
QTL Analysis and Heterosis Loci of Effective Tiller Using Three Genetic Populations Derived from Indica-Japonica Crosses in Rice

 , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Population Construction
2.2. Trait Statistics
2.3. Bin Mapping-Based QTL Analysis
2.4. Heterosis Analysis
2.5. Candidate Gene Prediction
3. Results
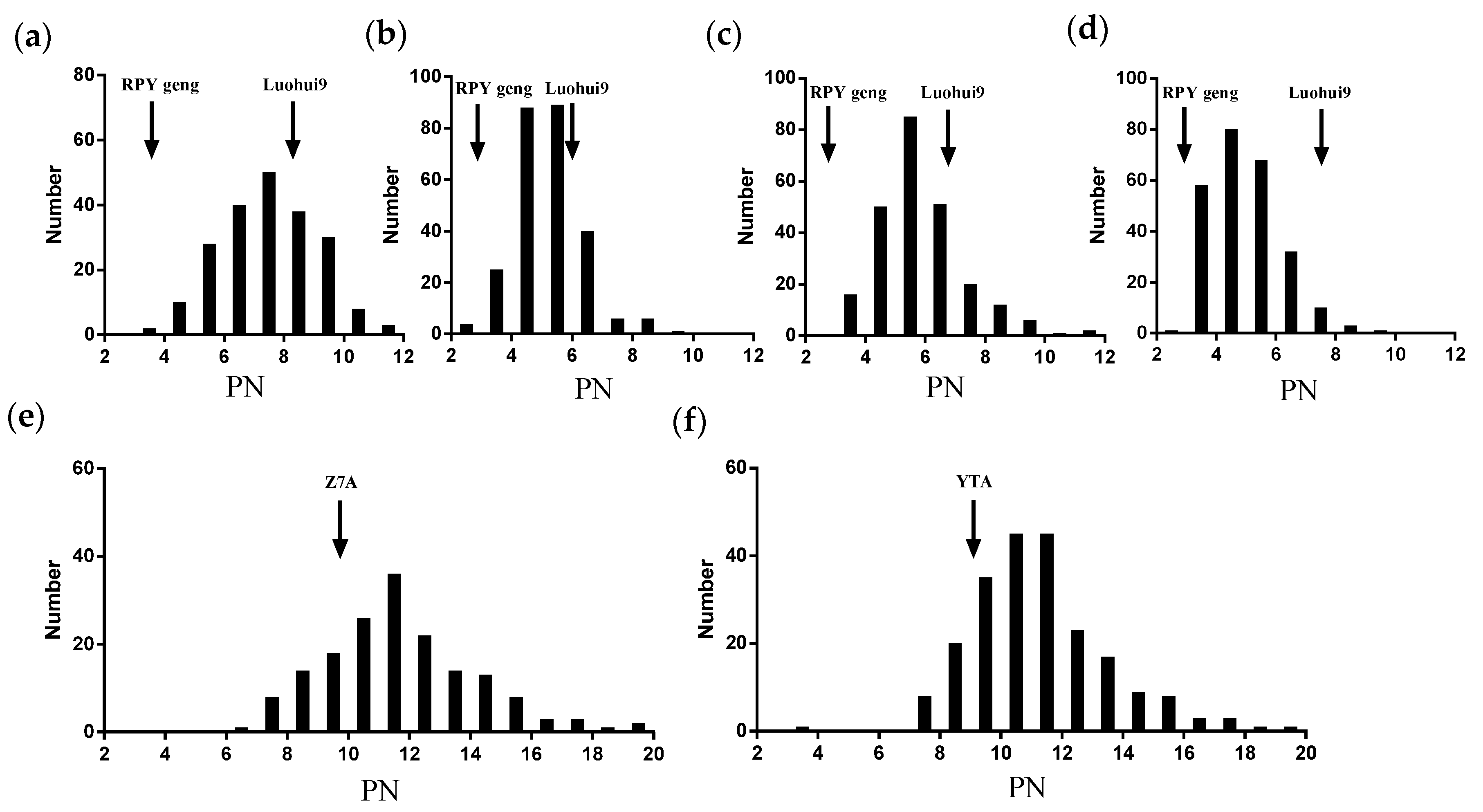
3.1. The Trait Performance of Effective Panicle Numbers per Plant (PN) from the RILs and the Testcross Hybrid Populations
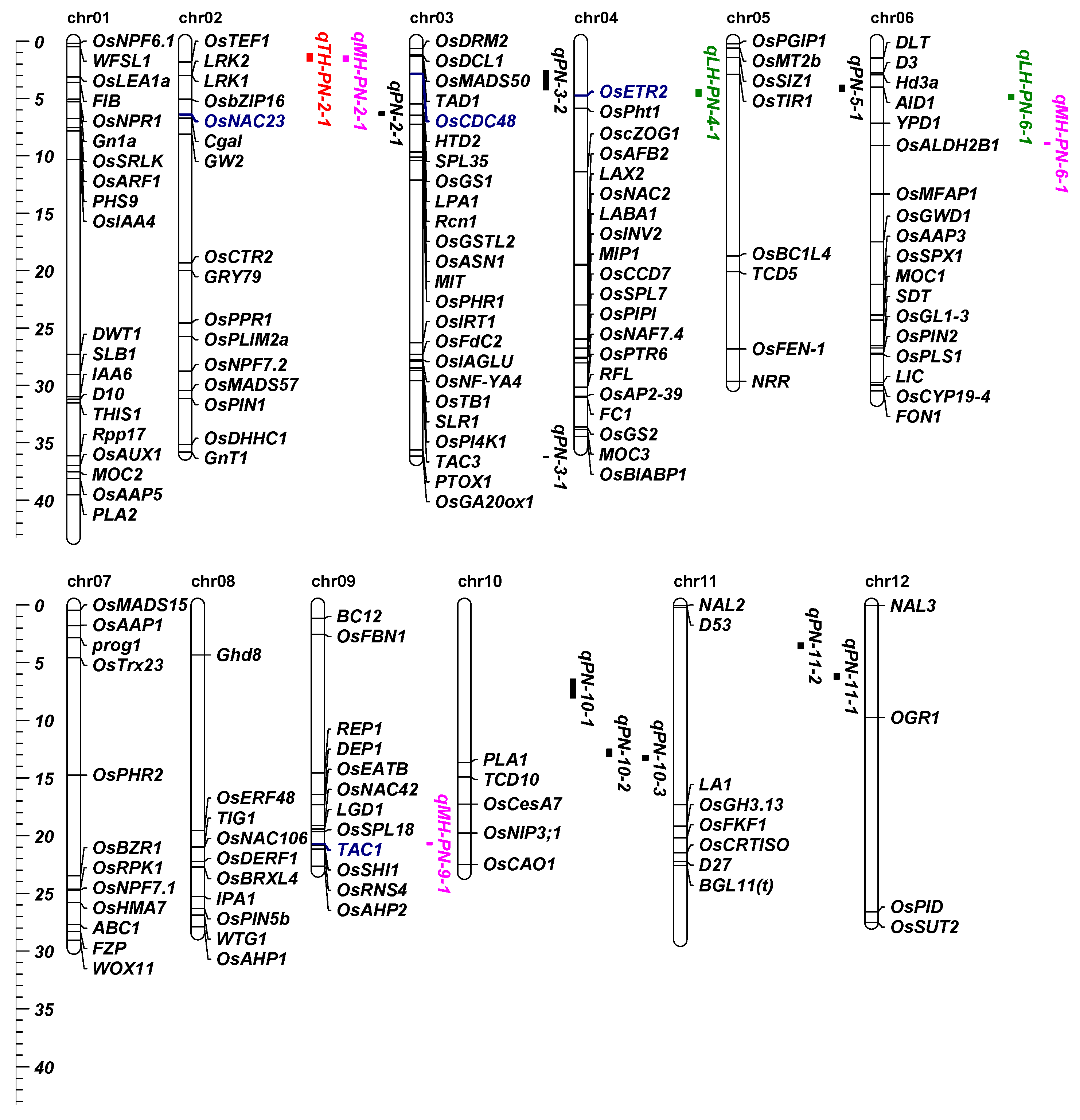
3.2. QTLs Detection in RILs and Heterosis-Related QTLs Detection in Two Testcross Hybrid Populations
3.3. Known Genes Affecting the PN of RILs in QTLs Interval
3.4. Kown Genes Acted as Heterosis-Related QTLs for PN
3.5. Prediction of Candidate Gene in the QTL Cluster
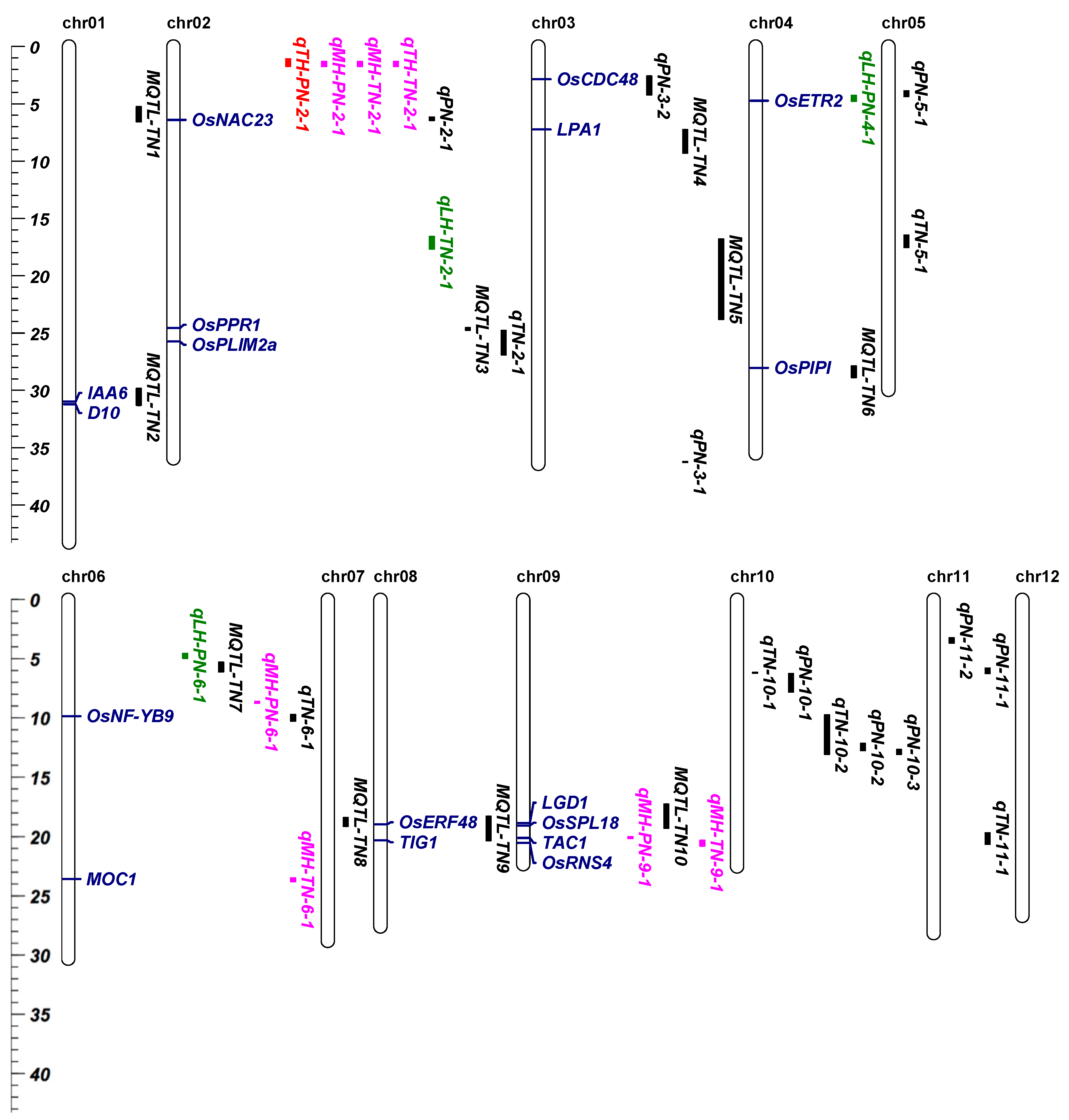
3.6. MQTLs and QTLs of Effective Tiller and Their Heterosis-Related Loci Cover Multiple Known Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jiao, Y.Q.; Wang, Y.H.; Xue, D.W.; Wang, J.; Yan, M.X.; Liu, G.F.; Dong, G.J.; Zeng, D.L.; Lu, Z.F.; Zhu, X.D.; et al. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice. Nat. Genet. 2010, 42, 541–544. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.Q.; Li, F.M.; Zhang, Q.; Wang, X.Q.; Guo, H.F.; Xie, J.Y.; Zhu, X.Y.; Ullah Khan, N.; Zhang, Z.Y.; Li, J.J.; et al. Genetic architecture to cause dynamic change in tiller and panicle numbers revealed by genome-wide association study and transcriptome profile in rice. Plant J. 2020, 104, 1603–1616. [Google Scholar] [CrossRef] [PubMed]
- Arcade, A.; Labourdette, A.; Falque, M.; Mangin, B.; Chardon, F.; Charcosset, A.; Joets, J. BioMercator: Integrating genetic maps and QTL towards discovery of candidate genes. Bioinformatics 2004, 20, 2324–2326. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.C.; Shabala, S.; Koutoulis, A.; Shabala, L.; Zhou, M.X. Meta-analysis of major QTL for abiotic stress tolerance in barley and implications for barley breeding. Planta 2017, 245, 283–295. [Google Scholar] [CrossRef]
- Martinez, A.K.; Soriano, J.M.; Tuberosa, R.; Koumproglou, R.; Jahrmann, T.; Salvi, S.; Yield, Q.T. Lome distribution correlates with gene density in maize. Plant Sci. 2016, 242, 300–309. [Google Scholar] [CrossRef]
- Kong, W.L.; Deng, X.X.; Liao, Z.Y.; Wang, Y.B.; Zhou, M.A.; Wang, Z.H.; Li, Y.S. De novo assembly of two chromosome-level rice genomes and bin-based QTL mapping reveal genetic diversity of grain weight trait in rice. Front. Plant Sci. 2022, 13, 995634. [Google Scholar] [CrossRef]
- Kong, W.L.; Zhang, C.H.; Qiang, Y.L.; Zhong, H.; Zhao, G.Q.; Li, Y.S. Integrated RNA-seq analysis and Meta-QTLs mapping provide insight into cold stress response in rice seeding roots. Int. J. Mol. Sci. 2020, 21, 4615. [Google Scholar] [CrossRef]
- Kong, W.L.; Zhong, H.; Gong, Z.Y.; Fang, X.Y.; Sun, T.; Deng, X.X.; Li, Y.S. Meta-analysis of salt stress transcriptome responses in different rice genotypes at the seedling stage. Plants 2019, 8, 64. [Google Scholar] [CrossRef]
- Khahani, B.; Tavakol, E.; Shariati, V.; Fornara, F. Genome wide screening and comparative genome analysis for Meta-QTLs, ortho-MQTLs and candidate genes controlling yield and yield-related traits in rice. BMC Genom. 2020, 21, 294–318. [Google Scholar] [CrossRef]
- Zhang, C.Y.; Liu, J.; Zhao, T.; Gomez, A.; Li, C.; Yu, C.S.; Li, H.Y.; Lin, J.Z.; Yang, Y.Z.; Liu, B.; et al. A drought-inducible transcription factor delays reproductive timing in rice. Plant Physiol. 2016, 171, 334–343. [Google Scholar] [CrossRef] [Green Version]
- Oikawa, T.; Kyozuka, J. Two-step regulation of LAX PANICLE1 protein accumulation in axillary meristem formation in rice. Plant Cell. 2009, 21, 1095–1108. [Google Scholar] [CrossRef] [PubMed]
- Shao, G.N.; Lu, Z.F.; Xiong, J.S.; Wang, B.; Jing, Y.H.; Meng, X.B.; Liu, G.F.; Ma, H.Y.; Liang, Y.; Chen, F.; et al. Tiller bud formation regulators MOC1 and MOC3 cooperatively promote tiller bud outgrowth by activating FON1 expression in rice. Mol. Plant 2019, 12, 1090–1102. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, H.; Zhang, Y.; Hattori, S.; Omae, M.; Shimizu-Sato, S.; Oikawa, T.; Qian, Q.; Nishimura, M.; Kitano, H.; Xie, H.; et al. LAX PANICLE2 of rice encodes a novel nuclear protein and regulates the formation of axillary meristems. Plant Cell. 2011, 23, 3276–3287. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Qian, Q.; Fu, Z.M.; Wang, Y.H.; Xiong, G.S.; Zeng, D.; Wang, X.Q.; Liu, X.F.; Teng, S.; Hiroshi, F.; et al. Control of tillering in rice. Nature 2003, 422, 618–621. [Google Scholar] [CrossRef]
- Koumoto, T.; Shimada, H.; Kusano, H.; She, K.C.; Iwamoto, M.; Takano, M. Rice monoculm mutation moc2, which inhibits outgrowth of the second tillers, is ascribed to lack of a fructose-1,6-bisphosphatase. Plant Biotechnol. 2013, 30, 47–56. [Google Scholar] [CrossRef]
- Li, Z.Y.; Wei, X.J.; Tong, X.H.; Zhao, J.; Liu, X.X.; Wang, H.M.; Tang, L.Q.; Shu, Y.Z.; Li, G.H.; Wang, Y.F.; et al. The OsNAC23-Tre6P-SnRK1a feed-forward loop regulates sugar homeostasis and grain yield in rice. Mol. Plant 2022, 15, 706–722. [Google Scholar] [CrossRef]
- Lei, L.; Zheng, H.L.; Wang, J.G.; Liu, H.L.; Sun, J.; Zhao, H.W.; Yang, L.M.; Zou, D. Genetic dissection of rice (Oryza sativa L.) tiller, plant height, and grain yield based on QTL mapping and metaanalysis. Euphytica 2018, 214, 109. [Google Scholar] [CrossRef]
- Arite, T.; Iwata, H.; Ohshima, K.; Maekawa, M.; Nakajima, M.; Kojima, M.; Sakakibara, H.; Kyozuka, J. DWARF10, an RMS1/MAX4/DAD1 ortholog, controls lateral bud outgrowth in rice. Plant J. 2007, 51, 1019–1029. [Google Scholar] [CrossRef]
- Zha, M.; Imran, M.; Wang, Y.; Xu, J.; Ding, Y.; Wang, S.H. Transcriptome analysis revealed the interaction among strigolactones, auxin, and cytokinin in controlling the shoot branching of rice. Plant Cell Rep. 2019, 38, 279–293. [Google Scholar] [CrossRef]
- Kong, W.L.; Deng, X.X.; Yang, J.; Zhang, C.H.; Sun, T.; Ji, W.J.; Zhong, H.; Fu, X.P.; Li, Y.S. High-resolution bin-based linkage mapping uncovers the genetic architecture and heterosis-related loci of plant height in indica-japonica derived populations. Plant J. 2022, 110, 814–827. [Google Scholar] [CrossRef]
- Deng, X.X.; Kong, W.L.; Sun, T.; Zhang, C.H.; Zhong, H.; Zhao, G.Q.; Liu, X.H.; Qiang, Y.L.; Li, Y.S. Bin mapping-based QTL analyses using three genetic populations derived from indica-japonica crosses uncover multiple grain shape heterosis-related loci in rice. Plant Genome 2022, 15, e20171. [Google Scholar] [PubMed]
- Arends, D.; Prins, P.; Jansen, R.C.; Broman, K.W. R/qtl: High-throughput multiple QTL mapping. Bioinformatics 2010, 26, 2990–2992. [Google Scholar] [CrossRef] [PubMed]
- Dupuis, J.; Siegmund, D. Statistical methods for mapping quantitative trait loci from a dense set of markers. Genetics 1999, 151, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Voorrips, R.E. Mapchart: Software for the graphical presenttion of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar]
- Patrick, S.S.; Nathan, M.S. Progress toward understanding heterosis in crop plants. Annu. Rev. Plant Biol. 2013, 64, 71–88. [Google Scholar]
- Kong, W.L.; Zhang, C.H.; Zhang, S.C.; Qiang, Y.L.; Zhang, Y.; Zhong, H.; Li, Y.S. Uncovering the novel QTLs and candidate genes of sale tolerance in rice with linkage mapping, RTM-GWAS, and RNA-seq. Rice 2021, 14, 93. [Google Scholar] [CrossRef]
- Wuriyanghan, H.; Zhang, B.; Cao, W.H.; Ma, B.; Lei, G.; Liu, Y.F.; Wei, W.; Wu, H.J.; Chen, L.J.; Chen, H.W.; et al. The ethylene receptor ETR2 delays floral transition and affects starch accumulation in rice. Plant Cell. 2009, 21, 1473–1494. [Google Scholar] [CrossRef]
- Yu, B.S.; Lin, Z.W.; Li, H.X.; Li, X.J.; Li, J.Y.; Wang, Y.H.; Zhang, X.; Zhu, Z.F.; Zhai, W.X.; Wang, X.K.; et al. TAC1, a major quantitative trait locus controlling tiller angle in rice. Plant J. 2007, 52, 891–898. [Google Scholar] [CrossRef]
- Na, J.K.; Huh, S.M.; Yoon, I.S.; Byun, M.O.; Lee, Y.H.; Lee, K.O.; Kim, D. Rice LIM protein OsPLIM2a is involved in rice seed and tiller development. Mol. Breed. 2014, 34, 569–581. [Google Scholar] [CrossRef]
- Das, S.; Parida, S.K.; Agarwal, P.; Tyagi, A.K. Transcription factor OsNF-YB9 regulates reproductive growth and development in rice. Planta 2019, 250, 1849–1865. [Google Scholar] [CrossRef]
- Mathew, I.E.; Das, S.; Mahto, A.; Agarwal, P. Three rice NAC transcription factors heteromerize and are associated with seed size. Front. Plant Sci. 2016, 7, 1638. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lu, Q.T.; Wen, X.G.; Lu, C.M. Enhanced Sucrose Loading Improves Rice Yield by Increasing Grain Size. Plant Physiol. 2015, 169, 2848–2862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, C.A.; Sagar, R.; Geng, Y.; Primavesi, L.F.; Patel, M.K.; Passarelli, M.K.; Gilmore, I.S.; Steven, R.T.; Bunch, J.; Paul, M.J.; et al. Chemical intervention in plant sugar signalling increases yield and resilience. Nature 2016, 540, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.; Lee, D.K.; Choi, Y.D.; Kim, J.K. OsIAA6, a member of the rice Aux/IAA gene family, is involved in drought tolerance and tiller outgrowth. Plant Sci. 2015, 236, 304–312. [Google Scholar] [CrossRef]
- Huang, X.H.; Yang, S.H.; Gong, J.Y.; Zhao, Q.; Feng, Q.; Zhan, Q.L.; Zhao, Y.; Li, W.J.; Cheng, B.Y.; Xia, J.H. Genomic architecture of heterosis for yield traits in rice. Nature 2016, 537, 629–633. [Google Scholar] [CrossRef]
- Li, D.Y.; Huang, Z.Y.; Song, S.H.; Xin, Y.Y.; Mao, D.H.; Lv, Q.M.; Zhou, M.; Tian, D.M.; Tang, M.F.; Wu, Q.; et al. Integrated analysis of phenome, genome, and transcriptome of hybrid rice uncovered multiple heterosis-related loci for yield increase. Proc. Natl. Acad. Sci. USA 2016, 113, E6026–E6035. [Google Scholar] [CrossRef]
- Lin, Z.C.; Qin, P.; Zhang, X.W.; Fu, C.J.; Deng, H.C.; Fu, X.X.; Hunag, Z.; Jiang, S.Q.; Tang, X.Y.; Wang, X.F.; et al. Divergent selection and genetic introgression shape the genome landscape of heterosis in hybrid rice. Proc. Natl. Acad. Sci. USA 2020, 117, 4623–4631. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Jia, H.Y.; Li, T.; Wu, J.Z.; Nagarajan, R.; Lei, L.; Powers, C.; Kan, C.C.; Hua, W.; Liu, Z.Y.; et al. TaCol-B5 modifies spike architecture and enhances grain yield in wheat. Science 2022, 376, 180–183. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Sun, X.M.; Ma, X.Q.; Xu, B.X.; Zhao, Y.; Ma, Z.Q.; Li, G.L.; Khan, N.U.; Pan, Y.H.; Liang, Y.T.; et al. GNP6, a novel allele of MOC1, regulates panicle and tiller development in rice. Crop J. 2020, 9, 57–67. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Position (bp) | Homologous Known Gene | QTLs | QTL Position (bp) | Male Parental Gene/Female Parental Gene | Difference of Protein Sequence |
|---|---|---|---|---|---|---|
| LOC_Os10g25720 | 13318548- 13322321 | D10 | qPN-10-2 | 13238715- 13251104 | FaEVM0003415.1/MoEVM0001835.1 | Yes |
| LOC_Os10g25830 | 13387338- 13388803 | MIT | qPN-10-2 | 13238715- 13251104 | FaEVM0043545.1/MoEVM0016401.1 | Yes |
| LOC_Os10g25890 | 13416622- 13421264 | D10 | qPN-10-2 | 13238715- 13251104 | FaEVM0032270.1/MoEVM0005566.1 | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, X.; Wang, J.; Liu, X.; Yang, J.; Zhou, M.; Kong, W.; Jiang, Y.; Ke, S.; Sun, T.; Li, Y. QTL Analysis and Heterosis Loci of Effective Tiller Using Three Genetic Populations Derived from Indica-Japonica Crosses in Rice. Agronomy 2022, 12, 2171. https://doi.org/10.3390/agronomy12092171
Deng X, Wang J, Liu X, Yang J, Zhou M, Kong W, Jiang Y, Ke S, Sun T, Li Y. QTL Analysis and Heterosis Loci of Effective Tiller Using Three Genetic Populations Derived from Indica-Japonica Crosses in Rice. Agronomy. 2022; 12(9):2171. https://doi.org/10.3390/agronomy12092171
Chicago/Turabian StyleDeng, Xiaoxiao, Jingzhang Wang, Xuhui Liu, Jian Yang, Mingao Zhou, Weilong Kong, Yifei Jiang, Shiming Ke, Tong Sun, and Yangsheng Li. 2022. "QTL Analysis and Heterosis Loci of Effective Tiller Using Three Genetic Populations Derived from Indica-Japonica Crosses in Rice" Agronomy 12, no. 9: 2171. https://doi.org/10.3390/agronomy12092171
APA StyleDeng, X., Wang, J., Liu, X., Yang, J., Zhou, M., Kong, W., Jiang, Y., Ke, S., Sun, T., & Li, Y. (2022). QTL Analysis and Heterosis Loci of Effective Tiller Using Three Genetic Populations Derived from Indica-Japonica Crosses in Rice. Agronomy, 12(9), 2171. https://doi.org/10.3390/agronomy12092171