

A New Approach for Timing Post-Emergence Weed Control Measures in Crops: The Use of the Differential Form of the Emergence Model


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Data
2.2. Fitting the Differential Form of the Gompertz Function
- A.
- O2: Smoothing weed emergence data (Ei).
- B.
- Estimating the numerical derivative of the raw (in O1) or smoothed (in O2) emergence
- C.
- Smoothing the estimated derivatives
- D.
- Fitting Gompertz function
- E.
- Determination of the percentage of emergence
- F.
- Implementation of the protocol
- G.
- Validation of the approach
3. Results
3.1. Implementation of the Protocol in One Data Set: How It Works
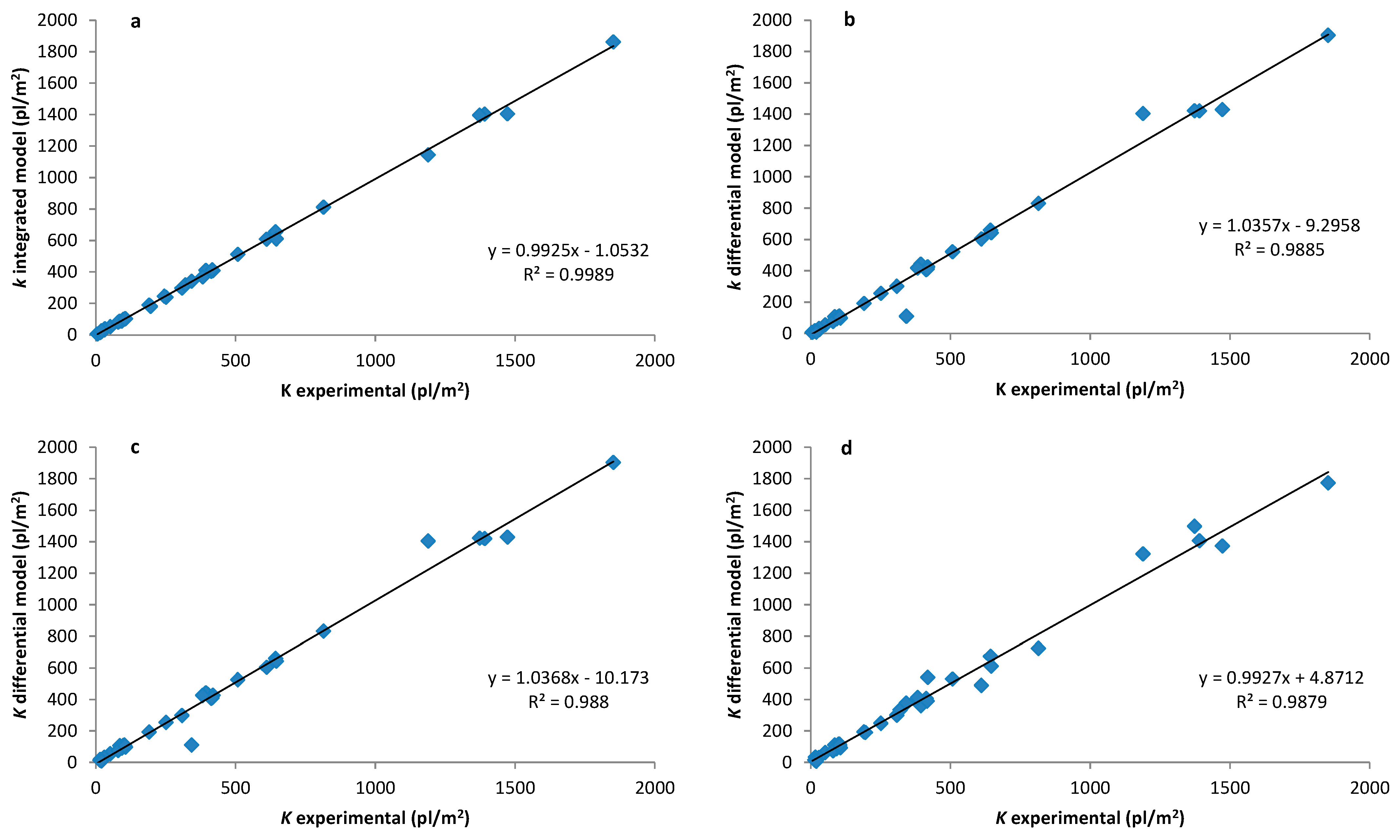
3.2. Implementation of the Protocol in All Data Sets and Accuracy Obtained
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghersa, C.M.; Benech-Arnold, R.L.; Satorre, E.H.; Martinez-Ghers, M.A. Advances in weed management strategies. Field Crops Res. 2000, 67, 95–104. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Gill, G.; Preston, C. Influence of environmental factors on seed germination and seedling emergence of rígid ryegrass (Lolium rigidum). Weed Sci. 2006, 54, 1004–1012. [Google Scholar] [CrossRef]
- Grundy, A.C.; Phelps, K.; Reader, R.J.; Burston, S. Modelling the germination of Stelllaria media using the concept of hydrothermal time. New Phytol. 2000, 148, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Vleeshouwers, L.M.; Kropff, M.J. Modelling field emergence patterns in arable weeds. New Phytol. 2000, 148, 445–457. [Google Scholar] [CrossRef]
- Colbach, N.; Durr, C.; Roger-Estrade, J.; Chauvel, B.; Caneill, J. ALOMYSYS: Modelling black-grass (Alopecurus myosuroides Huds.) germination and emergence, in interaction with seed characteristics, tillage and soil climate–I. Construction. Eur. J. Agron. 2006, 24, 95–112. [Google Scholar] [CrossRef]
- Gardarin, A.; Dürr, C.; Colbach, N. Modeling the dynamics and emergence of a multispecies weed seed bank with species traits. Ecol. Model. 2012, 240, 123–138. [Google Scholar] [CrossRef]
- Grundy, A.C. Predicting weed emergence: A review of approaches and future challenges. Weed Res. 2003, 43, 1–11. [Google Scholar] [CrossRef]
- Schwinghamer, T.D.; Van Acker, R.C. Emergence timing and persistence of Kochia (Kochia scoparia). Weed Sci. 2008, 56, 37–41. [Google Scholar] [CrossRef]
- Izquierdo, J.; González-Andújar, J.L.; Bastida, F.; Lezaun, J.A.; Sanchez del Arco, M.J. A thermal time model to predict corn poppy (Papaver rhoeas) emergence in cereal fields. Weed Sci. 2009, 57, 660–664. [Google Scholar] [CrossRef]
- Leon, R.G.; Izquierdo, J.; González-Andújar, J.L. Characterization and modeling of itchgrass (Rottboellia cochinchinensis) biphasic seedling emergence patterns in the tropics. Weed Sci. 2015, 63, 623–630. [Google Scholar] [CrossRef]
- Royo-Esnal, A.; Torra, J.; Conesa, J.A.; Recasen, J. Emergence and early growth of Galium aparine and Galium spurium. Weed Res. 2012, 52, 458–466. [Google Scholar] [CrossRef]
- Garcia, A.L.; Recasens, J.; Forcella, F.; Torra, J.; Royo-Esnal, A. Hydrothermal emergence model for ripgut brome (Bromus diandrus). Weed Sci. 2013, 61, 146–153. [Google Scholar] [CrossRef]
- Barnes, E.R.; Werle, R.; Sandell, L.D.; Lindquist, J.L.; Knezevic, S.Z.; Sikkema, P.H.; Jhala, A.J. Influence of tillage on common ragweed (Ambrosia artemisiifolia) emergence pattern in Nebraska. Weed Technol. 2017, 31, 623–631. [Google Scholar] [CrossRef]
- Brown, R.F.; Mayer, D.G. Representing cumulative germination. 2. The use of the Weibull function and other empirically derived curves. Ann. Bot. 1988, 61, 127–138. [Google Scholar] [CrossRef]
- Forcella, F. Real-time assessment of seed dormancy and seedling growth for weed management. Seed Sci. Res. 1998, 8, 201–209. [Google Scholar] [CrossRef]
- Masin, R.; Loddo, D.; Gasparini, V.; Otto, S.; Zanin, G. Evaluation of weed emergence model AlertInf for maize and soybean. Weed Sci. 2014, 62, 360–369. [Google Scholar] [CrossRef]
- Loddo, D.; Bozic, D.; Calha, I.M.; Dorado, J.; Izquierdo, J.; Šćepanović, M.; Barić, K.; Carlesi, S.; Leskovsek, R.; Peterson, D.; et al. Variability in seedling emergence for European and North American populations of Abutilon theophrasti. Weed Res. 2019, 59, 15–27. [Google Scholar] [CrossRef]
- Sousa-Ortega, C.; Chamber, E.; Urbano, J.M.; Izquierdo, J.; Loureiro, I.; Marí, A.I.; Cordero, C.; Vargas, M.; Saavedra, M.; Lezaun, J.A.; et al. Should emergence models for Lolium rigidum be changed throughout climatic conditions? The case of Spain. Crop Prot. 2020, 128, 105012. [Google Scholar] [CrossRef]
- González-Andújar, J.L.; Francisco-Fernandez, M.; Cao, R.; Reyes, M.; Urbano, J.M.; Forcella, F.; Bastida, F. A comparative study between nonlinear regression and non-parametric approaches for modelling Phalaris paradoxa seedling emergence. Weed Sci. 2016, 56, 367–376. [Google Scholar]
- Onofri, A.; Benincasa, P.; Mesgara, M.B.; Ritz, C. Hydrothermal-time-to-event models for seed germination. Eur. J. Agron. 2018, 101, 129–139. [Google Scholar] [CrossRef]
- González-Andújar, J.L.; Chantre, G.R.; Morvillo, C.; Blanco, A.M.; Forcella, F. Predicting field weed emergence with empirical models and soft computing techniques. Weed Sci. 2016, 56, 415–423. [Google Scholar] [CrossRef]
- Hadi, M.R.H.S.; Gonzalez-Andujar, J.L. Comparison of fitting weed seedling emergence models with nonlinear regression and genetic algorithm. Comput. Electron. Agric. 2009, 65, 19–25. [Google Scholar] [CrossRef]
- Blanco, A.M.; Chantre, G.R.; Lodovichi, M.V.; Bandoni, J.A.; Lopez, R.L.; Vigna, M.R.; Gigon, R.; Sabbatini, M.R. Modeling seed dormancy release and germination for predicting Avena fatua L. field emergence: A genetic algorithm approach. Ecol. Model. 2014, 272, 293–300. [Google Scholar] [CrossRef]
- Chantre, G.R.; Vigna, M.R.; Renzi, J.P.; Blanco, A.M. A flexible and practical approach for real-time weed emergence prediction based on Artificial Neural Networks. Biosyst. Eng. 2018, 170, 51–60. [Google Scholar] [CrossRef]
- Onofri, A.; Gresta, F.; Tei, F. A new method for the analysis of germination and emergence data of weed species. Weed Res. 2010, 50, 187–198. [Google Scholar] [CrossRef]
- Dorado, J.; Sousa, E.; Calha, I.M.; González-Andújar, J.L.; Fernández-Quintanilla, C. Predicting weed emergence in maize crops under two contrasting climatic conditions. Weed Res. 2009, 49, 251–260. [Google Scholar] [CrossRef]
- Izquierdo, J.; Bastida, F.; Lezaun, J.M.; Sanchez Del Arco, M.J.; González-Andújar, J.L. Development and evaluation of a model for predicting Lolium rigidum emergence in winter cereal crops in the Mediterranean area. Weed Res. 2013, 53, 269–278. [Google Scholar] [CrossRef]
- Bradford, K.J. Application of hydrothermal time to quantifying and modeling seed germination and dormancy. Weed Sci. 2002, 50, 248–260. [Google Scholar] [CrossRef]
- Mohler, C.L. Weed life history: Identifying vulnerabilities. In Ecological Management of Agricultural Weeds; Liebman, M., Mohler, C.L., Staver, C.P., Eds.; Cambridge University Press: Cambridge, UK, 2001; pp. 40–98. [Google Scholar]
- Smith, S.W. Moving Average Filters. In The Scientist and Engineer’s Guide to Digital Signal Processing; California Technical Publishing: Carlsbad, CA, USA, 1997; pp. 277–284. [Google Scholar]
- Levy, D. Introduction to Numerical Analysis; Dept. Mathematics and Center for Scientific Computation and Mathematical Modeling, University of Maryland: College Park, MD, USA, 2010; 121p. [Google Scholar]
- Swanton, C.J.; Weaver, S.; Cowan, P.; Van Acker, R.; Deen, W.; Shreshta, A. Weed thresholds. J. Crop Prod. 1999, 2, 9–29. [Google Scholar] [CrossRef]
- Das, B.S.; Sarathjith, M.C.; Santra, P.; Sahoo, R.N.; Srivastava, R.; Routray, A.; Ray, S.S. Hyperspectral remote sensing: Opportunities, status and challenges for rapid soil assessment in India. Curr. Sci. 2015, 108, 860–868. [Google Scholar]



{kind=link}
{kind=link}
| Weed | Crop | Sites | Year | Water Regime | Source * |
|---|---|---|---|---|---|
| Digitaria sanguinalis (L.) Scop. | Citrus | Huelva1 | 2008 | Irrigated | 4 |
| Huelva2 | 2008 | Irrigated | 4 | ||
| Corn | Arganda | 2005, 2006 | Irrigated | 1 | |
| Golega + | 2007 | Irrigated | 1 | ||
| La Roca1 | 2007 | Irrigated | 4 | ||
| Miralcamp | 2010 | Irrigated | 4 | ||
| Mollerussa | 2010 | Irrigated | 4 | ||
| Echinochloa crus-galli (L.) P. Beauv. | Corn | Golega + | 2006 | Irrigated | 1 |
| La Roca1 | 2006, 2007 | Irrigated | 4 | ||
| La Roca2 | 2008 | Irrigated | 4 | ||
| La Roca3 | 2009 | Irrigated | 4 | ||
| Miralcamp | 2010 | Irrigated | 4 | ||
| Mollerussa | 2010 | Irrigated | 4 | ||
| Lolium rigidum Gaudin | Cereal † | Albacete | 2007, 2008 | Dryland | 4 |
| Calaf | 2006, 2007, 2008 | Dryland | 3 | ||
| El Encín | 2008 | Dryland | 3 | ||
| Huelva | 2006 | Dryland | 4 | ||
| Igualada | 2006, 2007, 2008 | Dryland | 3 | ||
| Murillo | 2008 | Dryland | 3 | ||
| Papaver rhoeas L. | Cereal † | Calaf | 2006, 2007, 2008 | Dryland | 2 |
| El Encín | 2008 | Dryland | 2 | ||
| Igualada | 2006, 2007, 2008 | Dryland | 2 | ||
| Murillo | 2008 | Dryland | 2 | ||
| Phalaris brachystachis Link | Cereal † | Huelva | 2008 | Dryland | 4 |
| Phalaris paradoxa L. | Cereal † | Tajonar | 2008 | Dryland | 4 |
| Portulaca oleracea L. | Citrus | Huelva1 | 2008 | Irrigated | 4 |
| Huelva2 | 2008 | Irrigated | 4 | ||
| Corn | La Roca1 | 2006 | Irrigated | 4 |
| A. Filter Applied to Smooth Emergence and Numerical Derivative | |||||
|---|---|---|---|---|---|
| Parameters | Week | ||||
| 7 | 8 | 9 | 10 | 11 | |
| Data points of regression | 2 | 3 | 4 | 5 | 6 |
| Numerical derivative (pl.MPa/m2.day) | 0.898 | 0.485 | 0.230 | 0.070 | 0.010 |
| K differential (pl/m2) | 204 [NA-NA] | 189 [85–293] | 190 [131–250] | 192 [145–240] | 193 [155–231] |
| E experimental (pl/m2) | 188 | 190 | 191 | 191 | 191 |
| B. Filter Applied to Smooth Numerical Derivative | |||||
| Parameters | Week | ||||
| 5 | 6 | 7 | 8 | 9 | |
| Data points of regression | 2 | 3 | 4 | 5 | 6 |
| Numerical derivative (pl.MPa/m2.day) | 1.061 | 0.941 | 0.571 | 0.183 | 0.030 |
| K differential (pl/m2) | 214 [NA-NA] | 200 [136–263] | 189 [124–254] | 192 [141–244] | 194 [150–238] |
| E experimental (pl/m2) | 148 | 174 | 188 | 190 | 191 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Izquierdo, J.; Prats, C.; Gallart, M.; López, D. A New Approach for Timing Post-Emergence Weed Control Measures in Crops: The Use of the Differential Form of the Emergence Model. Agronomy 2022, 12, 2896. https://doi.org/10.3390/agronomy12112896
Izquierdo J, Prats C, Gallart M, López D. A New Approach for Timing Post-Emergence Weed Control Measures in Crops: The Use of the Differential Form of the Emergence Model. Agronomy. 2022; 12(11):2896. https://doi.org/10.3390/agronomy12112896
Chicago/Turabian StyleIzquierdo, Jordi, Clara Prats, Montserrat Gallart, and Daniel López. 2022. "A New Approach for Timing Post-Emergence Weed Control Measures in Crops: The Use of the Differential Form of the Emergence Model" Agronomy 12, no. 11: 2896. https://doi.org/10.3390/agronomy12112896
APA StyleIzquierdo, J., Prats, C., Gallart, M., & López, D. (2022). A New Approach for Timing Post-Emergence Weed Control Measures in Crops: The Use of the Differential Form of the Emergence Model. Agronomy, 12(11), 2896. https://doi.org/10.3390/agronomy12112896