
Comparative Study on the Physio-Biochemical Responses of Spring and Winter Barley Genotypes under Vernalized and Greenhouse Conditions


 , ,
, , 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Material and Growth Conditions
2.2. Analysis of Morphological and Photosynthetic Parameters
2.3. Detection of Antioxidant Activity and Lipid Peroxidation Assay
2.4. Gene Expression Analysis
2.5. Statistical Analysis
3. Results
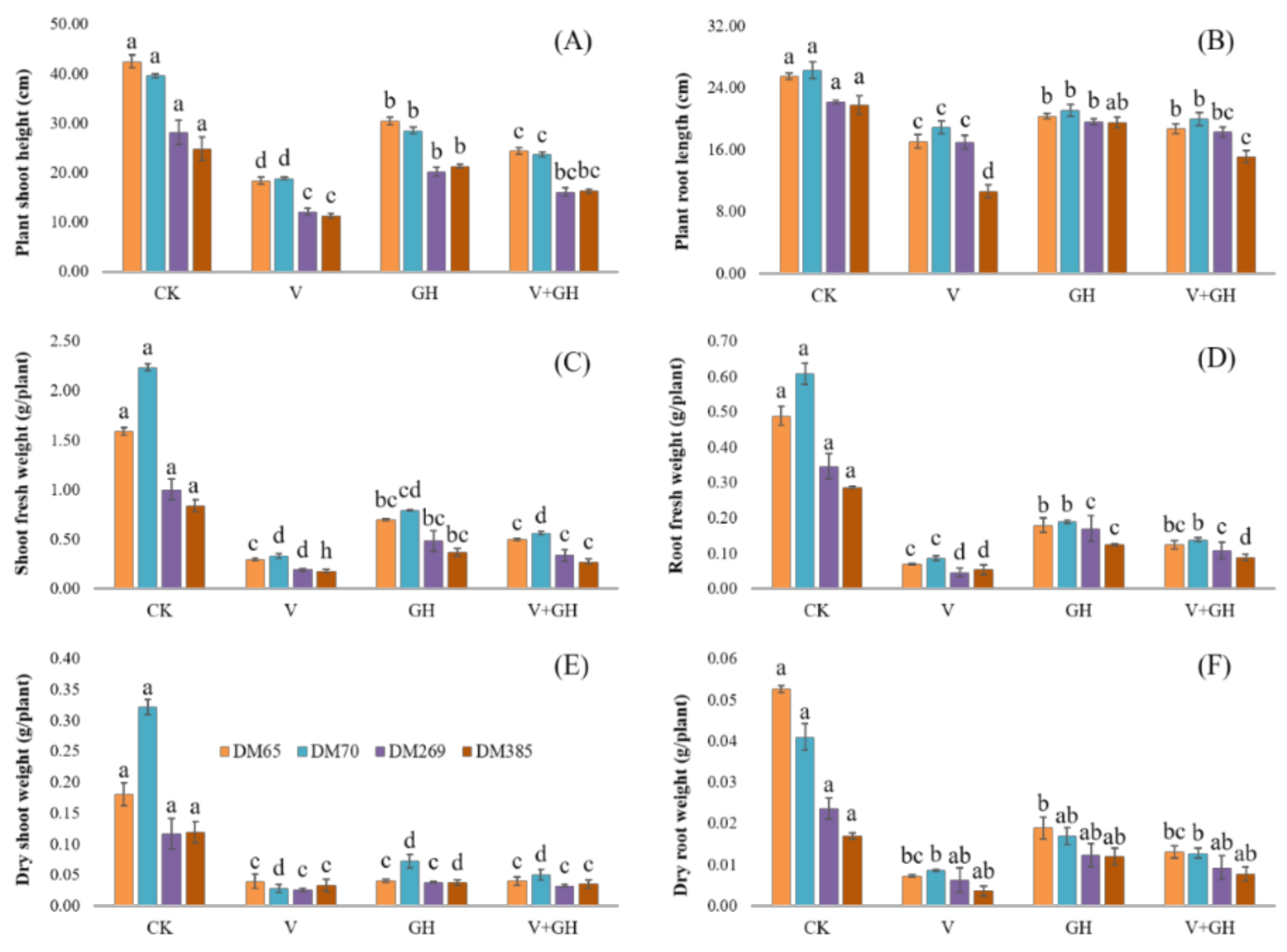
3.1. Plant Growth Attributes
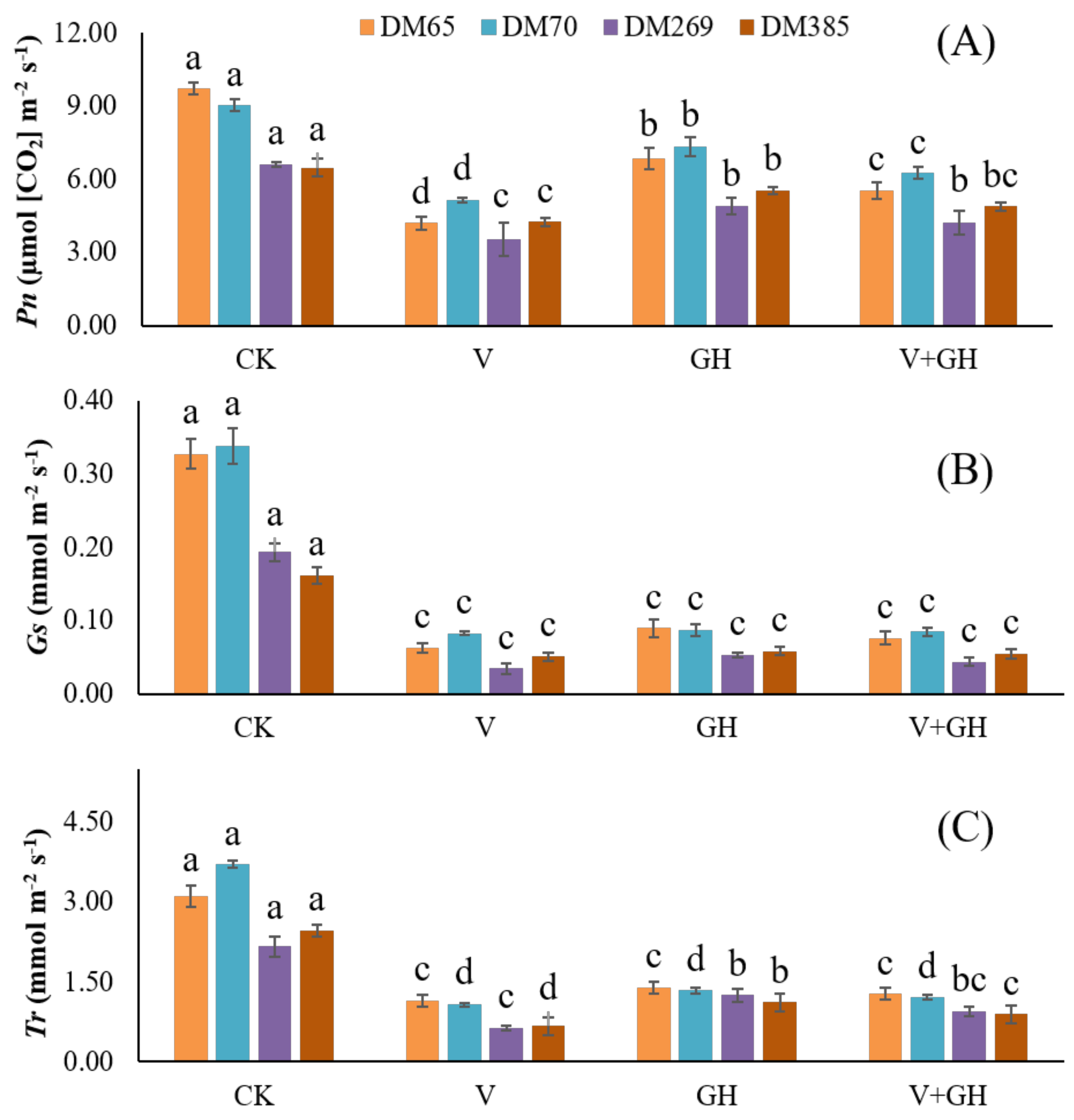
3.2. Photosynthetic Parameters
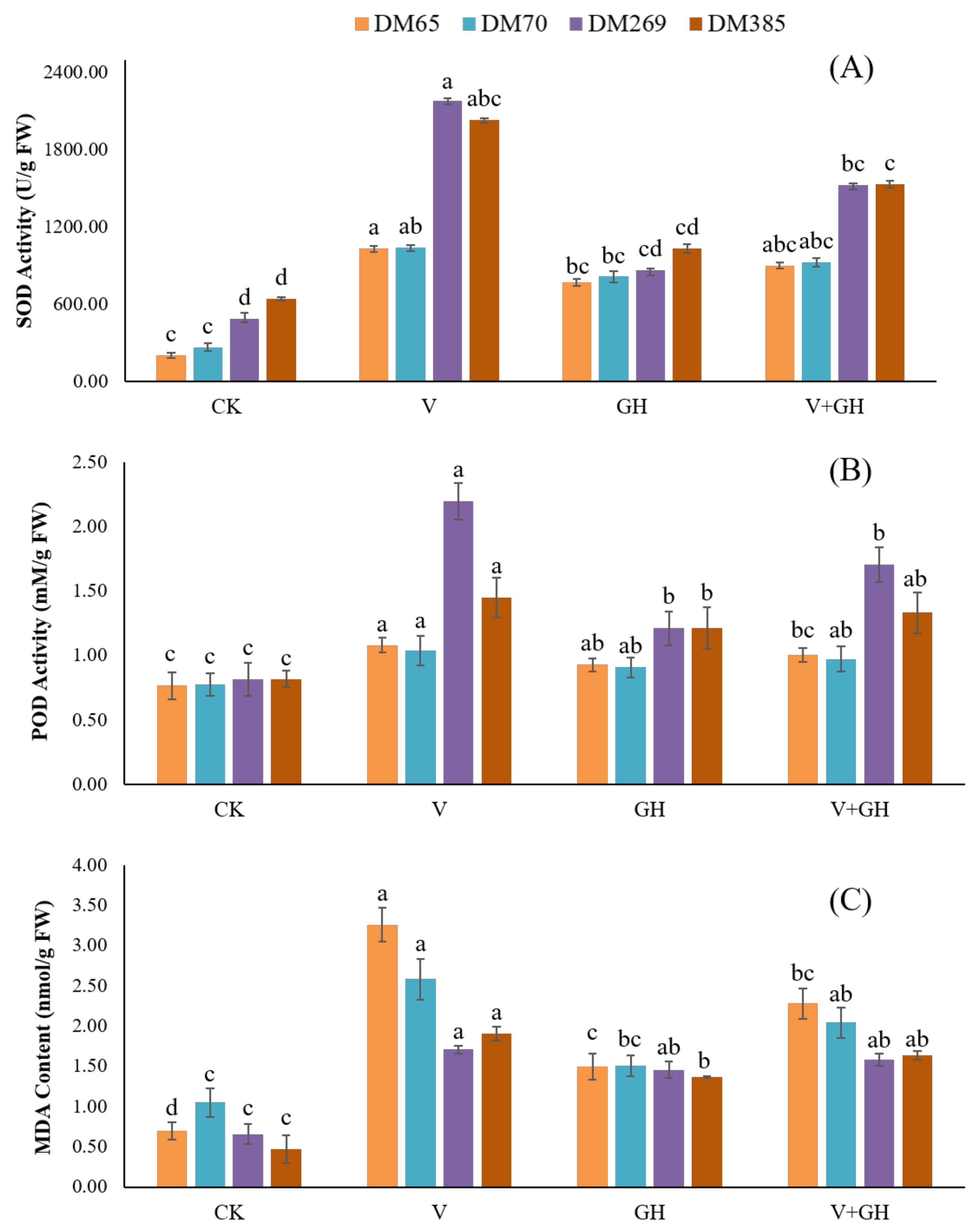
3.3. Membrane Lipid Peroxidation and Antioxidant Enzyme Activity
3.4. Expression of Candidate Genes in Different Environments
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Ethics Statement
References
- Chen, A.; Li, C.; Hu, W.; Lau, M.Y.; Lin, H.; Rockwell, N.C.; Martin, S.S.; Jernstedt, J.A.; Lagarias, J.C.; Dubcovsky, J. PHYTOCHROME C plays a major role in the acceleration of wheat flowering under long-day photoperiod. Proc. Natl. Acad. Sci. USA 2014, 111, 10037–10044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehar, S.; Adil, M.F.; Zeeshan, M.; Holford, P.; Cao, F.; Wu, F.; Wang, Y. Mechanistic insights into potassium-conferred drought stress tolerance in cultivated and tibetan wild barley: Differential osmoregulation, nutrient retention, secondary metabolism and antioxidative defense capacity. Int. J. Mol. Sci. 2021, 22, 13100. [Google Scholar] [CrossRef]
- Shcherban, A.; Khlestkina, E.; Efremova, T.; Salina, E. The effect of two differentially expressed wheat VRN-B1 alleles on the heading time is associated with structural variation in the first intron. Genetica 2013, 141, 133–141. [Google Scholar] [CrossRef]
- Acevedo, E.; Silva, P.; Silva, H. Wheat growth and physiology. Bread Wheat Improv. Prod. 2002, 30, 39–70. [Google Scholar]
- Wigge, P.A.; Kim, M.C.; Jaeger, K.E.; Busch, W.; Schmid, M.; Lohmann, J.U.; Weigel, D. Integration of spatial and temporal information during floral induction in Arabidopsis. Science 2005, 309, 1056–1059. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Fu, D.; Li, C.; Blechl, A.; Tranquilli, G.; Bonafede, M.; Sanchez, A.; Valarik, M.; Yasuda, S.; Dubcovsky, J. The wheat and barley vernalization gene VRN3 is an orthologue of FT. Proc. Natl. Acad. Sci. USA 2006, 103, 19581–19586. [Google Scholar] [CrossRef] [Green Version]
- Fu, D.; Szűcs, P.; Yan, L.; Helguera, M.; Skinner, J.S.; Von Zitzewitz, J.; Hayes, P.M.; Dubcovsky, J. Large deletions within the first intron in VRN-1 are associated with spring growth habit in barley and wheat. Mol. Genet. Genom. 2005, 273, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Díaz, A.; Zikhali, M.; Turner, A.S.; Isaac, P.; Laurie, D.A. Copy number variation affecting the Photoperiod-B1 and Vernalization-A1 genes is associated with altered flowering time in wheat (Triticum aestivum). PLoS ONE 2012, 7, e33234. [Google Scholar] [CrossRef] [Green Version]
- Guedira, M.; Maloney, P.; Xiong, M.; Petersen, S.; Murphy, J.P.; Marshall, D.; Johnson, J.; Harrison, S.; Brown-Guedira, G. Vernalization duration requirement in soft winter wheat is associated with variation at the VRN-B1 locus. Crop Sci. 2014, 54, 1960–1971. [Google Scholar] [CrossRef]
- Snape, J.; Butterworth, K.; Whitechurch, E.; Worland, A. Waiting for fine times: Genetics of flowering time in wheat. In Wheat in a Global Environment; Springer: Berlin/Heidelberg, Germany, 2001; pp. 67–74. [Google Scholar]
- Turner, A.; Beales, J.; Faure, S.; Dunford, R.P.; Laurie, D.A. The pseudo-response regulator Ppd-H1 provides adaptation to photoperiod in barley. Science 2005, 310, 1031–1034. [Google Scholar] [CrossRef]
- Beales, J.; Turner, A.; Griffiths, S.; Snape, J.W.; Laurie, D.A. A pseudo-response regulator is misexpressed in the photoperiod insensitive Ppd-D1a mutant of wheat (Triticum aestivum L.). Theor. Appl. Genet. 2007, 115, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Shaw, L.M.; Turner, A.S.; Laurie, D.A. The impact of photoperiod insensitive Ppd1a mutations on the photoperiod pathway across the three genomes of hexaploid wheat (Triticum aestivum). Plant J. 2012, 71, 71–84. [Google Scholar] [CrossRef]
- Alqudah, A.M.; Sharma, R.; Pasam, R.K.; Graner, A.; Kilian, B.; Schnurbusch, T. Genetic dissection of photoperiod response based on GWAS of pre-anthesis phase duration in spring barley. PLoS ONE 2014, 9, e113120. [Google Scholar] [CrossRef] [Green Version]
- Dyck, J.; Matus-Cádiz, M.; Hucl, P.; Talbert, L.; Hunt, T.; Dubuc, J.; Nass, H.; Clayton, G.; Dobb, J.; Quick, J. Agronomic performance of hard red spring wheat isolines sensitive and insensitive to photoperiod. Crop Sci. 2004, 44, 1976–1981. [Google Scholar] [CrossRef]
- Woods, D.P.; Ream, T.S.; Minevich, G.; Hobert, O.; Amasino, R.M. PHYTOCHROME C is an essential light receptor for photoperiodic flowering in the temperate grass, Brachypodium distachyon. Genetics 2014, 198, 397–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Möglich, A.; Yang, X.; Ayers, R.A.; Moffat, K. Structure and function of plant photoreceptors. Annu. Rev. Plant Biol. 2010, 61, 21–47. [Google Scholar] [CrossRef] [Green Version]
- Ruberti, I.; Sessa, G.; Ciolfi, A.; Possenti, M.; Carabelli, M.; Morelli, G. Plant adaptation to dynamically changing environment: The shade avoidance response. Biotechnol. Adv. 2012, 30, 1047–1058. [Google Scholar] [CrossRef]
- McClung, C.R. Plant circadian rhythms. Plant Cell 2006, 18, 792–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pankin, A.; Campoli, C.; Dong, X.; Kilian, B.; Sharma, R.; Himmelbach, A.; Saini, R.; Davis, S.J.; Stein, N.; Schneeberger, K. Mapping-by-sequencing identifies HvPHYTOCHROME C as a candidate gene for the early maturity 5 locus modulating the circadian clock and photoperiodic flowering in barley. Genetics 2014, 198, 383–396. [Google Scholar] [CrossRef] [Green Version]
- Somers, D.E.; Devlin, P.F.; Kay, S.A. Phytochromes and cryptochromes in the entrainment of the Arabidopsis circadian clock. Science 1998, 282, 1488–1490. [Google Scholar] [CrossRef] [Green Version]
- Suárez-López, P.; Wheatley, K.; Robson, F.; Onouchi, H.; Valverde, F.; Coupland, G. CONSTANS mediates between the circadian clock and the control of flowering in Arabidopsis. Nature 2001, 410, 1116–1120. [Google Scholar] [CrossRef]
- Trevaskis, B.; Hemming, M.N.; Dennis, E.S.; Peacock, W.J. The molecular basis of vernalization-induced flowering in cereals. Trends Plant Sci. 2007, 12, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Szűcs, P.; Karsai, I.; Von Zitzewitz, J.; Mészáros, K.; Cooper, L.; Gu, Y.; Chen, T.; Hayes, P.; Skinner, J. Positional relationships between photoperiod response QTL and photoreceptor and vernalization genes in barley. Theor. Appl. Genet. 2006, 112, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.B.; Zhang, G.P.; Dominy, P. Four barley genotypes respond differently to cadmium: Lipid peroxidation and activities of antioxidant capacity. Environ. Exp. Bot. 2003, 50, 67–78. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Jiang, Y.N.; He, Z.Y.; Ma, M. Cadmium accumulation and oxidative stress burst in garlic (Allium sativum). J. Plant Physiol. 2005, 162, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C T method. Nat. Protoc. 2008, 3, 1101. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Hotta, Y.; Tanaka, T.; Takaoka, H.; Takeuchi, Y.; Konnai, M. New physiological effects of 5-aminolevulinic acid in plants: The increase of photosynthesis, chlorophyll content, and plant growth. Biosci. Biotechnol. Biochem. 1997, 61, 2025–2028. [Google Scholar] [CrossRef]
- Adil, M.F.; Sehar, S.; Han, Z.; Lwalaba, L.J.; Jillani, G.; Zeng, F.; Chen, Z.; Shamsi, I.H. Zinc alleviates cadmium toxicity by modulating photosynthesis, ROS homeostasis, and cation flux kinetics in rice. Environ. Pollut. 2020, 265 Pt B, 115979. [Google Scholar] [CrossRef]
- Mitra, R.; Bhatia, C.R. Bioenergetic cost of heat tolerance in wheat crop. Curr. Sci. 2008, 54, 1049–1053. [Google Scholar]
- Koini, M.A.; Alvey, L.; Allen, T.; Tilley, C.A.; Harberd, N.P.; Whitelam, G.C.; Franklin, K.A. High temperature-mediated adaptations in plant architecture require the bHLH transcription factor PIF4. Curr. Biol. 2009, 19, 408–413. [Google Scholar] [CrossRef] [Green Version]
- Quint, M.; Delker, C.; Franklin, K.A.; Wigge, P.A.; Halliday, K.J.; Zanten, M.V. Molecular and genetic control of plant thermomorphogenesis. Nat. Plants 2016, 2, 15190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahar, N.; Sultana, R.; Gurley, E.S.; Hossain, M.J.; Luby, S.P. Date palm sap collection: Exploring opportunities to prevent Nipah transmission. EcoHealth 2010, 7, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Mishra, S.N. Putrescine alleviation of growth in salt stressed Brassica juncea by inducing antioxidative defense system. J. Plant Physiol. 2005, 162, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Kolupaev, Y.; Ryabchun, N.; Vayner, A.; Yastreb, T.; Oboznyi, A. Antioxidant enzyme activity and osmolyte content in winter cereal seedlings under hardening and cryostress. Russ. J. Plant Physiol. 2015, 62, 499–506. [Google Scholar] [CrossRef]
- Baek, K.H.; Skinner, D.Z. Production of reactive oxygen species by freezing stress and the protective roles of antioxidant enzymes in plants. J. Agric. Chem. Environ. 2012, 1, 34–40. [Google Scholar] [CrossRef]
- Baek, K.H.; Skinner, D.Z. Differential expression of manganese superoxide dismutase sequence variants in near isogenic lines of wheat during cold acclimation. Plant Cell Rep. 2006, 25, 223–230. [Google Scholar] [CrossRef]
- Lukatkin, A.S. Contribution of oxidative stress to the development of cold-induced damage to leaves of chilling-sensitive plants: 1. Reactive oxygen species formation during plant chilling. Russ. J. Plant Physiol. 2002, 49, 622–627. [Google Scholar] [CrossRef]
- Dai, F.; Huang, Y.; Zhou, M.; Zhang, G.P. The influence of cold acclimation on antioxidative enzymes and antioxidants in sensitive and tolerant barley cultivars. Biol. Plant. 2009, 53, 257–262. [Google Scholar] [CrossRef]
- Janmohammadi, M.; Enayati, V.; Sabaghnia, N. Impact of cold acclimation, de-acclimation and re-acclimation on carbohydrate content and antioxidant enzyme activities in spring and winter wheat. Icel. Agric. Sci. 2021, 25, 3–11. [Google Scholar]
- Oquist, G.; Hurry, V.M.; Huner, N.P.A. Low-temperature effects on photosynthesis and correlation with freezing tolerance in spring and winter cultivars of wheat and rye. Plant Physiol. 1993, 101, 245–250. [Google Scholar] [CrossRef] [Green Version]
- Delucia, E.H. The effect of freezing nights on photosynthesis, stomatal conductance, and internal CO2 concentration in seedlings of Engelmann spruce (Picea engelmannii Parry). Plant Cell Environ. 1987, 10, 333–338. [Google Scholar] [CrossRef]
- Zhu, X.G.; Long, S.P.; Ort, D.R. Improving photosynthetic efficiency for greater yield. Annu. Rev. Plant Biol. 2010, 61, 235–261. [Google Scholar] [CrossRef] [Green Version]
- Nishida, H.; Yoshida, T.; Kawakami, K.; Fujita, M.; Long, B.; Akashi, Y.; Laurie, D.A.; Kato, K. Structural variation in the 5′ upstream region of photoperiod-insensitive alleles Ppd-A1a and Ppd-B1a identified in hexaploid wheat (Triticum aestivum L.), and their effect on heading time. Mol. Breed. 2013, 31, 27–37. [Google Scholar] [CrossRef]
- Hemming, M.N.; Fieg, S.; Peacock, W.J.; Dennis, E.S.; Trevaskis, B. Regions associated with repression of the barley (Hordeum vulgare) VERNALIZATION1 gene are not required for cold induction. Mol. Genet. Genom. 2009, 282, 107–117. [Google Scholar] [CrossRef]
- Wilhelm, E.P.; Turner, A.S.; Laurie, D.A. Photoperiod insensitive Ppd-A1a mutations in tetraploid wheat (Triticum durum Desf.). Theor. Appl. Genet. 2009, 118, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Adrian, J.; Farrona, S.; Reimer, J.J.; Albani, M.C.; Coupland, G.; Turck, F. cis-Regulatory elements and chromatin state coordinately control temporal and spatial expression of FLOWERING LOCUS T in Arabidopsis. Plant Cell 2010, 22, 1425–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flood, R.G.; Halloran, G. Genetics and physiology of vernalization response in wheat. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 1986; pp. 87–125. [Google Scholar]
- D’Ottone Di Lorenzo, F. Calibración y Validación del Modelo CropSyst para un Cultivar de Trigo para las Condiciones de Producción del Litoral-Oeste de Uruguay. Diploma Thesis, Universidad de la República, Montevideo, Uruguay, 2011. [Google Scholar]
- Thomas, B.; Vince-Prue, D. Photoperiodism in Plants; Academic Press: New York, NY, USA, 1997; pp. 1–26. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | GenBank ID | Primer Pair Sequence (5′ to 3′) | Length (bp) | PCR Product (bp) |
|---|---|---|---|---|
| HvActin | AY145451 | AGGCCCCTTTGAACCCAAAA | 20 | 88 |
| ATAGCGACGTACATGGCAGG | 20 | |||
| HvVRN1 | AY750995.1 | CTTCACCAACAACTTGGGCG | 20 | 124 |
| CGTGGTGCTGGTGACAGTAT | 20 | |||
| HvPHY-C | ABB13327.1 | AGCCAGAAGCTCGTGAAGAC | 20 | 143 |
| TTCGACCACTTGCTCGTC | 18 | |||
| HvPpd-H1 | ACL78688.1 | TGAAGGTCATGCTGCCAACT | 20 | 149 |
| GCTCCCGTTTCATGTGCATC | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rasheed, A.; Feng, Q.; Adil, M.F.; Ahsan, M.; Han, Z.; Zeng, F.; Shamsi, I.H. Comparative Study on the Physio-Biochemical Responses of Spring and Winter Barley Genotypes under Vernalized and Greenhouse Conditions. Agronomy 2022, 12, 339. https://doi.org/10.3390/agronomy12020339
Rasheed A, Feng Q, Adil MF, Ahsan M, Han Z, Zeng F, Shamsi IH. Comparative Study on the Physio-Biochemical Responses of Spring and Winter Barley Genotypes under Vernalized and Greenhouse Conditions. Agronomy. 2022; 12(2):339. https://doi.org/10.3390/agronomy12020339
Chicago/Turabian StyleRasheed, Adeela, Qidong Feng, Muhammad Faheem Adil, Muhammad Ahsan, Zhigang Han, Fanrong Zeng, and Imran Haider Shamsi. 2022. "Comparative Study on the Physio-Biochemical Responses of Spring and Winter Barley Genotypes under Vernalized and Greenhouse Conditions" Agronomy 12, no. 2: 339. https://doi.org/10.3390/agronomy12020339
APA StyleRasheed, A., Feng, Q., Adil, M. F., Ahsan, M., Han, Z., Zeng, F., & Shamsi, I. H. (2022). Comparative Study on the Physio-Biochemical Responses of Spring and Winter Barley Genotypes under Vernalized and Greenhouse Conditions. Agronomy, 12(2), 339. https://doi.org/10.3390/agronomy12020339