Warm-Season Pasture Species Respond to Subsurface Placement of Phosphorus Fertiliser


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Plant Growth Conditions
2.3. Harvest and Analysis
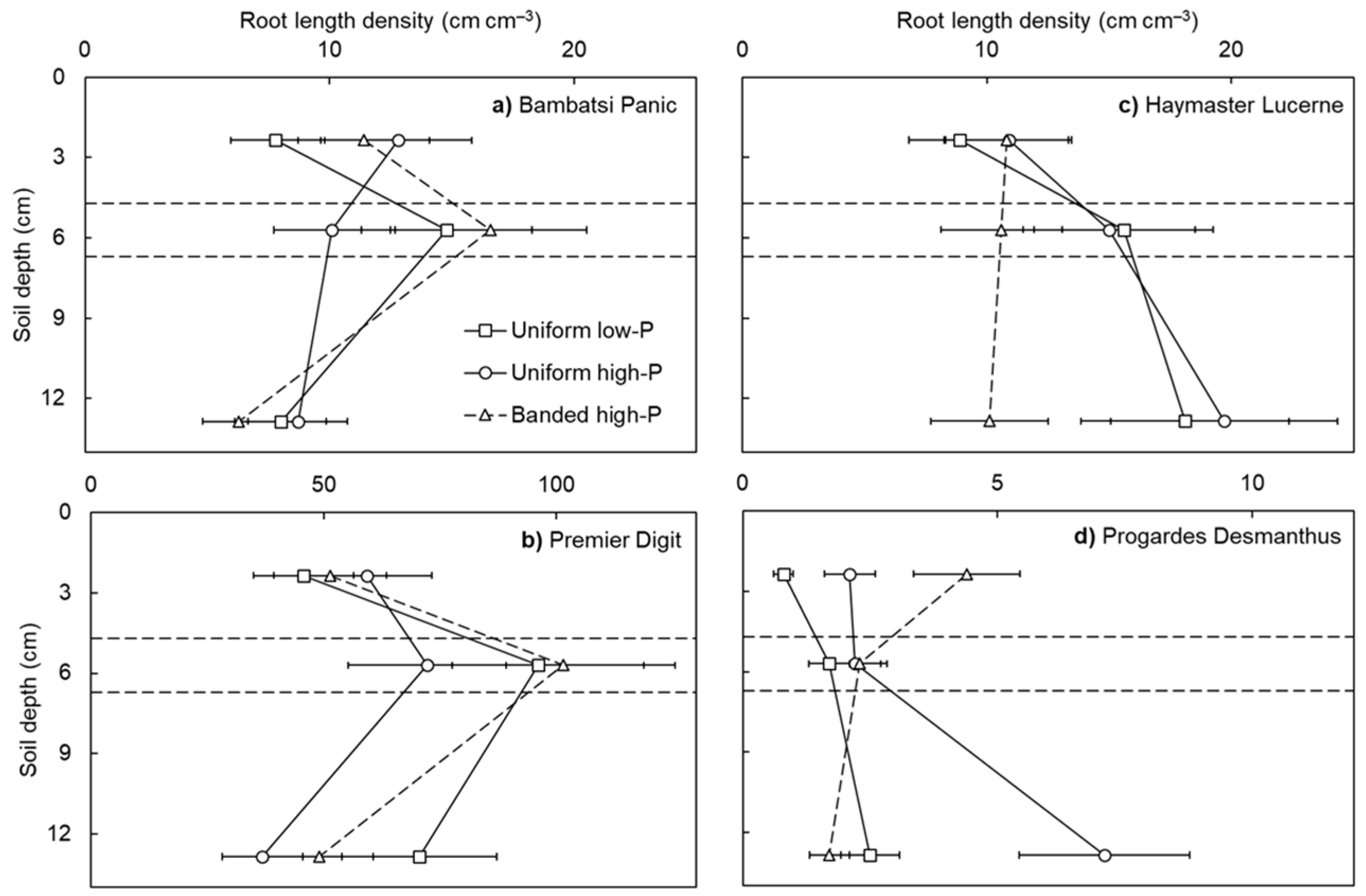
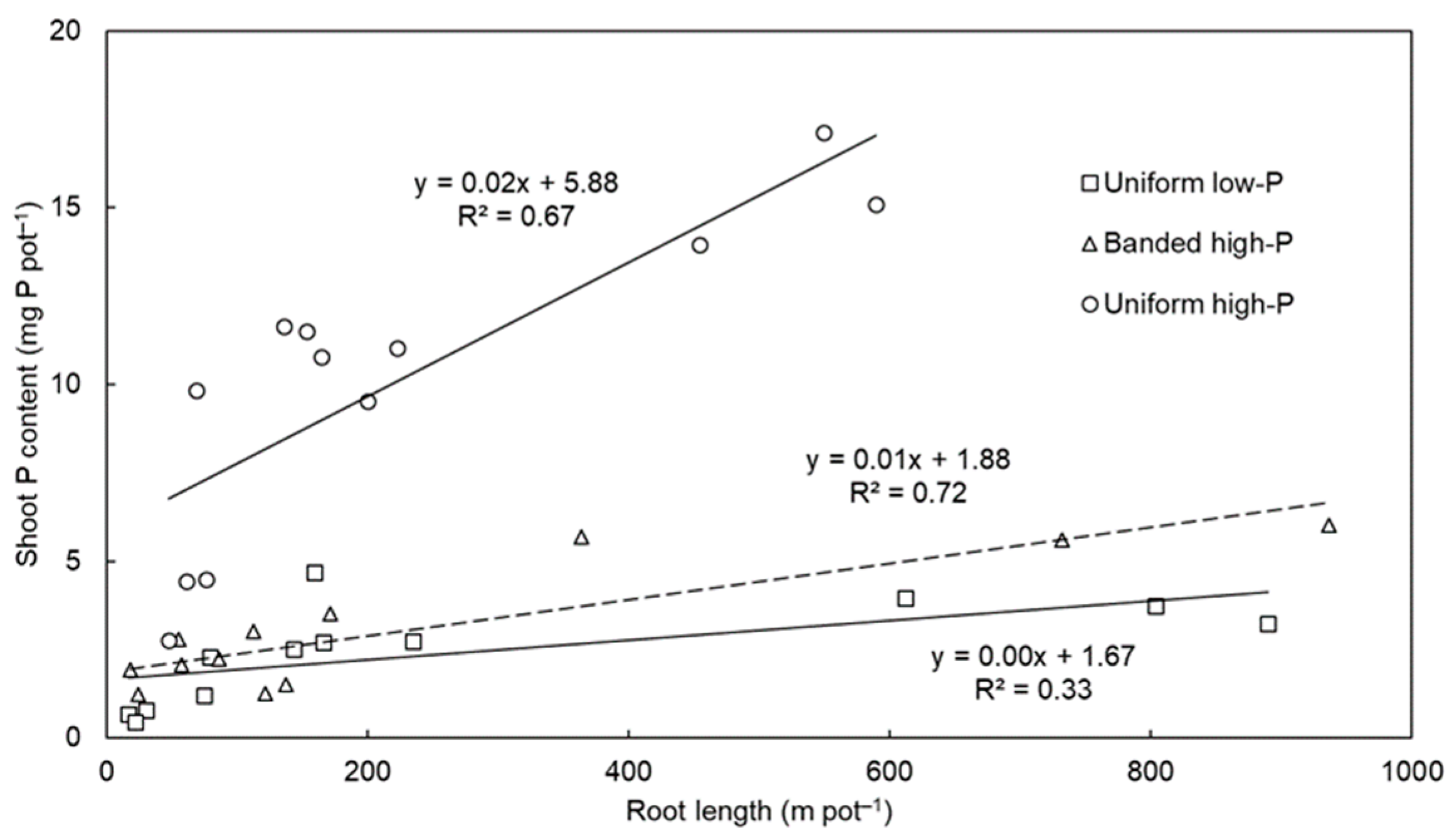
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, R.M.; Rees, M.C. Evaluation of tropical legumes on clay soils at four sites in southern inland Queensland. Trop. Grassl. 1997, 31, 95–106. [Google Scholar]
- Clem, R.L.; Hall, T.J. Persistence and productivity of tropical pasture legumes on three cracking clay soils (Vertisols) in north-eastern Queensland. Aust. J. Exp. Agric. 1994, 34, 161–171. [Google Scholar] [CrossRef]
- Haling, R.E.; Campbell, C.D.; Tighe, M.K.; Guppy, C.N. Effect of competition from a C4 grass on the phosphorus response of a subtropical legume. Crop Pasture Sci. 2013, 64, 985–992. [Google Scholar] [CrossRef]
- Robertson, F.A.; Myers, R.J.K.; Saffigna, P.G. Nitrogen cycling in brigalow clay soils under pasture and cropping. Aust. J. Soil Res. 1997, 35, 1323–1340. [Google Scholar] [CrossRef]
- Peck, G.; Hall, T.; Silcock, R.; Clem, B.; Buck, S.; Kedzlie, G. Persistence of pasture legumes in southern and central Queensland. In Proceedings of the 16th Australian Society of Agronomy Conference, Armidale, Australia, 14–18 October 2012. [Google Scholar]
- McCaskill, M.R.; Mitchell, M.L.; Zollinger, R.; Armstrong, R.D.; Partington, D. Dry matter and nutritive value responses of native, naturalised and sown pasture species to soil Olsen P. Crop Pasture Sci. 2019, 70, 1097–1109. [Google Scholar] [CrossRef]
- Muir, J.P.; Pitman, W.D.; Dubeux, J.C., Jr.; Foster, J.L. The future of warm-season, tropical and subtropical forage legumes in sustainable pastures and rangelands. Afr. J. Range Forage Sci. 2014, 31, 187–198. [Google Scholar] [CrossRef]
- Evans, P.S. Comparative root morphology of some pasture grasses and clovers. N. Z. J. Agric. Res. 1977, 20, 331–335. [Google Scholar] [CrossRef]
- Hill, J.O.; Simpson, R.J.; Moore, A.D.; Chapman, D.F. Morphology and response of roots of pasture species to phosphorus and nitrogen nutrition. Plant Soil 2006, 286, 7–19. [Google Scholar] [CrossRef]
- Lynch, J.P. Roots of the second green revolution. Aust. J. Bot. 2007, 55, 493–512. [Google Scholar] [CrossRef]
- Haling, R.E.; Yang, Z.; Shadwell, N.; Culvenor, R.A.; Stefanski, A.; Ryan, M.H.; Sandral, G.A.; Kidd, D.R.; Lambers, H.; Simpson, R.J. Root morphological traits that determine phosphorus-acquisition efficiency and critical external phosphorus requirement in pasture species. Funct. Plant Biol. 2016, 43, 815–826. [Google Scholar] [CrossRef]
- Yang, Z.; Culvenor, R.A.; Haling, R.E.; Stefanski, A.; Ryan, M.H.; Sandral, G.A.; Kidd, D.R.; Lambers, H.; Simpson, R.J. Variation in root traits associated with nutrient foraging among temperate pasture legumes and grasses. Grass Forage Sci. 2017, 72, 93–103. [Google Scholar] [CrossRef]
- Simpson, R.J.; Richardson, A.E.; Nichols, S.N.; Crush, J.R. Pasture plants and soil fertility management to improve the efficiency of phosphorus fertiliser use in temperate grassland systems. Crop Pasture Sci. 2014, 65, 556–575. [Google Scholar] [CrossRef]
- Hodge, A. Plastic plants and patchy soils. J. Exp. Bot. 2006, 57, 401–411. [Google Scholar] [CrossRef]
- Hodge, A. The plastic plant: Root responses to heterogeneous supplies of nutrients. New Phytol. 2004, 162, 9–24. [Google Scholar] [CrossRef]
- Drew, M.C. Comparison of the effects of a localised supply of phosphate, nitrate, ammonium and potassium on the growth of the seminal root system, and the shoot, in barley. New Phytol. 1975, 75, 479–490. [Google Scholar] [CrossRef]
- Lynch, J.P. Root phenes for enhanced soil exploration and phosphorus acquisition: Tools for future crops. Plant Physiol. 2011, 156, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P.; Wojciechowski, T. Opportunities and challenges in the subsoil: Pathways to deeper rooted crops. J. Exp. Bot. 2015, 66, 2199–2210. [Google Scholar] [CrossRef]
- Ho, M.D.; Rosas, J.C.; Brown, K.M.; Lynch, J.P. Root architectural tradeoffs for water and phosphorus acquisition. Funct. Plant Biol. 2005, 32, 737–748. [Google Scholar] [CrossRef]
- McLachlan, J.W.; Guppy, C.N.; Flavel, R.J. Differences in phosphorus acquisition and critical phosphorus requirements among nine Desmanthus spp. genotypes. Crop Pasture Sci. 2021, 72, 742–753. [Google Scholar] [CrossRef]
- McIvor, J.G.; Guppy, C.; Probert, M.E. Phosphorus requirements of tropical grazing systems: The northern Australian experience. Plant Soil 2011, 349, 55–67. [Google Scholar] [CrossRef]
- McLachlan, J.W.; Guppy, C.N.; Flavel, R.J. Banded application improves the recovery of phosphorus fertiliser in a temperate pasture sward containing red clover. In Proceedings of the 19th Australian Society of Agronomy Conference, Wagga Wagga, Australia, 25–29 August 2019. [Google Scholar]
- Isbell, R.F. The Australian Soil Classification; CSIRO Publishing: Melbourne, Australia, 1996. [Google Scholar]
- Rayment, G.E.; Lyons, D.J. Soil Chemical Methods—Australasia; CSIRO Publishing: Melbourne, Australia, 2011. [Google Scholar]
- Hopkinson, J.M.; English, B.H. Germination and hardseededness in desmanthus. Trop. Grassl. 2004, 38, 1–16. [Google Scholar]
- Bouma, T.J.; Nielsen, K.L.; Koutstaal, B. Sample preparation and scanning protocol for computerised analysis of root length and diameter. Plant Soil 2000, 218, 185–196. [Google Scholar] [CrossRef]
- IAEA. Use of Isotope and Radiation Methods in Soil and Water Management and Crop Nutrition; Training Course Series No. 14; IAEA: Vienna, Austria, 2001. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Team, R.C. Nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-148. 2020. Available online: https://CRAN.R-project.org/package=nlme (accessed on 12 September 2023).
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 1.5.0. 2020. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 12 September 2023).
- Simpson, R.J.; Stefanski, A.; Marshall, D.J.; Moore, A.D.; Richardson, A.E. Management of soil phosphorus fertility determines the phosphorus budget of a temperate grazing system and is the key to improving phosphorus efficiency. Agric. Ecosyst. Environ. 2015, 212, 263–277. [Google Scholar] [CrossRef]
- McLaughlin, M.J.; McBeath, T.M.; Smernik, R.; Stacey, S.P.; Ajiboye, B.; Guppy, C. The chemical nature of P accumulation in agricultural soils—Implications for fertiliser management and design: An Australian perspective. Plant Soil 2011, 349, 69–87. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P Treatment and Species | Shoot Dry Mass | Shoot P Concentration | Shoot P Content | Shoot P Content/RL |
|---|---|---|---|---|
| (g DM pot−1) | (mg P g DM−1) | (mg P pot−1) | (µg m−1) | |
| Uniform low-P | ||||
| Bambatsi Panic | 1.28 ± 0.29 | 2.04 ± 0.35 | 2.73 ± 0.52 | 23.9 ± 4.82 |
| Premier Digit | 4.39 ± 0.29 | 0.83 ± 0.14 | 3.64 ± 0.52 | 4.80 ± 0.96 |
| Haymaster Lucerne | 1.94 ± 0.29 | 1.38 ± 0.24 | 2.66 ± 0.52 | 15.0 ± 3.02 |
| Progardes® Desmanthus | 0.35 ± 0.29 | 1.76 ± 0.30 | 0.64 ± 0.52 | 27.6 ± 5.58 |
| Uniform high-P | ||||
| Bambatsi Panic | 2.34 ± 0.29 | 4.74 ± 0.82 | 10.99 ± 0.52 | 97.3 ± 19.7 |
| Premier Digit | 4.07 ± 0.29 | 3.77 ± 0.65 | 15.39 ± 0.52 | 29.0 ± 5.86 |
| Haymaster Lucerne | 2.95 ± 0.29 | 3.54 ± 0.61 | 10.44 ± 0.52 | 53.7 ± 10.8 |
| Progardes® Desmanthus | 1.26 ± 0.29 | 3.07 ± 0.53 | 3.89 ± 0.52 | 62.5 ± 12.6 |
| Banded high-P | ||||
| Bambatsi Panic | 1.81 ± 0.29 | 0.91 ± 0.16 | 1.62 ± 0.52 | 16.0 ± 3.23 |
| Premier Digit | 3.99 ± 0.29 | 1.49 ± 0.26 | 5.77 ± 0.52 | 9.20 ± 1.85 |
| Haymaster Lucerne | 1.78 ± 0.29 | 1.64 ± 0.28 | 2.93 ± 0.52 | 24.3 ± 4.91 |
| Progardes® Desmanthus | 0.54 ± 0.29 | 5.00 ± 0.86 | 1.98 ± 0.52 | 64.8 ± 13.1 |
| ANOVA | ||||
| Species | p < 0.001 | p = 0.003 | p < 0.001 | p < 0.001 |
| P treatment | p = 0.004 | p < 0.001 | p < 0.001 | p < 0.001 |
| Species × P treatment | p = 0.176 | p < 0.001 | p < 0.001 | p = 0.004 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McLachlan, J.W.; Staker, B.J.; Flavel, R.J.; Guppy, C.N. Warm-Season Pasture Species Respond to Subsurface Placement of Phosphorus Fertiliser. Agronomy 2023, 13, 2524. https://doi.org/10.3390/agronomy13102524
McLachlan JW, Staker BJ, Flavel RJ, Guppy CN. Warm-Season Pasture Species Respond to Subsurface Placement of Phosphorus Fertiliser. Agronomy. 2023; 13(10):2524. https://doi.org/10.3390/agronomy13102524
Chicago/Turabian StyleMcLachlan, Jonathan W., Benjamin J. Staker, Richard J. Flavel, and Chris N. Guppy. 2023. "Warm-Season Pasture Species Respond to Subsurface Placement of Phosphorus Fertiliser" Agronomy 13, no. 10: 2524. https://doi.org/10.3390/agronomy13102524
APA StyleMcLachlan, J. W., Staker, B. J., Flavel, R. J., & Guppy, C. N. (2023). Warm-Season Pasture Species Respond to Subsurface Placement of Phosphorus Fertiliser. Agronomy, 13(10), 2524. https://doi.org/10.3390/agronomy13102524