
Microbial Metabolomics Interaction and Ecological Challenges of Trichoderma Species as Biocontrol Inoculant in Crop Rhizosphere

 , ,
, ,  ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Trichoderma Species as a Commercial Biofungicide
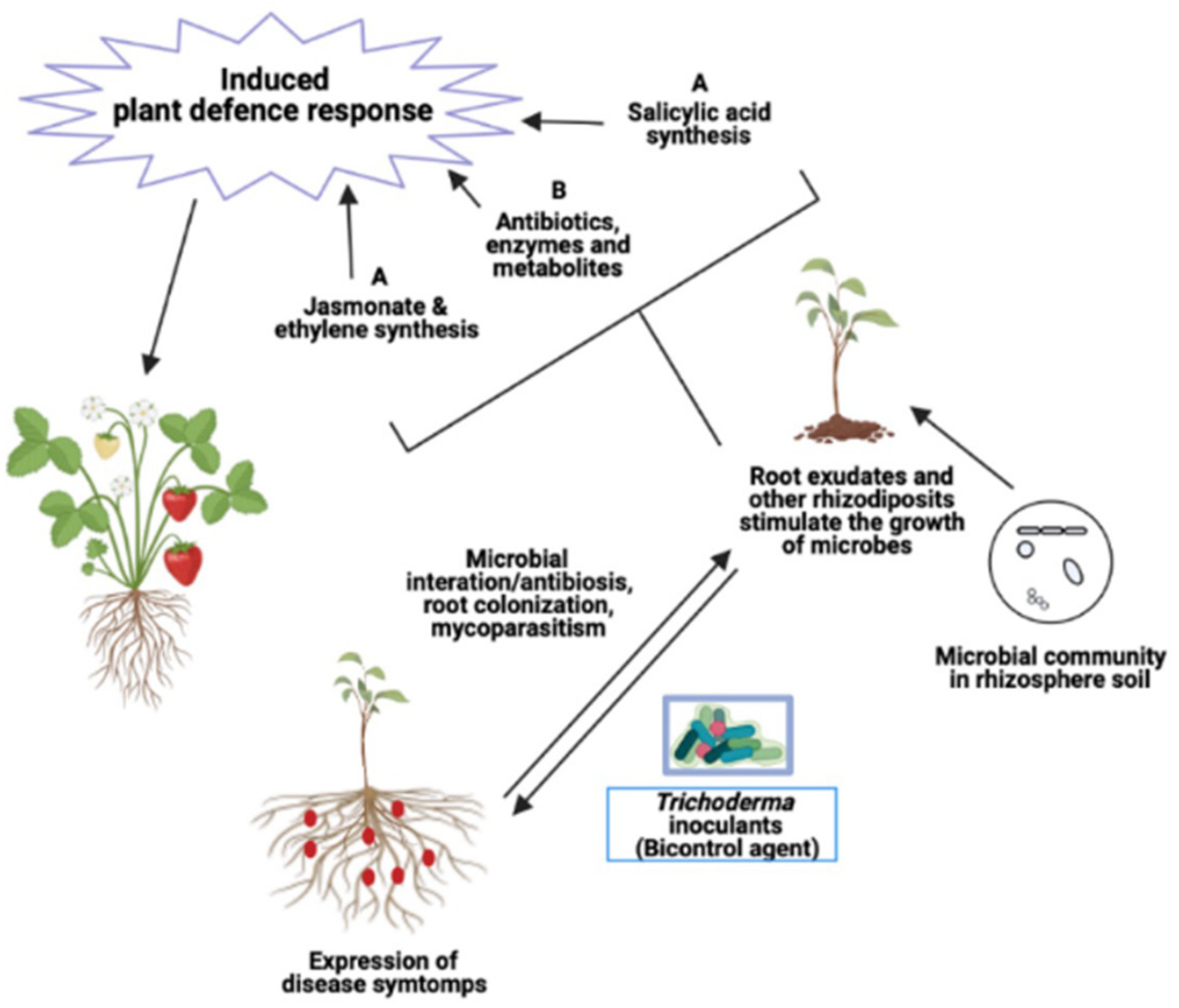
3. Effects of Trichoderma Metabolites on Plant Root Exudates
4. Effects of Trichoderma Metabolites on Soil and Root Pathogens

{kind=link}
{kind=link}
| Metabolites | Compound | Trichoderma Species | Target Fungal Pathogens | References |
|---|---|---|---|---|
| Anthraquinones | 1,8-dihydroxy-3-methylanthraquinone, 1-hydroxy-3-methylanthraquinone and 6-methyl-1,3,8-trihydroxyanthraquinone | T. harzianum | Fusarium oxysporum, Macrophomina phaseolina, Rizoctonia solani, and Sclerotium rolfsii | [55] |
| 1,8-dihydroxy-3-methylanthraquinone and 1-hydroxy-3-methylanthraquinone | T. harzianum | Gaeumannomyces graminis var. tritici, and Pythium ultimum | [56] | |
| Azaphilones | Harziphilone, Fleephilone and T22azaphilone | T. harzianum | G. graminis var. tritici, P. ultimum, and R. solani | [57] |
| T22azaphilone | T. harzianum | Leptosphaeria maculans, and Phytophthora cinnamomi | [58] | |
| Epipolythiodio-xopiperazines | Gliotoxins | T. virens | M. phaseolina, Pythium aphanidermatum, Pythium deharyanum, Rizoctonia bataticola, R. solani, and S. rolfsii | [1] |
| Gliovirin | T. longibrachiatum | R. solani | [1] | |
| Gliovirin | T. virens | P. ultimum | [1] | |
| Koninginins | koninginins A, B, D, E, and G | T. aureoviride T. harzianum and T. koningii | G. graminis var. tritici | [59] |
| koninginins A, B, and D | T. koningiopsis | F. oxysporum, F. solani, and S. rolfsii | [59] | |
| koninginin D | T. harzianum and T. koningii | Bipolaris sorokiniana, F. oxysporum, P. cinnamomi, and Pythium middletonii | [6] | |
| Lactones | aspinolide C | T. arundinaceum | Fusarium sporotrichioides | [60] |
| Cerinolactone | T. cerinum | Rosellinia necatrix | [60] | |
| Cremenolide | T. cremeum. | F. oxysporum and R. solani | [60] | |
| Peptaibols | trichokonins VI, VII, and VIII | T. koningii | F. oxysporum, R. solani, and Verticillium dahliae | [55] |
| Trichokonin VI | T. pseudokoningii | F. oxysporum, Phytophthora parasitica, and V. dahlia | [55] | |
| Polyketides | Harzianolide and Dehydro Harzianolide | T. harzianum | F. oxysporum and R. solani | [61] |
| 6-pentyl-α-pyrone | T. harzianum T. koningii T. viride and Trichoderma spp. | R. solani | [61] | |
| 6-pent-1-enyl-α-pyrone | T. harzianum and T. viride | R. solani | [61] | |
| Massoilactone δ-decenolactone | Trichoderma spp. | R. solani and S. rolfsii | [61] | |
| Koninginia E, B, and A | T. harzianum and T. koningii | G. graminis var. tritici | [57] | |
| Koninginin D and Seco-koninginin | T. harzianum | G. graminis var. tritici | [57] | |
| Koninginin C | T. koningii | G. graminis var. tritici | [57] | |
| Pyridones | Harzianopyridone | T. harzianum | G. graminis var. tritici, L. maculans, P. cinnamomi, P. ultimum, and R. solani | [57] |
| Pyrones | 6-Pentyl pyrone (6-PP) | T. harzianum, T. koningii, and T. viride | F. oxysporum and R. solani | [62] |
| Viridepyronone | T. viride | S. rolfsii | [62] | |
| Steroids | Stigmasterol | T. harzianum and T. koningii | F. oxysporum, M. phaseolina, R. solani, and S. rolfsii, | [57] |
| Terpenes (trichothecenes) | Trichodermin | T. polysporum T. sporulosum T. reesei and T. virens | R. solani | [63] |
| Harzianum A | T. harzianum | F. oxysporum | [63] | |
| Mycotoxin T2 | T. lignorum | R. solani and S. rolfsii | [63] | |
| Terpenes (triterpenes) (sterols) | Ergokonin A | T. koningii T. longibrachiatum and T. viride | Phoma spp. | [64] |
| Viridin | T. koningii T. virens and T. viride | F. oxysporum and R. solani | [64] | |
| Trichothecenes | Trichodermin | T. brevicompactum | R. solani | [65] |
| Trichodermin | T. harzianum | F. oxysporum and R. solani | [66] |
5. Effects of Trichoderma Metabolites on Soil Non-Pathogenic Fungal Communities
6. Effects of Trichoderma Metabolites on Soil Bacterial Communities
7. Effects of Trichoderma Metabolites on Soil Enzyme Activities
| Metabolites (Enzymes) | Trichoderma Strains | Bacteria | Fungi | Host Plant | References |
|---|---|---|---|---|---|
| 1,8-dihydroxy-3-methylanthraquinone | Trichoderma hamatum TR1-4 | Bacillus spp. | Gaeumannaomyces graminis var. tritici, and Rhizoctonia solani | Wheat and Eggplant | [92,93] |
| 1-hydroxy-3-methylanthraquinone | T. harzianum 2413 | Pseudomonas spp. | Phytophthora capsici | Pepper | [94] |
| 1,8-dihydroxy-3-methylanthraquinone | T. harzianum T-22 | Pseudomonas fluorescens Q8r1-96 | Gaeumannaomyces graminis var. tritici, and Pyrenophora triticis-repentis | Wheat | [95,96] |
| Harzianopyridone | T. harzianum T-1 | Pseudomonas aureofaciens AB244 | Fusarium oxysporum; Pythium ultimum, and R. solani | Bean and Tomato | [97,98] |
| 6-pentyl-α-pyrone | T. harzianum 1295-22 | Pseudomonas fluorescens VO61 | R. solani | Creeping bent grass and rice | [99,100] |
| Trichorzianin TA Trichorzianin TB | T. harzianum Th-87 | Stenotrophomonas maltophilia C3 | R. solani | Eggplant and Tall fescue | [92,101] |
| Trichodermin | T. harzianum BAFC 742 | Bacillus subtilis GB03 | Sclerotinia sclerotiorum | Soybean | [102] |
| β-1,4 endoglucanase | T. longibrachiatum CECT 2606 | Serratia plymuthica | P. ultimum | Cucumber | [101,103] |
| 6-pent-1-enyl-α-pyrone | T. viride WT-6 | Bacillus subtilis GB03 | R. solani | Eggplant | [96] |
| 3,4-dihydroxycarotane | T. virens GL-21 | Pseudomonas fluorescens VO61 | P. ultimum and R. solani | Cucumber and pea | [104] |
| 6-Pentyl pyrone (6-PP) | T. virens GL-1, GL-21, GL-23 | Bacillus subtilis BACT-D | R. solani | Eggplant | [92] |
| β-1,4 glucanase 3,4-dihydroxycarotane Viridin | T. virens GL-3 | Burkholderia cepacia A3R, B. cepacia PHQM 100, Pseudomonas aureofaciens 63-28, and P. aureofaciens AB244 | Fusarium graminearum, Pythium aphanidermatum, and P. ultimum | Barley, Cucumber, Maize, and wheat | [105,106,107,108,109] |
8. Challenges and Future Research Directions
8.1. Efficacy of Trichoderma–Based Bioformulations
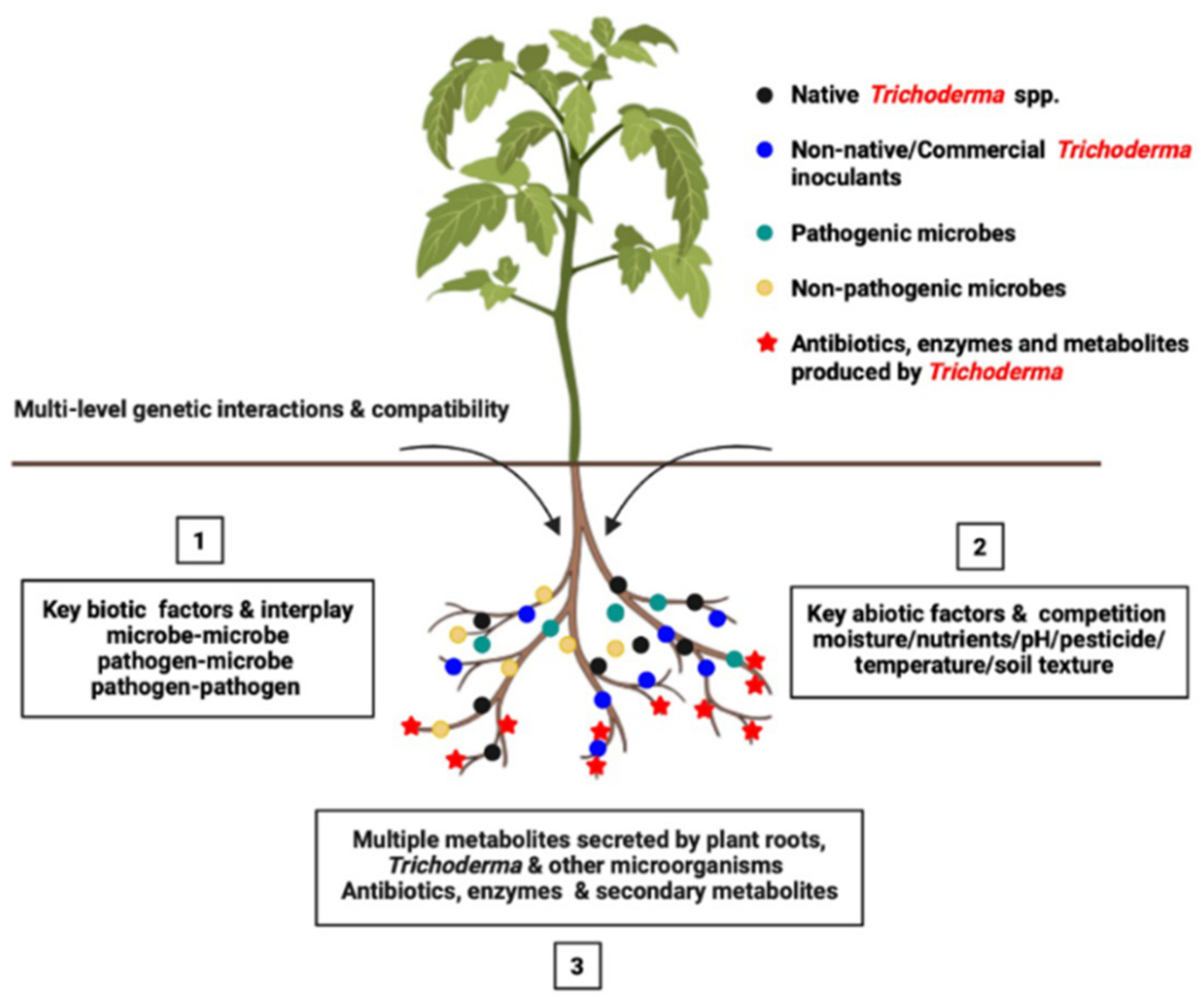
8.2. Ecological Challenges of Trichoderma Inoculant in Crop Rhizosphere
8.2.1. Survival Fitness of Microbial Communities in Crop Rhizosphere
8.2.2. Trichoderma Affects Chemical Signals in Crop Rhizosphere
8.2.3. Trichoderma Challenges Abiotic and Biotic Factors in Crop Rhizosphere
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ghorbanpour, M.; Omidvari, M.; Abbaszadeh-Dahaji, P.; Omidvar, R.; Kariman, K. Mechanisms underlying the protective effects of beneficial fungi against plant diseases. Biol. Control. 2018, 117, 147–157. [Google Scholar] [CrossRef]
- Harman, G.; Khadka, R.; Doni, F.; Uphoff, N. Benefits to Plant Health and Productivity From Enhancing Plant Microbial Symbionts. Front. Plant Sci. 2021, 11, 2001. [Google Scholar] [CrossRef] [PubMed]
- Rajesh, R.W.; Rahul, M.S.; Ambalal, N.S. Trichoderma: A significant fungus for agriculture and environment. Afric. J. Agric. Res. 2016, 11, 1952–1965. [Google Scholar] [CrossRef]
- Jin-lian, C.; Kai, L.; Cui-ping, M.; Hui-lin, G.; Li-xing, Z.; Shi-zhong, S. Chemical Constituents with Siderophores Activities from Trichoderma koningiopsis YIM PH30002. Nat. Prod. Res. Develop. 2015, 27, 1878–1883. [Google Scholar]
- Asis, A.; Shahriar, S.A.; Naher, L.; Saallah, S.; Nur Fatihah, H.N.; Kumar, V.; Siddiquee, S. Species pattern identification of Trichoderma strains using morphological characteristics, phylogenetic analyses and lignocellulolytic activities. Mol. Biol. Rep. 2021, 48, 3285–3301. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Horwitz, B.A.; Kenerley, C.M. Secondary metabolism in Trichoderma—A genomic perspective. Microbiology 2012, 158, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Guzmán, P.; Porras-Troncoso, M.D.; Olmedo-Monfil, V.; Herrera-Estrella, A. Trichoderma species: Versatile plant symbionts. Phytopathology 2019, 109, 6–16. [Google Scholar] [CrossRef]
- Jangir, M.; Sharma, S.; Sharma, S. Non-target effects of Trichoderma on plants and soil microbial communities. In Plant Microbe Interface; Springer: Cham, Switzerland, 2019; pp. 239–251. [Google Scholar]
- Rout, M.E.; Southworth, D. The root microbiome influences scales from molecules to ecosystems: The unseen majority. Am. J. Bot. 2013, 100, 1689–1691. [Google Scholar] [CrossRef]
- Sani, M.N.H.; Yong, J.W.H. Harnessing Synergistic Biostimulatory Processes: A Plausible Approach for Enhanced Crop Growth and Resilience in Organic Farming. Biology 2022, 11, 41. [Google Scholar] [CrossRef]
- Craney, A.; Ahmed, S.; Nodwell, J. Towards a new science of secondary metabolism. J. Antibiot. 2013, 66, 387–400. [Google Scholar] [CrossRef]
- Choudhary, D.K.; Sharma, K.P.; Gaur, R.K. Biotechnological perspectives of microbes in agro ecosystems. Biotechnol. lett. 2011, 33, 1905–1910. [Google Scholar] [CrossRef] [PubMed]
- Sani, M.N.H.; Hasan, M.; Uddain, J.; Subramaniam, S. Impact of application of Trichoderma and biochar on growth, productivity and nutritional quality of tomato under reduced N-P-K fertilization. Ann. Agric. Sci. 2020, 65, 107–115. [Google Scholar] [CrossRef]
- Sani, M.N.H.; Islam, M.N.; Uddain, J.; Chowdhury, M.S.N.; Subramaniam, S. Synergistic effect of microbial and nonmicrobial biostimulants on growth, yield, and nutritional quality of organic tomato. Crop Sci. 2020, 60, 2102–2114. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.; Bardgett, R.D.; Van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Naseby, D.C.; Pascual, J.A.; Lynch, J.M. Effect of biocontrol strains of Trichoderma on plant growth, Pythium ultimum populations, soil microbial communities and soil enzyme activities. J. Appl. Microbiol. 2000, 88, 161–169. [Google Scholar] [CrossRef]
- Brimner, T.A.; Boland, G.J. A review of the non-target effects of fungi used to biologically control plant diseases. Agric. Ecosyst. Environ. 2003, 100, 3–16. [Google Scholar] [CrossRef]
- Islam, M.N.; Germida, J.J.; Walley, F.L. Survival of a commercial AM fungal inoculant and its impact on indigenous AM fungal communities in field soils. Appl. Soil Ecol. 2021, 166, 103979. [Google Scholar] [CrossRef]
- McAllister, C.B.; Garcia-Romera, I.; Godeas, A.; Ocampo, J.A. In vitro interactions between Trichoderma koningii, Fusarium solani and Glomus mosseae. Soil Biol. Biochem. 1994, 26, 1369–1374. [Google Scholar] [CrossRef]
- Dubey, A.; Malla, M.A.; Khan, F.; Chowdhary, K.; Yadav, S.; Kumar, A.; Sharma, S.; Khare, P.K.; Khan, M.L. Soil microbiome: A key player for conservation of soil health under changing climate. Biodivers Conserv. 2019, 28, 2405–2429. [Google Scholar] [CrossRef]
- Schlatter, D.; Kinkel, L.; Thomashow, L.; Weller, D.; Paulitz, T. Disease suppressive soils: New insights from the soil microbiome. Phytopathology 2017, 107, 1284–1297. [Google Scholar] [CrossRef]
- Canarini, A.; Kaiser, C.; Merchant, A.; Richter, A.; Wanek, W. Root Exudation of Primary Metabolites: Mechanisms and Their Roles in Plant Responses to Environmental Stimuli. Front. Plant Sci. 2019, 10, 157. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, R.; Peukert, M.; Succurro, A.; Koprivova, A.; Kopriva, S. The Role of Soil Microorganisms in Plant Mineral Nutrition—Current Knowledge and Future Directions. Front. Plant Sci. 2017, 8, 1617. [Google Scholar] [CrossRef] [PubMed]
- Gougoulias, C.; Clark, J.M.; Shaw, L.J. The role of soil microbes in the global carbon cycle: Tracking the below-ground microbial processing of plant-derived carbon for manipulating carbon dynamics in agricultural systems. J. Sci. Food Agric. 2014, 94, 2362–2371. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Meng, H.; Gu, J.-D. Microbial extracellular enzymes in biogeochemical cycling of ecosystems. J. Environ. Manag. 2017, 197, 539–549. [Google Scholar] [CrossRef]
- Yang, T.; Lupway, N.; Marc, S.-A.; Siddique, K.H.M.; Bainard, L.D. Anthropogenic drivers of soil microbial communities and impacts on soil biological functions in agroecosystems. Global Ecol. Conserv. 2021, 27, e01521. [Google Scholar] [CrossRef]
- Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Jaroszuk-Ściseł, J. Trichoderma: The Current Status of Its Application in Agriculture for the Biocontrol of Fungal Phytopathogens and Stimulation of Plant Growth. Int. J. Mol. Sci. 2022, 23, 2329. [Google Scholar] [CrossRef]
- Mukhopadhyay, R.; Kumar, D. Trichoderma: A beneficial antifungal agent and insights into its mechanism of biocontrol potential. Egypt. J. Biol. Pest Control 2020, 30, 133. [Google Scholar] [CrossRef]
- Chaverri, P.; Samuels, G.J. Evolution of habitat preference and nutrition mode in a cosmopolitan fungal genus with evidence of interkingdom host jumps and major shifts in ecology. Evolution 2013, 67, 2823–2837. [Google Scholar] [CrossRef]
- Chaverri, P.; Branco-Rocha, F.; Jaklitsch, W.; Gazis, R.; Degenkolb, T.; Samuels, G.J. Systematics of the Trichoderma harzianum species complex and the re-identification of commercial biocontrol strains. Mycologia 2015, 107, 558–590. [Google Scholar] [CrossRef]
- Sudantha, I.M.; Sudirman; Ernawati, N.M.L. The effect of method and dosage application of biofungicide extract of Legundi leaf fermented with Trichoderma harzianum fungus for control of Fusarium wilt disease on shallots. IOP Conf. Ser. Earth Environ. Sci. 2021, 913, 012014. [Google Scholar] [CrossRef]
- Bailey, B.A.; Bae, H.; Strem, M.D.; Crozier, J.; Thomas, S.E.; Samuels, G.J.; Vinyard, B.T.; Holmes, K.A. Antibiosis, mycoparasitism, and colonization success for endophytic Trichoderma isolates with biological control potential in Theobroma cacao. Biol. Control 2008, 46, 24–35. [Google Scholar] [CrossRef]
- Adetunji, C.O.; Anani, O.A. Bio-fertilizer from Trichoderma: Boom for Agriculture Production and Management of Soil- and Root-Borne Plant Pathogens. In Innovations in Food Technology; Springer: Singapore, 2020; pp. 245–256. [Google Scholar] [CrossRef]
- Fraceto, L.F.; Maruyama, C.R.; Guilger, M.; Mishra, S.; Keswani, C.; Singh, H.B.; de Lima, R. Trichoderma harzianum-based novel formulations: Potential applications for management of Next-Gen agricultural challenges. J. Chem. Technol. Biotechnol. 2018, 93, 2056–2063. [Google Scholar] [CrossRef]
- Woo, S.L.; Ruocco, M.; Vinale, F.; Nigro, M.; Marra, R.; Lombardi, N.; Pascale, A.; Lanzuise, S.; Manganiello, G.; Lorito, M. Trichoderma-based Products and their Widespread Use in Agriculture. Open Mycol. J. 2014, 8, 71–126. [Google Scholar] [CrossRef]
- Srivastava, M.; Kumar, V.; Shahid, M.; Pandey, S.; Singh, A. Trichoderma—A potential and effective biofungicide and alternative source against notable phytopathogens: A review. Afr. J. Agric. Res. 2016, 11, 310–316. [Google Scholar] [CrossRef]
- Guerrieri, A.; Dong, L.; Bouwmeester, H.J. Role and exploitation of underground chemical signaling in plants. Pest Manag. Sci. 2019, 75, 2455–2463. [Google Scholar] [CrossRef] [PubMed]
- Vargas, W.A.; Crutcher, F.K.; Kenerley, C.M. Functional characterization of a plant-like sucrose transporter from the beneficial fungus Trichoderma virens. Regulation of the symbiotic association with plants by sucrose metabolism inside the fungal cells. New Phytol. 2011, 189, 777–789. [Google Scholar] [CrossRef] [PubMed]
- Samolski, I.; Rincón, A.M.; Pinzón, L.M.; Viterbo, A.; Monte, E. The qid74 gene from Trichoderma harzianum has a role in root architecture and plant biofertilization. Microbiology 2012, 158, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; del-Val, E.; Larsen, J. Ecological functions of Trichoderma spp. and their secondary metabolites in the rhizosphere: Interactions with plants. FEMS Microbiol. Ecol. 2016, 92, fiw036. [Google Scholar] [CrossRef]
- Chen, L.; Yang, X.; Raza, W.; Li, J.; Liu, Y.; Qiu, M.; Zhang, F.; Shen, Q. Trichoderma harzianum SQR-T037 rapidly degrades allelochemicals in rhizospheres of continuously cropped cucumbers. Appl. Microbiol. Biotechnol. 2011, 89, 1653–1663. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Roldán, A.; Albacete, A.; Pascual, J.A. The interaction with arbuscular mycorrhizal fungi or Trichoderma harzianum alters the shoot hormonal profile in melon plants. Phytochemistry 2011, 72, 223–229. [Google Scholar] [CrossRef]
- Chakraborty, B.N.; Chakraborty, U.; Sunar, K. Induced Immunity Developed by Trichoderma Species in Plants. In Trichoderma. Rhizosphere Biology; Sharma, A., Sharma, P., Eds.; Springer: Singapore, 2020; pp. 125–147. [Google Scholar] [CrossRef]
- Srivastava, M.; Shahid, M.; Pandey, S.; Kumar, V.; Singh, A.; Trivedi, S.; Srivastava, Y.K.; Shivram. Trichoderma: A scientific approach against soil borne pathogens. Afr. J. Microbiol. Res. 2015, 9, 2377–2384. [Google Scholar] [CrossRef]
- Błaszczyk, L.; Siwulski, M.; Sobieralski, K.; Lisiecka, J.; Jędryczka, M. Trichoderma spp.-application and prospects for use in organic farming and industry. J. Plant Prot. Res. 2014, 54, 309–317. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Steindorff, A.S.; Chenthamara, K.; Manganiello, G.; Henrissat, B.; Zhang, J.; Cai, F.; Kopchinskiy, A.G.; Kubicek, E.M.; Kuo, A.; et al. Evolution and comparative genomics of the most common Trichoderma species. BMC Genom. 2019, 20, 485. [Google Scholar] [CrossRef] [PubMed]
- Schuster, A.; Schmoll, M. Biology and biotechnology of Trichoderma. Appl. Microbiol. Biotechnol. 2010, 87, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.E.; Howell, C.R.; Viterbo, A. Trichoderma species opportunistic, a virulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Yedidia, I.; Shoresh, M.; Kerem, Z.; Benhamou, N.; Kapulnik, Y.; Chet, I. Concomitant induction of systemic resistance to Pseudomonas syringae pv. lachrymans in cucumber by Trichoderma asperellum (T-203) and accumulation of phytoalexins. Appl. Environ. Microbiol. 2003, 69, 7343–7353. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Valdespino, C.A.; Casas-Flores, S.; Olmedo-Monfil, V. Trichoderma as a model to study effector-like molecules. Front. Microbiol. 2019, 10, 1030. [Google Scholar] [CrossRef] [PubMed]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “secrets” of a multitalented biocontrol agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Kumar, M.; Brar, A.; Yadav, M.; Chawade, A.; Vivekanand, V.; Pareek, N. Chitinases—Potential candidates for enhanced plant resistance towards fungal pathogens. Agriculture 2018, 8, 88. [Google Scholar] [CrossRef]
- Esse, H.P.; Reuber, T.L.; Does, D. Genetic modification to improve disease resistance in crops. New Phytol. 2020, 225, 70–86. [Google Scholar] [CrossRef]
- Alfiky, A.; Weisskopf, L. Deciphering Trichoderma–plant–pathogen interactions for better development of biocontrol applications. J. Fungi. 2021, 7, 61. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.A.A.; Najeeb, S.; Hussain, S.; Xie, B.; Li, Y. Bioactive secondary metabolites from Trichoderma spp. against phytopathogenic fungi. Microorganisms 2020, 8, 817. [Google Scholar] [CrossRef] [PubMed]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Woo, S.L.; Nigro, M.; Marra, R.; Lombardi, N.; Pascale, A.; Ruocco, M.; Lanzuise, S.; et al. Trichoderma Secondary Metabolites Active on Plants and Fungal Pathogens. Open Mycol. J. 2014, 8, 127–139. [Google Scholar] [CrossRef]
- Keswani, C.; Singh, H.B.; Hermosa, R.; García-Estrada, C.; Caradus, J.; He, Y.W.; Mezaache-Aichour, S.; Glare, T.R.; Borriss, R.; Vinale, F.; et al. Antimicrobial secondary metabolites from agriculturally important fungi as next biocontrol agents. Appl. Microbiol. Biotechnol. 2019, 103, 9287–9303. [Google Scholar] [CrossRef] [PubMed]
- Daguerre, Y.; Siegel, K.; Edel-Hermann, V.; Steinberg, C. Fungal proteins and genes associated with biocontrol mechanisms of soil-borne pathogens: A review. Fungal. Biol. Rev. 2014, 28, 97–125. [Google Scholar] [CrossRef]
- Zeilinger, S.; Gruber, S.; Bansal, R.; Mukherjee, P.K. Secondary metabolism in Trichoderma—Chemistry meets genomics. Fungal. Biol. Rev. 2016, 30, 74–90. [Google Scholar] [CrossRef]
- Vargas, W.A.; Mukherjee, P.K.; Laughlin, D.; Wiest, A.; Moran-Diez, M.E.; Kenerley, C.M. Role of gliotoxin in the symbiotic and pathogenic interactions of Trichoderma virens. Microbiology 2014, 160, 2319–2330. [Google Scholar] [CrossRef]
- Shi, M.; Chen, L.; Wang, X.W.; Zhang, T.; Zhao, P.B.; Song, X.Y.; Sun, C.Y.; Chen, X.L.; Zhou, B.C.; Zhang, Y.Z. Antimicrobial peptaibols from Trichoderma pseudokoningii induce programmed cell death in plant fungal pathogens. Microbiology 2012, 158, 166–175. [Google Scholar] [CrossRef]
- Degenkolb, T.; Fog Nielsen, K.; Dieckmann, R.; Branco-Rocha, F.; Chaverri, P.; Samuels, G.J.; Thrane, U.; von Döhren, H.; Vilcinskas, A.; Brückner, H. Peptaibol, Secondary-Metabolite, and Hydrophobin Pattern of Commercial Biocontrol Agents Formulated with Species of the Trichoderma harzianum Complex. Chem. Biodivers. 2015, 12, 662–684. [Google Scholar] [CrossRef]
- Brakhage, A.A.; Schroeckh, V. Fungal secondary metabolites–Strategies to activate silent gene clusters. Fungal. Genet. Biol. 2011, 48, 15–22. [Google Scholar] [CrossRef]
- Netzker, T.; Fischer, J.; Weber, J.; Mattern, D.J.; König, C.C.; Valiante, V.; Schroeckh, V.; Brakhage, A.A. Microbial communication leading to the activation of silent fungal secondary metabolite gene clusters. Front. Microbiol. 2015, 6, 299. [Google Scholar] [CrossRef] [PubMed]
- Stoppacher, N.; Kluger, B.; Zeilinger, S.; Krska, R.; Schuhmacher, R. Identification and profiling of volatile metabolites of the biocontrol fungus Trichoderma atroviride by HS-SPME-GC-MS. J. Microbiol Methods 2010, 81, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Rokas, A.; Wisecaver, J.H.; Lind, A.L. The birth, evolution and death of metabolic gene clusters in fungi. Nat. Rev. Microbiol. 2018, 16, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Bell, T.; Newman, J.A.; Silverman, B.W.; Turner, S.L.; Lilley, A.K. The contribution of species richness and composition to bacterial services. Nature 2005, 436, 1157–1160. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, B.J.; Srivastava, D.S.; Duffy, J.E.; Wright, J.P.; Downing, A.L.; Sankaran, M.; Jouseau, C. Effects of biodiversity on the functioning of trophic groups and ecosystems. Nature 2006, 443, 989–992. [Google Scholar] [CrossRef]
- Assigbetsé, K.; Ciss, I.; Bakhoum, N.; Dieng, L. Effect of inoculation of Acacia senegal mature trees with mycorrhiza and rhizobia on soil properties and microbial community structure. In Proceedings of the EGU General Assembly Conference Abstracts, Vienna, Austria, 22–27 April 2012; pp. 2012–8004. [Google Scholar]
- Zhang, F.G. The Affects and Mechanisms of Puta Five Trichoredma harzianum Mutant and ITS Bio-Organic Fertilizer on Growth of Cucumber; Nanjing Agricultural University: Nanjing, China, 2015; pp. 15–18. [Google Scholar]
- Degenkolb, T.; Aghcheh, K.R.; Dieckmann, R.; Neuhof, T.; Baker, S.E.; Druzhinina, I.S.; Kubicek, C.P.; Brückner, H.; Von Döhren, H. The production of multiple small peptaibol families by single 14-module peptide synthetases in Trichoderma/hypocrea. Chem. Biodivers 2012, 9, 499–535. [Google Scholar] [CrossRef]
- Abegaz, B.M.; Kinfe, H.H. Secondary metabolites, their structural diversity, bioactivity, and ecological functions: An overview. Physic. Sci. Rev. 2019, 4, 4. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma-plant-pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Baazeem, A.; Almanea, A.; Manikandan, P.; Alorabi, M.; Vijayaraghavan, P.; Abdel-Hadi, A. In Vitro Antibacterial, Antifungal, Nematocidal and Growth Promoting Activities of Trichoderma hamatum FB10 and Its Secondary Metabolites. J. Fungi 2021, 7, 331. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; del-Val, E.; Larsen, J. Interactions of Trichoderma with Plants, Insects, and Plant Pathogen Microorganisms: Chemical and Molecular Bases. In Co-Evolution of Secondary Metabolites; Mérillon, J.M., Ramawat, K., Eds.; Springer: Cham, Switzerland, 2020; pp. 263–290. [Google Scholar] [CrossRef]
- Zin, N.A.; Badaluddin, N.A. Biological functions of Trichoderma spp. for agriculture applications. Ann. Agric. Sci. 2020, 65, 168–178. [Google Scholar] [CrossRef]
- Vázquez, M.M.; César, S.; Azcón, R.; Barea, J.M. Interactions between arbuscular mycorrhizal fungi and other microbial inoculants (Azospirillum, Pseudomonas, Trichoderma) and their effects on microbial population and enzyme activities in the rhizosphere of maize plants. Appl. Soil Ecol. 2000, 15, 261–272. [Google Scholar] [CrossRef]
- Umadevi, P.; Anandaraj, M.; Srivastav, V.; Benjamin, S. Trichoderma harzianum MTCC 5179 impacts the population and functional dynamics of microbial community in the rhizosphere of black pepper (Piper nigrum L.). Braz. J. Microbiol. 2018, 49, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Halifu, S.; Deng, X.; Song, X.; Song, R. Effects of Two Trichoderma Strains on Plant Growth, Rhizosphere Soil Nutrients, and Fungal Community of Pinus sylvestris var. mongolica Annual Seedlings. Forests 2019, 10, 758. [Google Scholar] [CrossRef]
- Cordier, C.; Alabouvette, C. Effects of the introduction of a biocontrol strain of Trichoderma atroviride on non-target soil microorganisms. Eur. J. Soil Biol. 2009, 7, 267–274. [Google Scholar] [CrossRef]
- Li, Q.R.; Tan, P.; Jiang, Y.L.; Hyde, K.D.; Mckenzie, E.H.C.; Bahkali, A.H.; Kang, J.C.; Wang, Y. A novel Trichoderma species isolated from soil in Guizhou, T. guizhouense. Mycol. Prog. 2013, 12, 167–172. [Google Scholar] [CrossRef]
- Crutcher, F.K.; Parich, A.; Schuhmacher, R.; Mukherjee, P.K.; Zeilinger, S.; Kenerley, C.M. A putative terpene cyclase, vir4, is responsible for the biosynthesis of volatile terpene compounds in the biocontrol fungus Trichoderma virens. Fungal. Genet. Biol. 2013, 56, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Reino, J.L.; Guerrero, R.F.; Hernandez-Galan, R.; Collado, I.G. Secondary metabolites from species of the biocontrol agent Trichoderma. Phytochem. Rev. 2008, 7, 89–123. [Google Scholar] [CrossRef]
- Saravanakumar, K.; Li, Y.; Yu, C.; Wang, Q.Q.; Wang, M.; Sun, J.; Gao, J.X.; Chen, J. Effect of Trichoderma harzianum on maize rhizosphere microbiome and biocontrol of Fusarium stalk rot. Sci. Rep. 2017, 7, 1771–1783. [Google Scholar] [CrossRef]
- Gupta, R.; Mathimaran, N.; Wiemken, A.; Boller, T.; Bisaria, V.S.; Sharma, S. Non-target effects of bioinoculants on rhizospheric microbial communities of Cajanus cajan. Appl. Soil Ecol. 2014, 76, 26–33. [Google Scholar] [CrossRef]
- Szczepaniak, Z.; Cyplik, P.; Juzwa, W.; Czarny, J.; Staninska, J.; Piotrowska-Cyplik, A. Antibacterial effect of the Trichoderma viride fungi on soil microbiome during PAH’s biodegradation. Int. Biodeterior. Biodegrad. 2015, 104, 170–177. [Google Scholar] [CrossRef]
- Gasoni, L.; Khan, N.; Yokoyama, K.; Chiessa, G.H.; Kobayashi, K. Impact of Trichoderma harzianum biocontrol agent on functional diversity of soil microbial community in tobacco monoculture in Argentina. World J. Agric. Sci. 2008, 4, 527–532. [Google Scholar]
- Naseby, D.C.; Lynch, J.M. Impact of wild-type and genetically modified Pseudomonas fluorescens on soil enzyme activities and microbial population structure in the rhizosphere of pea. Mol. Ecol. 1998, 7, 617–625. [Google Scholar] [CrossRef]
- Mbarki, S.; Cerdà, A.; Brestic, M.; Mahendra, R.; Abdelly, C.; Pascual, J.A. Vineyard compost supplemented with Trichoderma harzianum T78 improve saline soil quality. Land Degrad. Develop. 2016, 28, 1028–1037. [Google Scholar] [CrossRef]
- Wagner, M.R.; Lundberg, D.S.; Del Rio, T.G.; Tringe, S.G.; Dangl, J.L.; Mitchell-Olds, T. Host genotype and age shape the leaf and root microbiomes of a wild perennial plant. Nat. Commun. 2016, 7, 12151. [Google Scholar] [CrossRef]
- Yadav, R.L.; Shukla, S.K.; Suman, A.; Singh, P.N. Trichoderma inoculation and trash management effects on soil microbial biomass, soil respiration, nutrient uptake and yield of ratoon sugarcane under subtropical conditions. Biol. Fertil. Soils 2009, 45, 461–468. [Google Scholar] [CrossRef]
- Lewis, J.A.; Larkin, R.P. Formulation of the biocontrol fungus Cladorrhinum foecundissimum to reduce damping-off diseases caused by Rhizoctonia solani and Pythium ultimim. Biol. Control. 1998, 12, 182–190. [Google Scholar] [CrossRef]
- Ryder, M.H.; Yan, Z.; Terrace, T.E.; Rovira, A.D.; Tong, W.; Correll, R.L. Use of strains of Bacillus isolated in China to suppress take-all and Rhizoctonia root rot and promote seedling growth of glasshouse-grown wheat in Australian soils. Soil. Biol. Biochem. 1999, 31, 19–29. [Google Scholar] [CrossRef]
- Ahmed, A.S.; Perez-Sanchez, C.; Egea, C.; Candela, M.E. Evaluation of Trichoderma harzianum for controlling root rot caused Phytophthora capsica in pepper plants. Plant. Pathol. 1999, 48, 58–65. [Google Scholar] [CrossRef]
- da Luz, W.C.; Bergstorm, G.C.; Stockwell, C.A. Seed-applied bioprotectants for control of seed-borne Pyrenophora tritici-repentis and agronomic enhancement of wheat. Can. J. Plant Pathol. 1998, 19, 384–386. [Google Scholar]
- Raaijmakers, J.M.; Weller, D.M. Natural plant protection by 2,4-diacetylphloroglucinol-producing Pseudomonas spp. in take-all decline soils. Mol. Plant-Microbe Interact 1998, 11, 144–152. [Google Scholar] [CrossRef]
- Warren, J.E.; Bennett, M.A. Bio-osmopriming tomato (Lycopersicon esculentum Mill.) seeds for improved stand establishment. Seed Sci. Technol. 1999, 27, 489–499. [Google Scholar]
- Woo, S.L.; Donzelli, B.; Scala, F.; Mach, R.; Harman, G.E.; Kubicek, C.P.; Del Sorbe, G.; Lorito, M. Disruption of the ech42 (endochitinase-encoding) gene affects biocontrol activity in in Trichoderma harzianum P1. Mol. Plant-Microbe Interact. 1999, 12, 419–429. [Google Scholar] [CrossRef]
- Lo, C.-T.; Nelson, E.B.; Hayes, C.K.; Harman, G.E. Ecological studies of transformed Trichoderma harzianum strain 1295-22 in the rhizosphere and on the phylloplane of creeping bentgrass. Phytopathology 1998, 88, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Vidhyasekaran, P.; Muthamilan, M. Evaluation of a powder formulation of Pseudomonas fluorescens Pf1 for control of rice sheath blight. Biocontrol. Sci. Technol. 1999, 9, 67–74. [Google Scholar] [CrossRef]
- Giesler, L.J.; Yuen, G.Y. Evaluation of Stenotrophomonas maltophilia strain C3 for biocontrol of brown patch disease. Crop Prot. 1998, 17, 509–513. [Google Scholar] [CrossRef]
- Menendez, A.B.; Godeas, A. Biological control of Sclerotinia sclerotiorum attacking soybean plants. Degradation of the cell walls of this pathogen by Trichoderma harzianum (BAFC 742). Mycopathologia 1998, 143, 153–160. [Google Scholar] [CrossRef]
- Migheli, Q.; Gonzalez-Candelas, L.; Dealessi, L.; Camponogara, A.; Ramon-Vidal, D. Transformants of Trichoderma longibrachiatum overexpressing the β-1,4-endoglucanase gene egl1 show enhanced biocontrol of Pythium ultimum on cucumber. Phytopathology 1998, 88, 673–677. [Google Scholar] [CrossRef]
- Koch, E. Evaluation of commercial products for microbial control of soil-borne plant diseases. Crop Prot. 1999, 18, 119–125. [Google Scholar] [CrossRef]
- Chen, C.; Belanger, R.R.; Benhamou, N.; Paulitz, T.C. Induced systemic resistance (ISR) by Pseudomonas spp. impairs pre- and post-infection development of Pythium aphanidermatum on cucumber roots. Eur. J. Plant Pathol. 1998, 104, 877–886. [Google Scholar] [CrossRef]
- Hebbar, K.P.; Martel, M.H.; Heulin, T. Suppression of pre- and post-emergence damping-off in corn by Burkholderia cepacia. Eur. J. Plant Pathol. 1998, 104, 29–36. [Google Scholar] [CrossRef]
- Huang, Y.; Wong, P.T.W. Effect of Burkholderia (Pseudomonas) cepacia and soil type on the control of crown rot in wheat. Plant Soil 1998, 203, 103–108. [Google Scholar] [CrossRef]
- Johnsson, L.; Hokeberg, M.; Gerhardson, B. Performance of the Pseudomonas chlororaphis biocontrol agent MA 342 against cereal seed-borne diseases in field experiments. Eur. J. Plant Pathol. 1998, 104, 701–711. [Google Scholar] [CrossRef]
- Mao, W.; Lewis, J.A.; Hebbar, P.K.; Lumsden, R.D. Seed treatment with a fungal or a bacterial antagonist for reducing corn damping-off caused by species of Pythium and Fusarium. Plant Dis. 1997, 81, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Rajput, V.D.; Minkina, T.; Feizi, M.; Kumari, A.; Khan, M.; Mandzhieva, S.; Sushkova, S.; El-Ramady, H.; Verma, K.K.; Singh, A.; et al. Effects of Silicon and Silicon-Based Nanoparticles on Rhizosphere Microbiome, Plant Stress and Growth. Biology 2021, 10, 791. [Google Scholar] [CrossRef]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of Action of Microbial Biological Control Agents against Plant Diseases: Relevance Beyond Efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef]
- Kour, D.; Kaur, T.; Devi, R.; Yadav, A.; Singh, M.; Joshi, D.; Singh, J.; Suyal, D.C.; Kumar, A.; Rajput, V.D.; et al. Beneficial microbiomes for bioremediation of diverse contaminated environments for environmental sustainability: Present status and future challenges. Environ. Sci. Pollut. Res. Int. 2021, 28, 24917–24939. [Google Scholar] [CrossRef]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef]
- Hashem, A.; Tabassum, B.; Abd_Allah, E.F. Bacillus subtilis: A plant-growth promoting rhizobacterium that also impacts biotic stress. Saudi J. Biol. Sci. 2019, 26, 1291–1297. [Google Scholar] [CrossRef]
- Berg, G.; Köberl, M.; Rybakova, D.; Müller, H.; Grosch, R.; Smalla, K. Plant microbial diversity is suggested as the key to future biocontrol and health trends. FEMS Microbiol. Ecol. 2017, 93, fix050. [Google Scholar] [CrossRef]
- Kaul, S.; Choudhary, M.; Gupta, S.; Dhar, M.K. Engineering Host Microbiome for Crop Improvement and Sustainable Agriculture. Front. Microbiol. 2021, 12, 635917. [Google Scholar] [CrossRef]
- Kumar, A.; Dubey, A. Rhizosphere microbiome: Engineering bacterial competitiveness for enhancing crop production. J. Adv. Res. 2020, 24, 337–352. [Google Scholar] [CrossRef] [PubMed]
- Whipps, J.M. Microbial interactions and biocontrol in the rhizosphere. J. Expt. Bot. 2001, 52, 487–511. [Google Scholar] [CrossRef]
- Chen, C.; Wang, M.; Zhu, J.; Tang, Y.; Zhang, H.; Zhao, Q.; Jing, M.; Chen, Y.; Xu, X.; Jiang, J.; et al. Long-term effect of epigenetic modification in plant–microbe interactions: Modification of DNA methylation induced by plant growth-promoting bacteria mediates promotion process. Microbiome 2022, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Xu, X.; Huo, Y.; Xiao, Y. Trichoderma–Inoculation and Mowing Synergistically Altered Soil Available Nutrients, Rhizosphere Chemical Compounds and Soil Microbial Community, Potentially Driving Alfalfa Growth. Front. Microbiol. 2019, 7, 3241. [Google Scholar] [CrossRef]


| Trichoderma–Based Bioformulation/Tradename | Trichoderma Species | Target Pathogens | Manufacturer | References |
|---|---|---|---|---|
| Agroguard WG™ | T. harzianum | Phoma, Pythium, Rhizoctonia, Sclerotinia, and Sclerotium | Life Systems Technology S.A. (Colombia) | [34] |
| Antagon WP™ | T. harzianum | Botrytis, Ceratocystis, Fusarium, Pythium, Rhizoctonia, Rosellinia, Sclerotinia, and Sclerotium | Bio Ecologico Ltd. (Colombia) | [34] |
| Binab TF | T. polysporum | Fusarium, Pythium, Rhizoctonia, Sclerotinia, and Sclerotium | BINAB Bio-Innovation AB (USA) | [35] |
| Bioderma H | T. harzianum | Alternaria, Ascochyta, Cercospora, Colletotrichum, Fusarium, Phytophthora, Pythium, Macrophomina, Myrothecium, and Ralstonia | Biotech International Ltd. (India) | [34] |
| BioFungo™ | T. virens | Botrytis cinerea and Sphaerotheca pannosa | Orius Biotecnologia (Colombia) | [34] |
| ECO-77™ | T. harzianum | Botrytis and Eutypa | Plant Health Products (South Africa) | [34] |
| ECO-T™ | T. harzianum | Fusarium, Phytophthora, Pythium, and Rhizoctonia | Plant Health Products (South Africa) | [34] |
| Ecoderma | T. virens | Botrytis, Fusarium, Pythium, Rhizoctonia, Rosellinia, Sclerotinia, and Sclerotium | BigHaat Agro Ltd. (India) | [35] |
| Ecotrich ES™ | T. harzianum | Rhizoctonia solani, Pythium, and Sclerotinia | Ballagro Agro Tecnologia Ltd. (Brazil) | [34] |
| Esquive | T. atroviride | Pythium and Rhizoctonia | Agrauxine (France) | [35] |
| Floragard | T. hamatum | Fusarium, Pythium, Rhizoctonia solani, and Sclerotinia homeocarpa | Sellew Associates LLC (USA) | [35] |
| FoliGuard™ | T. hamatum | Alternaria, Botrytis cinerea, Cladosporium, Oidium, and Sphaeroteca pannosa | Live Systems Technology S.A. (Colombia) | [34] |
| Lycomax | T. viride | Soil-borne pathogens | Russell IPM (UK) | [34] |
| Natibiol™ | T. viride | Rhizoctonia | Probiagro S.A. (Venezuela) | [34] |
| PlantShield™/RootShield™ | T. harzianum | Fusarium, Pythium, Rhizoctonia solani, and Sclerotinia homeocarpa | Bioworks (USA) | [34] |
| T-Gro | T. harzianum | Botrytis, Ceratocystis, Fusarium, Pythium, and Rhizoctonia | Dagutat Biolab (USA) | [35] |
| Trianum™ | T. harzianum | Soil-borne pathogens | Koppert BV (The Netherlands) | [34,35] |
| Tricho™ | T. harzianum | Alternaria, Arrmilaria, Botrytis, Fusarium, Pythium, Rhizoctonia, Rosellinia, Sclerotinia, and Sclerotium | Orius Biotecnologia (Colombia) | [34] |
| Trichodermil™ | T. harzianum | Botrytis ricini, Fusarium, Phytophthora capsici, Phytophthora palmivora, Rhizoctonia, and Sclerotinia sclerotiorum | Itaforte BioProdutos (Brazil) | [34] |
| Trichodex | T. harzianum | Colletotrichum, Fusarium, Phytophthora, and Pythium, Macrophomina | Makhteshim chemical works Ltd. (USA) | [35] |
| Trichosav | T. harzianum | Soil-borne pathogens | Centros de Reproduccion de Medios Biologicos (Cuba) | [34] |
| Trichosoil | T. harzianum | Fusarium | Lage S.A. (Uruguay) | |
| Tusal | T. asperellum | Fusarium, Pythium, and Rhizoctonia solani | Isagro (USA) | [35] |
| Virisan | T. asperellum | Phytophthora, and Pythium | Isagro (USA) | [35] |
| VinevaxTM–Trichoprotection™ | T. harzianum | Armillaria, Botryosphaeria stevensii, Chondrostereum purpureum, Eutypa lata, and Phaeomoniella chlamydospor | Agrimm Technologies Ltd. (New Zealand) | [34,35] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahriar, S.A.; Islam, M.N.; Chun, C.N.W.; Kaur, P.; Rahim, M.A.; Islam, M.M.; Uddain, J.; Siddiquee, S. Microbial Metabolomics Interaction and Ecological Challenges of Trichoderma Species as Biocontrol Inoculant in Crop Rhizosphere. Agronomy 2022, 12, 900. https://doi.org/10.3390/agronomy12040900
Shahriar SA, Islam MN, Chun CNW, Kaur P, Rahim MA, Islam MM, Uddain J, Siddiquee S. Microbial Metabolomics Interaction and Ecological Challenges of Trichoderma Species as Biocontrol Inoculant in Crop Rhizosphere. Agronomy. 2022; 12(4):900. https://doi.org/10.3390/agronomy12040900
Chicago/Turabian StyleShahriar, Saleh Ahmed, M. Nazrul Islam, Charles Ng Wai Chun, Parwinder Kaur, Md. Abdur Rahim, Md. Mynul Islam, Jasim Uddain, and Shafiquzzaman Siddiquee. 2022. "Microbial Metabolomics Interaction and Ecological Challenges of Trichoderma Species as Biocontrol Inoculant in Crop Rhizosphere" Agronomy 12, no. 4: 900. https://doi.org/10.3390/agronomy12040900
APA StyleShahriar, S. A., Islam, M. N., Chun, C. N. W., Kaur, P., Rahim, M. A., Islam, M. M., Uddain, J., & Siddiquee, S. (2022). Microbial Metabolomics Interaction and Ecological Challenges of Trichoderma Species as Biocontrol Inoculant in Crop Rhizosphere. Agronomy, 12(4), 900. https://doi.org/10.3390/agronomy12040900