
Combined Biostimulant Applications of Trichoderma spp. with Fatty Acid Mixtures Improve Biocontrol Activity, Horticultural Crop Yield and Nutritional Quality

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Fatty Acid Mixture Features (FAm)
2.2. FAm Biocontrol Activity In Vitro
2.3. FAm Biocontrol Activity In Vivo
2.4. Compatibility between FAm and Trichoderma spp.
2.5. Biocontrol Assay of FAm + Trichoderma spp. Combinations
2.6. Growth Promotion Assay of Trichoderma spp. + FAm Combinations
2.7. Evaluation of Trichoderma spp. + FAm Combinations on Agronomic Parameters
2.8. Assessment of Trichoderma spp. + FAm Combination Effects on Nutritional Quality of Crops
2.9. Assessment of Trichoderma spp. + FAm Combination Effects on Tomato Plant Metabolism
2.10. Statistical Analysis
3. Results
3.1. In Vitro and In Vivo FAm Biocontrol Activity
3.2. Compatibility between FAm and Trichoderma spp.
3.3. Biocontrol Activity of FAm + Trichoderma spp. Combinations
3.4. Growth Promotion Effect of Trichoderma spp. +FAm Combinations
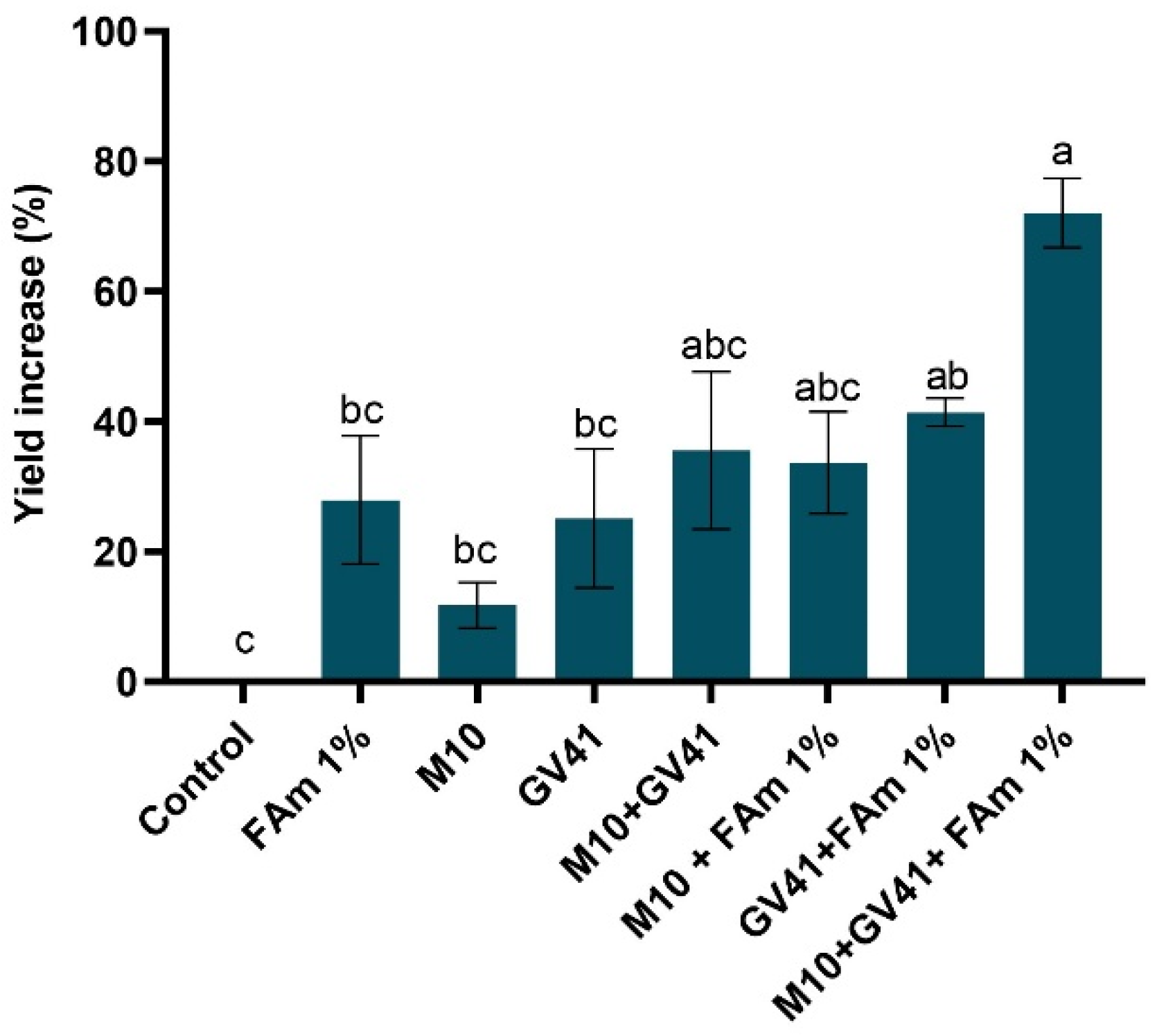
3.5. Effectiveness of Trichoderma spp. + FAm Combinations on Agronomic Parameters
3.6. Effect of Trichoderma spp. + FAm Combinations on Nutritional Quality of Crops
3.7. Effect of Trichoderma spp. + FAm Combinations on Tomato Plant Metabolism
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carillo, P.; Woo, S.L.; Comite, E.; El-Nakhel, C.; Rouphael, Y.; Fusco, G.M.; Borzacchiello, A.; Lanzuise, S.; Vinale, F. Application of Trichoderma harzianum, 6-Pentyl-α-Pyrone and Plant Biopolymer Formulations Modulate Plant Metabolism and Fruit Quality of Plum Tomatoes. Plants 2020, 9, 771. [Google Scholar] [CrossRef]
- Hertel, T.W. The Global Supply and Demand for Agricultural Land in 2050: A Perfect Storm in the Making. Am. J. Agric. Econ. 2011, 93, 259–275. [Google Scholar] [CrossRef]
- Santos, M.S.; Rodrigues, T.F.; Nogueira, M.A.; Hungria, M. The Challenge of Combining High Yields with Environmentally Friendly Bioproducts: A Review on the Compatibility of Pesticides with Microbial Inoculants. Agronomy 2021, 11, 870. [Google Scholar] [CrossRef]
- Tudi, M.; Daniel Ruan, H.; Wang, L.; Lyu, J.; Sadler, R.; Connell, D.; Chu, C.; Phung, D.T. Agriculture Development, Pesticide Application and Its Impact on the Environment. Int. J. Environ. Res. Public Health 2021, 18, 1112. [Google Scholar] [CrossRef]
- Guzmán-Guzmán, P.; Porras-Troncoso, M.D.; Olmedo-Monfil, V.; Herrera-Estrella, A. Trichoderma Species: Versatile Plant Symbionts. Phytopathology 2019, 109, 6–16. [Google Scholar] [CrossRef]
- Comite, E.; El-Nakhel, C.; Rouphael, Y.; Ventorino, V.; Pepe, O.; Borzacchiello, A.; Vinale, F.; Rigano, D.; Staropoli, A.; Lorito, M.; et al. Bioformulations with Beneficial Microbial Consortia, a Bioactive Compound and Plant Biopolymers Modulate Sweet Basil Productivity, Photosynthetic Activity and Metabolites. Pathogens 2021, 10, 870. [Google Scholar] [CrossRef]
- Woo, S.L.; Ruocco, M.; Vinale, F.; Nigro, M.; Marra, R.; Lombardi, N.; Pascale, A.; Lanzuise, S.; Manganiello, G.; Lorito, M. Trichoderma-Based Products and Their Widespread Use in Agriculture. Open Mycol. J. 2014, 8, 71–126. [Google Scholar] [CrossRef]
- Fiorentino, N.; Ventorino, V.; Woo, S.L.; Pepe, O.; de Rosa, A.; Gioia, L.; Romano, I.; Lombardi, N.; Napolitano, M.; Colla, G.; et al. Trichoderma-Based Biostimulants Modulate Rhizosphere Microbial Populations and Improve N Uptake Efficiency, Yield, and Nutritional Quality of Leafy Vegetables. Front. Plant Sci. 2018, 9, 743. [Google Scholar] [CrossRef]
- Schirawski, J.; Perlin, M. Plant—Microbe Interaction 2017—The Good, the Bad and the Diverse. Int. J. Mol. Sci. 2018, 19, 1374. [Google Scholar] [CrossRef]
- Stringlis, I.A.; Yu, K.; Feussner, K.; de Jonge, R.; van Bentum, S.; van Verk, M.C.; Berendsen, R.L.; Bakker, P.A.H.M.; Feussner, I.; Pieterse, C.M.J. MYB72-Dependent Coumarin Exudation Shapes Root Microbiome Assembly to Promote Plant Health. Proc. Natl. Acad. Sci. USA 2018, 115, E5213–E5222. [Google Scholar] [CrossRef] [PubMed]
- Silletti, S.; di Stasio, E.; van Oosten, M.J.; Ventorino, V.; Pepe, O.; Napolitano, M.; Marra, R.; Woo, S.L.; Cirillo, V.; Maggio, A. Biostimulant Activity of Azotobacter Chroococcum and Trichoderma harzianum in Durum Wheat under Water and Nitrogen Deficiency. Agronomy 2021, 11, 380. [Google Scholar] [CrossRef]
- Lombardi, N.; Caira, S.; Troise, A.D.; Scaloni, A.; Vitaglione, P.; Vinale, F.; Marra, R.; Salzano, A.M.; Lorito, M.; Woo, S.L. Trichoderma Applications on Strawberry Plants Modulate the Physiological Processes Positively Affecting Fruit Production and Quality. Front. Microbiol. 2020, 11, 1364. [Google Scholar] [CrossRef]
- Rouphael, Y.; Carillo, P.; Colla, G.; Fiorentino, N.; Sabatino, L.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cirillo, V.; Shabani, E.; et al. Appraisal of Combined Applications of Trichoderma Virens and a Biopolymer-Based Biostimulant on Lettuce Agronomical, Physiological, and Qualitative Properties under Variable N Regimes. Agronomy 2020, 10, 196. [Google Scholar] [CrossRef]
- Manganiello, G.; Nicastro, N.; Caputo, M.; Zaccardelli, M.; Cardi, T.; Pane, C. Functional Hyperspectral Imaging by High-Related Vegetation Indices to Track the Wide-Spectrum Trichoderma Biocontrol Activity Against Soil-Borne Diseases of Baby-Leaf Vegetables. Front. Plant Sci. 2021, 12, 630059. [Google Scholar] [CrossRef]
- Mishra, J.; Arora, N.K. Bioformulations for Plant Growth Promotion and Combating Phytopathogens: A Sustainable Approach. In Bioformulations: For Sustainable Agriculture; Springer: New Delhi, India, 2016; pp. 3–33. [Google Scholar]
- Aamir, M.; Rai, K.K.; Zehra, A.; Dubey, M.K.; Kumar, S.; Shukla, V.; Upadhyay, R.S. Microbial Bioformulation-Based Plant Biostimulants: A Plausible Approach toward next Generation of Sustainable Agriculture. In Microbial Endophytes; Elsevier: Amsterdam, The Netherlands, 2020; pp. 195–225. [Google Scholar]
- Castiglione, A.M.; Mannino, G.; Contartese, V.; Bertea, C.M.; Ertani, A. Microbial Biostimulants as Response to Modern Agriculture Needs: Composition, Role and Application of These Innovative Products. Plants 2021, 10, 1533. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; del Mar Alguacil, M.; Pascual, J.A.; van Wees, S.C.M. Phytohormone Profiles Induced by Trichoderma Isolates Correspond with Their Biocontrol and Plant Growth-Promoting Activity on Melon Plants. J. Chem. Ecol. 2014, 40, 804–815. [Google Scholar] [CrossRef] [PubMed]
- Gravel, V.; Antoun, H.; Tweddell, R.J. Growth Stimulation and Fruit Yield Improvement of Greenhouse Tomato Plants by Inoculation with Pseudomonas Putida or Trichoderma Atroviride: Possible Role of Indole Acetic Acid (IAA). Soil Biol. Biochem. 2007, 39, 1968–1977. [Google Scholar] [CrossRef]
- MacKenzie, A.J.; Starman, T.W.; Windham, M.T. Enhanced Root and Shoot Growth of Chrysanthemum Cuttings Propagated with the Fungus Trichoderma harzianum. HortScience 1995, 30, 496–498. [Google Scholar] [CrossRef]
- Ousley, M.A.; Lynch, J.M.; Whipps, J.M. Potential of Trichoderma Spp. as Consistent Plant Growth Stimulators. Biol. Fertil. Soils 1994, 17, 85–90. [Google Scholar] [CrossRef]
- Yedidia, I.; Benhamou, N.; Chet, I. Induction of Defense Responses in Cucumber Plants (Cucumis sativus L.) by the Biocontrol Agent Trichoderma harzianum. Appl. Environ. Microbiol. 1999, 65, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; El-Nakhel, C.; Rouphael, Y.; Comite, E.; Lombardi, N.; Cuciniello, A.; Woo, S.L. Diplotaxis tenuifolia (L.) DC. Yield and Quality as Influenced by Cropping Season, Protein Hydrolysates, and Trichoderma Applications. Plants 2020, 9, 697. [Google Scholar] [CrossRef] [PubMed]
- Elad, Y. Biological Control of Foliar Pathogens by Means of Trichoderma harzianum and Potential Modes of Action. Crop Prot. 2000, 19, 709–714. [Google Scholar] [CrossRef]
- Howell, C.R. Mechanisms Employed by Trichoderma Species in the Biological Control of Plant Diseases: The History and Evolution of Current Concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef]
- Markovich, N.A.; Kononova, G.L. Lytic Enzymes of Trichoderma and Their Role in Plant Defense from Fungal Diseases: A Review. Appl. Biochem. Microbiol. 2003, 39, 341–351. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma Species—Opportunistic, Avirulent Plant Symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Manganiello, G.; Sacco, A.; Ercolano, M.R.; Vinale, F.; Lanzuise, S.; Pascale, A.; Napolitano, M.; Lombardi, N.; Lorito, M.; Woo, S.L. Modulation of Tomato Response to Rhizoctonia solani by Trichoderma harzianum and Its Secondary Metabolite Harzianic Acid. Front. Microbiol. 2018, 9, 1966. [Google Scholar] [CrossRef]
- Sarma, B.K.; Yadav, S.K.; Singh, S.; Singh, H.B. Microbial Consortium-Mediated Plant Defense against Phytopathogens: Readdressing for Enhancing Efficacy. Soil Biol. Biochem. 2015, 87, 25–33. [Google Scholar] [CrossRef]
- Stockwell, V.O.; Johnson, K.B.; Sugar, D.; Loper, J.E. Mechanistically Compatible Mixtures of Bacterial Antagonists Improve Biological Control of Fire Blight of Pear. Phytopathology 2011, 101, 113–123. [Google Scholar] [CrossRef]
- Duffy, B.K.; Weller, D.M. Biological Control of Take-All of Wheat in the Pacific Northwest of the USA Using Hypovirulent Gaeumannomyces graminis Var. tritici and Fluorescent Pseudomonads. J. Phytopathol. 1996, 144, 585–590. [Google Scholar] [CrossRef]
- Jetiyanon, K. Defensive-Related Enzyme Response in Plants Treated with a Mixture of Bacillus Strains (IN937a and IN937b) against Different Pathogens. Biol. Control 2007, 42, 178–185. [Google Scholar] [CrossRef]
- Kannan, V.; Sureendar, R. Synergistic Effect of Beneficial Rhizosphere Microflora in Biocontrol and Plant Growth Promotion. J. Basic Microbiol. 2009, 49, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.; Khalid, A.; Singh, U.S.; Sharma, A.K. Evaluation of Arbuscular Mycorrhizal Fungus, Fluorescent Pseudomonas and Trichoderma harzianum Formulation against Fusarium oxysporum f. sp. lycopersici for the Management of Tomato Wilt. Biol. Control 2010, 53, 24–31. [Google Scholar] [CrossRef]
- Singh, P.K.; Vijay, K. Biological Control of Fusarium Wilt of Chrysanthemum with Trichoderma and Botanicals. J. Agric. Technol. 2011, 7, 1603–1613. [Google Scholar]
- Řiháková, Z.; Plocková, M.; Filip, V.; Šmidrkal, J. Antifungal Activity of Lauric Acid Derivatives against Aspergillus niger. Eur. Food Res. Technol. 2001, 213, 488–490. [Google Scholar] [CrossRef]
- Dawson, T.E.; Mambelli, S.; Plamboeck, A.H.; Templer, P.H.; Tu, K.P. Stable Isotopes in Plant Ecology. Annu. Rev. Ecol. Syst. 2002, 33, 507–559. [Google Scholar] [CrossRef]
- Yff, B.T.S.; Lindsey, K.L.; Taylor, M.B.; Erasmus, D.G.; Jäger, A.K. The Pharmacological Screening of Pentanisia Prunelloides and the Isolation of the Antibacterial Compound Palmitic Acid. J. Ethnopharmacol. 2002, 79, 101–107. [Google Scholar] [CrossRef]
- Desbois, A.P.; Lebl, T.; Yan, L.; Smith, V.J. Isolation and Structural Characterisation of Two Antibacterial Free Fatty Acids from the Marine Diatom, Phaeodactylum tricornutum. Appl. Microbiol. Biotechnol. 2008, 81, 755–764. [Google Scholar] [CrossRef]
- Dilika, F.; Bremner, P.D.; Meyer, J.J.M. Antibacterial Activity of Linoleic and Oleic Acids Isolated from Helichrysum Pedunculatum: A Plant Used during Circumcision Rites. Fitoterapia 2000, 71, 450–452. [Google Scholar] [CrossRef]
- Walters, D.R.; Walker, R.L.; Walker, K.C. Lauric Acid Exhibits Antifungal Activity Against Plant Pathogenic Fungi. J. Phytopathol. 2003, 151, 228–230. [Google Scholar] [CrossRef]
- Kabara, J.J.; Swieczkowski, D.M.; Conley, A.J.; Truant, J.P. Fatty Acids and Derivatives as Antimicrobial Agents. Antimicrob. Agents Chemother. 1972, 2, 23–28. [Google Scholar] [CrossRef]
- Cerdeiras, M.P.; Fernández, J.; Soubes, M.; Vero, S.; Ferreira, F.; Moyna, P.; Olano, I.; Vázquez, A. A New Antibacterial Compound from Ibicella Lutea. J. Ethnopharmacol. 2000, 73, 521–525. [Google Scholar] [CrossRef]
- Wang, L.L.; Johnson, E.A. Inhibition of Listeria Monocytogenes by Fatty Acids and Monoglycerides. Appl. Environ. Microbiol. 1992, 58, 624–629. [Google Scholar] [CrossRef]
- Thompson, P.A.; Calvert, S.E. Carbon-Isotope Fractionation by a Marine Diatom: The Influence of Irradiance, Daylength, PH, and Nitrogen Source. Limnol. Oceanogr. 1994, 39, 1835–1844. [Google Scholar] [CrossRef]
- Shin, S.Y.; Bajpai, V.K.; Kim, H.R.; Kang, S.C. Antibacterial Activity of Bioconverted Eicosapentaenoic (EPA) and Docosahexaenoic Acid (DHA) against Foodborne Pathogenic Bacteria. Int. J. Food Microbiol. 2007, 113, 233–236. [Google Scholar] [CrossRef]
- Vinale, F.; Flematti, G.; Sivasithamparam, K.; Lorito, M.; Marra, R.; Skelton, B.W.; Ghisalberti, E.L. Harzianic Acid, an Antifungal and Plant Growth Promoting Metabolite from Trichoderma harzianum. J. Nat. Prod. 2009, 72, 2032–2035. [Google Scholar] [CrossRef] [PubMed]
- Vinale, F.; Nigro, M.; Sivasithamparam, K.; Flematti, G.; Ghisalberti, E.L.; Ruocco, M.; Varlese, R.; Marra, R.; Lanzuise, S.; Eid, A.; et al. Harzianic Acid: A Novel Siderophore from Trichoderma harzianum. FEMS Microbiol. Lett. 2013, 347, 123–129. [Google Scholar] [CrossRef]
- Viterbo, A.; Wiest, A.; Brotman, Y.; Chet, I.; Kenerley, C. The 18mer Peptaibols from Trichoderma Virens Elicit Plant Defence Responses. Mol. Plant Pathol. 2007, 8, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Cortés-Penagos, C.; López-Bucio, J. Trichoderma Virens, a Plant Beneficial Fungus, Enhances Biomass Production and Promotes Lateral Root Growth through an Auxin-Dependent Mechanism in Arabidopsis. Plant Physiol. 2009, 149, 1579–1592. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Petropoulos, S.A.; El-Nakhel, C.; Pannico, A.; Kyriacou, M.C.; Giordano, M.; Troise, A.D.; Vitaglione, P.; de Pascale, S. Reducing Energy Requirements in Future Bioregenerative Life Support Systems (BLSSs): Performance and Bioactive Composition of Diverse Lettuce Genotypes Grown Under Optimal and Suboptimal Light Conditions. Front. Plant Sci. 2019, 10, 1305. [Google Scholar] [CrossRef] [PubMed]
- Caporale, A.G.; Vitaglione, P.; Troise, A.D.; Pigna, M.; Ruocco, M. Influence of Three Different Soil Types on the Interaction of Two Strains of Trichoderma harzianum with Brassica rapa subsp. Sylvestris cv. Esculenta, under Soil Mineral Fertilization. Geoderma 2019, 350, 11–18. [Google Scholar] [CrossRef]
- Lorito, M.; Woo, S.L.; Harman, G.E.; Monte, E. Translational Research on Trichoderma: From ’Omics to the Field. Annu. Rev. Phytopathol. 2010, 48, 395–417. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, M.; Mukherjee, P.K.; Horwitz, B.A.; Zachow, C.; Berg, G.; Zeilinger, S. Trichoderma–Plant–Pathogen Interactions: Advances in Genetics of Biological Control. Indian J. Microbiol. 2012, 52, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Kumari, I.; Hussain, R.; Sharma, S.; Geetika; Ahmed, M. Microbial Biopesticides for Sustainable Agricultural Practices. In Biopesticides; Woodhead Publishing: Cambridge, UK, 2022; pp. 301–317. [Google Scholar]
- Das, T.; Tudu, C.K.; Nandy, S.; Pandey, D.K.; Dey, A. Role of Fungal Metabolites as Biopesticides: An Emerging Trend in Sustainable Agriculture. In Volatiles and Metabolites of Microbes; Academic Press: San Diego, CA, USA, 2021; pp. 385–407. [Google Scholar]
- Altieri, C.; Cardillo, D.; Bevilacqua, A.; Sinigallia, M. Inhibition of Aspergillus spp. and Penicillium spp. by Fatty Acids and Their Monoglycerides. J. Food Prot. 2007, 70, 1206–1212. [Google Scholar] [CrossRef] [PubMed]
- Avis, T.J.; Anguenot, R.; Neveu, B.; Bolduc, S.; Zhao, Y.; Cheng, Y.; Labbè, C.; Belzile, F.; Belanger, R.R. Usefulness of Heterologous Promoters in the Pseudozyma Flocculosa Gene Expression System. Biosci. Biotechnol. Biochem. 2008, 72, 456–462. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Liu, S.; Ruan, W.; Li, J.; Xu, H.; Wang, J.; Gao, Y.; Wang, J. Biological Control of Phytopathogenic Fungi by Fatty Acids. Mycopathologia 2008, 166, 93–102. [Google Scholar] [CrossRef]
- Benyagoub, M.; Bel Rhlid, R.; Bélanger, R.R. Purification and Characterization of New Fatty Acids with Antibiotic Activity Produced BySporothrix Flocculosa. J. Chem. Ecol. 1996, 22, 405–413. [Google Scholar] [CrossRef]
- McDonough, V.; Stukey, J.; Cavanagh, T. Mutations in Erg4 Affect the Sensitivity of Saccharomyces Cerevisiae to Medium-Chain Fatty Acids. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2002, 1581, 109–118. [Google Scholar] [CrossRef]
- Aranega-Bou, P.; de la O Leyva, M.; Finiti, I.; García-Agustín, P.; González-Bosch, C. Priming of Plant Resistance by Natural Compounds. Hexanoic Acid as a Model. Front. Plant Sci. 2014, 5, 488. [Google Scholar] [CrossRef]
- Leyva, M.O.; Vicedo, B.; Finiti, I.; Flors, V.; del Amo, G.; Real, M.D.; García-Agustín, P.; González-Bosch, C. Preventive and Post-Infection Control of Botrytis Cinerea in Tomato Plants by Hexanoic Acid. Plant Pathol. 2008, 57, 1038–1046. [Google Scholar] [CrossRef]
- Preisig, C.L.; Kuć, J.A. Metabolism by Potato Tuber of Arachidonic Acid, an Elicitor of Hypersensitive Resistance. Physiol. Mol. Plant Pathol. 1988, 32, 77–88. [Google Scholar] [CrossRef]
- Bostock, R.M.; Quatrano, R.S. Regulation of Em Gene Expression in Rice. Plant Physiol. 1992, 98, 1356–1363. [Google Scholar] [CrossRef]
- Ruan, W.; Wang, J.; Pan, J.; Li, H.; Wang, J.; Zhang, F.; Gao, Y. Effects of Sesame Seed Cake Allelochemicals on the Growth Cucumber (Cucumis sativus L. cv. Jinchun 4). Allelopath. J. 2005, 16, 217–225. [Google Scholar]
- Weibin, R.; Mohan, L.; Jie, P.; Jing, W.; Wenlong, L.; Chengcang, M.; Jingguo, W.; Yubao, G. Effect of Oil Cakes on the Growth of Cucumber (Cucumis sativus L.) under Continuous Cropping System and Its Mechanism. Sci. Agric. Sinica. 2003, 36, 1519–1524. [Google Scholar]
- Kinderlerer, J.L. Fungal Strategies for Detoxification of Medium Chain Fatty Acids. Int. Biodeterior. Biodegrad. 1993, 32, 213–224. [Google Scholar] [CrossRef]
- Aneja, M.; Gianfagna, T.J.; Hebbar, P.K. Trichoderma harzianum Produces Nonanoic Acid, an Inhibitor of Spore Germination and Mycelial Growth of Two Cacao Pathogens. Physiol. Mol. Plant Pathol. 2005, 67, 304–307. [Google Scholar] [CrossRef]
- Calvo-Garrido, C.; Elmer, P.A.G.; Viñas, I.; Usall, J.; Bartra, E.; Teixidó, N. Biological Control of Botrytis Bunch Rot in Organic Wine Grapes with the Yeast Antagonist Candida Sake CPA-1. Plant Pathol. 2013, 62, 510–519. [Google Scholar] [CrossRef]
- Cañamás, T.P.; Viñas, I.; Torres, R.; Usall, J.; Solsona, C.; Teixidó, N. Field Applications of Improved Formulations of Candida Sake CPA-1 for Control of Botrytis Cinerea in Grapes. Biol. Control 2011, 56, 150–158. [Google Scholar] [CrossRef]
- Weber, H.; Chételat, A.; Caldelari, D.; Farmer, E.E. Divinyl Ether Fatty Acid Synthesis in Late Blight–Diseased Potato Leaves. Plant Cell 1999, 11, 485–493. [Google Scholar] [CrossRef]
- Creelman, R.A.; Mullet, J.E. Biosynthesis and Action of Jasmonates in Plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 355–381. [Google Scholar] [CrossRef]
- Farmer, E.E.; Weber, H.; Vollenweider, S. Fatty Acid Signaling in Arabidopsis. Planta 1998, 206, 167–174. [Google Scholar] [CrossRef]
- Weber, H. Fatty Acid-Derived Signals in Plants. Trends Plant Sci. 2002, 7, 217–224. [Google Scholar] [CrossRef]
- Baker, R. Trzchoderma spp. as Plant-Growth Stimulants. Crit. Rev. Biotechnol. 1988, 7, 97–106. [Google Scholar] [CrossRef]
- Tucci, M.; Ruocco, M.; de Masi, L.; de Palma, M.; Lorito, M. The Beneficial Effect of Trichoderma spp. on Tomato Is Modulated by the Plant Genotype. Mol. Plant Pathol. 2011, 12, 341–354. [Google Scholar] [CrossRef]
- Mannino, G.; Gentile, C.; Ertani, A.; Serio, G.; Bertea, C.M. Anthocyanins: Biosynthesis, Distribution, Ecological Role, and Use of Biostimulants to Increase Their Content in Plant Foods—A Review. Agriculture 2021, 11, 212. [Google Scholar] [CrossRef]
- Mannino, G.; Campobenedetto, C.; Vigliante, I.; Contartese, V.; Gentile, C.; Bertea, C.M. The Application of a Plant Biostimulant Based on Seaweed and Yeast Extract Improved Tomato Fruit Development and Quality. Biomolecules 2020, 10, 1662. [Google Scholar] [CrossRef]
- Kocira, S. Effect of Amino Acid Biostimulant on the Yield and Nutraceutical Potential of Soybean. Chil. J. Agric. Res. 2019, 79, 17–25. [Google Scholar] [CrossRef]
- Graziani, G.; Ritieni, A.; Cirillo, A.; Cice, D.; di Vaio, C. Effects of Biostimulants on Annurca Fruit Quality and Potential Nutraceutical Compounds at Harvest and during Storage. Plants 2020, 9, 775. [Google Scholar] [CrossRef] [PubMed]
- Szeto, Y.T.; Kwok, T.C.Y.; Benzie, I.F.F. Effects of a Long-Term Vegetarian Diet on Biomarkers of Antioxidant Status and Cardiovascular Disease Risk. Nutrition 2004, 20, 863–866. [Google Scholar] [CrossRef]
- Higdon, J.; Delage, B.; Williams, D.; Dashwood, R. Cruciferous Vegetables and Human Cancer Risk: Epidemiologic Evidence and Mechanistic Basis. Pharmacol. Res. 2007, 55, 224–236. [Google Scholar] [CrossRef]
- Sesso, H.D.; Liu, S.; Gaziano, J.M.; Buring, J.E. Dietary Lycopene, Tomato-Based Food Products and Cardiovascular Disease in Women. J. Nutr. 2003, 133, 2336–2341. [Google Scholar] [CrossRef]
- Omoni, A.O.; Aluko, R.E. The Anti-Carcinogenic and Anti-Atherogenic Effects of Lycopene: A Review. Trends Food Sci. Technol. 2005, 16, 344–350. [Google Scholar] [CrossRef]
- Stahl, W.; Sies, H. Bioactivity and Protective Effects of Natural Carotenoids. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2005, 1740, 101–107. [Google Scholar] [CrossRef]
- Burton-Freeman, B.; Reimers, K. Tomato Consumption and Health: Emerging Benefits. Am. J. Lifestyle Med. 2011, 5, 182–191. [Google Scholar] [CrossRef]
- Mayne, S.T. Beta-carotene, Carotenoids, and Disease Prevention in Humans. FASEB J. 1996, 10, 690–701. [Google Scholar] [CrossRef]
- Pascale, A.; Vinale, F.; Manganiello, G.; Nigro, M.; Lanzuise, S.; Ruocco, M.; Marra, R.; Lombardi, N.; Woo, S.L.; Lorito, M. Trichoderma and Its Secondary Metabolites Improve Yield and Quality of Grapes. Crop Prot. 2017, 92, 176–181. [Google Scholar] [CrossRef]
- Khan, M.Y.; Haque, M.M.; Molla, A.H.; Rahman, M.M.; Alam, M.Z. Antioxidant Compounds and Minerals in Tomatoes by Trichoderma-Enriched Biofertilizer and Their Relationship with the Soil Environments. J. Integr. Agric. 2017, 16, 691–703. [Google Scholar] [CrossRef]
- Molla, A.H.; Manjurul Haque, M.; Amdadul Haque, M.; Ilias, G.N.M. Trichoderma-Enriched Biofertilizer Enhances Production and Nutritional Quality of Tomato (Lycopersicon esculentum Mill.) and Minimizes NPK Fertilizer Use. Agric. Res. 2012, 1, 265–272. [Google Scholar] [CrossRef]
- Halkier, B.A.; Gershenzon, J. Biology and Biochemistry of Glucosinolates. Annu. Rev. Plant Biol. 2006, 57, 303–333. [Google Scholar] [CrossRef] [PubMed]
- Njoroge, S.M.C.; Vallad, G.E.; Park, S.-Y.; Kang, S.; Koike, S.T.; Bolda, M.; Burman, P.; Polonik, W.; Subbarao, K.V. Phenological and Phytochemical Changes Correlate with Differential Interactions of Verticillium Dahliae with Broccoli and Cauliflower. Phytopathology 2011, 101, 523–534. [Google Scholar] [CrossRef]
- Sandrock, R.W.; VanEtten, H.D. Fungal Sensitivity to and Enzymatic Degradation of the Phytoanticipin α-Tomatine. Phytopathology 1998, 88, 137–143. [Google Scholar] [CrossRef]
- Stringlis, I.A.; de Jonge, R.; Pieterse, C.M.J. The Age of Coumarins in Plant–Microbe Interactions. Plant Cell Physiol. 2019, 60, 1405–1419. [Google Scholar] [CrossRef] [PubMed]
- Wojtaszek, P.; Roshchina, V.V. Neurotransmitters in Plant Life. Ann. Bot. 2003, 92, 166. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | [M-H]− m/z | Production m/z |
|---|---|---|
| Sinigrin | 358 | 97 |
| Glucoraphanin | 436 | 97 |
| Glucoalyssin | 450 | 97 |
| Glucoiberin | 422 | 97 |
| 4-Hydroxyglucobrassicin | 463 | 97 |
| 4-Methoxyglucobrassicin | 477 | 97 |
| Glucobrassicin | 447 | 97 |
| Neoglucobrassicin | 477 | 97 |
| Glucoiberverin | 406 | 97 |
| Glucoerucin | 420 | 97 |
| Glucotropaleoin | 408 | 97 |
| Dehydro Phytosfingosine | Bufotenine | A−Tomatidan−3−One | Tomatidine | Coumarin | Dehydrotomatine | Tomatine | |
|---|---|---|---|---|---|---|---|
| FAm 0.3 % | −0.1456 | −0.1456 | −0.12479 | −0.11483 | −4.82014 | −0.1238 | −0.1456 |
| FAm 0.5 % | −0.34555 | −0.34555 | −20.1518 | −0.32685 | 4.396446 | −0.23038 | −0.34555 |
| FAm 1 % | −0.22871 | −0.22871 | −20.035 | −0.19447 | 4.629767 | −0.21834 | −0.22871 |
| M10 | −0.22533 | −0.22533 | −20.0316 | −0.45842 | 4.568698 | −0.20868 | −0.22533 |
| GV41 | −0.28736 | −0.28736 | −20.0936 | −0.6035 | 0.218449 | 0.19646 | 0.28736 |
| M10 + GV41 | 16.81207 | −0.45301 | −20.2593 | −0.12306 | −0.12754 | −0.61138 | 20.77753 |
| M10 + FAm 0.3% | 0.232552 | 0.232552 | −19.5737 | −0.10955 | −4.44199 | 0.413509 | 0.232552 |
| M10 + FAm 0.5% | −0.48115 | −0.48115 | −20.2874 | −0.29364 | −5.15569 | −0.48943 | 6.805254 |
| M10 + FAm 1% | 15.85772 | −1.03429 | −20.8406 | −0.42998 | 9.630247 | −1.09503 | 21.37197 |
| GV41 + FAm 0.3% | 16.29603 | −0.64539 | −20.4517 | −0.52558 | 9.144241 | 0.72401 | 21.7276 |
| GV41 + FAm 0.5% | 16.271 | 17.01381 | −20.4952 | −0.50401 | 4.304544 | 0.74871 | 21.61315 |
| GV41 + FAm 1% | 16.02612 | −0.68987 | −20.4961 | −0.70976 | 4.336587 | −0.7407 | 19.50915 |
| M10 + GV41 + FAm 0.3% | 16.17436 | 17.29442 | −20.5374 | −0.30001 | 8.920802 | −0.87419 | 21.44018 |
| M10 + GV41 + FAm 0.5% | 16.20062 | −0.72027 | −0.5265 | −0.3068 | 9.610603 | −0.88789 | 21.98139 |
| M10 + GV41 + FAm 1% | 15.98161 | 16.83258 | −20.6264 | −0.6515 | −5.49467 | 0.84643 | 21.90122 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lanzuise, S.; Manganiello, G.; Guastaferro, V.M.; Vincenzo, C.; Vitaglione, P.; Ferracane, R.; Vecchi, A.; Vinale, F.; Kamau, S.; Lorito, M.; et al. Combined Biostimulant Applications of Trichoderma spp. with Fatty Acid Mixtures Improve Biocontrol Activity, Horticultural Crop Yield and Nutritional Quality. Agronomy 2022, 12, 275. https://doi.org/10.3390/agronomy12020275
Lanzuise S, Manganiello G, Guastaferro VM, Vincenzo C, Vitaglione P, Ferracane R, Vecchi A, Vinale F, Kamau S, Lorito M, et al. Combined Biostimulant Applications of Trichoderma spp. with Fatty Acid Mixtures Improve Biocontrol Activity, Horticultural Crop Yield and Nutritional Quality. Agronomy. 2022; 12(2):275. https://doi.org/10.3390/agronomy12020275
Chicago/Turabian StyleLanzuise, Stefania, Gelsomina Manganiello, Valentino M. Guastaferro, Cono Vincenzo, Paola Vitaglione, Rosalia Ferracane, Alfeo Vecchi, Francesco Vinale, Solomon Kamau, Matteo Lorito, and et al. 2022. "Combined Biostimulant Applications of Trichoderma spp. with Fatty Acid Mixtures Improve Biocontrol Activity, Horticultural Crop Yield and Nutritional Quality" Agronomy 12, no. 2: 275. https://doi.org/10.3390/agronomy12020275
APA StyleLanzuise, S., Manganiello, G., Guastaferro, V. M., Vincenzo, C., Vitaglione, P., Ferracane, R., Vecchi, A., Vinale, F., Kamau, S., Lorito, M., & Woo, S. L. (2022). Combined Biostimulant Applications of Trichoderma spp. with Fatty Acid Mixtures Improve Biocontrol Activity, Horticultural Crop Yield and Nutritional Quality. Agronomy, 12(2), 275. https://doi.org/10.3390/agronomy12020275