 0.02 | −0.17 | −0.04 | −0.46 | −0.09 | −0.22 | −0.23 | −1.04 |
| TH-48 | 0.00 | −0.51 | −0.07 | −0.32 | −0.07 | −0.48 | −0.50 | 0.10 | −1.84 |
| TH-149 | 0.28 | −0.16 | −0.50 | −0.22 | −0.40 | −0.50 | −0.64 | −0.51 | −2.64 |
| TH-12 | −0.76 | −0.46 | −0.17 | −0.57 | −0.55 | 0.25 | −0.17 | −0.46 | −2.88 |
| GHB-538 | −1.40 | −0.49 | −0.01 | −0.36 | −0.38 | −0.91 | 0.46 | −0.23 | −3.32 |
| TH-108 | −1.14 | −0.52 | −0.51 | −0.59 | −0.26 | −0.17 | 0.03 | −0.56 | −3.70 |
| TH-13 | −0.59 | −0.45 | −0.57 | −0.51 | −0.42 | −0.17 | −0.39 | −0.65 | −3.75 |
| TH-103 | −0.01 | −0.82 | −0.90 | −0.54 | −0.18 | −0.14 | −1.03 | −0.36 | −3.98 |
| TH-129 | 0.08 | −0.80 | −0.72 | −0.38 | −0.65 | −0.59 | −1.20 | −0.63 | −4.90 |
| RHB 177 | −1.56 | −0.72 | −0.79 | −0.68 | −0.62 | −0.26 | 0.05 | −0.79 | −5.37 |
| TH-139 | −1.02 | −0.72 | −0.40 | −0.91 | −0.79 | −0.92 | −0.42 | −0.80 | −5.98 |
| Dhanashakti | −1.11 | −0.78 | −0.47 | −0.94 | −1.08 | −0.66 | −0.36 | −0.70 | −6.09 |
| Raj 171 | −1.23 | −1.05 | −1.24 | −0.78 | −0.97 | −1.27 | −0.55 | −0.68 | −7.77 |
| TH-42 | −1.13 | −1.21 | −1.09 | −1.12 | −1.01 | −0.53 | −0.98 | −1.01 | −8.10 |
| TH-137 | −0.50 | −0.95 | −1.08 | −0.98 | −1.12 | −0.79 | −1.34 | −1.39 | −8.16 |
| TH-96 | −0.88 | −1.10 | −1.28 | −1.24 | −0.73 | −1.16 | −0.99 | −0.91 | −8.27 |
| TH-147 | 0.66 | −1.23 | −1.24 | −0.92 | −1.26 | −1.62 | −2.09 | −0.95 | −8.66 |
| TH-77 | −1.21 | −0.97 | −1.22 | −1.44 | −1.11 | −1.26 | −1.00 | −1.66 | −9.86 |
| HHB-67-Imp | −1.40 | −2.10 | −2.00 | −2.05 | −2.35 | −2.42 | −2.20 | −1.98 | −16.50 |
| Mean | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| St Dev | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Hybrid | Standard Heterosis | Stability Parameter | Cluster Analysis | GYT Biplot | Promising for Number of Methods |
|---|---|---|---|---|---|
| TH-1 | √ | 1 | |||
| TH-2 | √ | √ | √ | 3 * | |
| TH-49 | √ | √ | √ | 3 | |
| TH-67 | √ | √ | √ | 3 | |
| TH-7 | √ | √ | 2 | ||
| TH-74 | √ | 1 | |||
| TH-79 | √ | √ | √ | 3 | |
| TH-91 | √ | 1 | |||
| TH-114 | √ | √ | √ | 3 | |
| TH-124 | √ | 1 | |||
| TH-138 | √ | √ | √ | √ | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kandarkar, K.; Palaniappan, V.; Gupta, P.C.; Rajasekaran, R.; Prabhakaran, J.; Sevugapperumal, N.; Gupta, S.K. Identification of Promising Three-Way Hybrids of Pearl Millet for Drought-Prone Environments of North-Western India. Agronomy 2023, 13, 2813. https://doi.org/10.3390/agronomy13112813
Kandarkar K, Palaniappan V, Gupta PC, Rajasekaran R, Prabhakaran J, Sevugapperumal N, Gupta SK. Identification of Promising Three-Way Hybrids of Pearl Millet for Drought-Prone Environments of North-Western India. Agronomy. 2023; 13(11):2813. https://doi.org/10.3390/agronomy13112813
Chicago/Turabian StyleKandarkar, Kuldeep, Viswanathan Palaniappan, Phool Chand Gupta, Ravikesavan Rajasekaran, Jeyakumar Prabhakaran, Nakkeeran Sevugapperumal, and Shashi Kumar Gupta. 2023. "Identification of Promising Three-Way Hybrids of Pearl Millet for Drought-Prone Environments of North-Western India" Agronomy 13, no. 11: 2813. https://doi.org/10.3390/agronomy13112813
APA StyleKandarkar, K., Palaniappan, V., Gupta, P. C., Rajasekaran, R., Prabhakaran, J., Sevugapperumal, N., & Gupta, S. K. (2023). Identification of Promising Three-Way Hybrids of Pearl Millet for Drought-Prone Environments of North-Western India. Agronomy, 13(11), 2813. https://doi.org/10.3390/agronomy13112813