


Strategy for Selection of Drought-Tolerant Arabica Coffee Genotypes in Brazil
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Experiment 1: Evaluation of Coffee Progenies in Field Conditions in a Region with a Long and Defined Dry Season
2.2.1. Productivity Analysis
2.2.2. Statistical Analysis
2.3. Experiment 2: Evaluation of Coffee Progenies under Water Deficit in Greenhouse
2.3.1. Phenotypic Analyses
2.3.2. Physiological Analyses
2.3.3. Anatomical Analyses
2.3.4. Statistical Analysis
Genotypic Correlations among Traits
3. Results
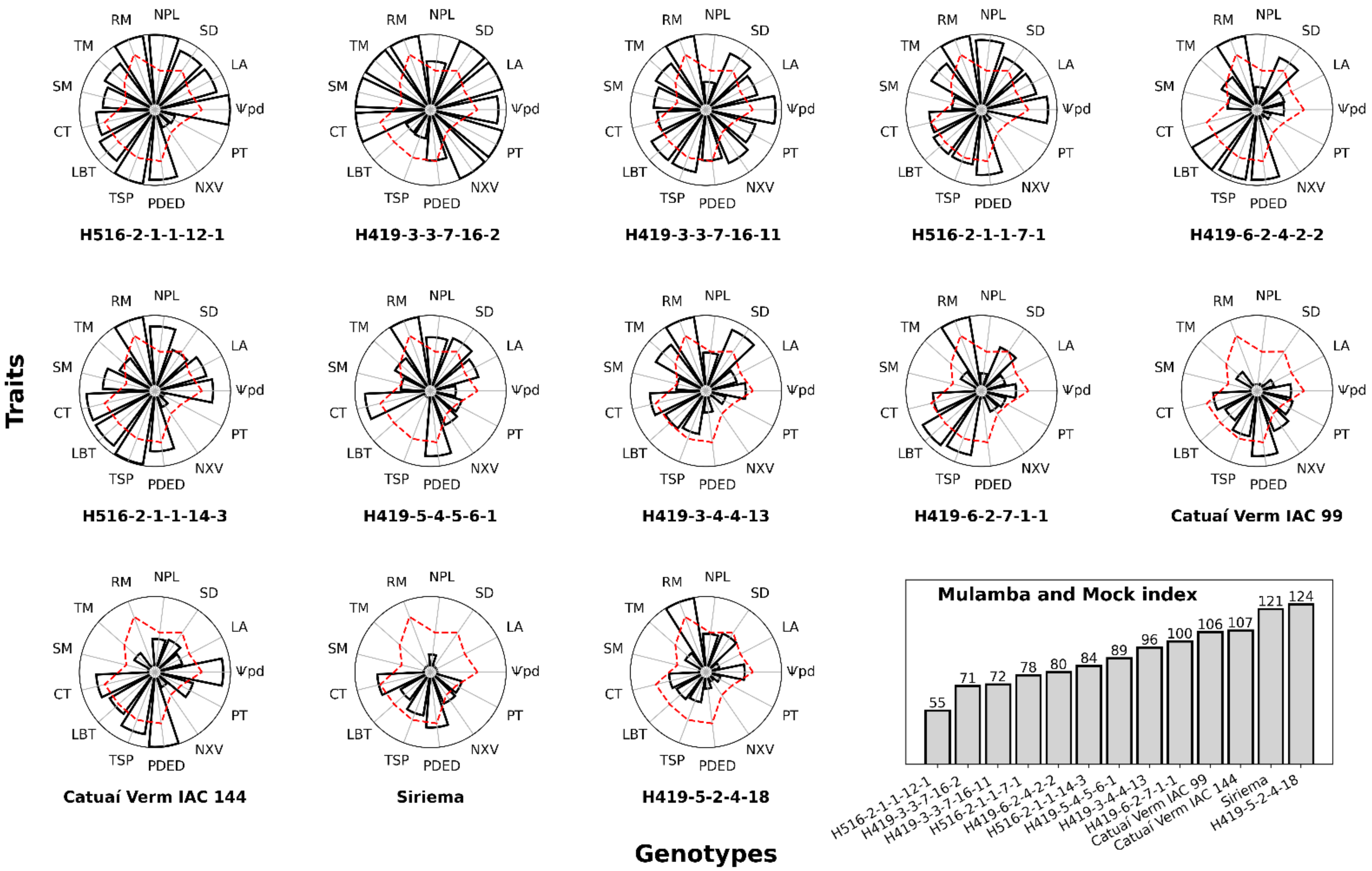
3.1. Selection under Greenhouse Conditions
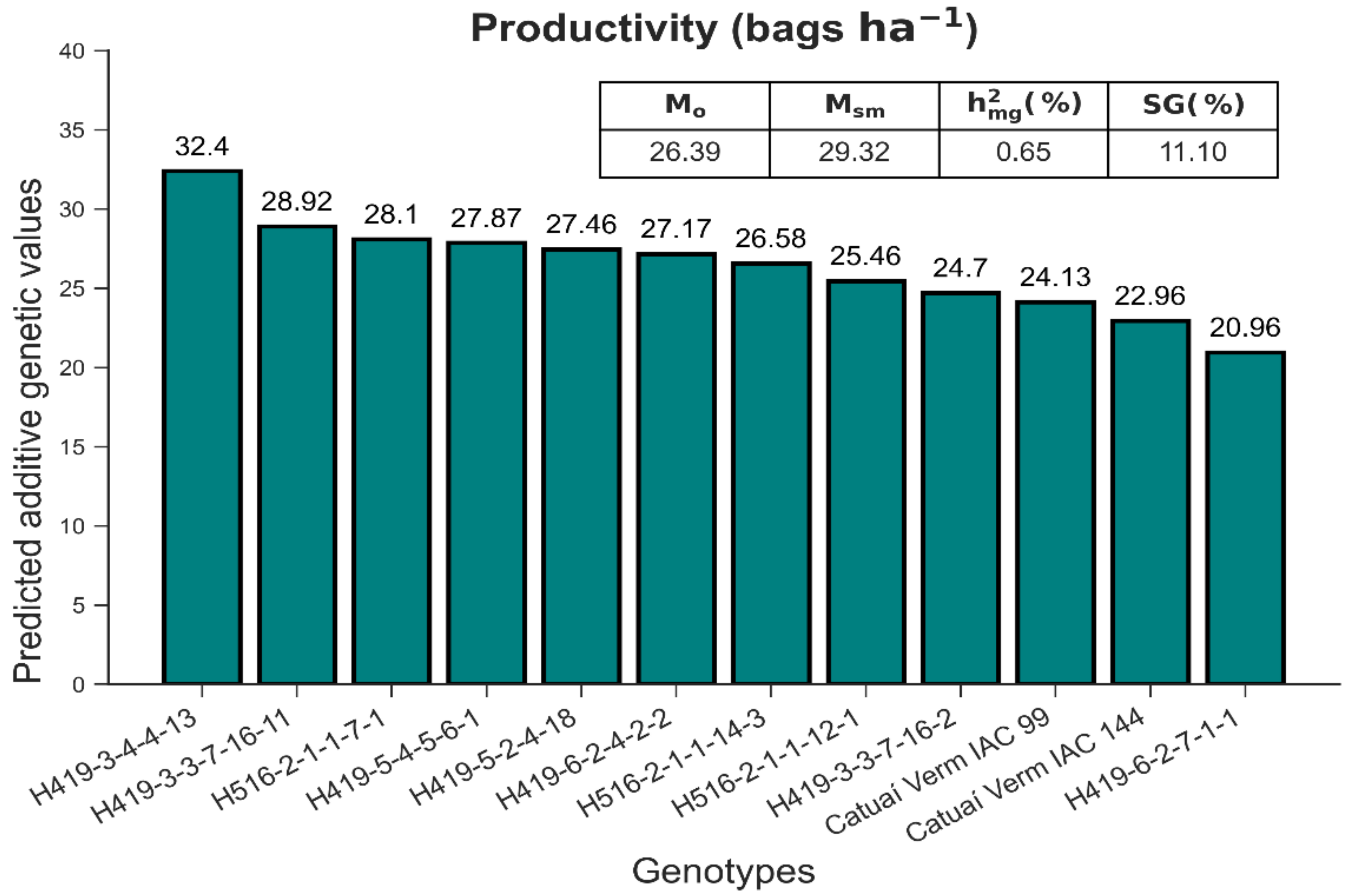
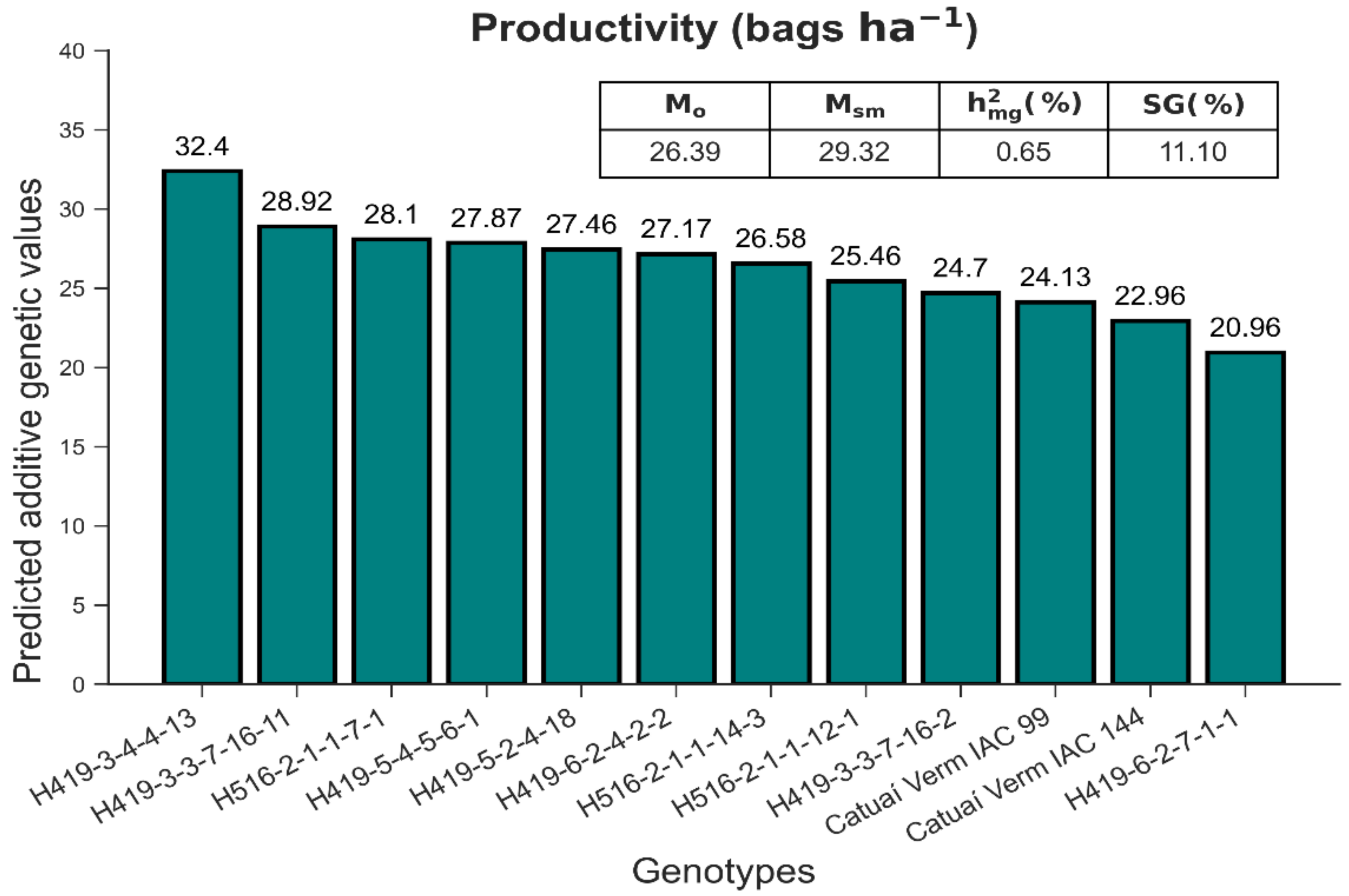
3.2. Selection under Field Conditions
3.3. Genotypic Correlations among Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nakabayashi, R.; Saito, K. Integrated Metabolomics for Abiotic Stress Responses in Plants. Curr. Opin. Plant Biol. 2015, 24, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Barchet, G.L.H.; Dauwe, R.; Guy, R.D.; Schroeder, W.R.; Soolanayakanahally, R.Y.; Campbell, M.M.; Mansfield, S.D. Investigating the Drought-Stress Response of Hybrid Poplar Genotypes by Metabolite Profiling. Tree Physiol. 2014, 34, 1203–1219. [Google Scholar] [CrossRef] [PubMed]
- Krasensky, J.; Jonak, C. Drought, Salt, and Temperature Stress-Induced Metabolic Rearrangements and Regulatory Networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Brinkley, S.; Smith, E.; Sela, A.; Theisen, M.; Thibodeau, C.; Warne, T.; Anderson, E.; Van Dusen, N.; Giuliano, P.; et al. Climate Change and Coffee Quality: Systematic Review on the Effects of Environmental and Management Variation on Secondary Metabolites and Sensory Attributes of Coffea arabica and Coffea canephora. Front. Plant Sci. 2021, 12, 708013. [Google Scholar] [CrossRef]
- Silva, V.A.; Prado, F.M.; Antunes, W.C.; Paiva, R.M.C.; Ferrão, M.A.G.; Andrade, A.C.; Di Mascio, P.; Loureiro, M.E.; DaMatta, F.M.; Almeida, A.M. Reciprocal Grafting between Clones with Contrasting Drought Tolerance Suggests a Key Role of Abscisic Acid in Coffee Acclimation to Drought Stress. Plant Growth Regul. 2018, 85, 221–229. [Google Scholar] [CrossRef]
- Lima, E.A.; Furlanetto, C.; Nicole, M.; Gomes, A.C.M.M.; Almeida, M.R.A.; Jorge-Júnior, A.; Correa, V.R.; Salgado, S.M.; Ferrão, M.A.G.; Carneiro, R.M.D.G. The Multi-Resistant Reaction of Drought-Tolerant Coffee ‘Conilon Clone 14’ to Meloidogyne Spp. and Late Hypersensitive-Like Response in Coffea canephora. Phytopathology 2015, 105, 805–814. [Google Scholar] [CrossRef]
- Mofatto, L.S.; de Carneiro, F.A.; Vieira, N.G.; Duarte, K.E.; Vidal, R.O.; Alekcevetch, J.C.; Cotta, M.G.; Verdeil, J.-L.; Lapeyre-Montes, F.; Lartaud, M.; et al. Identification of Candidate Genes for Drought Tolerance in Coffee by High-Throughput Sequencing in the Shoot Apex of Different Coffea arabica Cultivars. BMC Plant Biol. 2016, 16, 94. [Google Scholar] [CrossRef]
- Alves, G.S.C.; Torres, L.F.; de Aquino, S.O.; Reichel, T.; Freire, L.P.; Vieira, N.G.; Vinecky, F.; This, D.; Pot, D.; Etienne, H.; et al. Nucleotide Diversity of the Coding and Promoter Regions of DREB1D, a Candidate Gene for Drought Tolerance in Coffea Species. Trop. Plant Biol. 2018, 11, 31–48. [Google Scholar] [CrossRef]
- Torres, L.F.; Reichel, T.; Déchamp, E.; de Aquino, S.O.; Duarte, K.E.; Alves, G.S.C.; Silva, A.T.; Cotta, M.G.; Costa, T.S.; Diniz, L.E.C.; et al. Expression of DREB-Like Genes in Coffea canephora and C. arabica Subjected to Various Types of Abiotic Stress. Trop. Plant Biol. 2019, 12, 98–116. [Google Scholar] [CrossRef]
- Kiwuka, C.; Goudsmit, E.; Tournebize, R.; de Aquino, S.O.; Douma, J.C.; Bellanger, L.; Crouzillat, D.; Stoffelen, P.; Sumirat, U.; Legnaté, H.; et al. Genetic Diversity of Native and Cultivated Ugandan Robusta Coffee (Coffea canephora Pierre Ex A. Froehner): Climate Influences, Breeding Potential and Diversity Conservation. PLoS ONE 2021, 16, e0245965. [Google Scholar] [CrossRef]
- De Oliveira Santos, M.; Coelho, L.S.; Carvalho, G.R.; Botelho, C.E.; Torres, L.F.; Vilela, D.J.M.; Andrade, A.C.; Silva, V.A. Photochemical Efficiency Correlated with Candidate Gene Expression Promote Coffee Drought Tolerance. Sci. Rep. 2021, 11, 7436. [Google Scholar] [CrossRef]
- Bertrand, B.; Marraccini, P.; Villain, L.; Breitler, J.-C.; Etienne, H. Healthy Tropical Plants to Mitigate the Impact of Climate Change—As Exemplified in Coffee. In Climate Change and Agriculture Worldwide; Torquebiau, E., Ed.; Springer: Dordrecht, The Netherlands, 2016; pp. 83–95. ISBN 9789401774604. [Google Scholar]
- De Souza, B.P.; Martinez, H.E.P.; de Carvalho, F.P.; Loureiro, M.E.; Sturião, W.P. Gas Exchanges and Chlorophyll Fluorescence of Young Coffee Plants Submitted to Water and Nitrogen Stresses. J. Plant Nutr. 2020, 43, 2455–2465. [Google Scholar] [CrossRef]
- Dias, P.C.; Araujo, W.L.; Moraes, G.A.B.K.; Barros, R.S.; DaMatta, F.M. Morphological and Physiological Responses of Two Coffee Progenies to Soil Water Availability. J. Plant Physiol. 2007, 164, 1639–1647. [Google Scholar] [CrossRef] [PubMed]
- Fazuoli, L.C.; Braghini, M.T.; Silvarolla, M.B.; Gonçalves, W.; Mistro, J.C.; Gallo, P.B.; Guerreiro Filho, O. IAC Catuaí SH3—A Dwarf Arabica Coffee Cultivar with Leaf Rust Resistance and Drought Tolerance. Crop Breed. Appl. Biotechnol. 2019, 19, 356–359. [Google Scholar] [CrossRef]
- Sera, T.; Sera, G.H.; Fazuoli, L.C. IPR 103—Rustic Dwarf Arabic Coffee Cultivar More Adapted to Hot Regions and Poor Soils. Crop Breed. Appl. Biotechnol. 2013, 13, 95–98. [Google Scholar] [CrossRef]
- Ferrão, R.G.; Fonseca, A.F.A.; Silveira, J.S.M.; Ferrão, M.A.G.; Bragança, S.M. EMCAPA 8141—Robustão Capixaba, Variedade Clonal De Café Conilon Tolerante À Seca Desenvolvida Para O Estado do Espírito Santo. Rev. Ceres 2000, 273, 555–560. [Google Scholar]
- Van der Vossen, H.; Bertrand, B.; Charrier, A. Next Generation Variety Development for Sustainable Production of Arabica Coffee (Coffea arabica L.): A Review. Euphytica 2015, 204, 243–256. [Google Scholar] [CrossRef]
- Surya Prakash, N.; Devasia, J.; Raghuramulu, Y.; Aggarwal, R.K. Genetic Diversity and Coffee Improvement in India. In Molecular Breeding for Sustainable Crop Improvement; Rajpal, V.R., Rao, S.R., Raina, S.N., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 11, pp. 231–268. ISBN 9783319270883. [Google Scholar]
- Silva, P.E.M.; Cavatte, P.C.; Morais, L.E.; Medina, E.F.; DaMatta, F.M. The Functional Divergence of Biomass Partitioning, Carbon Gain and Water Use in Coffea canephora in Response to the Water Supply: Implications for Breeding Aimed at Improving Drought Tolerance. Environ. Exp. Bot. 2013, 87, 49–57. [Google Scholar] [CrossRef]
- De Resende, M.D.V. Software Selegen-REML/BLUP: A Useful Tool for Plant Breeding. Crop Breed. Appl. Biotechnol. 2016, 16, 330–339. [Google Scholar] [CrossRef]
- Barros, R.S.; Maestri, M.; Vieira, M.; Braga-Filho, L.J. Determinação De Área De Folhas Do Café (Coffea arabica L. Cv. ‘Bourbon Amarelo’). Rev. Ceres 1973, 20, 44–52. [Google Scholar]
- Scholander, P.F.; Hammel, H.T.; Hemmingsen, E.A.; Bradstreet, E.D. Hydrostatic pressure and osmotic potential in leaves of mangroves and some other plants. Proc. Natl. Acad. Sci. USA 1964, 52, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Mulamba, N.N.; Mock, J.J. Improvement of Yield Potential of the ETO Blanco Maize (Zea mays L.) Population by Breeding for Plant Traits [Mexico]. Egypt. J. Genet. Cytol. 1978, 7, 40–45. [Google Scholar]
- Cruz, C.D. GENES—A Software Package for Analysis in Experimental Statistics and Quantitative Genetics. Acta Sci. Agron. 2013, 35, 271–276. [Google Scholar] [CrossRef]
- Yan, W.; Zhong, Y.; Shangguan, Z. Responses of Different Physiological Parameter Thresholds to Soil Water Availability in Four Plant Species during Prolonged Drought. Agric. For. Meteorol. 2017, 247, 311–319. [Google Scholar] [CrossRef]
- Menezes-Silva, P.E.; Sanglard, L.M.V.P.; Ávila, R.T.; Morais, L.E.; Martins, S.C.V.; Nobres, P.; Patreze, C.M.; Ferreira, M.A.; Araújo, W.L.; Fernie, A.R.; et al. Photosynthetic and Metabolic Acclimation to Repeated Drought Events Play Key Roles in Drought Tolerance in Coffee. J. Exp. Bot. 2017, 68, 4309–4322. [Google Scholar] [CrossRef] [PubMed]
- Ogunbayo, S.A.; Sie, M.; Ojo, D.K.; Sanni, K.A.; Akinwale, M.G.; Toulou, B.; Shittu, A.; Idehen, E.O.; Popoola, A.R.; Daniel, I.O.; et al. Genetic Variation and Heritability of Yield and Related Traits in Promising Rice Genotypes (Oryza sativa L.). J. Plant Breed. Crop Sci. 2014, 6, 153–159. [Google Scholar] [CrossRef]
- Dubberstein, D.; Oliveira, M.G.; Aoyama, E.M.; Guilhen, J.H.; Ferreira, A.; Marques, I.; Ramalho, J.C.; Partelli, F.L. Diversity of Leaf Stomatal Traits among Coffea canephora Pierre Ex A. Froehner Genotypes. Agronomy 2021, 11, 1126. [Google Scholar] [CrossRef]
- Anim-Kwapong, E.; Anim-Kwapong, G.J.; Adomako, B. Indirect Versus Direct Selection of Adaptable Genotypes of Coffea Canephora for Drought Stress and Non-Stress Conditions. J. Crop Improv. 2011, 25, 356–370. [Google Scholar] [CrossRef]
- King’oro, M. Effect of Different Watering Regimes on Agro-Morphology of Selected Coffee Genotypes. AJEA 2014, 4, 1016–1026. [Google Scholar] [CrossRef]
- De Resende, M.D.V.; Duarte, J.B. Precision and Quality Control in Variety Trials. Pesq. Agropec. Trop. 2007, 37, 182–194. [Google Scholar]
- Rocha, R.B.; Vieira, A.H.; Gama, M.M.B.; Rossi, L.M.B. Avaliação Genética De Procedências De Bandarra (Schizolobium amazonicum) Utilizando REML/BLUP (Máxima Verossimilhança Restrita/Melhor Predição Linear Não Viciada). Sci. For. 2009, 37, 351–358. [Google Scholar]
- De Carvalho, A.M.; Mendes, A.N.G.; Carvalho, G.R.; Botelho, C.E.; Gonçalves, F.M.A.; Ferreira, A.D. Correlação Entre Crescimento e Produtividade de Cultivares de Café Em Diferentes Regiões de Minas Gerais, Brasil. Pesq. Agropec. Bras. 2010, 45, 269–275. [Google Scholar] [CrossRef]
- Castro, E.D.; Pereira, F.J.; Paiva, R. Histologia Vegetal: Estrutura E Função De Órgãos Vegetativos, 9th ed.; Editora UFLA: Lavras, Brazil, 2009; 244p. [Google Scholar]
- Prince, S.J.; Murphy, M.; Mutava, R.N.; Durnell, L.A.; Valliyodan, B.; Grover Shannon, J.; Nguyen, H.T. Root Xylem Plasticity to Improve Water Use And Yield In Water-Stressed Soybean. J. Exp. Bot. 2017, 68, 2027–2036. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, P.; Wang, G.; Augstein, F.; de Vries, J.; Carlsbecker, A. Continuous Root Xylem Formation and Vascular Acclimation To Water Deficit Involves Endodermal ABA Signalling Via miR165. Development 2018, 145, dev.159202. [Google Scholar] [CrossRef]
- Hazman, M.; Brown, K.M. Progressive Drought Alters Architectural and Anatomical Traits of Rice Roots. Rice 2018, 11, 62. [Google Scholar] [CrossRef] [Green Version]
- Velikova, V.; Arena, C.; Izzo, L.G.; Tsonev, T.; Koleva, D.; Tattini, M.; Roeva, O.; De Maio, A.; Loreto, F. Functional and Structural Leaf Plasticity Determine Photosynthetic Performances during Drought Stress and Recovery in Two Platanus Orientalis Populations from Contrasting Habitats. Int. J. Mol. Sci. 2020, 21, 3912. [Google Scholar] [CrossRef]
- DaMatta, F.M.; Avila, R.T.; Cardoso, A.A.; Martins, S.C.V.; Ramalho, J.C. Physiological and Agronomic Performance of the Coffee Crop in the Context of Climate Change and Global Warming: A Review. J. Agric. Food Chem. 2018, 66, 5264–5274. [Google Scholar] [CrossRef]
- Thioune, E.-H.; Strickler, S.; Gallagher, T.; Charpagne, A.; Decombes, P.; Osborne, B.; McCarthy, J. Temperature Impacts the Response of Coffea canephora to Decreasing Soil Water Availability. Trop. Plant Biol. 2020, 13, 236–250. [Google Scholar] [CrossRef]
- Tounekti, T.; Mahdhi, M.; Al-Turki, T.A.; Khemira, H. Water Relations and Photo-Protection Mechanisms during Drought Stress in Four Coffee (Coffea arabica) Cultivars from Southwestern Saudi Arabia. S. Afr. J. Bot. 2018, 117, 17–25. [Google Scholar] [CrossRef]
- Fernandes, I.; Marques, I.; Paulo, O.S.; Batista, D.; Partelli, F.L.; Lidon, F.C.; DaMatta, F.M.; Ramalho, J.C.; Ribeiro-Barros, A.I. Understanding the Impact of Drought in Coffea Genotypes: Transcriptomic Analysis Supports a Common High Resilience to Moderate Water Deficit but a Genotype Dependent Sensitivity to Severe Water Deficit. Agronomy 2021, 11, 2255. [Google Scholar] [CrossRef]
- De Guedes, F.A.F.; Nobres, P.; Rodrigues Ferreira, D.C.; Menezes-Silva, P.E.; Ribeiro-Alves, M.; Correa, R.L.; DaMatta, F.M.; Alves-Ferreira, M. Transcriptional Memory Contributes to Drought Tolerance in Coffee (Coffea canephora) Plants. Environ. Exp. Bot. 2018, 147, 220–233. [Google Scholar] [CrossRef]
- Marques, I.; Gouveia, D.; Gaillard, J.-C.; Martins, S.; Semedo, M.C.; Lidon, F.C.; DaMatta, F.M.; Ribeiro-Barros, A.I.; Armengaud, J.; Ramalho, J.C. Next-Generation Proteomics Reveals a Greater Antioxidative Response to Drought in Coffea arabica Than in Coffea Canephora. Agronomy 2022, 12, 148. [Google Scholar] [CrossRef]
- Sales, C.R.G.; Molero, G.; Evans, J.R.; Taylor, S.H.; Joynson, R.; Furbank, R.T.; Hall, A.; Carmo-Silva, E. Phenotypic Variation in Photosynthetic Traits in Wheat Grown under Field versus Glasshouse Conditions. J. Exp. Bot. 2022, 73, 3221–3237. [Google Scholar] [CrossRef] [PubMed]
- DaMatta, F.M.; Ramalho, J.D.C. Impacts of drought and temperature stress on coffee physiology and production: A review. Braz. J. Plant Physiol. 2006, 18, 55–81. [Google Scholar] [CrossRef]
- De Oliveira, A.C.B.; Pereira, A.A.; da Silva, F.L.; de Rezende, J.C.; Botelho, C.E.; Carvalho, G.R. Prediction of Genetic Gains from Selection in Arabica Coffee Progenies. Crop Breed. Appl. Biotechnol. 2011, 11, 106–113. [Google Scholar] [CrossRef] [Green Version]
- Reis, A.M.; Mendes, A.N.G.; Abrahão, J.C.R.; de Santos, M.O.; Silva, V.A. Early Selection of Drought-Tolerant Coffea Arabica Genotypes At The Seedling Stage Using Functional Divergence. Pesq. Agropec. Trop. 2022, 52, e72412. [Google Scholar]

{kind=link}
{kind=link}
| Trait | SG (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| Physiological parameters 1 | Control (irrigated) | Ψpd | 0.002 ± 0.02 | 0.01 | 0.10 | −0.14 | - | - |
| A | 0.10 ± 0.12 | 0.31 | 0.55 | 7.61 | - | - | ||
| gs | 0.11 ± 0.12 | 0.31 | 0.56 | 0.12 | - | - | ||
| E | 0.08 ± 0.11 | 0.27 | 0.52 | 2.52 | - | - | ||
| Drought stress | Ψpd | 0.48 ± 0.23 * | 0.79 * | 0.88 | −2.33 | −3.01 | −28.8 | |
| A | 0.06 ± 0.10 | 0.22 | 0.47 | 2.92 | - | - | ||
| gs | 0.07 ± 0.10 | 0.22 | 0.46 | 0.03 | - | - | ||
| E | 0.11 ± 0.13 | 0.34 | 0.58 | 0.57 | - | - | ||
| Morphological parameters | LA | 0.49 ± 0.19 * | 0.85 * | 0.89 | 0.45 | 0.52 | 14.35 | |
| H | 0.00 ± 0.02 | 0.02 | 0.02 | 47.81 | - | - | ||
| SD | 0.53 ± 0.20 * | 0.90 * | 1 | 10.94 | 11.54 | 5.5 | ||
| NPB | 0.41 ± 0.17 * | 0.81 * | 0.87 | 5.65 | 5.96 | 5.4 | ||
| ÂPL | 0.23 ± 0.13 | 0.65 | 0.77 | 71.28 | - | - | ||
| LPL | 0.14 ± 0.10 | 0.46 | 0.47 | 13.55 | - | - | ||
| RL | 0.09 ± 0.08 | 0.43 | 0.73 | 48.99 | - | - | ||
| RM | 0.31 ± 0.16 * | 0.73 * | 0.79 | 0.04 | 0.04 | 8.07 | ||
| TM | 0.35 ± 0.17 * | 0.76 * | 0.82 | 0.13 | 0.14 | 9.19 | ||
| SM | 0.35 ± 0.16 * | 0.77 * | 0.83 | 0.09 | 0.1 | 9.56 | ||
| RMSM | 0.21 ± 0.13 | 0.67 | 0.95 | 0.43 | - | - | ||
| Anatomical parameters | AdE | 0.29 ± 0.15 | 0.77 | 0.99 | 26.95 | - | - | |
| CT | 0.32 ± 0.16 | 0.79 * | 0.99 | 5.01 | 4.8 | −4.25 | ||
| AbE | 0.11 ± 0.09 | 0.5 | 0.98 | 19.77 | - | - | ||
| LBT | 0.35 ± 0.16 | 0.81 * | 0.99 | 322 | 324.76 | 0.86 | ||
| TPP | 0.03 ± 0.05 | 0.2 | 0.93 | 74.24 | - | - | ||
| TSP | 0.41 ± 0.18 | 0.85 * | 1 | 200.38 | 202.91 | 1.26 | ||
| PDED | 0.38 ± 0.17 | 0.83 * | 1 | 1.47 | 1.49 | 1.18 | ||
| DEN | 0.21 ± 0.13 | 0.69 | 0.99 | 174.86 | - | - | ||
| NXV | 0.51 ± 0.20 | 0.89 * | 1 | 165.69 | 177.51 | 7.13 | ||
| PT | 0.60 ± 0.21 | 0.92 * | 1 | 78.64 | 81.87 | 4.11 | ||
| XVT | 0.17 ± 0.11 | 0.62 | 0.99 | 18.84 | - | - | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, V.A.; Abrahão, J.C.d.R.; Reis, A.M.; Santos, M.d.O.; Pereira, A.A.; Botelho, C.E.; Carvalho, G.R.; Castro, E.M.d.; Barbosa, J.P.R.A.D.; Botega, G.P.; et al. Strategy for Selection of Drought-Tolerant Arabica Coffee Genotypes in Brazil. Agronomy 2022, 12, 2167. https://doi.org/10.3390/agronomy12092167
Silva VA, Abrahão JCdR, Reis AM, Santos MdO, Pereira AA, Botelho CE, Carvalho GR, Castro EMd, Barbosa JPRAD, Botega GP, et al. Strategy for Selection of Drought-Tolerant Arabica Coffee Genotypes in Brazil. Agronomy. 2022; 12(9):2167. https://doi.org/10.3390/agronomy12092167
Chicago/Turabian StyleSilva, Vânia Aparecida, Juliana Costa de Rezende Abrahão, André Moraes Reis, Meline de Oliveira Santos, Antônio Alves Pereira, Cesar Elias Botelho, Gladyston Rodrigues Carvalho, Evaristo Mauro de Castro, João Paulo Rodrigues Alves Delfino Barbosa, Gustavo Pucci Botega, and et al. 2022. "Strategy for Selection of Drought-Tolerant Arabica Coffee Genotypes in Brazil" Agronomy 12, no. 9: 2167. https://doi.org/10.3390/agronomy12092167
APA StyleSilva, V. A., Abrahão, J. C. d. R., Reis, A. M., Santos, M. d. O., Pereira, A. A., Botelho, C. E., Carvalho, G. R., Castro, E. M. d., Barbosa, J. P. R. A. D., Botega, G. P., & Oliveira, A. C. B. d. (2022). Strategy for Selection of Drought-Tolerant Arabica Coffee Genotypes in Brazil. Agronomy, 12(9), 2167. https://doi.org/10.3390/agronomy12092167