
Preference of Pollinators over Various Forage Mixtures and Microelement Treatments

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Locations
2.2. Meadow Mixtures
2.3. Design of Experiment
- (a)
- Control (without application of Zn and Se);
- (b)
- Foliar application of Se: 20 g Se ha−1 (in 250 L water ha−1) (form Na2SeO3) [38];
- (c)
- Foliar application of Zn: 200 g Zn ha−1 (in 250 L water ha−1) (form ZnSO4×7H2O) [45];
- (d)
- Foliar application of Se and Zn: 20 g Se ha−1 (form Na2SeO3) + 200 g Zn ha−1 (all in 250 l water ha−1) (form ZnSO4×7H2O).
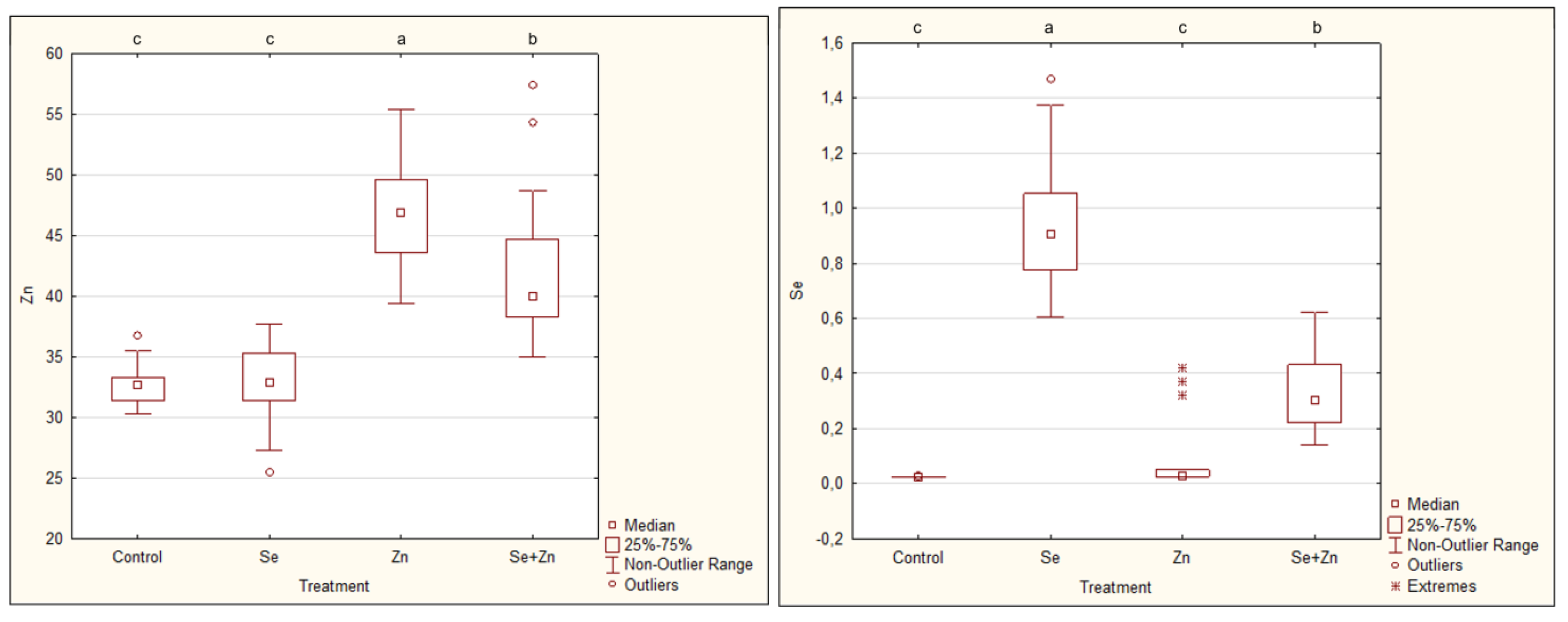
2.4. Determination of Zn and Se Content in Flowers
2.5. Statistical Analysis
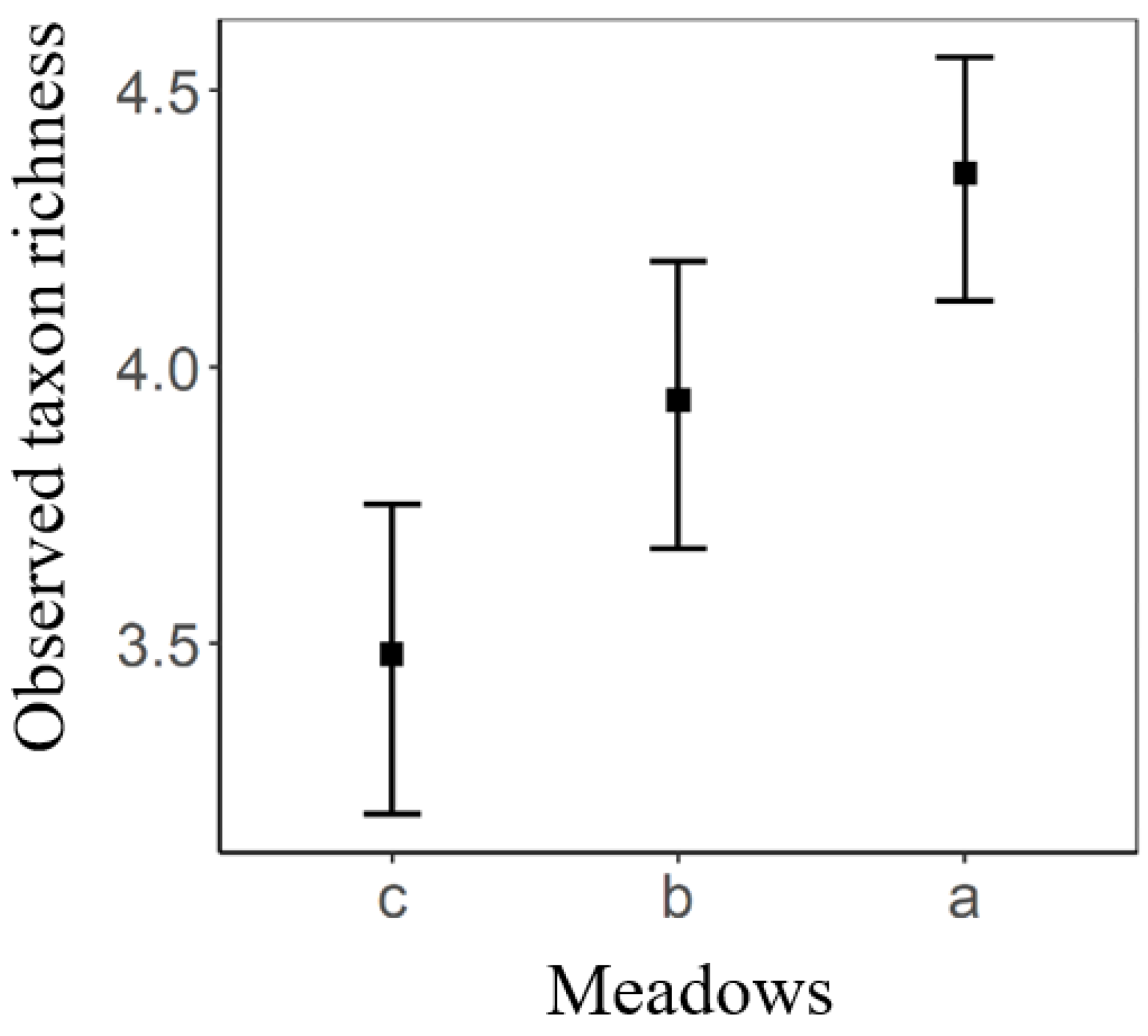
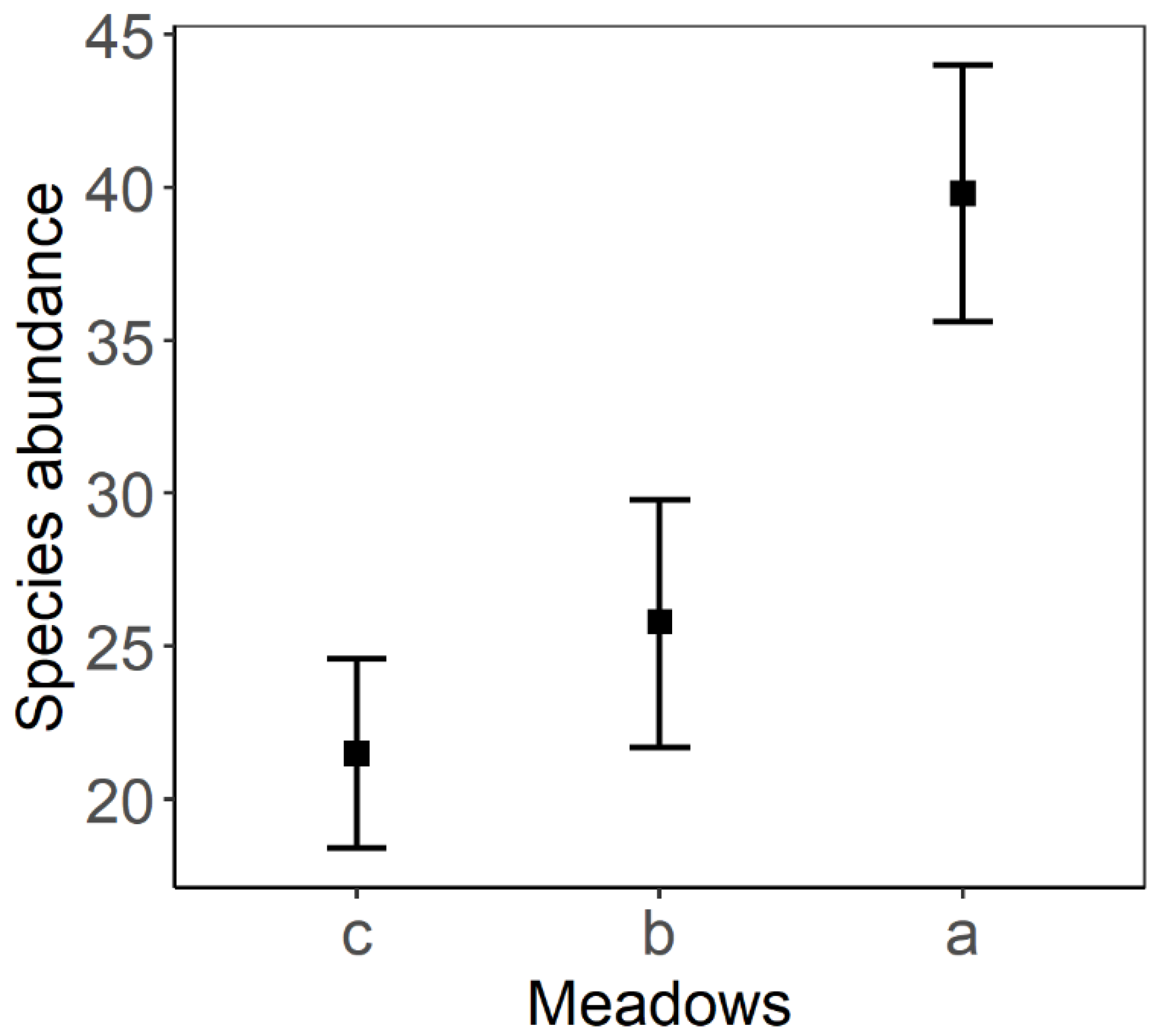
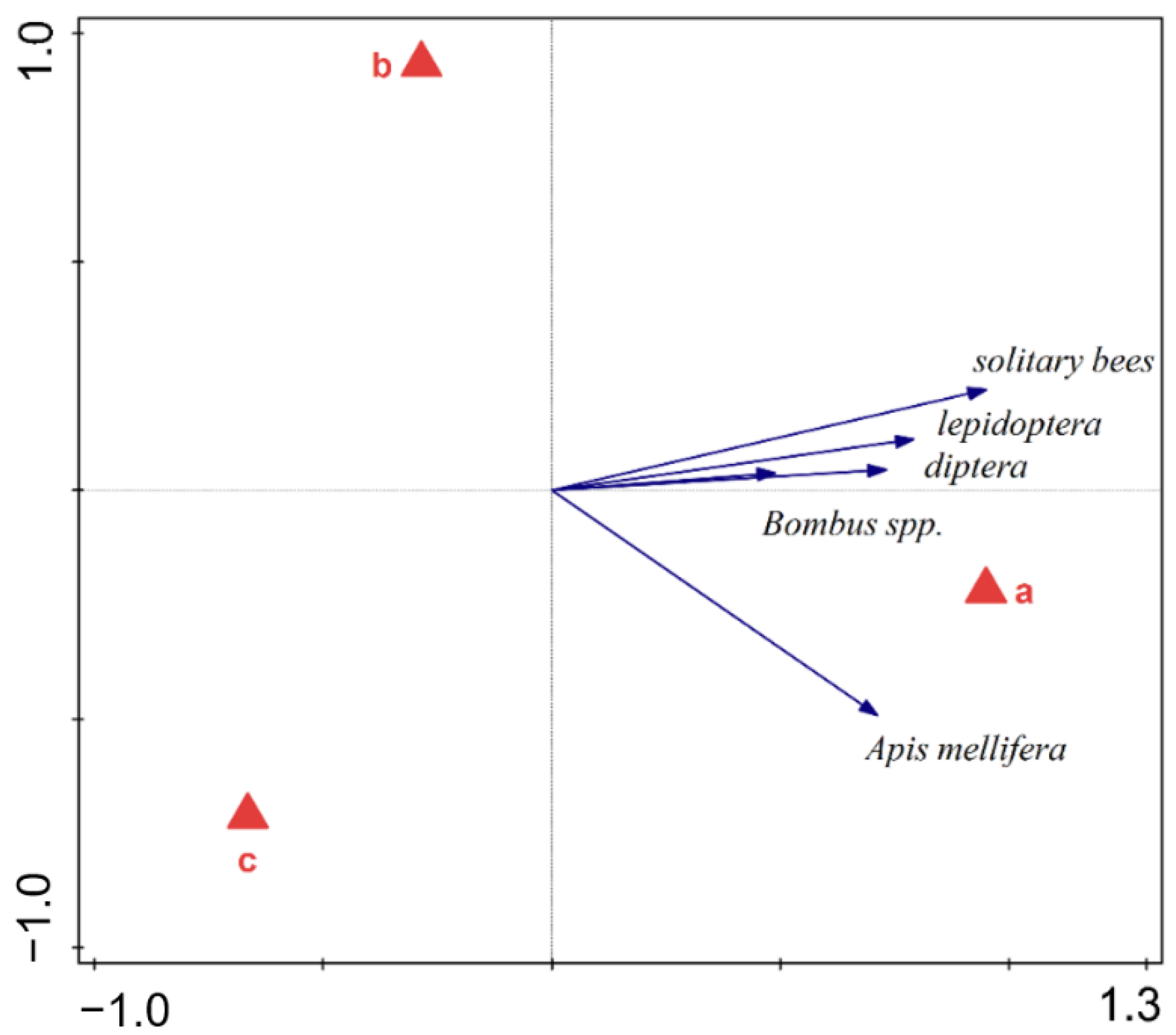
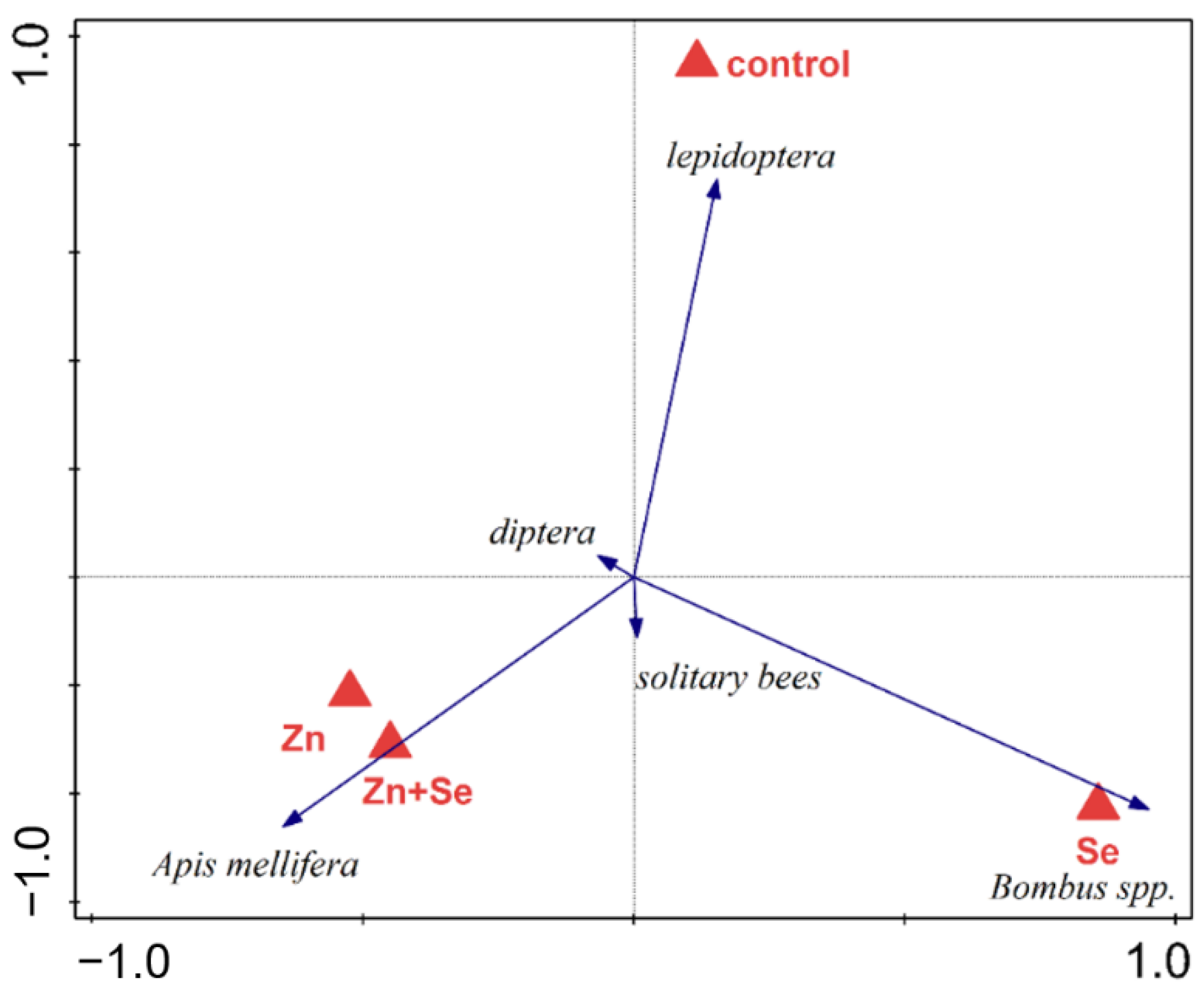
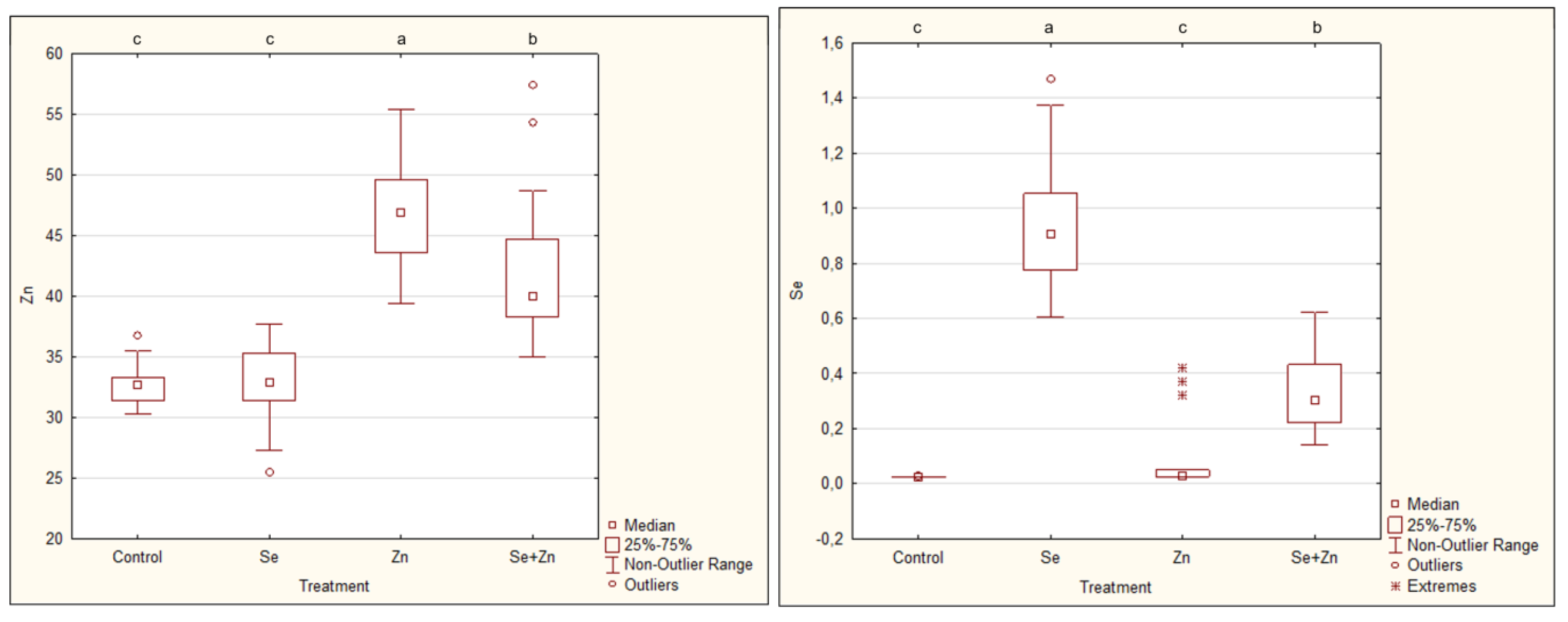
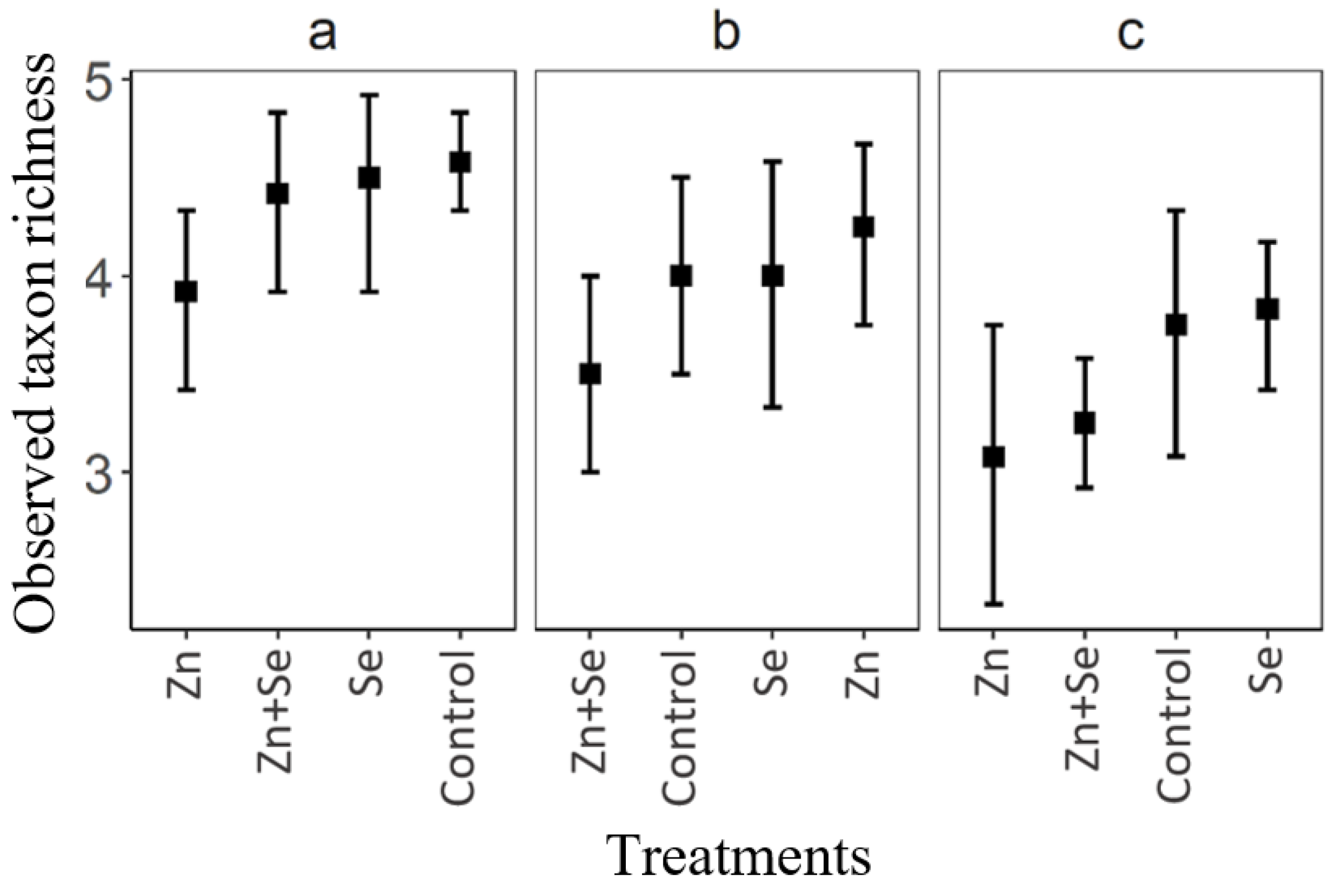
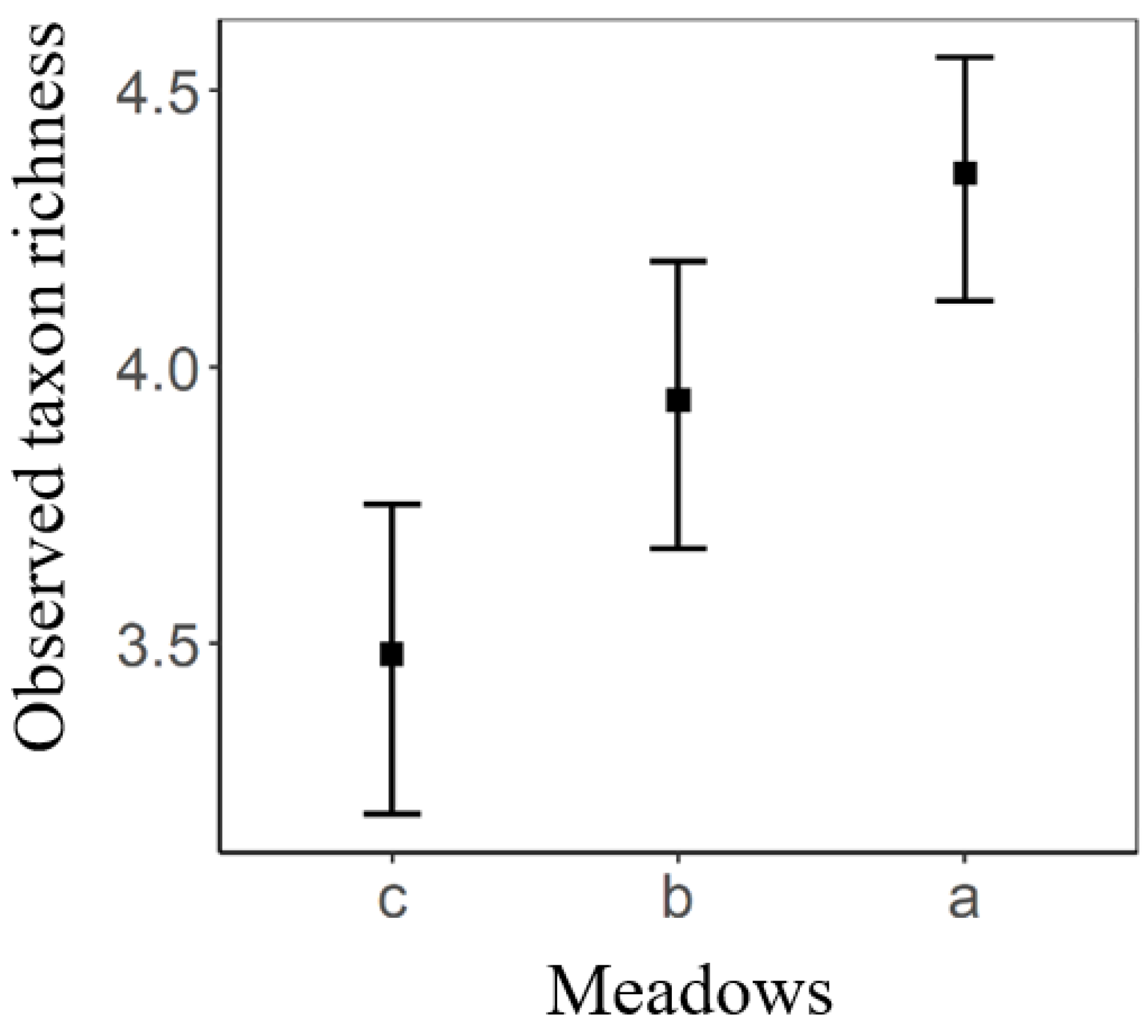
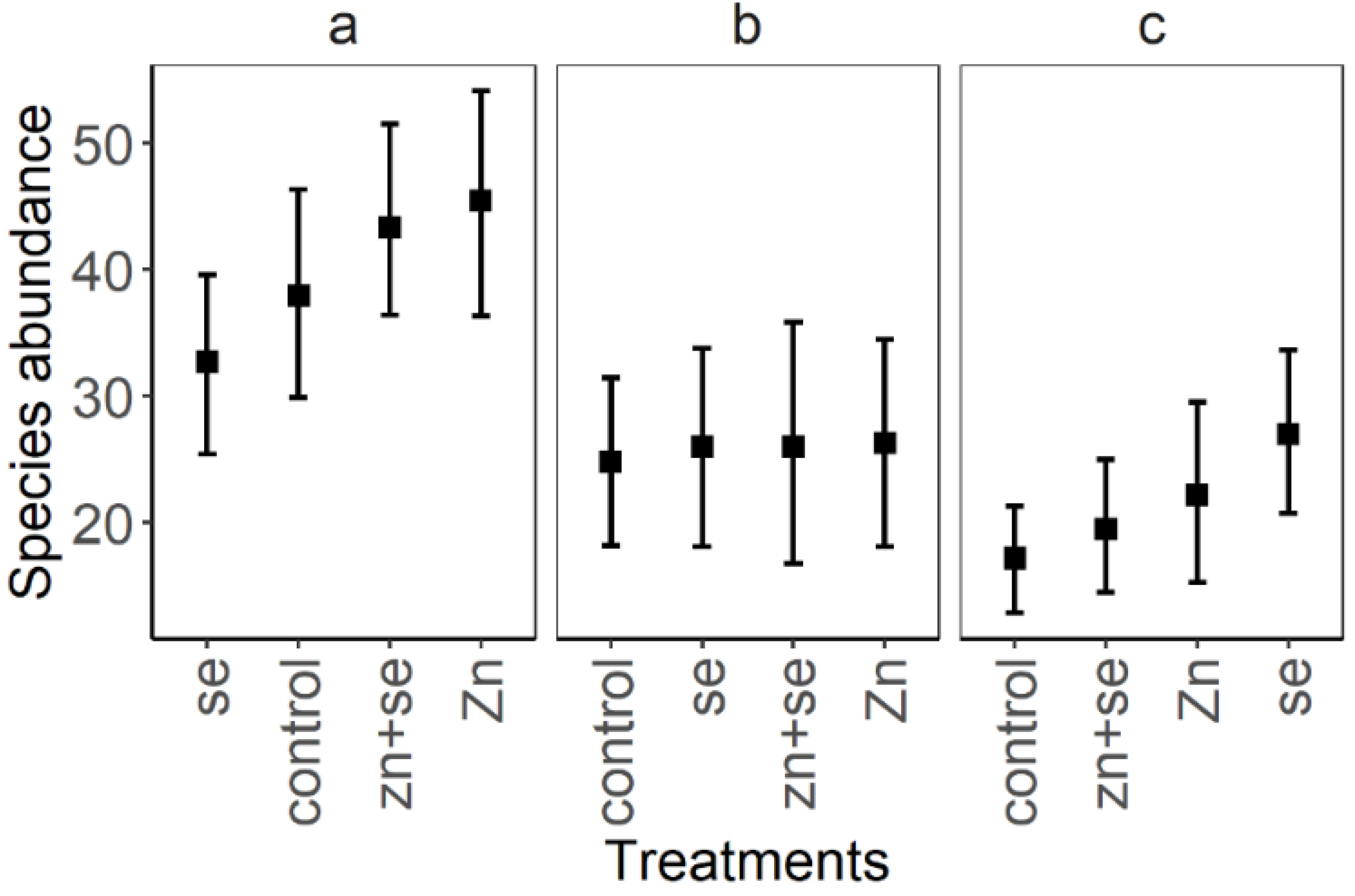
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ollerton, J. Pollinator Diversity: Distribution, Ecological Function, and Conservation. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 353–376. [Google Scholar] [CrossRef] [Green Version]
- Ollerton, J.; Winfree, R.; Tarrant, S. How Many Flowering Plants Are Pollinated by Animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Rodger, J.G.; Bennett, J.M.; Razanajatovo, M.; Knight, T.M.; van Kleunen, M.; Ashman, T.-L.; Steets, J.A.; Hui, C.; Arceo-Gómez, G.; Burd, M. Widespread Vulnerability of Flowering Plant Seed Production to Pollinator Declines. Sci. Adv. 2021, 7, eabd3524. [Google Scholar] [CrossRef] [PubMed]
- Wei, N.; Kaczorowski, R.L.; Arceo-Gómez, G.; O’Neill, E.M.; Hayes, R.A.; Ashman, T.-L. Pollinators Contribute to the Maintenance of Flowering Plant Diversity. Nature 2021, 597, 688–692. [Google Scholar] [CrossRef] [PubMed]
- Vladek, A.; Hybl, M.; Pridal, A. Pollination and Pollinators of Haskap (Lonicera Caerulea). Proceedings of International PhD Students Conference MendelNet, New York, NY, USA, 28 January 2018; pp. 211–216. [Google Scholar]
- Dmitruk, M.; Wrzesień, M.; Strzałkowska-Abramek, M.; Denisow, B. Pollen Food Resources to Help Pollinators. A Study of Five Ranunculaceae Species in Urban Forest. Urban For. Urban Green. 2021, 60, 127051. [Google Scholar] [CrossRef]
- Woodard, S.H.; Jha, S. Wild Bee Nutritional Ecology: Predicting Pollinator Population Dynamics, Movement, and Services from Floral Resources. Curr. Opin. Insect Sci. 2017, 21, 83–90. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Salignon, M.; Le Conte, Y.; Belzunces, L.P.; Decourtye, A.; Kretzschmar, A.; Suchail, S.; Brunet, J.-L.; Alaux, C. Influence of Pollen Nutrition on Honey Bee Health: Do Pollen Quality and Diversity Matter? PLoS ONE 2013, 8, e72016. [Google Scholar] [CrossRef] [Green Version]
- Biesmeijer, J.C.; Roberts, S.P.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D. Parallel Declines in Pollinators and Insect-Pollinated Plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef]
- Winfree, R.; Bartomeus, I.; Cariveau, D.P. Native Pollinators in Anthropogenic Habitats. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Bennett, J.M.; Steets, J.A.; Burns, J.H.; Burkle, L.A.; Vamosi, J.C.; Wolowski, M.; Arceo-Gómez, G.; Burd, M.; Durka, W.; Ellis, A.G. Land Use and Pollinator Dependency Drives Global Patterns of Pollen Limitation in the Anthropocene. Nat. Commun. 2020, 11, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global Pollinator Declines: Trends, Impacts and Drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Burkle, L.A.; Marlin, J.C.; Knight, T.M. Plant-Pollinator Interactions over 120 Years: Loss of Species, Co-Occurrence, and Function. Science 2013, 339, 1611–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naug, D. Nutritional Stress Due to Habitat Loss May Explain Recent Honeybee Colony Collapses. Biol. Conserv. 2009, 142, 2369–2372. [Google Scholar] [CrossRef]
- Alaux, C.; Ducloz, F.; Crauser, D.; Le Conte, Y. Diet Effects on Honeybee Immunocompetence. Biol. Lett. 2010, 6, 562–565. [Google Scholar] [CrossRef] [Green Version]
- Mráz, P.; Hýbl, M.; Kopecký, M.; Bohatá, A.; Hoštičková, I.; Šipoš, J.; Vočadlová, K.; Čurn, V. Screening of Honey Bee Pathogens in the Czech Republic and Their Prevalence in Various Habitats. Insects 2021, 12, 1051. [Google Scholar] [CrossRef]
- Wahl, O.; Ulm, K. Influence of Pollen Feeding and Physiological Condition on Pesticide Sensitivity of the Honey Bee Apis Mellifera Carnica. Oecologia 1983, 59, 106–128. [Google Scholar] [CrossRef]
- Roulston, T.H.; Goodell, K. The Role of Resources and Risks in Regulating Wild Bee Populations. Annu. Rev. Entomol. 2011, 56, 293–312. [Google Scholar] [CrossRef] [Green Version]
- Keller, I.; Fluri, P.; Imdorf, A. Pollen Nutrition and Colony Development in Honey Bees: Part 1. Bee World 2005, 86, 3–10. [Google Scholar] [CrossRef]
- Carvell, C.; Westrich, P.; Meek, W.R.; Pywell, R.F.; Nowakowski, M. Assessing the Value of Annual and Perennial Forage Mixtures for Bumblebees by Direct Observation and Pollen Analysis. Apidologie 2006, 37, 326–340. [Google Scholar] [CrossRef] [Green Version]
- Ziaja, M.; Denisow, B.; Wrzesień, M.; Wójcik, T. Availability of Food Resources for Pollinators in Three Types of Lowland Meadows. J. Apic. Res. 2018, 57, 467–478. [Google Scholar] [CrossRef]
- Bhandari, K.B.; West, C.; Longing, S.; Brown, C.; Green, P.; Barkowsky, E. Pollinator Abundance in Semiarid Pastures as Affected by Forage Species. Crop Sci. 2018, 58, 2665–2671. [Google Scholar] [CrossRef] [Green Version]
- Boehmel, C.; Lewandowski, I.; Claupein, W. Comparing Annual and Perennial Energy Cropping Systems with Different Management Intensities. Agric. Syst. 2008, 96, 224–236. [Google Scholar] [CrossRef]
- Dumbrovský, M.; Sobotková, V.; Šarapatka, B.; Chlubna, L.; Váchalová, R. Cost-Effectiveness Evaluation of Model Design Variants of Broad-Base Terrace in Soil Erosion Control. Ecol. Eng. 2014, 68, 260–269. [Google Scholar] [CrossRef]
- Arora, A.; Sairam, R.; Srivastava, G. Oxidative Stress and Antioxidative System in Plants. Curr. Sci. 2002, 1227–1238. [Google Scholar]
- Choi, H.W.; Hwang, B.K. The Pepper Extracellular Peroxidase CaPO2 Is Required for Salt, Drought and Oxidative Stress Tolerance as Well as Resistance to Fungal Pathogens. Planta 2012, 235, 1369–1382. [Google Scholar] [CrossRef]
- Petanidou, T.; Goethals, V.; Smets, E. The Effect of Nutrient and Water Availability on Nectar Secretion and Nectary Structure of the Dominant Labiatae Species of Phrygana. Syst. Geogr. Plants 1999, 233–244. [Google Scholar] [CrossRef]
- Quesada, M.; Bollman, K.; Stephenson, A.G. Leaf Damage Decreases Pollen Production and Hinders Pollen Performance in Cucurbita Texana. Ecology 1995, 76, 437–443. [Google Scholar] [CrossRef]
- Hanson, B.; Garifullina, G.F.; Lindblom, S.D.; Wangeline, A.; Ackley, A.; Kramer, K.; Norton, A.P.; Lawrence, C.B.; Pilon-Smits, E.A. Selenium Accumulation Protects Brassica Juncea from Invertebrate Herbivory and Fungal Infection. New Phytol. 2003, 159, 461–469. [Google Scholar] [CrossRef] [Green Version]
- Broadley, M.R.; White, P.J.; Hammond, J.P.; Zelko, I.; Lux, A. Zinc in Plants. New Phytol. 2007, 173, 677–702. [Google Scholar] [CrossRef]
- Gardener, M.C.; Gillman, M.P. The Effects of Soil Fertilizer on Amino Acids in the Floral Nectar of Corncockle, Agrostemma Githago (Caryophyllaceae). Oikos 2001, 92, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Jansen, M.A. 10 Ultraviolet-B Radiation: Stressor and Regulatory Signal. In Plant Stress Physiology, 2nd ed.; CAB International: Oxfordshire, UK, 2017; p. 253. [Google Scholar]
- Fazekas, C.; Péntek, A.; Vojnich, V.J. effect of foliar fertilizers applications on the nectar production of sunflower (Heliathus Annuus L.) and on the foraging behaviour of honeybees (Apis Mellifera L.). Ann. Fac. Eng. Hunedoara 2019, 17, 153–160. [Google Scholar]
- Sawidis, T.; Papadopoulou, A.; Voulgaropoulou, M. Effect of Zinc on Nectar Secretion of Hibiscus Rosa-Sinensis L. Protoplasma 2014, 251, 575–589. [Google Scholar] [CrossRef]
- Woo, H.R.; Kim, J.H.; Nam, H.G.; Lim, P.O. The Delayed Leaf Senescence Mutants of Arabidopsis, Ore1, Ore3, and Ore9 Are Tolerant to Oxidative Stress. Plant Cell Physiol. 2004, 45, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Karim, M.R.; Zhang, Y.; Zhao, R.; Chen, X.; Zhang, F.; Zou, C. Alleviation of Drought Stress in Winter Wheat by Late Foliar Application of Zinc, Boron, and Manganese. J. Plant Nutr. Soil Sci. 2012, 175, 142–151. [Google Scholar] [CrossRef]
- da Silva Folli-Pereira, M.; Ramos, A.C.; Canton, G.C.; da Conceição, J.M.; de Souza, S.B.; Cogo, A.J.D.; Figueira, F.F.; Eutrópio, F.J.; Rasool, N. Foliar Application of Trace Elements in Alleviating Drought Stress. Water Stress Crop Plants A Sustain. Approach 2016, 2, 669–681. [Google Scholar]
- Ducsay, L.; Ložek, O.; Marček, M.; Varényiová, M.; Hozlár, P.; Lošák, T. Possibility of Selenium Biofortification of Winter Wheat Grain. Plant Soil Environ. 2016, 62, 379–383. [Google Scholar] [CrossRef] [Green Version]
- Chilimba, A.D.; Young, S.D.; Black, C.R.; Meacham, M.C.; Lammel, J.; Broadley, M.R. Agronomic Biofortification of Maize with Selenium (Se) in Malawi. Field Crops Res. 2012, 125, 118–128. [Google Scholar] [CrossRef]
- Jiang, Y.; Zeng, Z.; Bu, Y.; Ren, C.; Li, J.; Han, J.; Tao, C.; Zhang, K.; Wang, X.; Lu, G. Effects of Selenium Fertilizer on Grain Yield, Se Uptake and Distribution in Common Buckwheat (Fagopyrum Esculentum Moench). Plant Soil Environ. 2015, 61, 371–377. [Google Scholar] [CrossRef] [Green Version]
- Cartes, P.; Gianfreda, L.; Mora, M. Uptake of Selenium and Its Antioxidant Activity in Ryegrass When Applied as Selenate and Selenite Forms. Plant Soil 2005, 276, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Zhang, W.; Cui, X.; Xu, B. Zinc Nutrition Increases the Antioxidant Defenses of Honey Bees. Entomol. Exp. Et Appl. 2015, 156, 201–210. [Google Scholar] [CrossRef]
- Sobolev, O.; Gutyj, B.; Petryshak, R.; Pivtorak, J.; Kovalskyi, Y.; Naumyuk, A.; Petryshak, O.; Semchuk, I.; Mateusz, V.; Shcherbatyy, A. Biological Role of Selenium in the Organism of Animals and Humans. Ukr. J. Ecol. 2018, 8, 654–665. [Google Scholar] [CrossRef]
- Hýbl, M.; Mráz, P.; Šipoš, J.; Hoštičková, I.; Bohatá, A.; Čurn, V.; Kopec, T. Polyphenols as Food Supplement Improved Food Consumption and Longevity of Honey Bees (Apis Mellifera) Intoxicated by Pesticide Thiacloprid. Insects 2021, 12, 572. [Google Scholar] [CrossRef]
- Wang, S.; Tian, X.; Liu, Q. The Effectiveness of Foliar Applications of Zinc and Biostimulants to Increase Zinc Concentration and Bioavailability of Wheat Grain. Agronomy 2020, 10, 178. [Google Scholar] [CrossRef] [Green Version]
- ter Braak, C.J.; Smilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, Version 5.0; Microcomputer Power: Ithaca, NY, USA, 2012.
- Viik, E.; Maend, M.; Karise, R.; Laeaeniste, P.; Williams, I.H.; Luik, A. The Impact of Foliar Fertilization on the Number of Bees (Apoidea) on Spring Oilseed Rape. Zemdirbyste 2012, 99, 41–46. [Google Scholar]
- Hendriksma, H.P.; Toth, A.L.; Shafir, S. Individual and Colony Level Foraging Decisions of Bumble Bees and Honey Bees in Relation to Balancing of Nutrient Needs. Front. Ecol. Evol. 2019, 7, 177. [Google Scholar] [CrossRef] [Green Version]
- Mader, E.; Spivak, M.; Evans, E. Managing Alternative Pollinators; NRAES: Ithaca, NY, USA, 2010. [Google Scholar]
- Galef Jr, B.G.; Giraldeau, L.-A. Social Influences on Foraging in Vertebrates: Causal Mechanisms and Adaptive Functions. Anim. Behav. 2001, 61, 3–15. [Google Scholar] [CrossRef]
- Kay, A. Applying Optimal Foraging Theory to Assess Nutrient Availability Ratios for Ants. Ecology 2002, 83, 1935–1944. [Google Scholar] [CrossRef]
- Wilson, E.O. The Insect Societies; Harvard University Press: Cambridge, MA, USA, 1971. [Google Scholar]
- Nisbet, C.; Guler, A.; Ormancı, N.; Cenesiz, S. Preventive Action of Zinc against Heavy Metals Toxicity in Honeybee. Afr. J. Biochem. Res. 2018, 12, 1–6. [Google Scholar]
- Hladun, K.R.; Kaftanoglu, O.; Parker, D.R.; Tran, K.D.; Trumble, J.T. Effects of Selenium on Development, Survival, and Accumulation in the Honeybee (Apis Mellifera L.). Environ. Toxicol. Chem. 2013, 32, 2584–2592. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Sparrow, K.R. Evidence for Competition between Honeybees and Bumblebees; Effects on Bumblebee Worker Size. J. Insect Conserv. 2009, 13, 177–181. [Google Scholar] [CrossRef] [Green Version]
- Walther-Hellwig, K.; Fokul, G.; Frankl, R.; Büchler, R.; Ekschmitt, K.; Wolters, V. Increased Density of Honeybee Colonies Affects Foraging Bumblebees. Apidologie 2006, 37, 517–532. [Google Scholar] [CrossRef] [Green Version]
- Tuzen, M.; Silici, S.; Mendil, D.; Soylak, M. Trace Element Levels in Honeys from Different Regions of Turkey. Food Chem. 2007, 103, 325–330. [Google Scholar] [CrossRef]
- Hladun, K.R.; Parker, D.R.; Tran, K.D.; Trumble, J.T. Effects of Selenium Accumulation on Phytotoxicity, Herbivory, and Pollination Ecology in Radish (Raphanus Sativus L.). Environ. Pollut. 2013, 172, 70–75. [Google Scholar] [CrossRef]
- Hladun, K.R.; Parker, D.R.; Trumble, J.T. Selenium Accumulation in the Floral Tissues of Two Brassicaceae Species and Its Impact on Floral Traits and Plant Performance. Environ. Exp. Bot. 2011, 74, 90–97. [Google Scholar] [CrossRef]
- Wu, L. Review of 15 Years of Research on Ecotoxicology and Remediation of Land Contaminated by Agricultural Drainage Sediment Rich in Selenium. Ecotoxicol. Environ. Saf. 2004, 57, 257–269. [Google Scholar] [CrossRef]
- Vickerman, D.B.; Trumble, J.T.; George, G.N.; Pickering, I.J.; Nichol, H. Selenium Biotransformations in an Insect Ecosystem: Effects of Insects on Phytoremediation. Environ. Sci. Technol. 2004, 38, 3581–3586. [Google Scholar] [CrossRef]
- Quinn, C.F.; Prins, C.N.; Freeman, J.L.; Gross, A.M.; Hantzis, L.J.; Reynolds, R.J.; in Yang, S.; Covey, P.A.; Bañuelos, G.S.; Pickering, I.J. Selenium Accumulation in Flowers and Its Effects on Pollination. New Phytol. 2011, 192, 727–737. [Google Scholar] [CrossRef]
- Varjú, D.; Núñez, J. What Do Foraging Honeybees Optimize? J. Comp. Physiol. A 1991, 169, 729–736. [Google Scholar] [CrossRef]
- Pankiw, T.; Page Jr, R. The Effect of Genotype, Age, Sex, and Caste on Response Thresholds to Sucrose and Foraging Behavior of Honey Bees (Apis Mellifera L.). J. Comp. Physiol. A 1999, 185, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.R. Division of Labor in Honeybees: Form, Function, and Proximate Mechanisms. Behav. Ecol. Sociobiol. 2010, 64, 305–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, M.; Shibata, A.; Yasui, T.; Nagamasu, H. Impact of Introduced Honeybees, Apis Mellifera, upon Native Bee Communities in the Bonin (Ogasawara) Islands. Popul. Ecol. 1999, 41, 217–228. [Google Scholar] [CrossRef]
- Sugden, E.A.; Thorp, R.W.; Buchmann, S.L. Honey Bee-Native Bee Competition: Focal Point for Environmental Change and Apicultural Response in Australia. Bee World 1996, 77, 26–44. [Google Scholar] [CrossRef]
- Thomson, D.M.; Page, M.L. The Importance of Competition between Insect Pollinators in the Anthropocene. Curr. Opin. Insect Sci. 2020, 38, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Goras, G.; Tananaki, C.; Dimou, M.; Tscheulin, T.; Petanidou, T.; Thrasyvoulou, A. Impact of Honeybee (Apis Mellifera L.) Density on Wild Bee Foraging Behaviour. J. Apic. Sci. 2016, 60, 49. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.D.; Hendrix, S.D.; Debinski, D.M.; Hemsley, C.J. Butterfly, Bee and Forb Community Composition and Cross-Taxon Incongruence in Tallgrass Prairie Fragments. J. Insect Conserv. 2008, 12, 69–79. [Google Scholar] [CrossRef]
- Fernández, V.; Brown, P.H. From Plant Surface to Plant Metabolism: The Uncertain Fate of Foliar-Applied Nutrients. Front. Plant Sci. 2013, 4, 289. [Google Scholar] [CrossRef] [Green Version]
- Kurešová, G.; Menšík, L.; Haberle, J.; Svoboda, P.; Raimanova, I. Influence of Foliar Micronutrients Fertilization on Nutritional Status of Apple Trees. Plant Soil Environ. 2019, 65, 320–327. [Google Scholar] [CrossRef] [Green Version]
- Zbíral, J. Determination of Plant-Available Micronutrients by the Mehlich 3 Soil Extractant–a Proposal of Critical Values. Plant Soil Environ. 2016, 62, 527–531. [Google Scholar] [CrossRef] [Green Version]
- Száková, J.; Tremlová, J.; Pegová, K.; Najmanová, J.; Tlustoš, P. Soil-to-Plant Transfer of Native Selenium for Wild Vegetation Cover at Selected Locations of the Czech Republic. Environ. Monit. Assess. 2015, 187, 1–9. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | Type of Soil | Longterm Average | 2021 | |||
|---|---|---|---|---|---|---|
| Soil Type (WRB) | Soil Texture Class (WRB) | Temparature (°C) | Precipitation (mm) | Temparature (°C) | Precipitation (mm) | |
| Vatín | cambisols | sundy loam | 6.9 | 617.5 | 7.2 | 658.7 |
| Troubsko | chernozems | clay loam | 9.5 | 506.5 | 9.4 | 565.3 |
| Plants | Latin Name | Composition (%) | ||
|---|---|---|---|---|
| Mixture a | Mixture b | Mixture c | ||
| Blossom herbs | Carum carvi | 1.70 | 0.60 | 0.20 |
| Centaurea jacea | 1.70 | 0.60 | 0.20 | |
| Fagopyrum esculentum | 17.10 | 5.70 | 2.00 | |
| Knautia arvensis | 4.30 | 1.40 | 0.50 | |
| Leucanthemum vulgare | 8.60 | 2.80 | 1.00 | |
| Phacelia tanacefolia | 17.10 | 5.70 | 2.00 | |
| Salvia pratensis | 6.00 | 2.00 | 0.70 | |
| Sanguisorba minor | 2.60 | 0.90 | 0.30 | |
| Silene vulgaris | 0.90 | 0.30 | 0.10 | |
| Total | 60.00 | 20.00 | 7.00 | |
| Fabaceae | Anthyllis vulneraria | 5.30 | 1.30 | 0.40 |
| Lotus corniculatus | 26.70 | 6.70 | 2.00 | |
| Onobrychis viciifolia | 6.70 | 1.70 | 0.50 | |
| Trifolium pratense | 1.30 | 0.30 | 0.10 | |
| Total | 40.00 | 10.00 | 3.00 | |
| Poaceae | Agrostis capillaris | 2.30 | 3.00 | |
| Anthoxanthum odoratum | 3.10 | 4.00 | ||
| Arrhenatherum elatius | 2.30 | 3.00 | ||
| Bromus erectus | 7.80 | 10.00 | ||
| Cynosorus cristatus | 6.20 | 8.00 | ||
| Festuca pratensis | 6.20 | 8.00 | ||
| Festuca rubra rubra | 11.70 | 15.00 | ||
| Festuca rubra trichophylla | 5.40 | 7.00 | ||
| Festuca rubra commutata | 3.90 | 5.00 | ||
| Festuca trachyphylla | 7.80 | 10.00 | ||
| Lolium perenne | 1.60 | 2.00 | ||
| Phleum pratense | 1.60 | 2.00 | ||
| Poa pratensís | 7.80 | 10.00 | ||
| Trisetum flavescens | 2.30 | 3.00 | ||
| Total | 70.00 | 90.00 | ||
| Sum Sq | Mean Sq | NumDF | DenDF | F-Value | p-Value | |
|---|---|---|---|---|---|---|
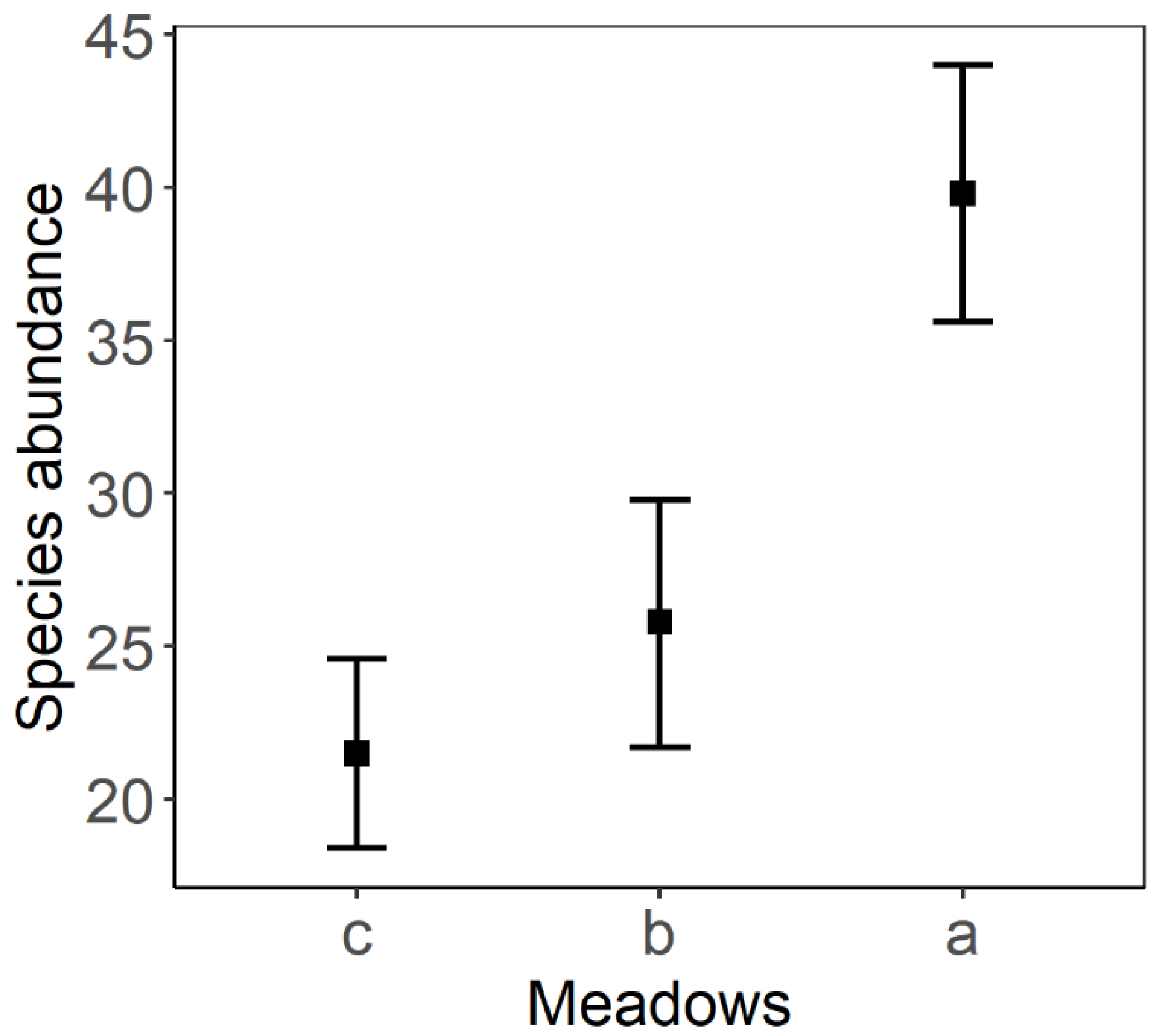
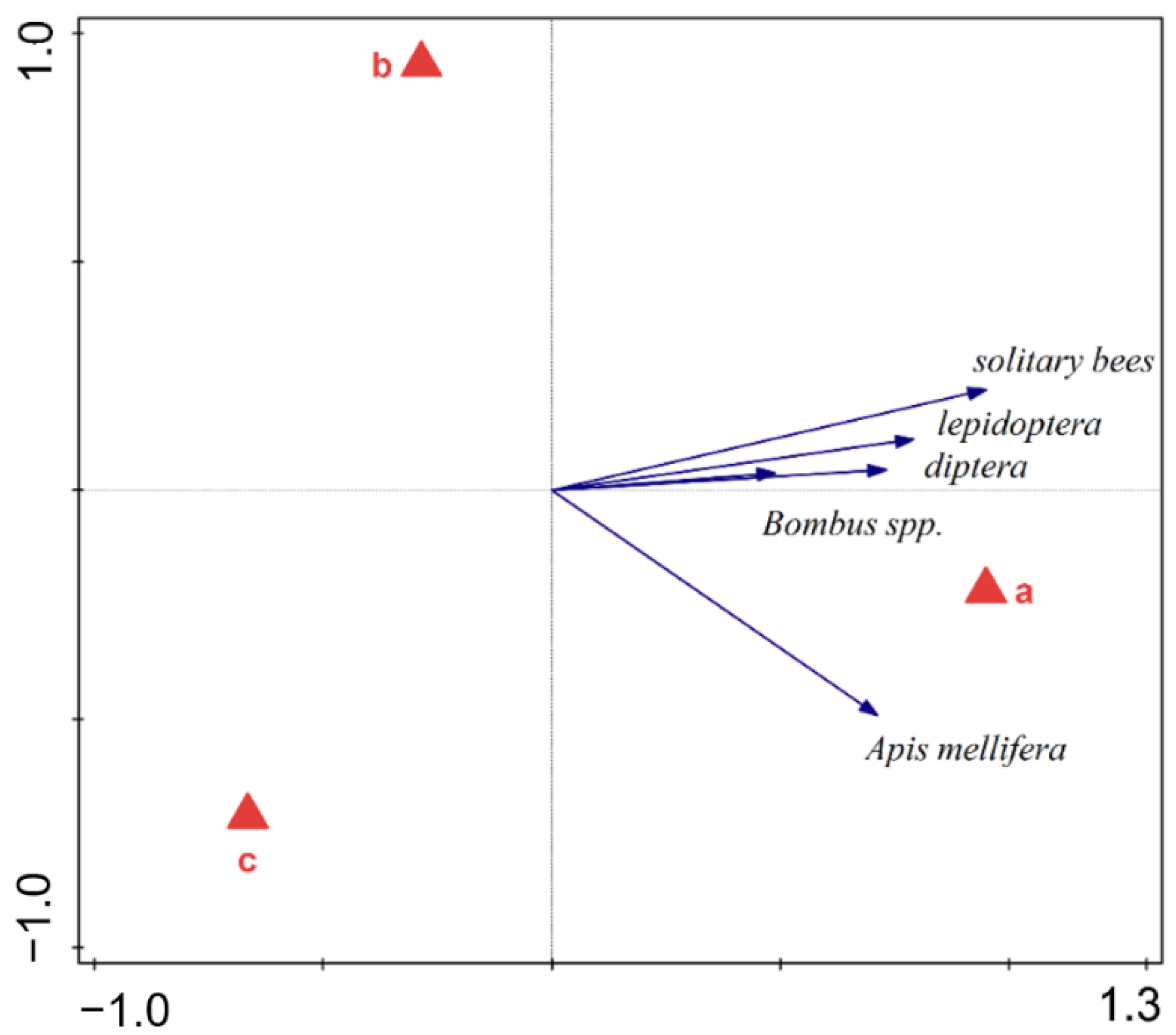
| Meadows | 85.386 | 42.693 | 2 | 8 | 11.673 | 0.004 |
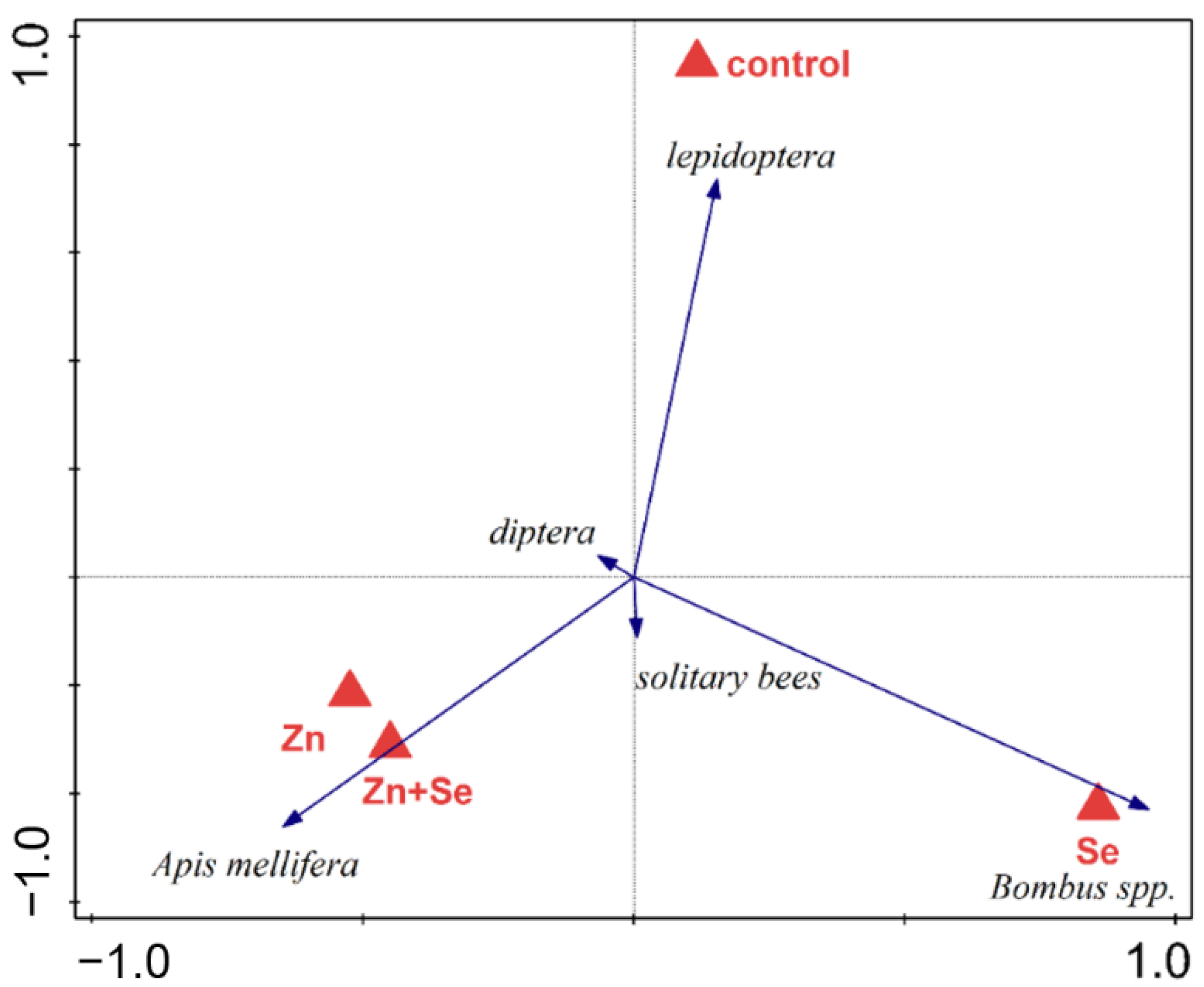
| Treatment | 18.671 | 6.224 | 3 | 3 | 1.702 | 0.337 |
| Meadows:Treatment | 32.061 | 5.344 | 6 | 8 | 1.461 | 0.302 |
| Contrasts | Estimate | SE | Df. | T.Ratio | p-Value |
|---|---|---|---|---|---|
| a–b | 1.07 | 0.403 | 126 | 2.654 | 0.024 |
| a–c | 2.15 | 0.403 | 126 | 5.343 | <0.001 |
| b–c | 1.08 | 0.403 | 126 | 2.689 | 0.022 |
| Sum Sq | Mean Sq | NumDF | DenDF | F-Value | p-Value | |
|---|---|---|---|---|---|---|
| Meadows | 133.925 | 66.962 | 2 | 8 | 20.661 | <0.001 |
| Treatment | 4.518 | 1.506 | 3 | 3 | 1.701 | 0.727 |
| Meadows:Treatment | 19.674 | 3.279 | 6 | 8 | 1.461 | 0.479 |
| Contrasts | Estimate | SE | Df. | T.Ratio | p-Value |
|---|---|---|---|---|---|
| a–b | 2.808 | 0.405 | 126 | 6.938 | <0.001 |
| a–c | 3.675 | 0.405 | 126 | 9.079 | <0.001 |
| b–c | 0.867 | 0.405 | 126 | 2.141 | 0.086 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hýbl, M.; Šipoš, J.; Krejčová, A.; Sodomová, K.; Polák, O.; Koláčková, I.; Mráz, P. Preference of Pollinators over Various Forage Mixtures and Microelement Treatments. Agronomy 2022, 12, 370. https://doi.org/10.3390/agronomy12020370
Hýbl M, Šipoš J, Krejčová A, Sodomová K, Polák O, Koláčková I, Mráz P. Preference of Pollinators over Various Forage Mixtures and Microelement Treatments. Agronomy. 2022; 12(2):370. https://doi.org/10.3390/agronomy12020370
Chicago/Turabian StyleHýbl, Marian, Jan Šipoš, Anna Krejčová, Karolína Sodomová, Ondřej Polák, Ivana Koláčková, and Petr Mráz. 2022. "Preference of Pollinators over Various Forage Mixtures and Microelement Treatments" Agronomy 12, no. 2: 370. https://doi.org/10.3390/agronomy12020370
APA StyleHýbl, M., Šipoš, J., Krejčová, A., Sodomová, K., Polák, O., Koláčková, I., & Mráz, P. (2022). Preference of Pollinators over Various Forage Mixtures and Microelement Treatments. Agronomy, 12(2), 370. https://doi.org/10.3390/agronomy12020370