
Seed-Soaking with Melatonin for the Improvement of Seed Germination, Seedling Growth, and the Antioxidant Defense System under Flooding Stress

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Material
2.2. Experimental Design
2.3. Determination Items and Methods
2.3.1. Determination of Seed Germination Indicators
2.3.2. Determination of Seed Shoots and Roots Growth Indicators
2.3.3. Determination of Seed Shoots and Roots Biomass Indicators
2.3.4. Determination of Physiological and Biochemical Indicators
2.3.5. Data Processing and Analysis
3. Results
3.1. Germination Characteristics
3.2. Biomass of Rice Seed Shoots and Roots
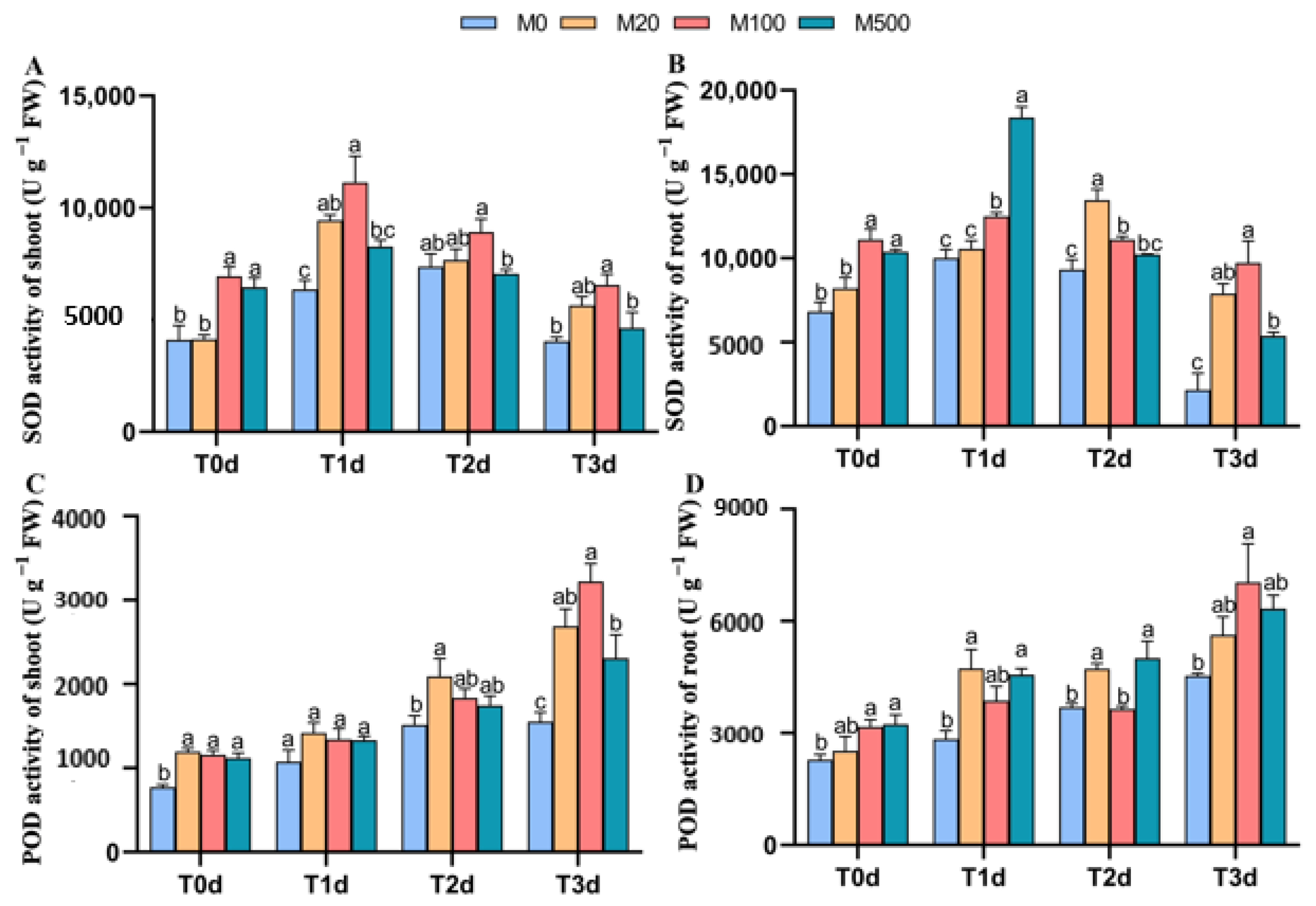
3.3. Antioxidant Enzyme Activities
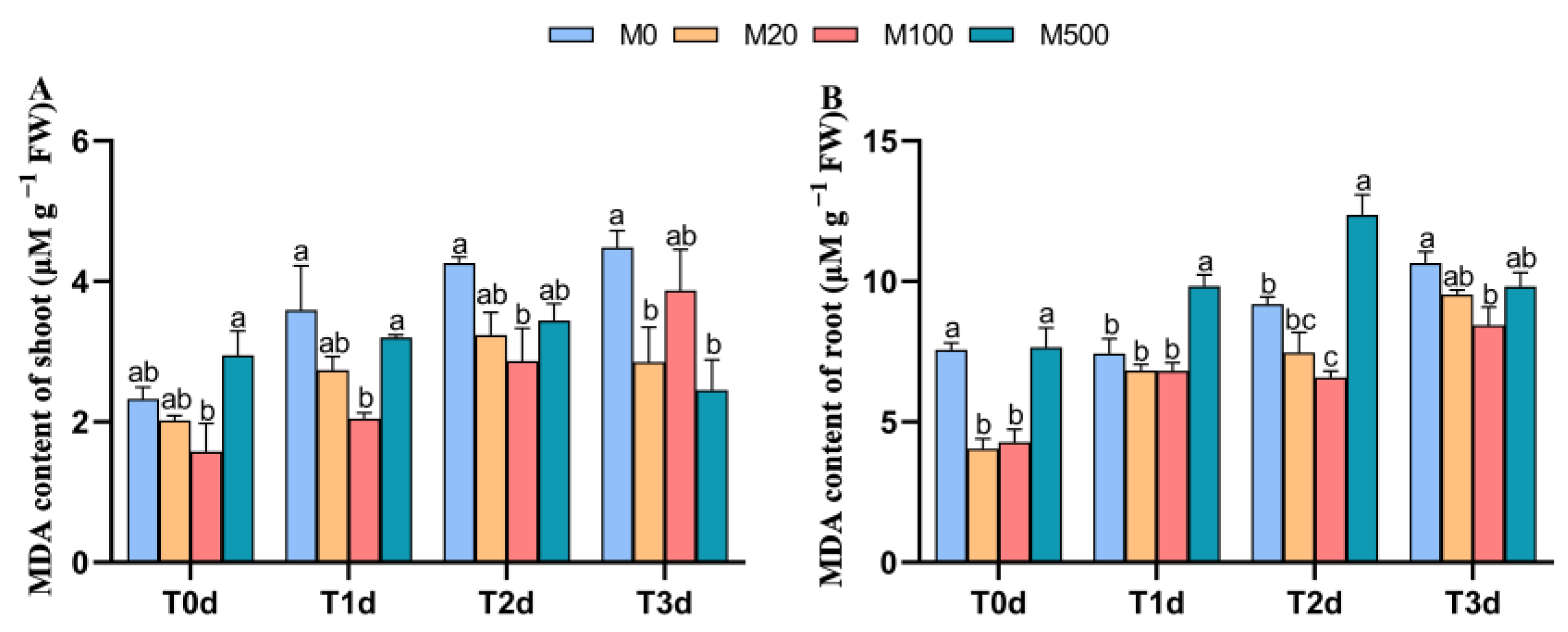
3.4. Malondialdehyde Content
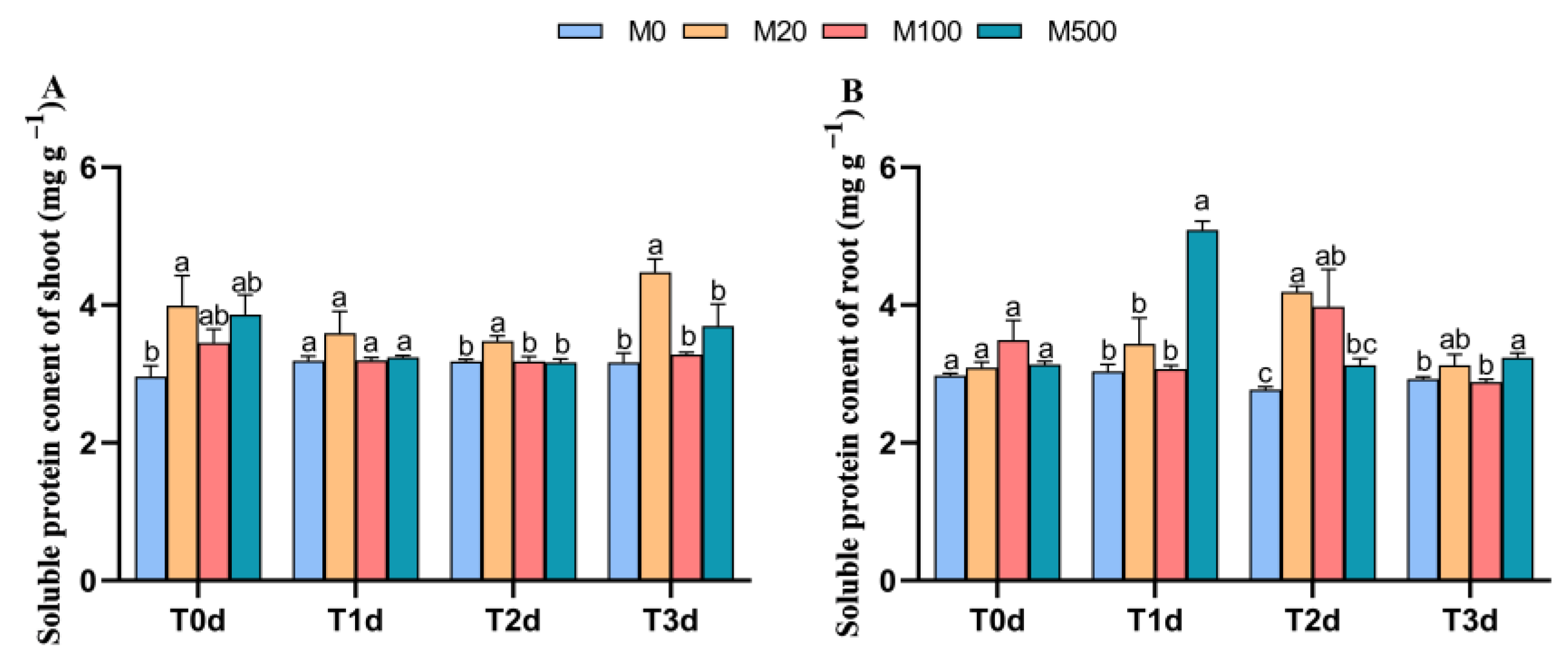
3.5. Soluble Protein Content
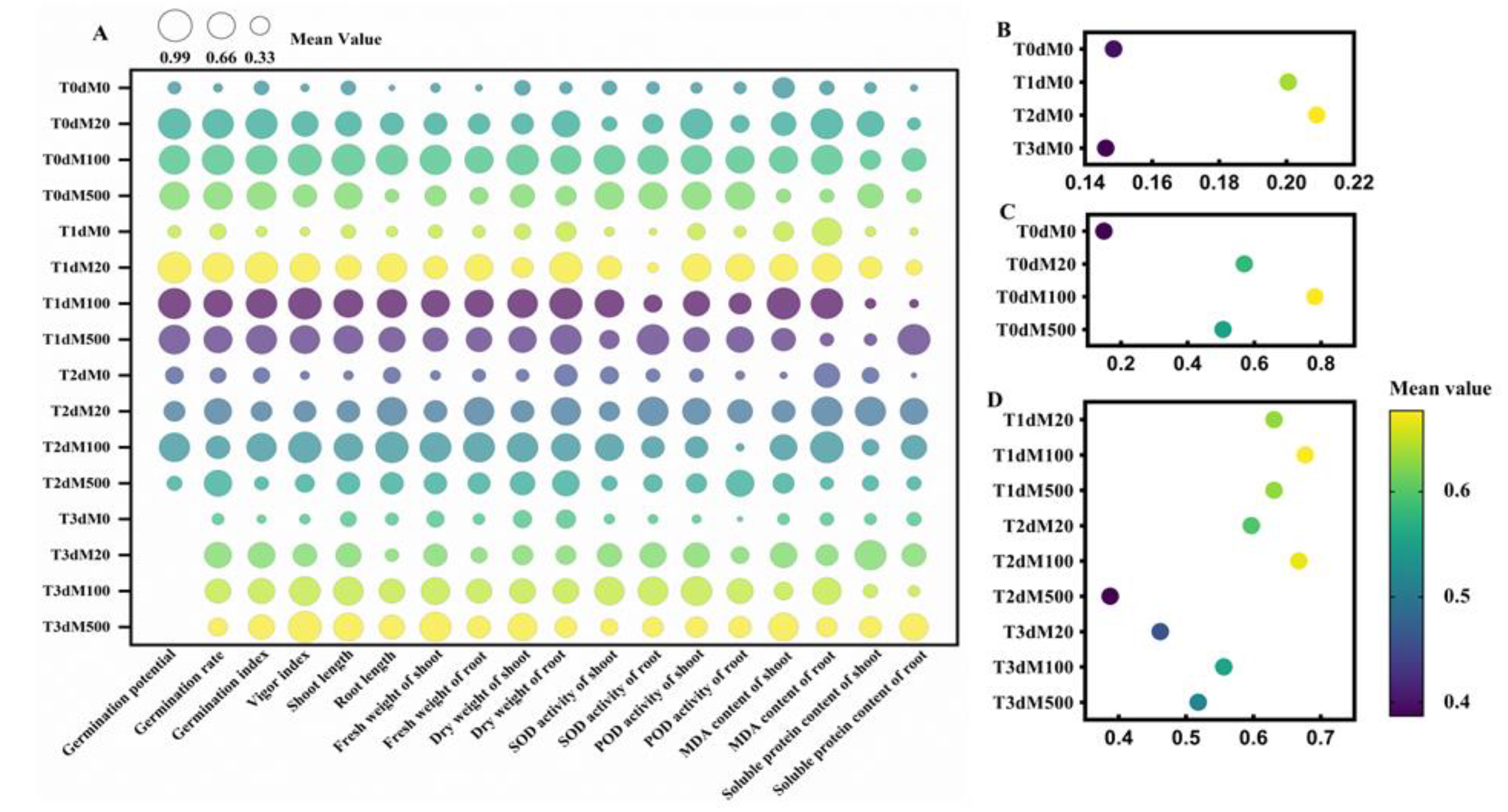
3.6. Grey Relational Grade of Exogenous Melatonin on Rice Seed Germination under Flooding Stress
3.7. Correlation Analysis of Exogenous Melatonin on Rice Seed Germination under Flooding Stress
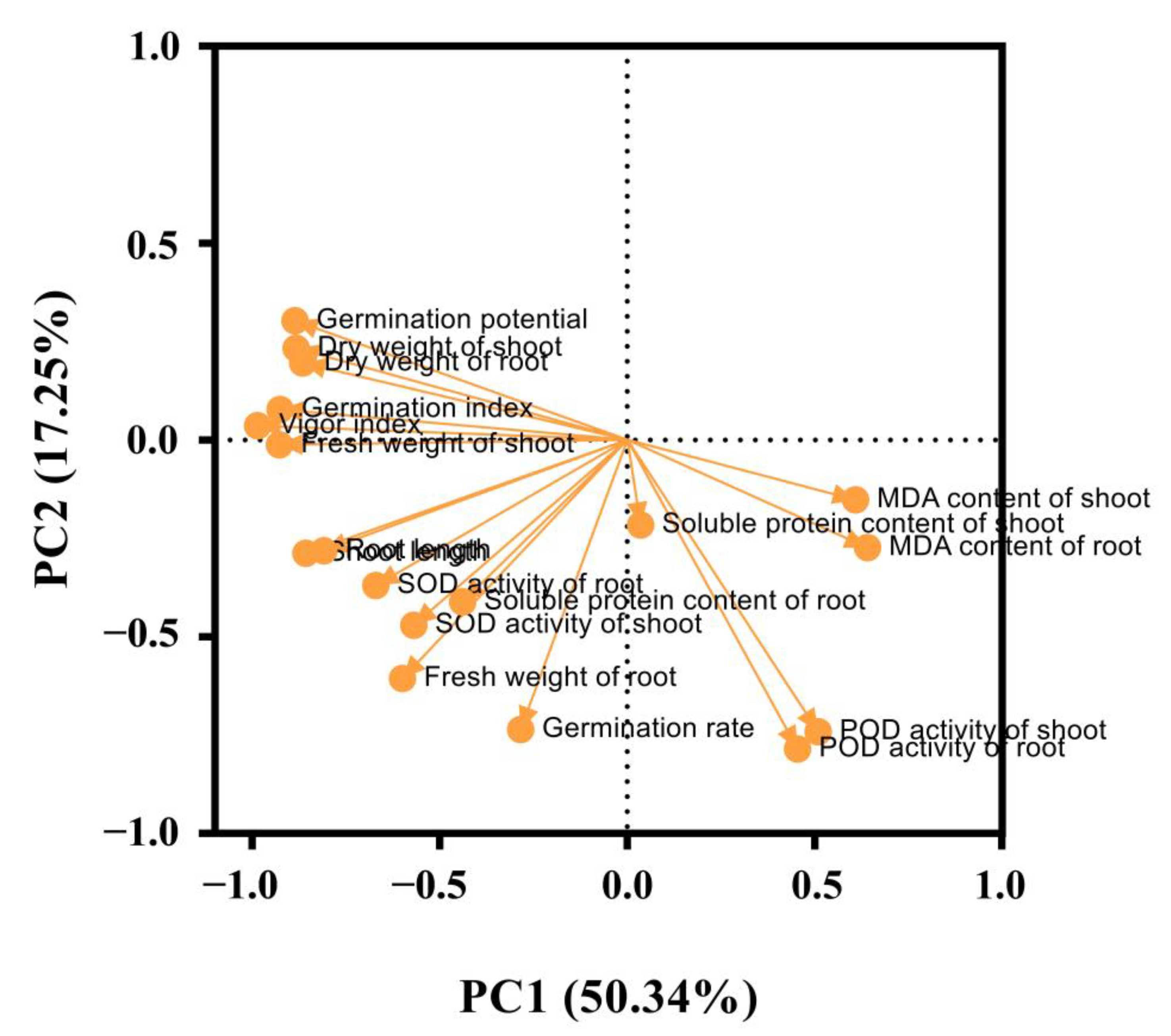
3.8. Principal Component Analysis of Exogenous Melatonin on Rice Seed Germination under Flooding Stress
3.9. Two-Way ANOVA of Exogenous Melatonin on Rice Seed Germination under Flooding Stress
3.10. Comprehensive Evaluation on the Tolerance of Exogenous Melatonin on Rice Seed Germination under Flooding Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, R.; Deng, Y.; Ding, Y.; Guo, J.; Qiu, J.; Wang, B.; Wang, C.; Xie, Y.; Zhang, Z.; Chen, J.; et al. Rice functional genomics: Decades’ efforts and roads ahead. Sci. China Life Sci. 2022, 65, 33–92. [Google Scholar] [CrossRef] [PubMed]
- Panneerselvam, P.; Kumar, V.; Banik, N.C.; Kumar, V.; Parida, N.; Wasim, I.; Das, A.; Pattnaik, S.; Roul, P.K.; Sarangi, D.R.; et al. Transforming labor requirement, crop yield, and profitability with precision dry-direct seeding of rice and integrated weed management in Eastern India. Field Crop. Res. 2020, 259, 107961. [Google Scholar] [CrossRef]
- Sandhu, N.; Subedi, S.R.; Singh, V.; Sinha, P.; Kumar, S.; Singh, S.P.; Ghimire, S.K.; Pandey, M.; Yadaw, R.B.; Varshney, R.K.; et al. Deciphering the genetic basis of root morphology, nutrient uptake, yield, and yield-related traits in rice under dry direct-seeded cultivation systems. Sci. Rep. 2019, 9, 9334. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.; Majhi, P.; Anandan, A.; Mahender, A.; Veludandi, S.; Bastia, D.; Guttala, S.; Singh, S.; Saha, S.; Ali, J. Proofing Direct-Seeded Rice with Better Root Plasticity and Architecture. Int. J. Mol. Sci. 2021, 22, 6058. [Google Scholar] [CrossRef]
- Wang, W.; Du, J.; Chen, L.; Zeng, Y.; Tan, X.; Shi, Q.; Pan, X.; Wu, Z.; Zeng, Y. Transcriptomic, proteomic, and physiological comparative analyses of flooding mitigation of the damage induced by low-temperature stress in direct seeded early indica rice at the seedling stage. BMC Genom. 2021, 22, 176. [Google Scholar] [CrossRef] [PubMed]
- van Dongen, J.T.; Gupta, K.J.; Ramírez-Aguilar, S.J.; Araújo, W.L.; Nunes-Nesi, A.; Fernie, A.R. Regulation of respiration in plants: A role for alternative metabolic pathways. J. Plant Physiol. 2011, 168, 1434–1443. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-Y.; Wu, Y.-S.; Chen, C.-T.; Lai, M.-H.; Yen, H.-M.; Yang, C.-Y. Physiological and molecular responses of seedlings of an upland rice (‘Tung Lu 3′) to total submergence compared to those of a submergence-tolerant lowland rice (‘FR13A’). Rice 2017, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Calleja-Cabrera, J.; Boter, M.; Oñate-Sánchez, L.; Pernas, M. Root Growth Adaptation to Climate Change in Crops. Front. Plant Sci. 2020, 11, 544. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Q.-Q.; Shen, T.-H.; Zhong, L.; Zhu, C.-L.; Peng, X.-S.; He, X.-P.; Fu, J.-R.; Ouyang, L.-J.; Bian, J.-M.; Hu, L.-F.; et al. Comprehensive metabolomic, proteomic and physiological analyses of grain yield reduction in rice under abrupt drought–flood alternation stress. Physiol. Plant. 2019, 167, 564–584. [Google Scholar] [CrossRef]
- Ismail, A.M.; Ella, E.S.; Vergara, G.V.; Mackill, D.J. Mechanisms associated with tolerance to flooding during germination and early seedling growth in rice (Oryza sativa). Ann. Bot. 2009, 103, 197–209. [Google Scholar] [CrossRef]
- Basu, S.; Kumari, S.; Kumar, P.; Kumar, G.; Rajwanshi, R. Redox imbalance impedes photosynthetic activity in rice by disrupting cellular membrane integrity and induces programmed cell death under submergence. Physiol. Plant. 2021, 172, 1764–1778. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Jiang, S.; Wang, Z.; Hu, K.; Xie, Y.; Zhou, L.; Zhu, J.; Xing, D.; Du, B. Seed priming with selenium: Effects on germination, seedling growth, biochemical attributes, and grain yield in rice growing under flooding conditions. Plant Direct 2022, 6, e378. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Ruiz, J.H. Melatonin: A New Plant Hormone and/or a Plant Master Regulator? Trends Plant Sci. 2019, 24, 38–48. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Ren, M.; Mora-Poblete, F.; Arnao, M.B.; Naz, S.; Anwar, M.; Shahid, S.; Shakoor, A.; Sohail, H.; et al. Phytomelatonin: An overview of the importance and mediating functions of melatonin against environmental stresses. Physiol. Plant. 2021, 172, 820–846. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wang, J.; Sun, Y.; Zhang, L.; Zheng, S. Versatile Roles of Melatonin in Growth and Stress Tolerance in Plants. J. Plant Growth Regul. 2022, 41, 507–523. [Google Scholar] [CrossRef]
- Cen, H.; Wang, T.; Liu, H.; Tian, D.; Zhang, Y. Melatonin Application Improves Salt Tolerance of Alfalfa (Medicago sativa L.) by Enhancing Antioxidant Capacity. Plants 2020, 9, 220. [Google Scholar] [CrossRef]
- Simlat, M.; Szewczyk, A.; Ptak, A. Melatonin promotes seed germination under salinity and enhances the biosynthesis of steviol glycosides in Stevia rebaudiana Bertoni leaves. PLoS ONE 2020, 15, e0230755. [Google Scholar] [CrossRef]
- Li, D.; Batchelor, W.D.; Zhang, D.; Miao, H.; Li, H.; Song, S.; Li, R. Analysis of melatonin regulation of germination and antioxidant metabolism in different wheat cultivars under polyethylene glycol stress. PLoS ONE 2020, 15, e0237536. [Google Scholar] [CrossRef]
- Bai, Y.; Xiao, S.; Zhang, Z.; Zhang, Y.; Sun, H.; Zhang, K.; Wang, X.; Bai, Z.; Li, C.; Liu, L. Melatonin improves the germination rate of cotton seeds under drought stress by opening pores in the seed coat. PeerJ 2020, 8, e9450. [Google Scholar] [CrossRef]
- Cao, Q.; Li, G.; Cui, Z.; Yang, F.; Jiang, X.; Diallo, L.; Kong, F. Seed Priming with Melatonin Improves the Seed Germination of Waxy Maize under Chilling Stress via Promoting the Antioxidant System and Starch Metabolism. Sci. Rep. 2019, 9, 15044. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Xu, W.; Liu, A.; Chen, S. Endogenous melatonin deficiency aggravates high temperature-induced oxidative stress in Solanum lycopersicum L. Environ. Exp. Bot. 2019, 161, 303–311. [Google Scholar] [CrossRef]
- Zhang, H.S.; Wang, Z.F. Seed Science, 3rd ed.; Science Press: Beijing, China, 2021; pp. 172–173. [Google Scholar]
- Vujicic, M.M.; Milosevic, S.; Sabovljevic, M.S.; Sabovljevic, A.D. Effect of ABA treatment on activities of antioxidative enzymes in selected bryophyte species. Bot. Serbica 2017, 41, 11–15. [Google Scholar] [CrossRef]
- Kochba, J.; Lavee, S.; Spiegel-Roy, P. Differences in peroxidase activity and isoenzymes in embryogenic ane non-embryogenic ‘Shamouti’ orange ovular callus lines. Plant Cell Physiol. 1977, 18, 463–467. [Google Scholar] [CrossRef]
- Li, R.; Xie, Y.; Tang, Y. Effect of Application Accumulator Plant Straw on the Osmotic Adjustment Substances and Malondialdehyde Content of Lettuce. IOP Conf. Ser. Earth Environ. Sci. 2019, 233, 042025. [Google Scholar] [CrossRef]
- Deans, C.A.; Sword, G.A.; Lenhart, P.A.; Burkness, E.; Hutchison, W.D.; Behmer, S.T. Quantifying Plant Soluble Protein and Digestible Carbohydrate Content, Using Corn (Zea mays) As an Exemplar. J. Vis. Exp. 2018, 138, e58164. [Google Scholar] [CrossRef] [PubMed]
- Su, D.W.; Mei, L.; Lin, H.; Luo, H.L.; Zhang, L.L.; Song, J.; Liu, J.; Lin, Z.X. Grey Correlation Analysis of Physiological Traits Related to Drought Tolerance in Pennisetum sp. Agric. Basic Sci. Technol. 2017, 18, 1158–1163. [Google Scholar] [CrossRef]
- Liu, J.X.; OU, X.B.; Liu, X.L.; Wang, J.C. Principal Component and Subordinate Function of the Alleviating Effects of Hydrogen Peroxide(H2O2) on Low-temperature Stress in Naked Oat (Avena nuda) Seedlings. Bull. Bot. Res. 2018, 38, 748–756. [Google Scholar] [CrossRef]
- Ma, M.; Cen, W.; Li, R.; Wang, S.; Luo, J. The Molecular Regulatory Pathways and Metabolic Adaptation in the Seed Germination and Early Seedling Growth of Rice in Response to Low O2 Stress. Plants 2020, 9, 1363. [Google Scholar] [CrossRef]
- Yu, Y.; Su, Y.H.; Zhang, C.L.; Yang, M.; Li, Z.H.; Zhang, L.J.; Pu, S.H.; Channarong, P.; Li, J.; Jin, S.L.; et al. Rice germination and biochemical indicator variation in the hypoxic condition. J. Yangzhou Univ. 2020, 41, 16–20. [Google Scholar] [CrossRef]
- Nurrahma, A.H.I.; Junaedi, A.; Purnamawati, H. Rice Tolerance to Flooding Stress on Germination Stage and Early Seedling Growth under 5 cm Water Depth. Agrosintesa J. Ilmu Budid. Pertan. 2019, 2, 40–47. [Google Scholar] [CrossRef]
- Li, H.; Guo, Y.; Lan, Z.; Zhang, Z.; Ahammed, G.J.; Chang, J.; Zhang, Y.; Wei, C.; Zhang, X. Melatonin antagonizes ABA action to promote seed germination by regulating Ca2+ efflux and H2O2 accumulation. Plant Sci. 2021, 303, 110761. [Google Scholar] [CrossRef]
- Chen, L.; Liu, L.; Lu, B.; Ma, T.; Jiang, D.; Li, J.; Zhang, K.; Sun, H.; Zhang, Y.; Bai, Z.; et al. Exogenous melatonin promotes seed germination and osmotic regulation under salt stress in cotton (Gossypium hirsutum L.). PLoS ONE 2020, 15, e0228241. [Google Scholar] [CrossRef]
- Xiao, S.; Liu, L.; Wang, H.; Li, D.; Bai, Z.; Zhang, Y.; Sun, H.; Zhang, K.; Li, C. Exogenous melatonin accelerates seed germination in cotton (Gossypium hirsutum L.). PLoS ONE 2019, 14, e0216575. [Google Scholar] [CrossRef]
- Ahmad, S.; Muhammad, I.; Wang, G.Y.; Zeeshan, M.; Yang, L.; Ali, I.; Zhou, X.B. Ameliorative effect of melatonin improves drought tolerance by regulating growth, photosynthetic traits and leaf ultrastructure of maize seedlings. BMC Plant Biol. 2021, 21, 368. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Zhao, X.; Liu, S.; Sun, F.; Zhang, C.; Xi, Y. Beneficial effects of melatonin in overcoming drought stress in wheat seedlings. Plant Physiol. Biochem. 2017, 118, 138–149. [Google Scholar] [CrossRef]
- Wu, S.; Wang, Y.; Zhang, J.; Gong, X.; Zhang, Z.; Sun, J.; Chen, X.; Wang, Y. Exogenous Melatonin Improves Physiological Characteristics and Promotes Growth of Strawberry Seedlings Under Cadmium Stress. Hortic. Plant J. 2021, 7, 13–22. [Google Scholar] [CrossRef]
- Mondal, S.; Khan, M.I.R.; Entila, F.; Dixit, S.; Cruz, P.C.S.; Ali, M.P.; Pittendrigh, B.; Septiningsih, E.M.; Ismail, A.M. Responses of AG1 and AG2 QTL introgression lines and seed pre-treatment on growth and physiological processes during anaerobic germination of rice under flooding. Sci. Rep. 2020, 10, 10214. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Chen, F.; Meng, Y.; Chandrasekaran, U.; Luo, X.; Yang, W.; Shu, K. Plant waterlogging/flooding stress responses: From seed germination to maturation. Plant Physiol. Biochem. 2020, 148, 228–236. [Google Scholar] [CrossRef]
- Basu, S.; Kumar, G.; Kumari, N.; Kumari, S.; Rajwanshi, R. Reactive oxygen species and reactive nitrogen species induce lysigenous aerenchyma formation through programmed cell death in rice roots under submergence. Environ. Exp. Bot. 2020, 177, 104118. [Google Scholar] [CrossRef]
- Shah, A.A.; Ahmed, S.; Ali, A.; Yasin, N.A. 2-Hydroxymelatonin mitigates cadmium stress in cucumis sativus seedlings: Modulation of antioxidant enzymes and polyamines. Chemosphere 2020, 243, 125308. [Google Scholar] [CrossRef]
- Gu, X.; Xue, L.; Lu, L.; Xiao, J.; Song, G.; Xie, M.; Zhang, H. Melatonin Enhances the Waterlogging Tolerance of Prunus persica by Modulating Antioxidant Metabolism and Anaerobic Respiration. J. Plant Growth Regul. 2021, 40, 1–13. [Google Scholar] [CrossRef]
- Chen, D.; Li, Q.; Peng, Y.; Wu, T.H.; Zhang, X.L.; Dong, J.Y.; Mao, B.G.; Zhao, B.R. Effect of Melatonin on Rice Seedling Growth under Submergence Stress. Acta Agric. Boreali-Sin. 2019, 34, 129–136. [Google Scholar] [CrossRef]
- Liu, N.; Lin, Z.; Guan, L.; Gaughan, G.; Lin, G. Antioxidant Enzymes Regulate Reactive Oxygen Species during Pod Elongation in Pisum sativum and Brassica chinensis. PLoS ONE 2014, 9, e87588. [Google Scholar] [CrossRef]
- Liao, B.T.; Chen, L.; Tang, K.L.; Wen, M.Y.; Liu, J.M.; Han, R. Germination and drought tolerance evaluation of pepper seeds under PEG simulated drought stress. China Cucurbits Veg. 2022, 35, 64–69. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Flooding Stress | Melatonin Treatment | Interactive Treatment | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Df | F | p | Df | F | p | Df | F | p | |
| Germination potential | 3 | 2167.27 | <0.001 | 3 | 59.35 | <0.001 | 9 | 17.61 | <0.001 |
| Germination rate | 3 | 8.05 | <0.001 | 3 | 74.56 | <0.001 | 9 | 8.14 | <0.001 |
| Germination index | 3 | 597.74 | <0.001 | 3 | 106.78 | <0.001 | 9 | 19.25 | <0.001 |
| Vigor index | 3 | 2588.13 | <0.001 | 3 | 883.81 | <0.001 | 9 | 79.89 | <0.001 |
| Shoot length | 3 | 79.03 | <0.001 | 3 | 123.04 | <0.001 | 9 | 3.67 | 0.001 |
| Root length | 3 | 55.71 | <0.001 | 3 | 92.52 | <0.001 | 9 | 15.84 | <0.001 |
| Fresh weight of shoot | 3 | 99.42 | <0.001 | 3 | 47.25 | <0.001 | 9 | 2.84 | 0.007 |
| Fresh weight of root | 3 | 17.06 | <0.001 | 3 | 47.8 | <0.001 | 9 | 4.75 | <0.001 |
| Dry weight of shoot | 3 | 141.87 | <0.001 | 3 | 30.79 | <0.001 | 9 | 1.6 | 0.134 |
| Dry weight of root | 3 | 123.89 | <0.001 | 3 | 29.15 | <0.001 | 9 | 5.49 | <0.001 |
| SOD activity of shoot | 3 | 154.71 | <0.001 | 3 | 71.79 | <0.001 | 9 | 9.98 | <0.001 |
| SOD activity of root | 3 | 331.47 | <0.001 | 3 | 151.56 | <0.001 | 9 | 63.02 | <0.001 |
| POD activity of shoot | 3 | 243.64 | <0.001 | 3 | 59.58 | <0.001 | 9 | 13.12 | <0.001 |
| POD activity of root | 3 | 139.78 | <0.001 | 3 | 34.04 | <0.001 | 9 | 7.45 | <0.001 |
| MDA content of shoot | 3 | 36.11 | <0.001 | 3 | 25.04 | <0.001 | 9 | 9.57 | <0.001 |
| MDA content of root | 3 | 166.63 | <0.001 | 3 | 154.93 | <0.001 | 9 | 18.32 | <0.001 |
| Soluble protein content of shoot | 3 | 13.57 | <0.001 | 3 | 37.8 | <0.001 | 9 | 5.62 | <0.001 |
| Soluble protein content of root | 3 | 29.47 | <0.001 | 3 | 33.02 | <0.001 | 9 | 32.83 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, H.; Liu, M.; Wang, X.; Liu, L.; Wu, H.; Chen, X.; Wang, H.; Shen, Q.; Chen, G.; Wang, Y. Seed-Soaking with Melatonin for the Improvement of Seed Germination, Seedling Growth, and the Antioxidant Defense System under Flooding Stress. Agronomy 2022, 12, 1918. https://doi.org/10.3390/agronomy12081918
Zeng H, Liu M, Wang X, Liu L, Wu H, Chen X, Wang H, Shen Q, Chen G, Wang Y. Seed-Soaking with Melatonin for the Improvement of Seed Germination, Seedling Growth, and the Antioxidant Defense System under Flooding Stress. Agronomy. 2022; 12(8):1918. https://doi.org/10.3390/agronomy12081918
Chicago/Turabian StyleZeng, Hongli, Minghao Liu, Xin Wang, Ling Liu, Huiyi Wu, Xuan Chen, Haodong Wang, Quansheng Shen, Guanghui Chen, and Yue Wang. 2022. "Seed-Soaking with Melatonin for the Improvement of Seed Germination, Seedling Growth, and the Antioxidant Defense System under Flooding Stress" Agronomy 12, no. 8: 1918. https://doi.org/10.3390/agronomy12081918
APA StyleZeng, H., Liu, M., Wang, X., Liu, L., Wu, H., Chen, X., Wang, H., Shen, Q., Chen, G., & Wang, Y. (2022). Seed-Soaking with Melatonin for the Improvement of Seed Germination, Seedling Growth, and the Antioxidant Defense System under Flooding Stress. Agronomy, 12(8), 1918. https://doi.org/10.3390/agronomy12081918