
Different Green Manures (Vicia villosa and Brassica juncea) Construct Different Fungal Structures, Including Plant-Growth-Promoting Effects, after Incorporation into the Soil


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Fungi
2.2. DNA Extraction and PCR Amplification for Isolated Fungi
2.3. Screening of Plant-Growth-Promoting Fungi (PGPF)
2.3.1. Microcosm Experiment under Axenic Conditions
2.3.2. Microcosm Experiment under Non-Axenic Conditions
2.4. Characterization of Selected Strains
2.4.1. Plant-Growth-Promoting Traits of Isolated Fungi
2.4.2. Extracellular Enzyme Activities of Isolates
2.5. Statistical Analysis
3. Results
3.1. Isolation and Identification of Fungi
3.2. Screening of Plant Growth Promoting Fungi (PGPF)
3.2.1. Microcosm Experiment under Axenic Conditions
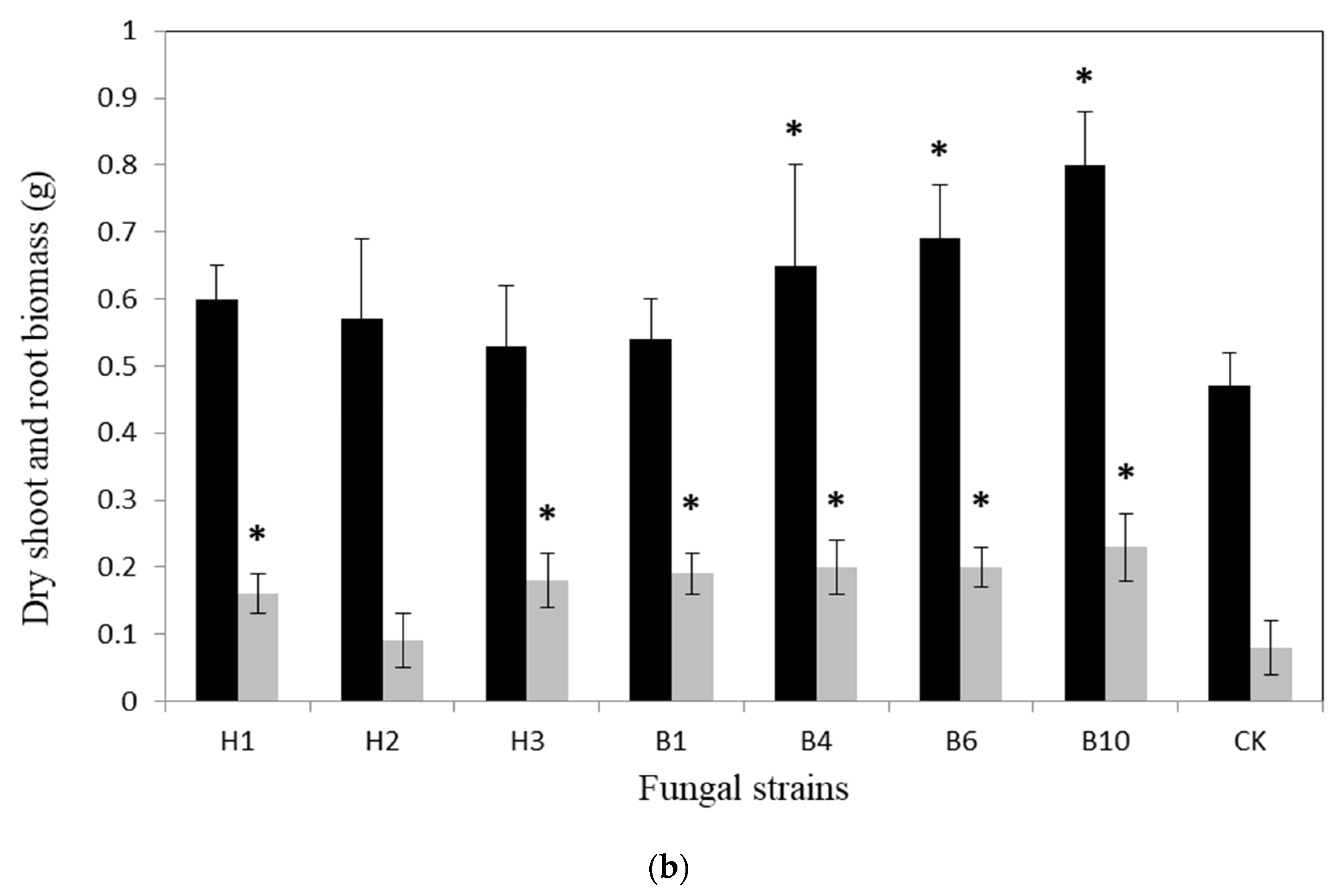
3.2.2. Microcosm Experiment under Non-Axenic Conditions
3.3. Characterization of Selected Strains
3.3.1. Plant-Growth-Promoting Traits
3.3.2. Extracellular Enzyme Activities of Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mahmood, A.; Iguchi, R.; Kataoka, R. Multifunctional food waste fertilizer having the capability of Fusarium-growth inhibition and phosphate solubility: A new horizon of food waste recycle using microorganisms. Waste Manag. 2019, 94, 77–84. [Google Scholar] [CrossRef]
- Babu, A.G.; Kim, S.W.; Yadav, D.R.; Hyum, U.; Adhikari, M.; Lee, Y.S. Penicillium menonorum: A Novel Fungus to Promote Growth and Nutrient Management in Cucumber Plants. Mycobiology 2015, 43, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Majeed, A.; Abbasi, M.K.; Hameed, S.; Imran, A.; Rahim, N. Isolation and characterization of plant growth-promoting rhizobacteria from wheat rhizosphere and their effect on plant growth promotion. Front. Microbiol. 2015, 6, 198. [Google Scholar] [CrossRef] [Green Version]
- Asghar, W.; Kondo, S.; Iguchi, R.; Mahmood, A.; Kataoka, R. Agricultural Utilization of Unused Resources: Liquid Food Waste Material as a New Source of Plant Growth-Promoting Microbes. Agronomy 2020, 10, 954. [Google Scholar] [CrossRef]
- Sinha, R.K.; Herat, S.; Bharambe, G.; Patil, S.; Bapat, P.; Chauhan, K.; Valani, D. Human waste-apotential resource: Converting trash into treasure by embracing the 5 r’s philosophy for safe and sustainable waste management. Environ. Res. J. 2009, 3, 143. [Google Scholar]
- Bowles, T.M.; Acosta-Martínez, V.; Calderón, F.; Jackson, L.E. Soil enzyme activities, microbial communities, and carbon and nitrogen availability in organic agroecosystems across an intensively-managed agricultural landscape. Soil Biol. Biochem. 2014, 68, 252–262. [Google Scholar] [CrossRef]
- Hai-Ming, T.; Xiao-Ping, X.; Wen-Guang, T.; Ye-Chun, L.; Ke, W.; Guang-Li, Y. Effects of winter cover crops residue returning on soil enzyme activities and soil microbial community in double-cropping rice fields. PLoS ONE 2014, 9, e100443. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, R.; Gao, J.; Wang, X.; Fan, F.; Ma, X.; Yin, H.; Zhang, C.; Feng, K.; Deng, Y. Thirty-one years of rice-rice-green manure rotations shape the rhizosphere microbial community and enrich beneficial bacteria. Soil Biol. Biochem. 2017, 104, 208–217. [Google Scholar] [CrossRef]
- Brennan, E.B.; Acosta-Martinez, V. Cover cropping frequency is the main driver of soil microbial changes during six years of organic vegetable production. Soil Biol. Biochem. 2017, 109, 188–204. [Google Scholar] [CrossRef]
- Chavarria, D.N.; Verdenelli, R.A.; Serri, D.L.; Restovich, S.B.; Andriulo, A.E.; Meriles, J.M.; Vargas-Gil, S. Effect of cover crops on microbial community structure and related enzyme activities and macronutrient availability. Eur. J. Soil Biol. 2016, 76, 74–82. [Google Scholar] [CrossRef]
- Asghar, W.; Kataoka, R. Green manure incorporation accelerates enzyme activity, plant growth, and changes in the fungal community of soil. Arch. Microbiol. 2022, 204, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mbuthia, L.W.; Acosta-Martínez, V.; DeBruyn, J.; Schaeffer, S.; Tyler, D.; Odoi, E.; Mpheshea, M.; Walker, F.; Eash, N. Long term tillage, cover crop, and fertilization effects on microbial community structure, activity: Implications for soil quality. Soil Biol. Biochem. 2015, 89, 24–34. [Google Scholar] [CrossRef]
- Khan, M.I.; Gwon, H.S.; Alam, M.A.; Song, H.J.; Das, S.; Kim, P.J. Short term effects of different green manure amendments on the composition of main microbial groups and microbial activity of a submerged rice cropping system. Appl. Soil Ecol. 2020, 147, 103400. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Van Der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Wang, J.; Li, R.; Zhang, H.; Wei, G.; Li, Z. Beneficial bacteria activate nutrients and promote wheat growth under conditions of reduced fertilizer application. BMC Microbiol. 2020, 20, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turbat, A.; Rakk, D.; Vigneshwari, A.; Kocsubé, S.; Thu, H.; Szepesi, Á.; Bakacsy, L.; Škrbić, B.D.; Jigjiddorj, E.-A.; Vágvölgyi, C. Characterization of the Plant Growth-Promoting Activities of Endophytic Fungi Isolated from Sophora flavescens. Microorganisms 2020, 8, 683. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Rahman, M.A.; Guo, J.; Kang, Z.; Babar, M.A. Comparative physiological and metabolic analysis reveals a complex mechanism involved in drought tolerance in chickpea (Cicer arietinum L.) induced by PGPR and PGRs. Sci. Rep. 2019, 9, 1–19. [Google Scholar] [CrossRef]
- Hossain, M.M.; Sultana, F. Application and mechanisms of plant growth promoting fungi (PGPF) for phytostimulation. Org. Agric. 2020, 1–31. [Google Scholar] [CrossRef]
- Naziya, B.; Murali, M.; Amruthesh, K.N. Plant growth-promoting fungi (PGPF) instigate plant growth and induce disease resistance in Capsicum annuum L. upon infection with Colletotrichum capsici (Syd.) Butler & Bisby. Biomolecules 2020, 10, 41. [Google Scholar]
- Mahdi, I.; Fahsi, N.; Hafidi, M.; Allaoui, A.; Biskri, L. Plant Growth Enhancement using Rhizospheric Halotolerant Phosphate Solubilizing Bacterium Bacillus licheniformis QA1 and Enterobacter asburiae QF11 Isolated from Chenopodium quinoa Willd. Microorganisms 2020, 8, 948. [Google Scholar] [CrossRef]
- Galeano, R.M.S.; Franco, D.G.; Chaves, P.O.; Giannesi, G.C.; Masui, D.C.; Ruller, R.; Corrêa, B.O.; da Silva Brasil, M.; Zanoelo, F.F. Plant growth promoting potential of endophytic Aspergillus niger 9-p isolated from native forage grass in Pantanal of Nhecolândia region, Brazil. Rhizosphere 2021, 18, 100332. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, F.-S.; Wu, X.-Q.; Luan, F.-G.; Zhang, L.-P.; Fang, X.-M.; Wan, S.-Z.; Hu, X.-F.; Ye, J.-R. Isolation and characterization of two phosphate-solubilizing fungi from rhizosphere soil of moso bamboo and their functional capacities when exposed to different phosphorus sources and pH environments. PLoS ONE 2018, 13, e0199625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, J.P. Use of acid, rose bengal, and streptomycin in the plate method for estimating soil fungi. Soil Sci. 1950, 69, 215–232. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Pikovskaya, R. Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Mikrobiologiya 1948, 17, 362–370. [Google Scholar]
- Schwyn, B.; Neilands, J. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Pérez-Miranda, S.; Cabirol, N.; George-Téllez, R.; Zamudio-Rivera, L.; Fernández, F. O-CAS, a fast and universal method for siderophore detection. J. Microbiol. Methods 2007, 70, 127–131. [Google Scholar] [CrossRef]
- Acuña, J.J.; Jorquera, M.A.; Martínez, O.A.; Menezes-Blackburn, D.; Fernández, M.T.; Marschner, P.; Greiner, R.; Mora, M. Indole acetic acid and phytase activity produced by rhizosphere bacilli as affected by pH and metals. J. Soil Sci. Plant Nutr. 2011, 11, 1–12. [Google Scholar]
- Shah, S.; Shrestha, R.; Maharjan, S.; Selosse, M.-A.; Pant, B. Isolation and characterization of plant growth-promoting endophytic fungi from the roots of Dendrobium moniliforme. Plants 2019, 8, 5. [Google Scholar] [CrossRef] [Green Version]
- Colonia, B.S.O.; Junior, A.C. Screening and detection of extracellular cellulases (endo-and exo-glucanases) secreted by filamentous fungi isolated from soils using rapid tests with chromogenic dyes. Afr. J. Biotechnol. 2014, 13, 52. [Google Scholar]
- Asghar, W.; Kataoka, R. Effect of co-application of Trichoderma spp. with organic composts on plant growth enhancement, soil enzymes and fungal community in soil. Arch. Microbiol. 2021, 203, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Y.; Li, J.; Bai, S.; Tian, T. Fungal community composition and diversity in the rhizosphere soils of Argentina (syn. Potentilla) anserina, on the Qinghai Plateau. Fungal Ecol. 2021, 54, 101107. [Google Scholar] [CrossRef]
- Qarni, A.; Billah, M.; Hussain, K.; Shah, S.H.; Ahmed, W.; Alam, S.; Sheikh, A.A.; Jafri, L.; Munir, A.; Malik, K.M. Isolation and Characterization of Phosphate Solubilizing Microbes from Rock Phosphate Mines and their Potential Effect for Sustainable Agriculture. Sustainability 2021, 13, 2151. [Google Scholar] [CrossRef]
- Khan, A.; Singh, P.; Srivastava, A. Synthesis, nature and utility of universal iron chelator–Siderophore: A review. Microbiol. Res. 2018, 212, 103–111. [Google Scholar] [CrossRef]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus 2013, 2, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Gontia-Mishra, I.; Sapre, S.; Sharma, A.; Tiwari, S. Alleviation of mercury toxicity in wheat by the interaction of mercury-tolerant plant growth-promoting rhizobacteria. J. Plant Growth Regul. 2016, 35, 1000–1012. [Google Scholar] [CrossRef]
- Purahong, W.; Wubet, T.; Lentendu, G.; Schloter, M.; Pecyna, M.J.; Kapturska, D.; Hofrichter, M.; Krüger, D.; Buscot, F. Life in leaf litter: Novel insights into community dynamics of bacteria and fungi during litter decomposition. Mol. Ecol. 2016, 25, 4059–4074. [Google Scholar] [CrossRef]
- Carson, C.M.; Jumpponen, A.; Blair, J.M.; Zeglin, L.H. Soil fungal community changes in response to long-term fire cessation and N fertilization in tallgrass prairie. Fungal Ecol. 2019, 41, 45–55. [Google Scholar] [CrossRef]
- Liu, J.; Sui, Y.; Yu, Z.; Shi, Y.; Chu, H.; Jin, J.; Liu, X.; Wang, G. Soil carbon content drives the biogeographical distribution of fungal communities in the black soil zone of northeast China. Soil Biol. Biochem. 2015, 83, 29–39. [Google Scholar] [CrossRef]
- Böhme, L.; Böhme, F. Soil microbiological and biochemical properties affected by plant growth and different long-term fertilisation. Eur. J. Soil Biol. 2006, 42, 1–12. [Google Scholar] [CrossRef]
- Tang, H.-M.; Xiao, X.-P.; Tang, W.-G.; Li, C.; Wang, K.; Cheng, K.-K.; Sun, G. Dynamic change of soil enzyme activities and soil microbe during rice main growth stages in different long-term fertilizer regimes. J. Pure Appl. Microbiol. 2017, 11, 649–661. [Google Scholar] [CrossRef]
- Hanschen, F.S.; Winkelmann, T. Biofumigation for fighting replant disease-A Review. Agronomy 2020, 10, 425. [Google Scholar] [CrossRef] [Green Version]
- Intanon, S.; Hulting, A.G.; Myrold, D.D.; Mallory-Smith, C.A. Short-term effects of soil amendment with meadowfoam seed meal on soil microbial composition and function. Appl. Soil Ecol. 2015, 89, 85–92. [Google Scholar] [CrossRef]
- Brown, P.D. Control of soil-borne plant pests using glucosinolate-containing plants. Adv. Agron. 1997, 61, 168–231. [Google Scholar]
- Hollister, E.B.; Hu, P.; Wang, A.S.; Hons, F.M.; Gentry, T.J. Differential impacts of brassicaceous and nonbrassicaceous oilseed meals on soil bacterial and fungal communities. FEMS Microbiol. Ecol. 2013, 83, 632–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plaszkó, T.; Szűcs, Z.; Vasas, G.; Gonda, S. Effects of glucosinolate-derived isothiocyanates on fungi: A comprehensive review on direct effects, mechanisms, structure-activity relationship data and possible agricultural applications. J. Fungi 2021, 7, 539. [Google Scholar] [CrossRef]
- Sarwar, M.; Kirkegaard, J.; Wong, P.; Desmarchelier, J. Biofumigation potential of brassicas. Plant Soil 1998, 201, 103–112. [Google Scholar] [CrossRef]
- Yim, B.; Hanschen, F.S.; Wrede, A.; Nitt, H.; Schreiner, M.; Smalla, K.; Winkelmann, T. Effects of biofumigation using Brassica juncea and Raphanus sativus in comparison to disinfection using Basamid on apple plant growth and soil microbial communities at three field sites with replant disease. Plant Soil 2016, 406, 389–408. [Google Scholar] [CrossRef]



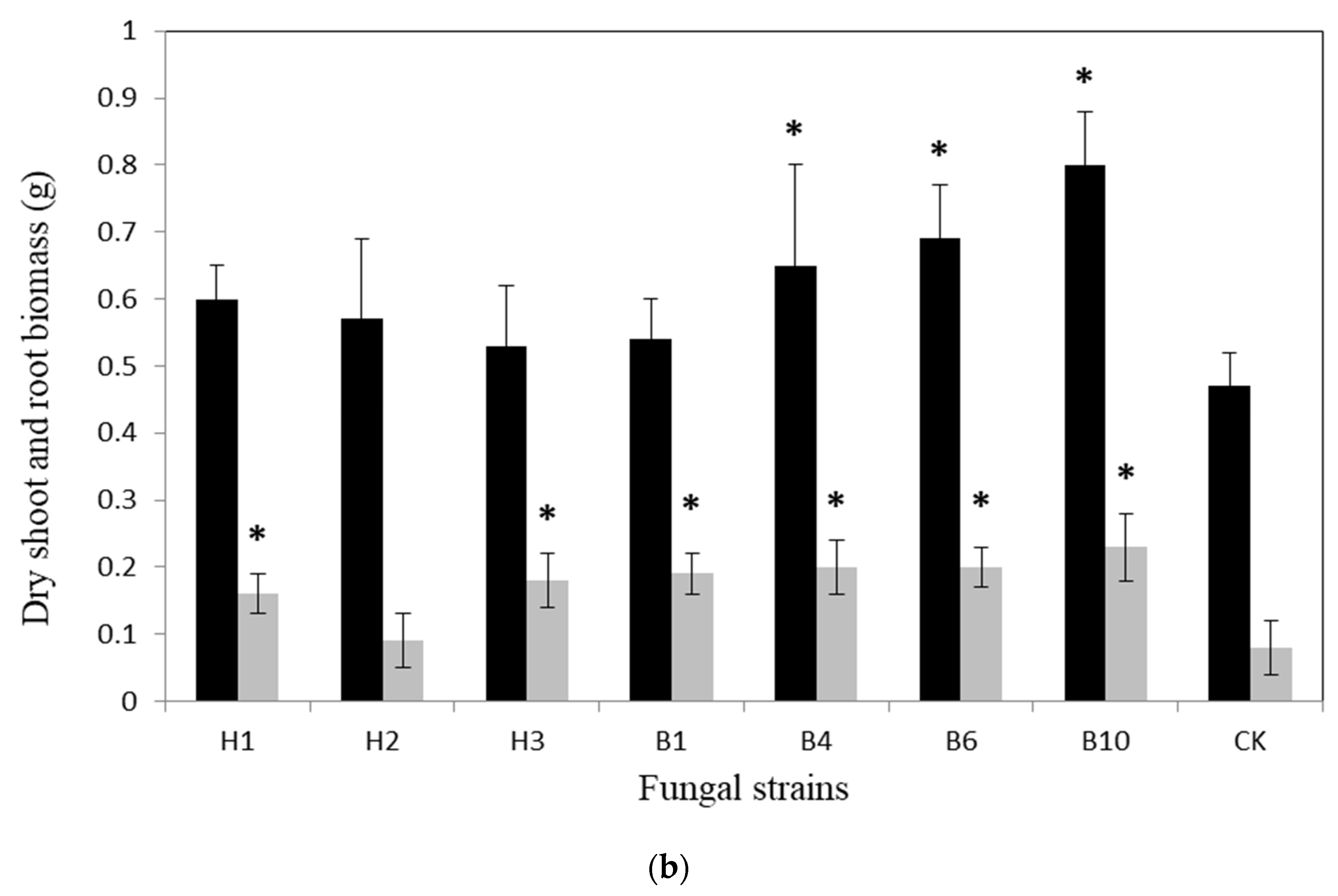

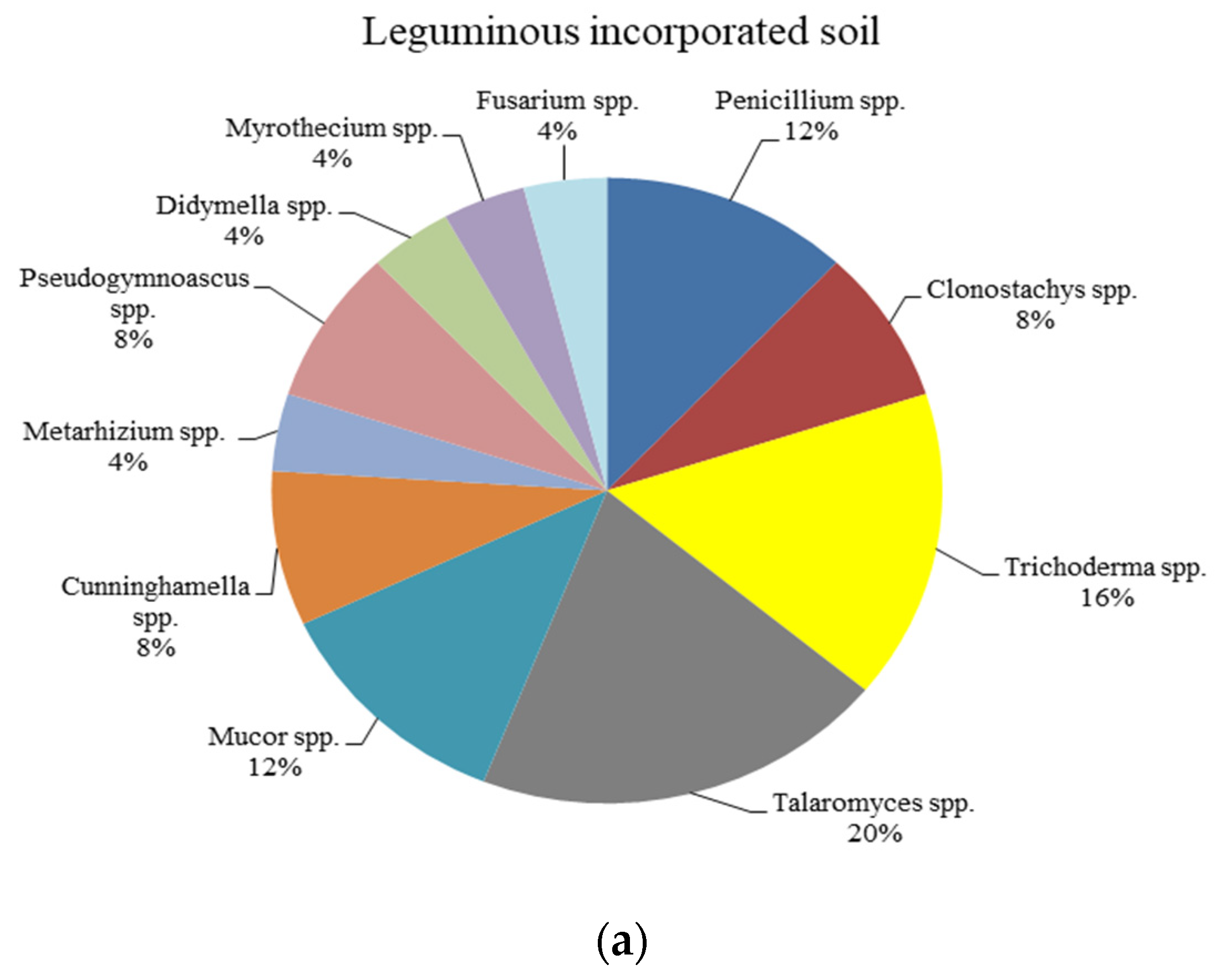{kind=link}
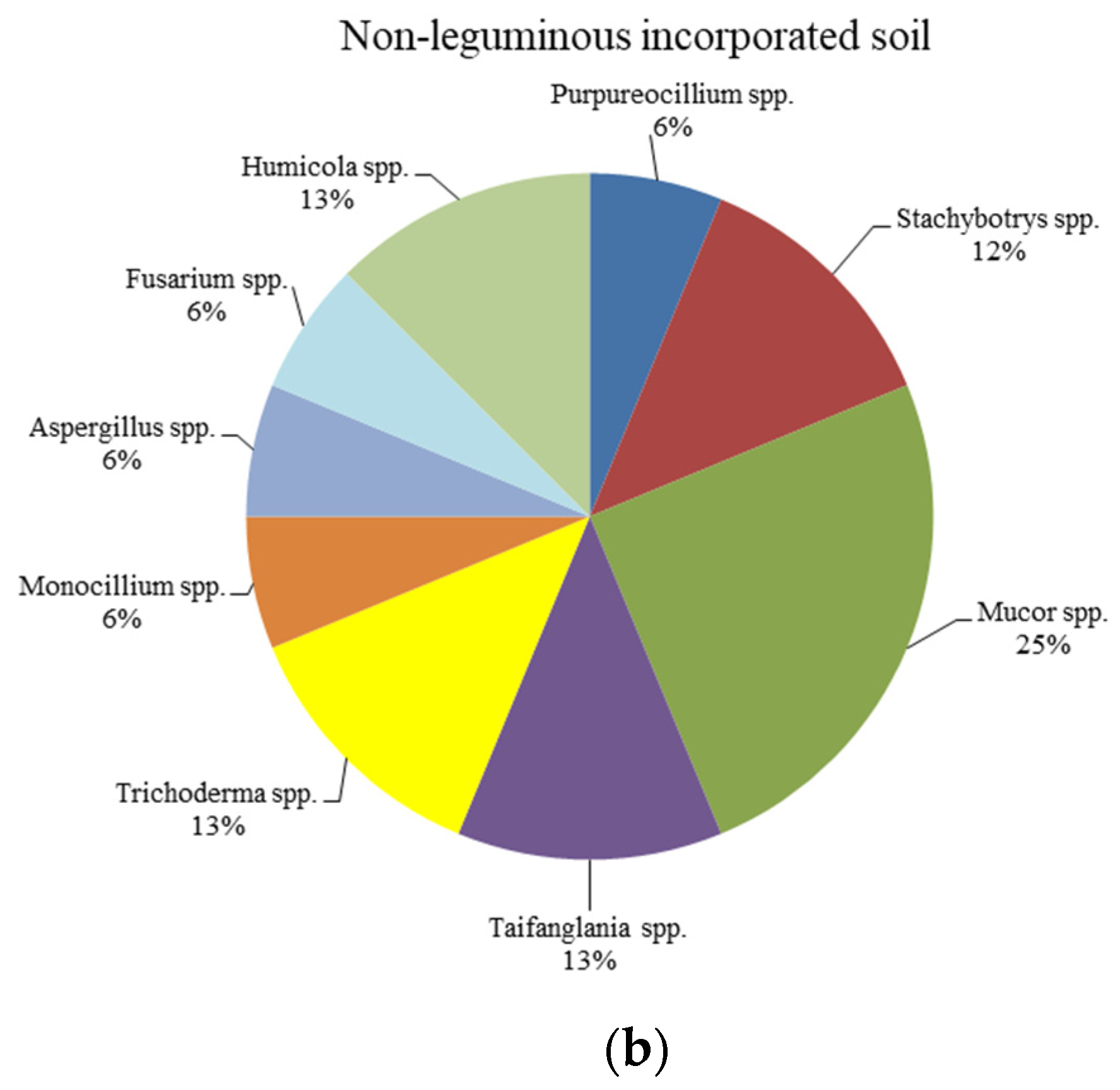{kind=link}
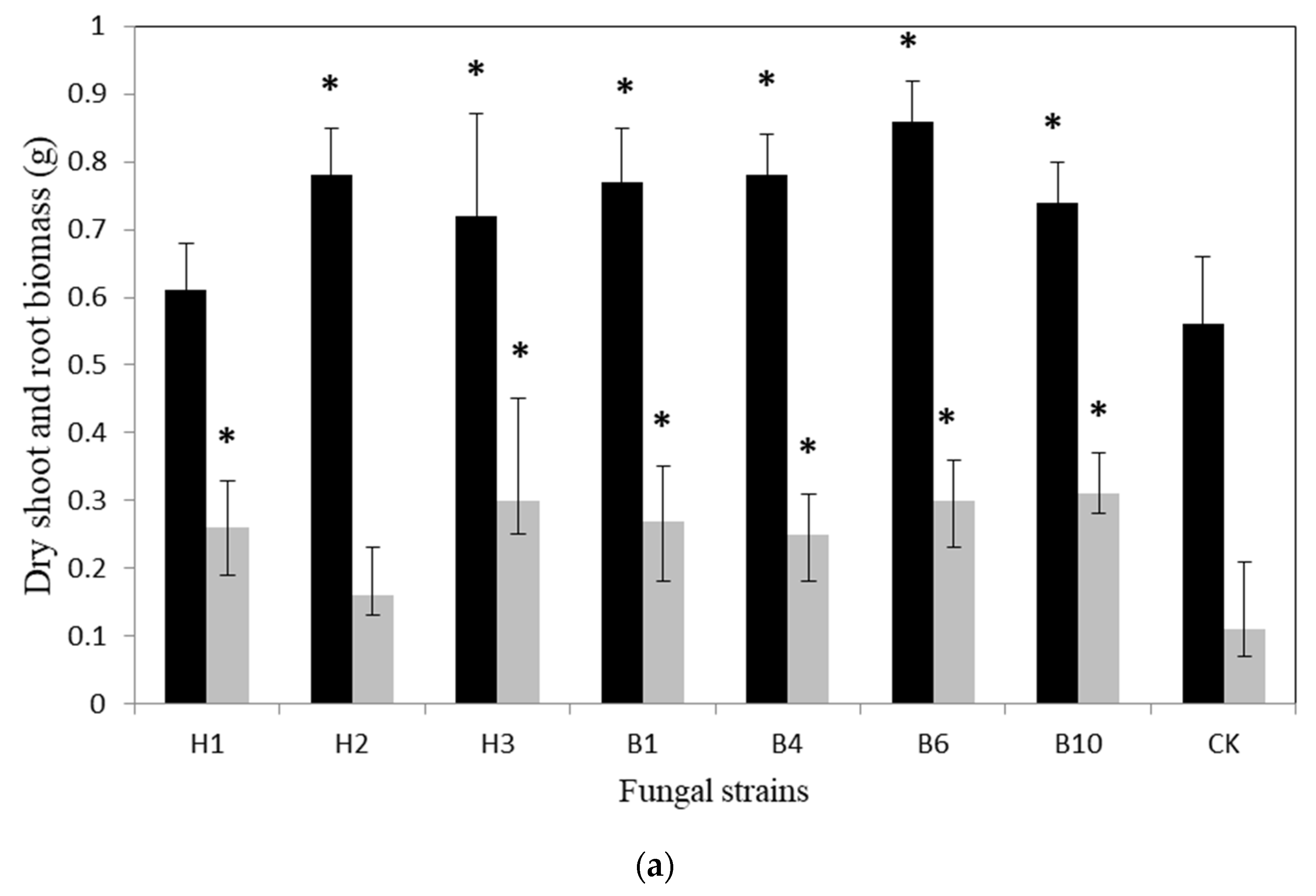{kind=link}
{kind=link}
| (a) | |||
|---|---|---|---|
| Leguminous Soil | Blast Closest Match | Identity | Accession Number |
| H1 | Penicillium brasilianum (KY701767) | 421/424 (99%) | LC638643 |
| H2 | Clonostachys rosea (KY810798) | 495/495 (100%) | LC638644 |
| H3 | Trichoderma harzianum (KY552264) | 555/555 (100%) | LC638645 |
| H4 | Talaromyces pinophilus (MF68681) | 515/516 (99%) | LC638646 |
| H5 | Talaromyces adpressus (KY425714) | 498/499 (99%) | LC638647 |
| H6 | Trichoderma tomentosum (KC576650) | 539/540 (99%) | LC638648 |
| H7 | Trichoderma tomentosum (FJ412070) | 557/558 (99%) | LC638649 |
| H8 | Penicillium brasilianum (MH857470) | 513/513 (100%) | LC638650 |
| H9 | Penicillium brasilianum (MH857470) | 508/508 (100%) | LC638651 |
| H10 | Trichoderma tomentosum (FJ412070) | 547/548 (99%) | LC638652 |
| H11 | Colletotrichum rosea (MK075014). | 494/494 (100%) | LC638653 |
| H12 | Mucor circinelloides (KR709187) | 563/563 (100%) | LC638654 |
| H13 | Mucor circinelloides (MN105451) | 574/574 (100%) | LC638655 |
| H14 | Cunninghamella bertholletiae (JN205877) | 691/696 (99%) | LC638656 |
| H15 | Cunninghamella bertholletiae (JN205877) | 689/700 (985) | LC638656 |
| H16 | Mucor circinelloides (MW577264) | 689/700 (98%) | LC638658 |
| H17 | Metarhizium marquandii (MK952275) | 579/579 (100%) | LC638659 |
| H18 | Pseudogymnoascus pannorum (MH864862) | 505/505 (100%) | LC638660 |
| H19 | Fusarium oxysporum (MW563929) | 487/487 (100%) | LC638661 |
| H20 | Didymella americana (KY099740) | 478/478 (100%) | LC638662 |
| H21 | Talaromyces sp. (MW670552) | 479/479 (100%) | LC638663 |
| H22 | Talaromyces pinophilus (MW363693) | 502/502 (100%) | LC638664 |
| H23 | Myrothecium sp. (MK775959) | 513/513 (100%) | LC638665 |
| H24 | Pseudogymnoascus pannorum (MT089975) | 491/495 (99%) | LC638666 |
| H25 | Talaromyces pinophilus (MT730086) | 503/503 (100%) | LC638667 |
| (b) | |||
| Non-Leguminous Soil | BLAST Closest Match | Identity | Accession Number |
| B1 | Purpureocillium lilacinum (MW113495) | 541/541 (100%) | LC638626 |
| B2 | Stachybotrys sp. (KC305351) | 501/501 (100%) | LC638627 |
| B3 | Mucor sp. (MW789351) | 568/568 (100%) | LC638628 |
| B4 | Taifanglania sp. (KP143091) | 468/473 (98%) | LC638629 |
| B5 | Mucor circinelloides (JN226939) | 571/572 (99%) | LC638630 |
| B6 | Trichoderma harzianum (MK594269) | 531/531 (100%) | LC638631 |
| B7 | Mucor circinelloides f. circinelloides (AY243943) | 558/563 (99%) | LC638632 |
| B8 | Humicola fuscoatra (KY310738) | 489/490 (99%) | LC638633 |
| B9 | Trichoderma harzianum (MT090002) | 552/552 (100%) | LC638634 |
| B10 | Aspergillus niger (MW931861) | 517/517 (100%) | LC638635 |
| B11 | Mucor circinelloides (MW763155) | 549/550 (99%) | LC638636 |
| B12 | Fusarium verticillioides (MN088603) | 482/482 (100%) | LC638637 |
| B13 | Monocillium mucidum (MH859551) | 519/519 (100%) | LC638638 |
| B14 | Stachybotrys sansevieriae voucher (KY587783) | 499/500 (99%) | LC638639 |
| B15 | Humicola grisea (KF876826) | 486/486 (100%) | LC638640 |
| B16 | Taifanglania sp. (KP143091) | 491/491 (100%) | LC638641 |
| B17 | Mucor circinelloides (JN226939) | 571/572 (99%) | LC638642 |
| Strain No. | Phosphate Solubilization | Siderophore | IAA | Endoglucanase | β-Glucosidase Units h−1 |
|---|---|---|---|---|---|
| H1 | + | + | − | + | 33.19 ± 1.03 |
| H2 | + | − | − | + | 27.61 ± 1.05 |
| H3 | − | − | − | − | 35.07 ± 1.65 |
| B1 | + | + | − | + | 31.89 ± 0.59 |
| B4 | − | − | − | + | 27.68 ± 2.71 |
| B6 | − | + | − | − | 25.19 ± 1.05 |
| B10 | + | + | − | + | 36.83 ± 1.48 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asghar, W.; Kataoka, R. Different Green Manures (Vicia villosa and Brassica juncea) Construct Different Fungal Structures, Including Plant-Growth-Promoting Effects, after Incorporation into the Soil. Agronomy 2022, 12, 323. https://doi.org/10.3390/agronomy12020323
Asghar W, Kataoka R. Different Green Manures (Vicia villosa and Brassica juncea) Construct Different Fungal Structures, Including Plant-Growth-Promoting Effects, after Incorporation into the Soil. Agronomy. 2022; 12(2):323. https://doi.org/10.3390/agronomy12020323
Chicago/Turabian StyleAsghar, Waleed, and Ryota Kataoka. 2022. "Different Green Manures (Vicia villosa and Brassica juncea) Construct Different Fungal Structures, Including Plant-Growth-Promoting Effects, after Incorporation into the Soil" Agronomy 12, no. 2: 323. https://doi.org/10.3390/agronomy12020323
APA StyleAsghar, W., & Kataoka, R. (2022). Different Green Manures (Vicia villosa and Brassica juncea) Construct Different Fungal Structures, Including Plant-Growth-Promoting Effects, after Incorporation into the Soil. Agronomy, 12(2), 323. https://doi.org/10.3390/agronomy12020323