
Susceptibility of Cassava Varieties to Disease Caused by Sri Lankan Cassava Mosaic Virus and Impacts on Yield by Use of Asymptomatic and Virus-Free Planting Material

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Measurements
3. Results
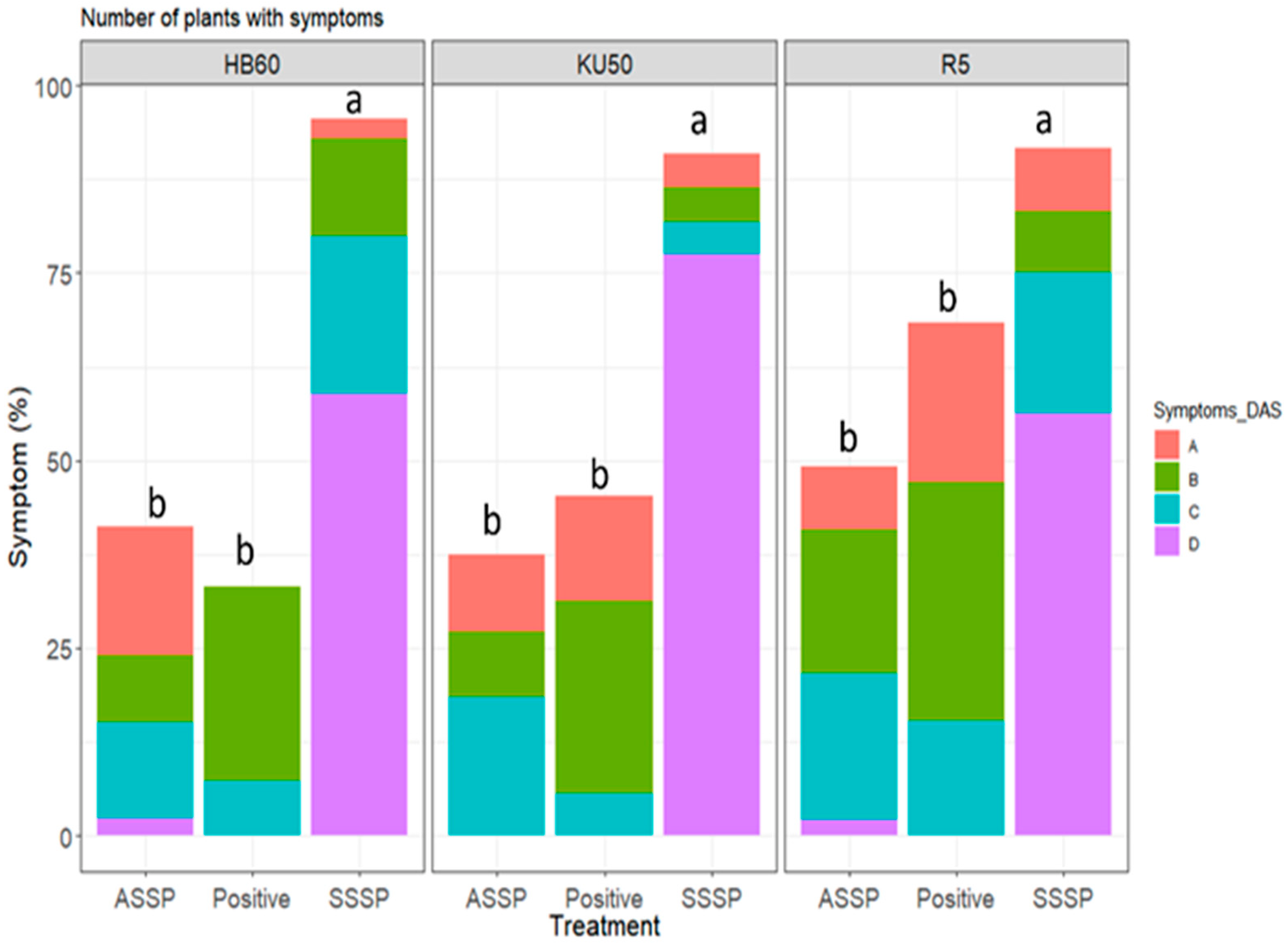
3.1. Disease Symptoms
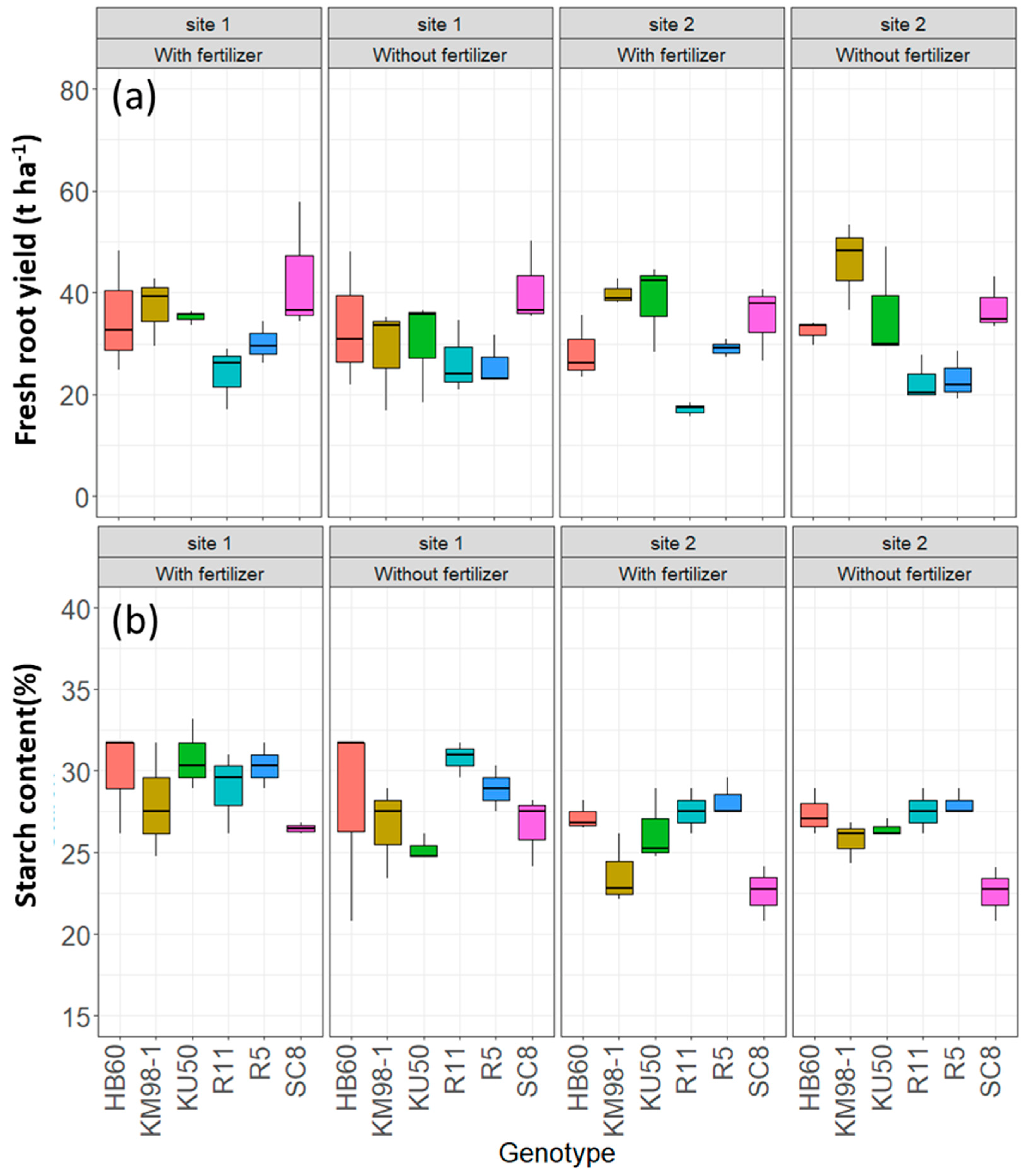
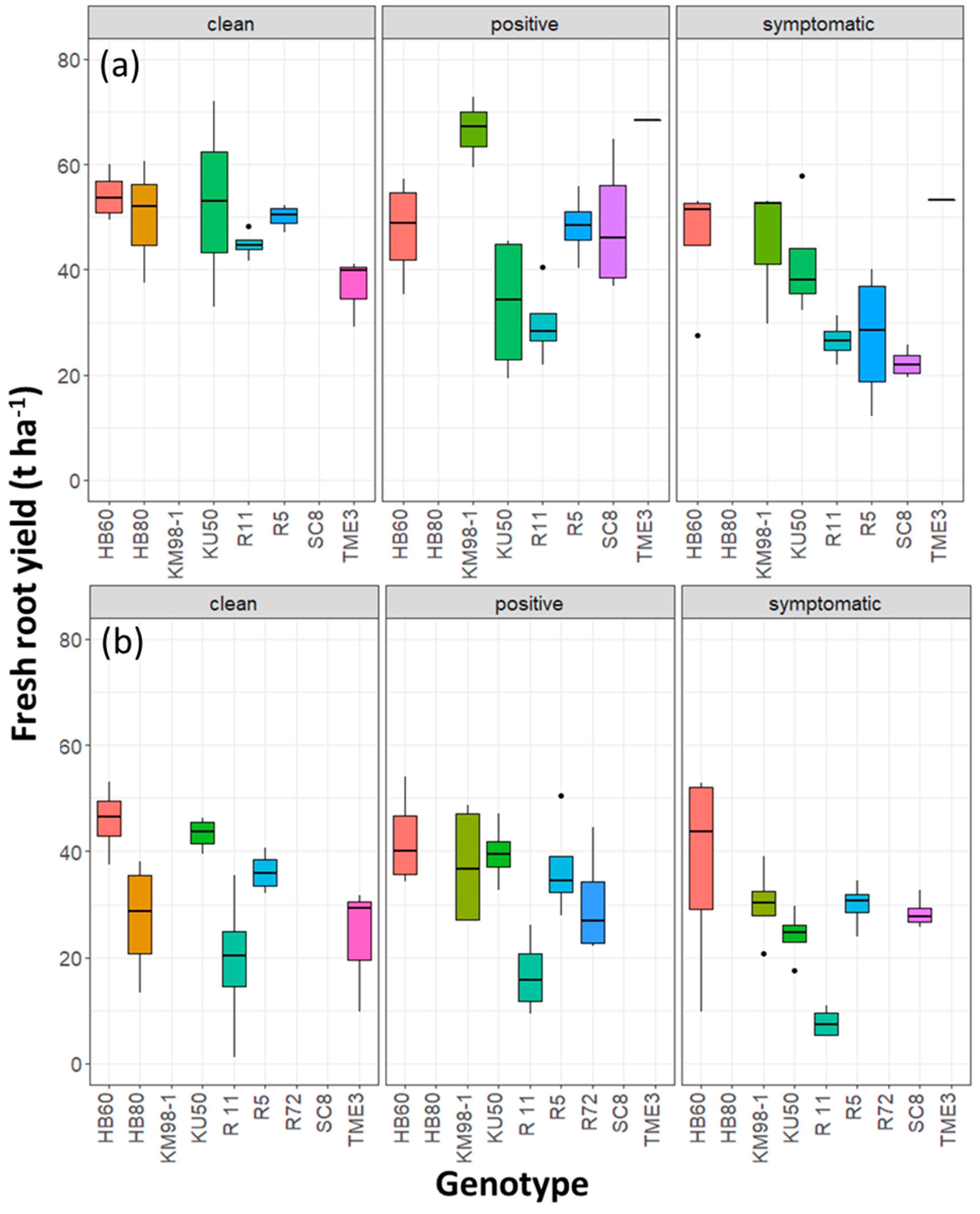
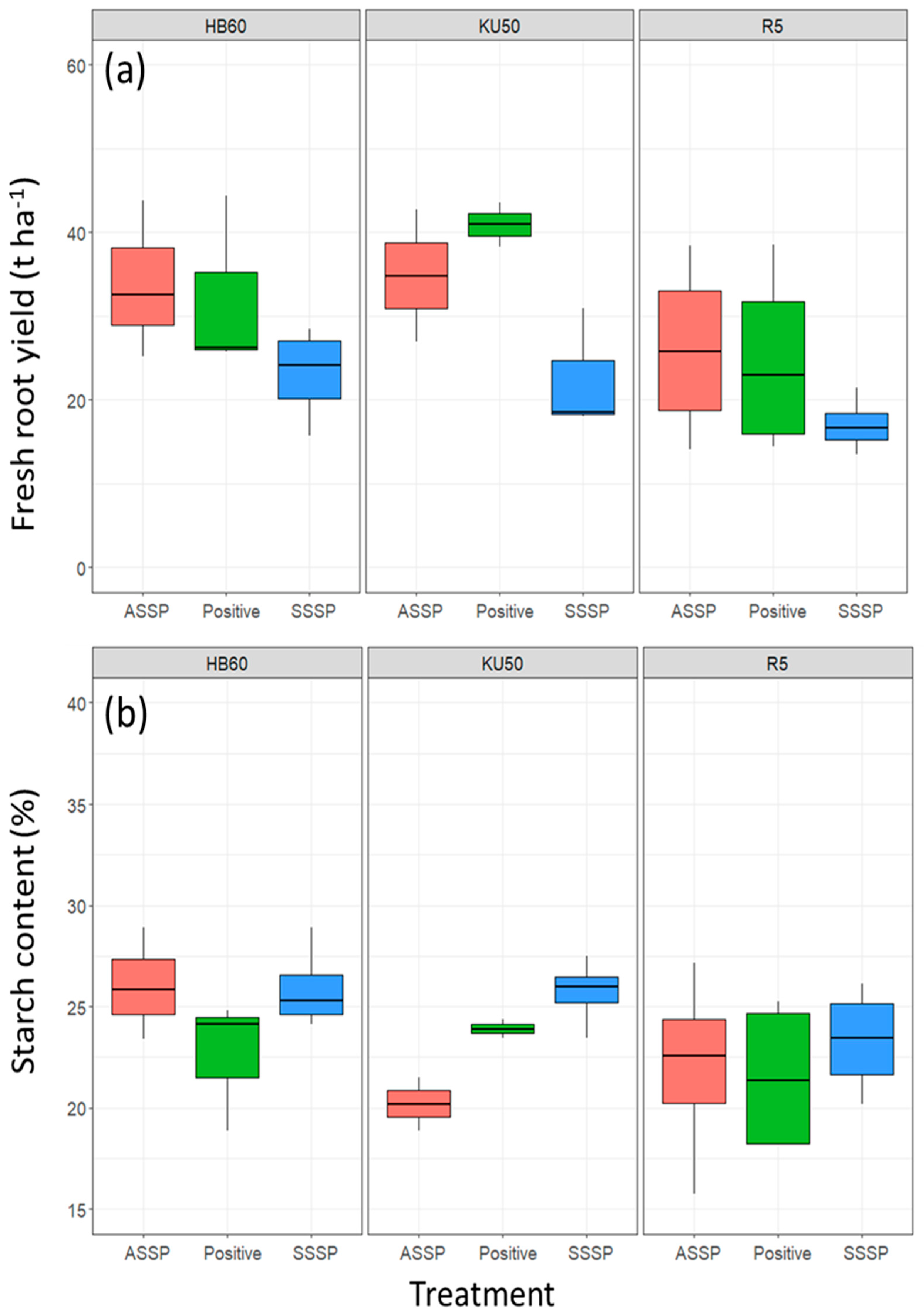
3.2. Yield and Starch Content
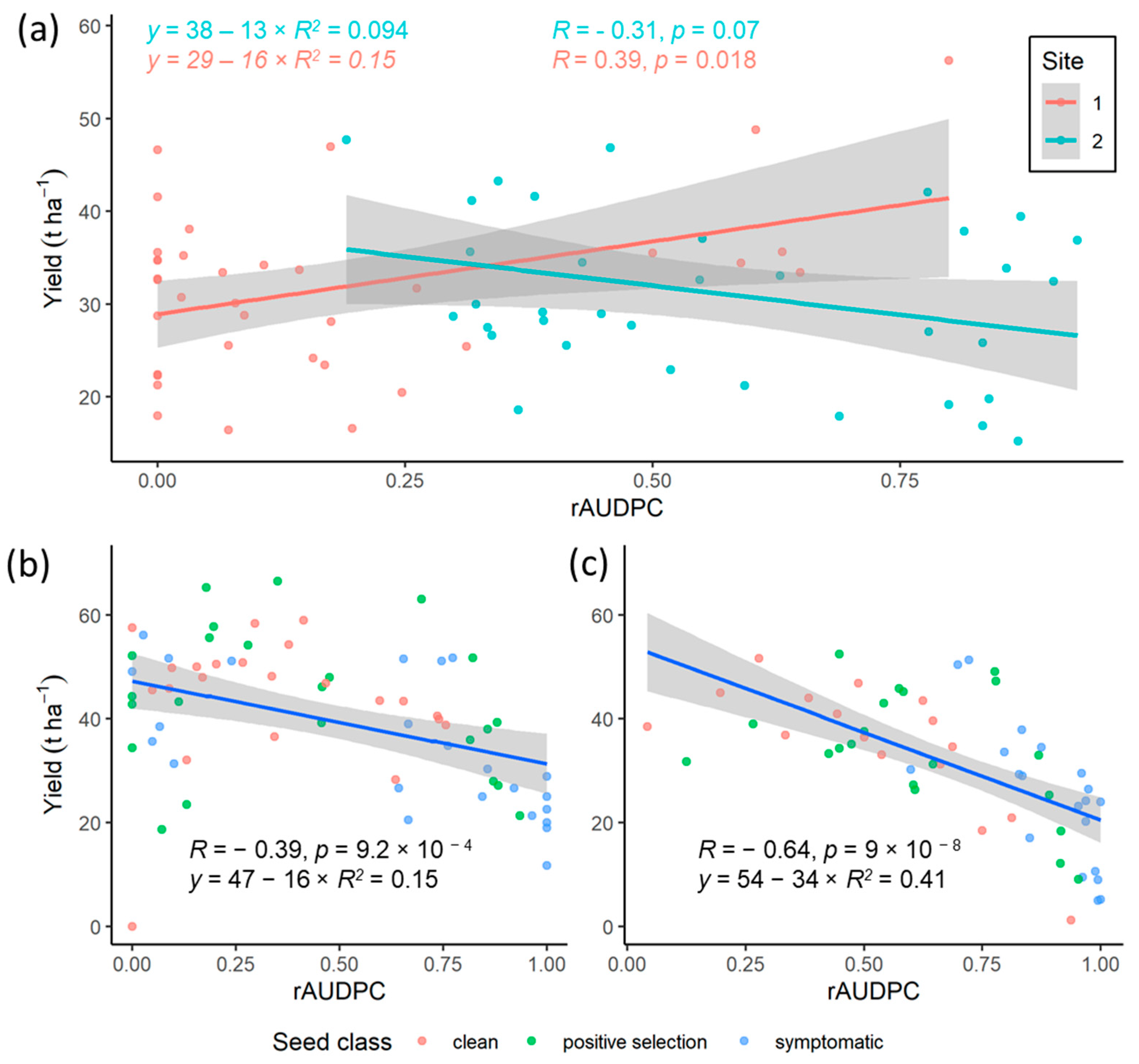
3.2.1. Relationship between Onset of Symptoms and Root Yield
3.2.2. Analysis of Asymptomatic Plants
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malik, A.I.; Kongsil, P.; Nguyễn, V.A.; Ou, W.; Sholihin; Srean, P.; Sheela, M.; López-Lavalle, L.A.B.; Utsumi, Y.; Lu, C.; et al. Cassava breeding and agronomy in Asia: 50 years of history and future directions. Breed. Sci. 2020, 70, 145–166. [Google Scholar] [CrossRef]
- Howeler, R.H.; Aye, T.M. Sustainable Management of Cassava in Asia—From Research to Practice; International Center for Tropical Agriculture (CIAT), The Nippon Foundation: Hanoi, Vietnam, 2014; p. 168. [Google Scholar]
- Graziosi, I.; Minato, N.; Alvarez, E.; Ngo, D.T.; Hoat, T.X.; Aye, T.M.; Pardo, J.M.; Wongtiem, P.; Wyckhuys, K.A. Emerging pests and diseases of South-east Asian cassava: A comprehensive evaluation of geographic priorities, management options and research needs. Pest Manag. Sci. 2016, 72, 1071–1089. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.; Cui, X.Y.; Wang, X.W.; Liu, S.S.; Zhang, Z.H.; Zhou, X.P. First Report of Sri Lankan cassava mosaic virus Infecting Cassava in Cambodia. Plant Dis. 2016, 100, 1029. [Google Scholar] [CrossRef]
- Uke, A.; Hoat, T.X.; Quan, M.V.; Liem, N.V.; Ugaki, M.; Natsuaki, K.T. First report of Sri Lankan cassava mosaic virus infecting cassava in Vietnam. Plant Dis. 2019, 102, 2669. [Google Scholar] [CrossRef]
- Wang, D.; Yao, X.M.; Huang, G.X.; Shi, T.; Wang, G.F.; Ye, J. First Report of Sri Lankan Cassava Mosaic Virus Infected Cassava in China. Plant Dis. 2019, 103, 1437. [Google Scholar] [CrossRef]
- Siriwan, W.; Jimenez, J.; Hemniam, N.; Saokham, K.; Lopez-Alvarez, D.; Leiva, A.M.; Martinez, A.; Mwanzia, L.; Lopez-Lavalle, L.A.B.; Cuellar, W.J. Surveillance and diagnostics of the emergent Sri Lankan cassava mosaic virus (Fam. Geminiviridae) in Southeast Asia. Virus Res. 2020, 285, 197959. [Google Scholar] [CrossRef] [PubMed]
- Chittarath, K.; Jimenez, J.; Vongphachanh, P.; Leiva, A.M.; Sengsay, S.; Lopez-Alvarez, D.; Bounvilayvong, T.; Lourido, D.; Vorlachith, V.; Cuellar, W.J. First Report of Cassava Mosaic Disease and Sri Lankan Cassava Mosaic Virus in Laos. Plant Dis. 2021, 105, 1861. [Google Scholar] [CrossRef] [PubMed]
- Delaquis, E.; Andersen, K.F.; Minato, N.; Le Cu, T.T.; Karssenberg, M.E.; Sok, S.; Wyckhuys, K.A.G.; Newby, J.C.; Burra, D.D.; Srean, P.; et al. Raising the Stakes: Cassava Seed Networks at Multiple Scales in Cambodia and Vietnam. Front. Sustain. Food Syst. 2018, 2, 73. [Google Scholar] [CrossRef]
- Götz, M.; Winter, S. Diversity of Bemisia tabaci in Thailand and Vietnam and indications of species replacement. J. Asia Pac. Èntomol. 2016, 19, 537–543. [Google Scholar] [CrossRef]
- Chi, Y.; Pan, L.-L.; Bouvaine, S.; Fan, Y.-Y.; Liu, Y.-Q.; Liu, S.-S.; Seal, S.; Wang, X.-W. Differential transmission of Sri Lankan cassava mosaic virus by three cryptic species of the whitefly Bemisia tabaci complex. Virology 2019, 540, 141–149. [Google Scholar] [CrossRef]
- FAOSTAT. Food and Agricultural Organization of the United Nations. Available online: www.fao.org/faostat/en (accessed on 13 July 2021).
- Alabi, O.J.; Ogbe, F.O.; Bandyopadhyay, R.; Kumar, P.L.; Dixon, A.G.O.; Hughes, J.D.; Naidu, R.A. Alternate hosts of African cassava mosaic virus and East African cassava mosaic Cameroon virus in Nigeria. Arch. Virol. 2008, 153, 1743–1747. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, H.; Rojanaridpiched, C.; Phumichai, C.; Becerra, L.A.; Kittipadakul, P.; Iglesias, C.; Gracen, V.E. Excellence in Cassava Breeding: Perspectives for the Future. Crop Breed. Genet. Genom. 2020, 2, e200008. [Google Scholar] [CrossRef]
- ACIAR (Australian Centre for International Agriculture). Establishing Sustainable Solutions to Cassava Diseases in Mainland Southeast Asia. Available online: https://www.aciar.gov.au/project/agb-2018-172 (accessed on 21 July 2021).
- Fauquet, C. African Cassava Mosaic Virus: Etiology, Epidemiology, and Control. Plant Dis. 1990, 74, 404. [Google Scholar] [CrossRef]
- McQuaid, C.F.; Sseruwagi, P.; Pariyo, A.; Bosch, F.V.D. Cassava brown streak disease and the sustainability of a clean seed system. Plant Pathol. 2015, 65, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Pérez, J.C.; Ceballos, H.; Ramírez, I.C.; Lenis, J.I.; Calle, F.; Morante, N.; Jaramillo, G.; Lentini, M. Adjustment for missing plants in cassava evaluation trials. Euphytica 2009, 172, 59–65. [Google Scholar] [CrossRef]
- Howeler, R.H. Effect of cassava production on soil fertility and the long-term fertilizer requirements to maintain high yields. In The Cassava Handbook: A Reference Manual Based on the Asian Regional Cassava Training Course, Held in Thailand; Howeler, R.H., Ed.; Centro Internacional de Agricultura Tropical (CIAT): Bangkok, Thailand, 2012; pp. 411–428. [Google Scholar]
- Chua, M.F.; Youbee, L.; Oudthachit, S.; Khanthavong, P.; Veneklaas, E.J.; Malik, A.I. Potassium Fertilisation Is Required to Sustain Cassava Yield and Soil Fertility. Agronomy 2020, 10, 1103. [Google Scholar] [CrossRef]
- Jeger, M.J.; Viljanen-Rollinson, S.L.H. The use of the area under the disease-progress curve (AUDPC) to assess quantitative disease resistance in crop cultivars. Theor. Appl. Genet. 2001, 102, 32–40. [Google Scholar] [CrossRef]
- Shaner, G. The Effect of Nitrogen Fertilization on the Expression of Slow-Mildewing Resistance in Knox Wheat. Phytopathology 1977, 67, 1051–1056. [Google Scholar] [CrossRef]
- R Development Core Team 2013. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: https://www.R-project.org/ (accessed on 2 August 2021).
- Jimenez, J.; Leiva, A.M.; Olaya, C.; Acosta-Trujillo, D.; Cuellar, W.J. An optimized nucleic acid isolation protocol for virus diagnostics in cassava (Manihot esculenta Crantz.). MethodsX 2021, 8, 101496. [Google Scholar] [CrossRef]
- Dutt, N.; Briddon, R.W.; Dasgupta, I. Identification of a second begomovirus, Sri Lankan cassava mosaic virus, causing cassava mosaic disease in India. Arch. Virol. 2005, 150, 2101–2108. [Google Scholar] [CrossRef] [PubMed]
- Leiva, A.M.; Siriwan, W.; Lopez-Alvarez, D.; Barrantes, I.; Hemniam, N.; Saokham, K.; Cuellar, W.J. Nanopore-Based Complete Genome Sequence of a Sri Lankan Cassava Mosaic Virus (Geminivirus) Strain from Thailand. Microbiol. Resour. Announc. 2020, 9, e01274-19. [Google Scholar] [CrossRef] [PubMed]
- Cuellar, W.J.; Mwanzia, L.; Lourido, D.; Martinez, A.F.; Rodriguez, R.; Garcia, C.; PestDisPlace: Monitoring the distribution of pests and diseases. International Center for Tropical Agriculture (CIAT). 2018. Version 3.0. Available online: https://pestdisplace.org (accessed on 8 July 2021).
- Minato, N.; Sok, S.; Chen, S.; Delaquis, E.; Phirun, I.; Le, V.X.; Burra, D.D.; Newby, J.C.; Wyckhuys, K.; De Haan, S. Surveillance for Sri Lankan cassava mosaic virus (SLCMV) in Cambodia and Vietnam one year after its initial detection in a single plantation in 2015. PLoS ONE 2019, 14, e0212780. [Google Scholar] [CrossRef]
- Abarshi, M.; Mohammed, I.; Wasswa, P.; Hillocks, R.; Holt, J.; Legg, J.; Seal, S.; Maruthi, M. Optimization of diagnostic RT-PCR protocols and sampling procedures for the reliable and cost-effective detection of Cassava brown streak virus. J. Virol. Methods 2010, 163, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Debibakas, S.; Rocher, S.; Garsmeur, O.; Toubi, L.; Roques, D.; D’Hont, A.; Hoarau, J.-Y.; Daugrois, J.H. Prospecting sugarcane resistance to Sugarcane yellow leaf virus by genome-wide association. Theor. Appl. Genet. 2014, 127, 1719–1732. [Google Scholar] [CrossRef]
- Sharma, R.; Bhardwaj, V.; Dalamu, D.; Kaushik, S.; Singh, B.; Sharma, S.K.; Umamaheshwari, R.; Baswaraj, R.; Kumar, V.; Gebhardt, C. Identification of elite potato genotypes possessing multiple disease resistance genes through molecular approaches. Sci. Hortic. 2014, 179, 204–211. [Google Scholar] [CrossRef]
- Aritua, V.; Legg, J.P.; Smit, N.E.J.M.; Gibson, R.W. Effect of local inoculum on the spread of sweet potato virus disease: Limited infection of susceptible cultivars following widespread cultivation of a resistant sweet potato cultivar. Plant Pathol. 1999, 48, 655–661. [Google Scholar] [CrossRef]
- Houngue, J.A.; Zandjanakou-Tachin, M.; Ngalle, H.B.; Pita, J.S.; Cacaï, G.H.T.; Ngatat, S.E.; Bell, J.M.; Ahanhanzo, C. Evaluation of resistance to cassava mosaic disease in selected African cassava cultivars using combined molecular and greenhouse grafting tools. Physiol. Mol. Plant Pathol. 2019, 105, 47–53. [Google Scholar] [CrossRef]
- Sseruwagi, P.; Sserubombwe, W.; Legg, J.; Ndunguru, J.; Thresh, J. Methods of surveying the incidence and severity of cassava mosaic disease and whitefly vector populations on cassava in Africa: A review. Virus Res. 2004, 100, 129–142. [Google Scholar] [CrossRef]
- Jose, A.; Makeshkumar, T.; Edison, S. Survey of cassava mosaic disease in Kerela. J. Root Crops 2011, 37, 41–47. [Google Scholar]
- Tembo, M.; Mataa, M.; Legg, J.; Chikoti, P.; Ntawuruhunga, P. Cassava Mosaic Disease: Incidence and Yield Performance of Cassava Cultivars in Zambia. J. Plant Pathol. 2017, 99, 681–689. [Google Scholar] [CrossRef]
- Milgram, M.; Cohen, J.; Loebenstein, G. Effects of sweet potato feathery mottle virus and sweet potato sunken vein virus on sweet potato yields and rates of reinfection of virus-Free planting material in Israel. Phytoparasitica 1996, 24, 189–193. [Google Scholar] [CrossRef]
- Ogero, K.O.; Kreuze, J.; McEwan, M.A.; Luambano, N.D.; Bachwenkizi, H.; Garrett, K.A.; Andersen, K.F.; Thomas-Sharma, S.; Van Der Vlugt, R.A.A. Efficiency of insect-proof net tunnels in reducing virus-related seed degeneration in sweet potato. Plant Pathol. 2019, 68, 1472–1480. [Google Scholar] [CrossRef] [PubMed]
- Muimba-Kankolongo, A.; Phuti, K. Relationship of Cassava Mosaic Severity in Planting Material to Mosaic Development, Growth and Yield of Cassava in Zaire. Exp. Agric. 1987, 23, 221–225. [Google Scholar] [CrossRef]
- McQuaid, C.F.; Bosch, F.V.D.; Szyniszewska, A.; Alicai, T.; Pariyo, A.; Chikoti, P.C.; Gilligan, C.A. Spatial dynamics and control of a crop pathogen with mixed-mode transmission. PLoS Comput. Biol. 2017, 13, e1005654. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, K.; Omondi, B.A.; Almekinders, C.; Alvarez, E.; Blomme, G.; Dita, M.; Iskra-Caruana, M.-L.; Ocimati, W.; Tinzaara, W.; Kumar, P.L.; et al. Seed degeneration of banana planting materials: Strategies for improved farmer access to healthy seed. Plant Pathol. 2018, 68, 207–228. [Google Scholar] [CrossRef]
- Thresh, J.M.; Fargette, D.; Otim-Nape, G.W. Effects of African cassava mosaic geminivirus on the yield of cassava. Trop. Sci. 1994, 34, 26–42. [Google Scholar]
- Gibson, R.W.; Otim-Nape, G.W. Factors determining recovery and reversion in mosaic-affected African cassava mosaic virus resistant cassava. Ann. Appl. Biol. 1997, 131, 259–271. [Google Scholar] [CrossRef]
- Fondong, V.N.; Thresh, J.M.; Fauquet, C. Field experiments in Cameroon on cassava mosaic virus disease and the reversion phenomenon in susceptible and resistant cassava cultivars. Int. J. Pest Manag. 2000, 46, 211–217. [Google Scholar] [CrossRef]
- Jennings, D.L. Further Studies in Breeding Cassava for Virus Resistance. East Afr. Agric. J. 1957, 22, 213–219. [Google Scholar] [CrossRef]
- Beyene, G.; Chauhan, R.D.; Wagaba, H.; Moll, T.; Alicai, T.; Miano, D.; Carrington, J.C.; Taylor, N.J. Loss of CMD2-mediated resistance to cassava mosaic disease in plants regenerated through somatic embryogenesis. Mol. Plant Pathol. 2016, 17, 1095–1110. [Google Scholar] [CrossRef]
- Fargette, D. Simulation of the Effects of Host Resistance, Reversion, and Cutting Selection on Incidence of African Cassava Mosaic Virus and Yield Losses in Cassava. Phytopathology 1995, 85, 370. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety (Long Name) | Variety (Code) | Origin | Genetic Background/ Pedigree * | Source of Planting Material | ||
|---|---|---|---|---|---|---|
| 2018–2019 | 2019–2020 | 2020–2021 | ||||
| Kasetsart University 50 | KU50 | Thailand | R1 × R 90 | HBRST | ||
| Rayong 11 | R11 | Thailand | R5 × OMR 29-20-118 | HBRST | ||
| South China 8 | SC8 | China | CMR38-120-10 | CLSKC | ||
| Huay Bong 60 | HB60 | Thailand | R5 × KU50 | CLSKC | ||
| KM98-1(6) | KM98-1 | Vietnam | R1 × R5 | CLSKC | ||
| Rayong 5 | R5 | Thailand | 22-27-10 XX × R3 | CLSKC | ||
| Tropical Manihot Esculenta 3 | TME3 | West Africa | Landrace | X | TTDI | TTDI |
| Huay Bong 80 | HB80 | Thailand | R5 × KU50 | X | TTDI | TTDI |
| Rayong 72 | R72 | Thailand | R1 × R5 | X | X | TTDI |
| Symptom Incidence (%) | |||||
|---|---|---|---|---|---|
| Site | Variety | 60 DAP | 150 DAP | 270 DAP | AUDPC |
| Site 1 | HB60 | 0 b | 14 b | 14 c | 2353 c |
| KM98-1 | 0 b | 3b c | 6 c | 736 cd | |
| KU50 | 0 b | 0 c | 2 c | 91 d | |
| R11 | 6 b | 12 b | 47 b | 4347 b | |
| R5 | 0 b | 5b c | 7 c | 868 cd | |
| SC8 | 16 a | 65 a | 94 a | 13201 a | |
| HSD c | 8 | 11 | 16 | 1886 | |
| Site 2 | HB60 | 20 ab | 44 b | 82 ab | 10442 b |
| KM98-1 | 19 ab | 35 b | 90 ab | 9938 b | |
| KU50 | 12 b | 26 b | 55 c | 6556 c | |
| R11 | 32 ab | 90 a | 100 a | 16825 a | |
| R5 | 17 ab | 35 b | 71b c | 8695b c | |
| SC8 | 44 a | 97 a | 100 a | 18106 a | |
| HSD | 28 | 20 | 23 | 10442 | |
| Symptom Incidence (%) | |||||||
|---|---|---|---|---|---|---|---|
| Year | Variety | Seed Class | 30 DAP | 60 DAP | 150 DAP | 270 DAP | AUDPC |
| 2019 | HB60 | Clean | 7 b | 25 cd | 48 cde | 5804 cd | |
| Positive | 0 b | 5 d | 8 f | 975 d | |||
| Symptomatic | 14 b | 26 cd | 29 def | 5095 cd | |||
| KU50 | Clean | 0 b | 4 d | 17 def | 1438 d | ||
| Positive | 0 b | 8 d | 14 ef | 1652 d | |||
| Symptomatic | 3 b | 5 d | 11 ef | 1255 d | |||
| R5 | Clean | 0 b | 18 cd | 29 def | 3565 cd | ||
| Positive | 16 b | 46 bc | 54 bcd | 8775 bc | |||
| Symptomatic | 73 a | 77 ab | 81 abc | 16,250 a | |||
| R11 | Clean | 2 b | 68 ab | 94 ab | 12,891 ab | ||
| Positive | 50 a | 100 a | 100 a | 18,749 a | |||
| Symptomatic | 65 a | 96 a | 98 a | 18,841 a | |||
| KM98-1 | Positive | 0 | 25 | 34 | 4699 | ||
| Symptomatic | 60 | 78 | 96 | 16,665 | |||
| HB80 | Clean | 3 | 31 | 59 | 6904 | ||
| TME3 | Clean | 5 | 65 | 69 | 11,195 | ||
| SC8 | Positive | 24 | 92 | 100 | 16,760 | ||
| Symptomatic | 100 | 100 | 100 | 21,000 | |||
| HSD | 27 | 38 | 40 | 6569 | |||
| 2020 | HB60 | Clean | 0 c | 14 ef | 57 bcde | 75 abc | 11,352 cde |
| Positive | 6 c | 29 def | 56 cde | 60 c | 11,327 cde | ||
| Symptomatic | 18 c | 61 abcde | 93 a | 97 a | 19,546 ab | ||
| KU50 | Clean | 0 c | 3 f | 29 e | 53 c | 6388 e | |
| Positive | 0 c | 16 ef | 39 de | 62 bc | 8787 de | ||
| Symptomatic | 77 a | 84 abc | 93 a | 100 a | 21,924 a | ||
| R5 | Clean | 0 c | 36 cdef | 62 bcd | 74 abc | 13,069 cd | |
| Positive | 0 c | 45 bcdef | 77 abc | 90 ab | 16,185 bc | ||
| Symptomatic | 48 b | 67 abcd | 85 ab | 90 ab | 19,063 ab | ||
| R11 | Clean | 0 c | 44 bcdef | 100 a | 100 a | 19,125 ab | |
| Positive | 6 c | 91 ab | 100 a | 100 a | 23,872 a | ||
| Symptomatic | 91 a | 100 a | 100 a | 100 a | 22,060 a | ||
| KM98-1 | Positive | 0 | 34 | 79 | 89 | 15,693 | |
| Symptomatic | 78 | 85 | 98 | 98 | 22,418 | ||
| HB80 | Clean | 0 | 55 | 95 | 98 | 19,108 | |
| TME3 | Clean | 8 | 58 | 92 | 100 | 19,250 | |
| SC8 | Symptomatic | 92 | 100 | 100 | 100 | 23,875 | |
| R72 | Positive | 2 | 31 | 62 | 70 | 12,616 | |
| HSD | 27 | 49 | 29 | 31 | 5441 | ||
| Varieties | Clean | Positive Selection | Clean | Positive Selection |
|---|---|---|---|---|
| 2019–2020 | 2020–2021 | |||
| KU50 | 6.7 ± 0.07 | 0 | 6.7 ± 0.07 | 0 |
| R11 | 12.5 ± 0.09 | 66.7 ± 0.13 | 20 ± 0.20 | 26.7 ± 0.12 |
| HB60 | 26.7 ± 0.12 | 23.5 ± 0.11 | 0 | 18.8 ± 0.10 |
| R5 | 46.7 ± 0.13 | 35.3 ± 0.12 | 6.7 ± 0.07 | 0 |
| SC8 | NA | 42.9 ± 0.14 | NA | NA |
| TME3 * | 30.8 ± 0.13 | NA | 15.4 ± 0.10 | NA |
| KM98-1 | NA | 31.3 ± 0.12 | NA | 18.8 ± 0.10 |
| HB80 ** | 31.3 ± 0.12 | NA | 26.7 ± 0.12 | NA |
| R72 | - | - | NA | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malik, A.I.; Sophearith, S.; Delaquis, E.; Cuellar, W.J.; Jimenez, J.; Newby, J.C. Susceptibility of Cassava Varieties to Disease Caused by Sri Lankan Cassava Mosaic Virus and Impacts on Yield by Use of Asymptomatic and Virus-Free Planting Material. Agronomy 2022, 12, 1658. https://doi.org/10.3390/agronomy12071658
Malik AI, Sophearith S, Delaquis E, Cuellar WJ, Jimenez J, Newby JC. Susceptibility of Cassava Varieties to Disease Caused by Sri Lankan Cassava Mosaic Virus and Impacts on Yield by Use of Asymptomatic and Virus-Free Planting Material. Agronomy. 2022; 12(7):1658. https://doi.org/10.3390/agronomy12071658
Chicago/Turabian StyleMalik, Al Imran, Sok Sophearith, Erik Delaquis, Wilmer J. Cuellar, Jenyfer Jimenez, and Jonathan C. Newby. 2022. "Susceptibility of Cassava Varieties to Disease Caused by Sri Lankan Cassava Mosaic Virus and Impacts on Yield by Use of Asymptomatic and Virus-Free Planting Material" Agronomy 12, no. 7: 1658. https://doi.org/10.3390/agronomy12071658
APA StyleMalik, A. I., Sophearith, S., Delaquis, E., Cuellar, W. J., Jimenez, J., & Newby, J. C. (2022). Susceptibility of Cassava Varieties to Disease Caused by Sri Lankan Cassava Mosaic Virus and Impacts on Yield by Use of Asymptomatic and Virus-Free Planting Material. Agronomy, 12(7), 1658. https://doi.org/10.3390/agronomy12071658