
Dissecting the Genetic Mechanisms of Hemicellulose Content in Rapeseed Stalk

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Experimental Design
2.3. Phenotypic Data Collectionion
2.4. Statistical Analysis
2.5. Genotyping, Population Structure Linkage and Disequilibrium Analysis
2.6. Genome-Wide Association Study
2.7. Functional Annotation and Tissue Expression Analysis of Candidate Genes
2.8. Transcriptome Sequencing and Identification of Differentially Expressed Genes
3. Results and Analysis
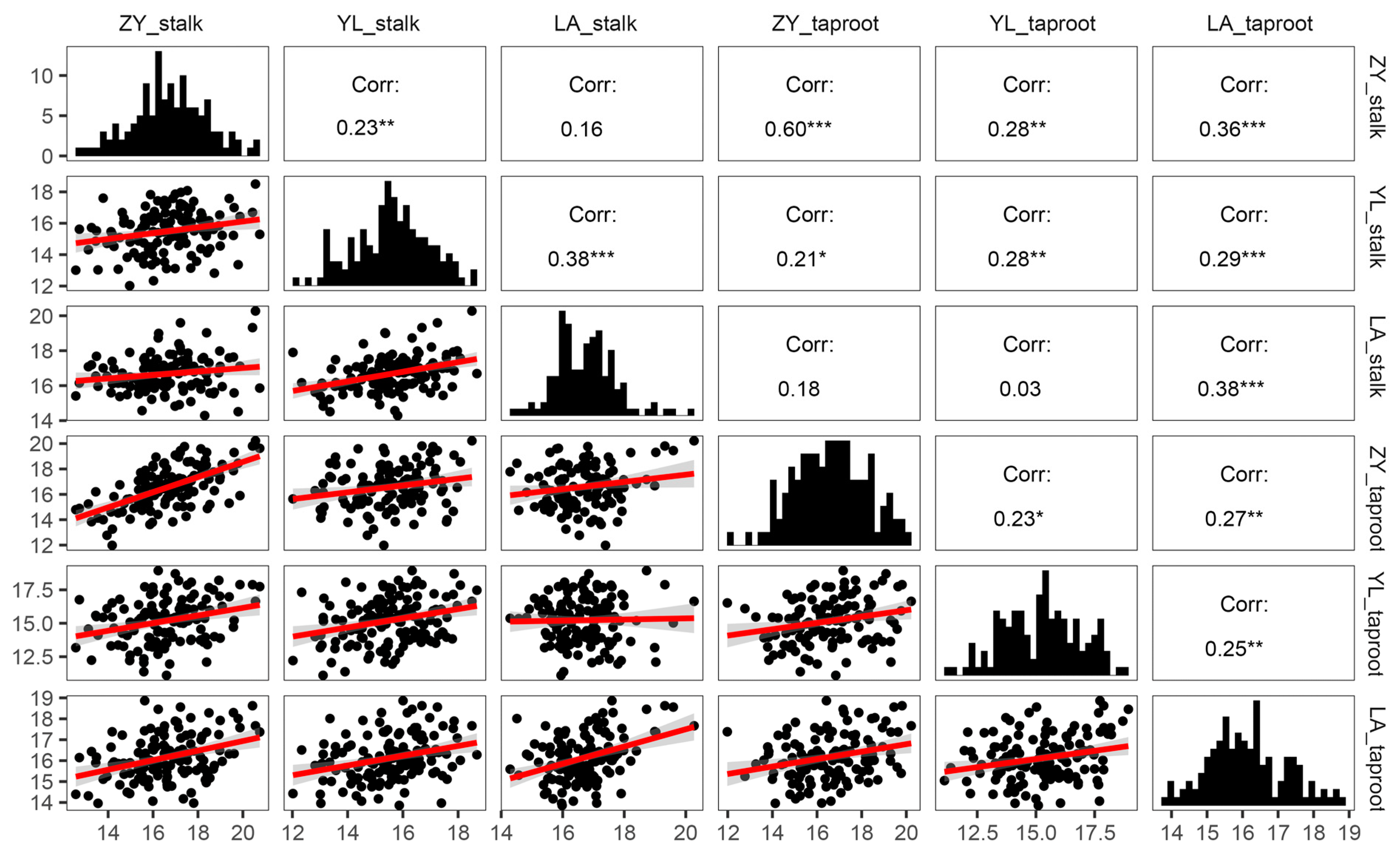
3.1. Evaluation of Hemicellulose Content in Rapeseed Core Natural Population
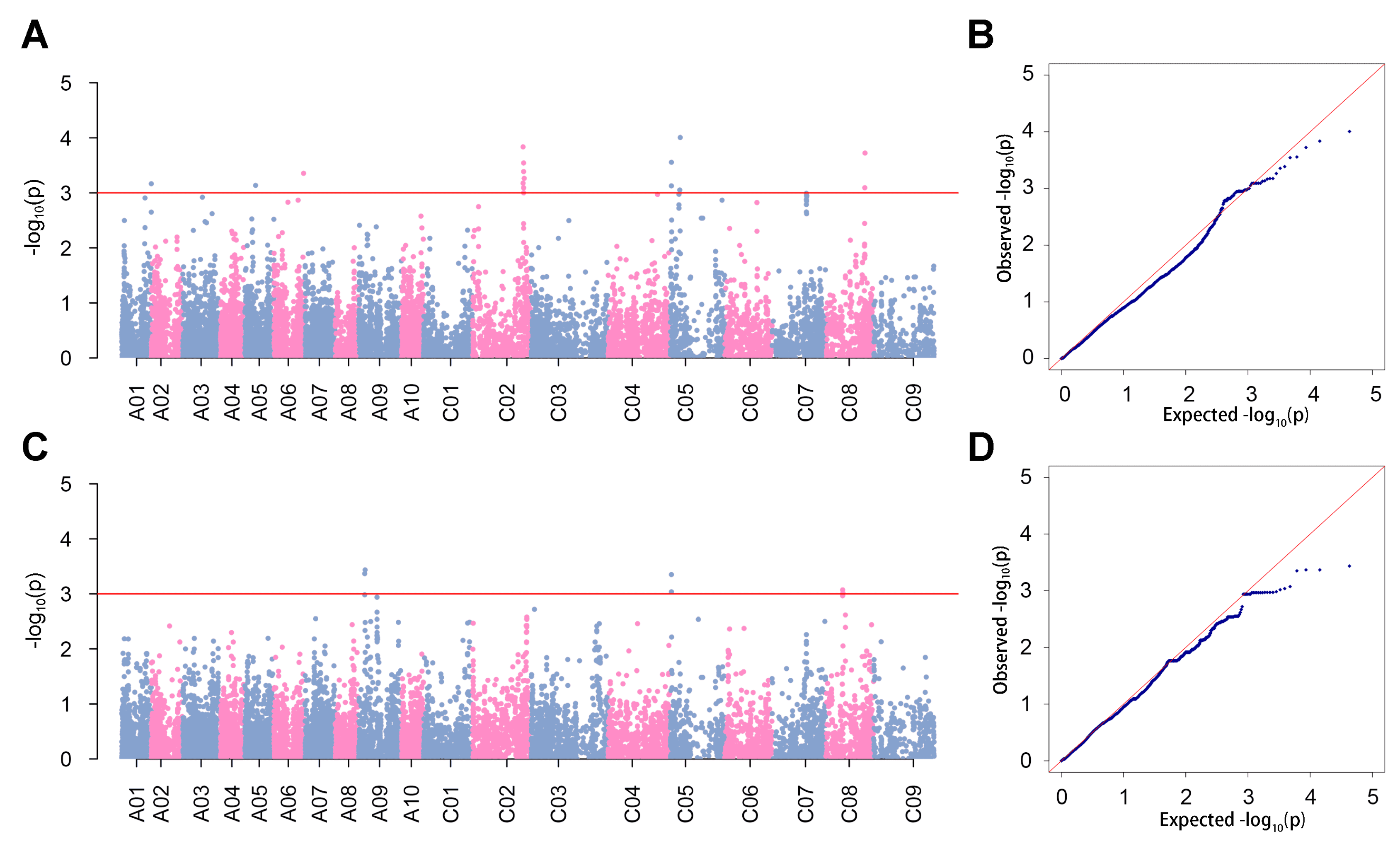
3.2. GWAS for Hemicellulose Content
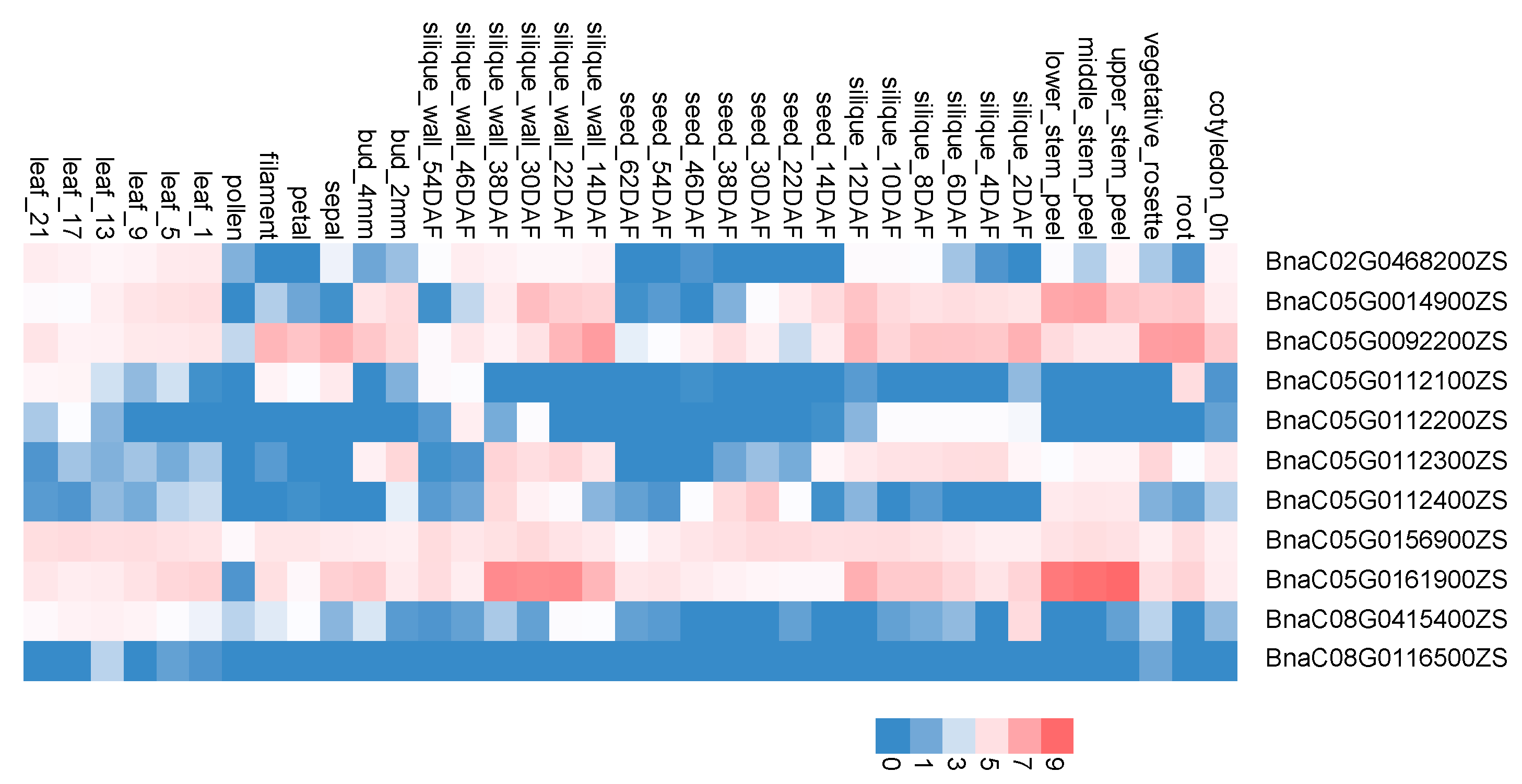
3.3. Screening Candidate Genes Associated with Hemicellulose Content
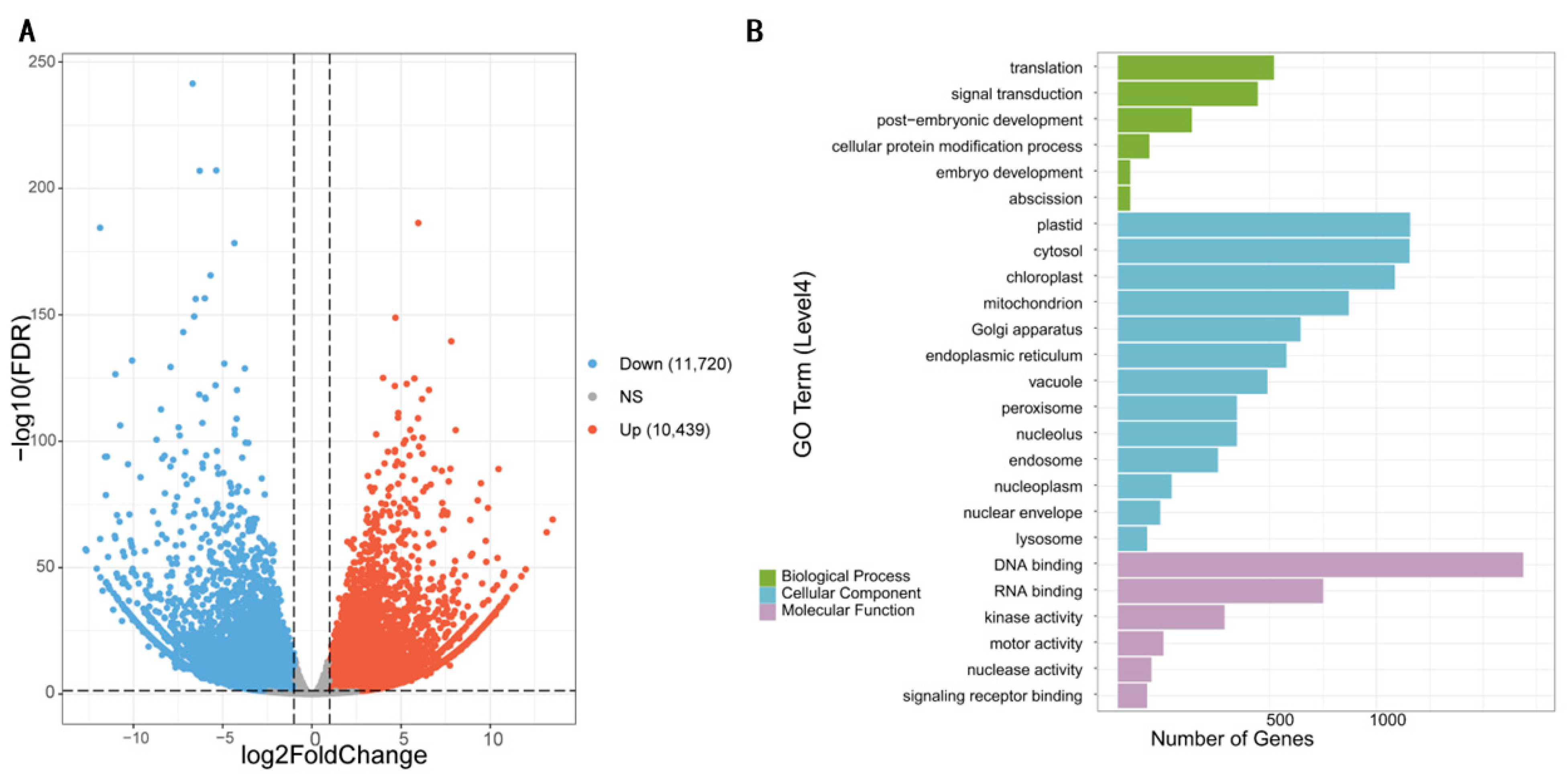
3.4. Identification of Differentially Expressed Genes and GO Enrichment Analysis
3.5. Combining GWAS and Transcriptome to Identify Candidate Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cosgrove, D.J. Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 2005, 6, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Gao, Y.; Zhang, L.; Zhou, Y. The plant cell wall: Biosynthesis, construction, and functions. J. Integr. Plant Biol. 2021, 63, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef] [PubMed]
- Schultink, A.; Liu, L.; Zhu, L.; Pauly, M. Structural diversity and function of xyloglucan sidechain substituents. Plants 2014, 3, 526–542. [Google Scholar] [CrossRef] [PubMed]
- Khodayari, A.; Thielemans, W.; Hirn, U.; Van Vuure, A.W.; Seveno, D. Cellulose-hemicellulose interactions-a nanoscale view. Carbohydr. Polym. 2021, 270, 118364. [Google Scholar] [CrossRef]
- Cuimei, L.; Xujie, W.; Jun, M.; Chusheng, Q. Effects of exogenous acid catalysis on the thermal degradation law of wood hemicellulose. J. Beijing For. Univ. 2022, 44, 147–156. [Google Scholar]
- Woolard, G.R.; Rathbone, E.B.; Novellie, L. Studies on the free sugars, water-soluble gums and hemicelluloses from barley grains and malts. J. Sci. Food Agric. 1977, 28, 469–476. [Google Scholar] [CrossRef]
- Liu, K.X.; Li, H.Q.; Zhang, J.; Zhang, Z.G.; Xu, J. The effect of non-structural components and lignin on hemicellulose extraction. Bioresour. Technol. 2016, 214, 755–760. [Google Scholar] [CrossRef]
- Olmedo, P.; Zepeda, B.; Rojas, B.; Silva-Sanzana, C.; Delgado-Rioseco, J.; Fernández, K.; Balic, I.; Arriagada, C.; Moreno, A.A.; Defilippi, B.G.; et al. Cell wall calcium and hemicellulose have a role in the fruit firmness during storage of blueberry (Vaccinium spp.). Plants 2021, 10, 553. [Google Scholar] [CrossRef]
- Brummell, D.A.; Harpster, M.H. Cell wall metabolism in fruit softening and quality and its manipulation in transgenic plants. Plant Mol. Biol. 2001, 47, 311–339. [Google Scholar] [CrossRef]
- Yang, H.; Yi, N.; Zhao, S.; Xiang, Z.; Qaseem, M.F.; Zheng, B.; Li, H.; Feng, J.X.; Wu, A.M. Characterization of hemicellulose in Cassava (Manihot esculenta Crantz) stem during xylogenesis. Carbohydr. Polym. 2021, 264, 118038. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S. The role of cell wall in plant resistance to nutritional stresses and the underlying physiological and molecular mechanisms. Sci. Sin. Vitae 2014, 44, 334–341. [Google Scholar] [CrossRef]
- Wan, J.X.; Zhu, X.F.; Wang, Y.Q.; Liu, L.Y.; Zhang, B.C.; Li, G.X.; Zhou, Y.H.; Zheng, S.J. Xyloglucan fucosylation modulates Arabidopsis cell wall hemicellulose aluminium binding capacity. Sci. Rep. 2018, 8, 428. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Jie, H.; Tang, Y.; Xing, H.; Jie, Y. The Role of Hemicellulose in Cadmium Tolerance in Ramie (Boehmeria nivea (L.) Gaud.). Plants 2022, 11, 1941. [Google Scholar] [CrossRef]
- Qaseem, M.F.; Shaheen, H.; Wu, A.M. Cell wall hemicellulose for sustainable industrial utilization. Renew. Sustain. Energy Rev. 2021, 144, 110996. [Google Scholar] [CrossRef]
- Yi, G.; Zhang, Y. One-Pot Selective Conversion of Hemicellulose (Xylan) to Xylitol under Mild Conditions. ChemSusChem 2012, 5, 1383–1387. [Google Scholar] [CrossRef]
- Palma, A.; Loaiza, J.M.; Díaz, M.J.; García, J.C.; Giráldez, I.; López, F. Tagasaste, leucaena and paulownia: Three industrial crops for energy and hemicelluloses production. Biotechnol. Biofuels 2021, 14, 89. [Google Scholar] [CrossRef]
- Gürbüz, E.I.; Gallo JM, R.; Alonso, D.M.; Wettstein, S.G.; Lim, W.Y.; Dumesic, J.A. Conversion of hemicellulose into furfural using solid acid catalysts in γ-valerolactone. Angew. Chem. Int. Ed. 2013, 52, 1270–1274. [Google Scholar] [CrossRef]
- Li, K.; Wang, H.; Hu, X.; Liu, Z.; Wu, Y.; Huang, C. Genome-wide association study reveals the genetic basis of stalk cell wall components in maize. PLoS ONE 2016, 11, e0158906. [Google Scholar] [CrossRef]
- Hernández-Espinosa, N.; Romano, G.P.; Crespo-Herrera, L.; Singh, R.; Guzmán, C.; Ibba, M.I. Endogenous arabinoxylans variability in refined wheat flour and its relationship with quality traits. J. Cereal Sci. 2020, 95, 103062. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Rehman, H.M.; Baloch, F.S.; Ijaz, B.; Ali, M.A.; Khan, I.A.; Lee, J.D.; Chung, G.; Yang, S.H. Genome and transcriptome-wide analyses of cellulose synthase gene superfamily in soybean. J. Plant Physiol. 2017, 215, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Colas, V.; Barre, P.; van Parijs, F.; Wolters, L.; Quitté, Y.; Ruttink, T.; Roldán-Ruiz, I.; Gutiérrez AJ, E.; Muylle, H. Seasonal differences in structural and genetic control of digestibility in perennial ryegrass. Front. Plant Sci. 2021, 12, 801145. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Li, J.; Wittkop, B.; Frauen, M.; Yan, X.; Liu, L.; Xiao, Y. QTL mapping for oil, protein, cellulose, and hemicellulose contents in seeds of Brassica napus L. Acta Agron. Sin. 2013, 39, 1214–1222. [Google Scholar] [CrossRef]
- Li, Q.; Fu, C.; Liang, C.; Ni, X.; Zhao, X.; Chen, M.; Ou, L. Crop Lodging and The Roles of Lignin, Cellulose, and Hemicellulose in Lodging Resistance. Agronomy 2022, 12, 1795. [Google Scholar] [CrossRef]
- Wang, H.Z. New-demand oriented oilseed rape industry developing strategy. Chin. J. Oil Crop Sci. 2018, 40, 613–617. [Google Scholar]
- Rashid, U.; Anwar, F. Production of biodiesel through optimized alkaline-catalyzed transesterification of rapeseed oil. Fuel 2008, 87, 265–273. [Google Scholar] [CrossRef]
- López-Linares, J.C.; Cara-Corpas, C.; Ruiz-Ramos, E.; Moya-Vilar, M.; Castro-Galiano, E.; Romero-Pulido, I. Hemicellulose-derived sugars solubilisation of rape straw. Cofermentation of pentoses and hexoses by Escherichia coli. Span. J. Agric. Res. 2015, 13, e0213. [Google Scholar] [CrossRef]
- Qaseem, M.F.; Wu, A.M. Marginal lands for bioenergy in China; an outlook in status, potential and management. GCB Bioenergy 2021, 13, 21–44. [Google Scholar] [CrossRef]
- Liu, J.; Wang, W.; Mei, D.; Wang, H.; Fu, L.; Liu, D.; Li, Y.; Hu, Q. Characterising variation of branch angle and genome-wide association mapping in rapeseed (Brassica napus L.). Front. Plant Sci. 7, 21.
- R Core Team. In R: A Language and Environment for Statistical Computing; R Core Team: Zurich, Switzerland, 2013.
- Wickham, H. Data analysis. In Ggplot2; Springer: Cham, Switzerland, 2016; pp. 189–201. [Google Scholar]
- Schloerke, B.; Crowley, J.; Cook, D.; Briatte, F.; Marbach, M.; Thoen, E.; Larmarange, J. GGally: Extension to ‘Ggplot2’; R Package Version, 1. 2018. Available online: https://cran.r-project.org/web/packages/GGally/index.html.
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef]
- Zhang, C.; Dong, S.S.; Xu, J.Y.; He, W.M.; Yang, T.L. PopLDdecay: A fast and effective tool for linkage disequilibrium decay analysis based on variant call format files. Bioinformatics 2019, 35, 1786–1788. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.D. qqman: An R package for visualizing GWAS results using QQ and manhattan plots. Biorxiv 2014, 005165. [Google Scholar] [CrossRef]
- Song, J.M.; Guan, Z.; Hu, J.; Guo, C.; Yang, Z.; Wang, S.; Liu, D.; Wang, B.; Lu, S.; Zhou, R.; et al. Eight high-quality genomes reveal pan-genome architecture and ecotype differentiation of Brassica napus. Nat. Plants. 2020, 6(1), 34–45. [Google Scholar] [CrossRef] [PubMed]
- Korf, I.; Yandell, M.; Bedell, J. Blast—O’reilly Media; Oreilly Media: Newton, MA, USA, 2003. [Google Scholar]
- Bischoff, V.; Nita, S.; Neumetzler, L.; Schindelasch, D.; Urbain, A.; Eshed, R.; Persson, S.; Deborah, D.; Scheible, W.R. TRICHOME BIREFRINGENCE and its homolog AT5G01360 encode plant-specific DUF231 proteins required for cellulose biosynthesis in Arabidopsis. Plant Physiol. 2010, 153, 590–602. [Google Scholar] [CrossRef]
- Avanthi, A.; Kumar, S.; Sherpa, K.C.; Banerjee, R. Bioconversion of hemicelluloses of lignocellulosic biomass to ethanol: An attempt to utilize pentose sugars. Biofuels 2017, 8, 431–444. [Google Scholar] [CrossRef]
- Luo, Y.; Li, Z.; Li, X.; Liu, X.; Fan, J.; Clark, J.H.; Hu, C. The production of furfural directly from hemicellulose in lignocellulosic biomass: A review. Catal. Today 2019, 319, 14–24. [Google Scholar] [CrossRef]
- Wang, N.; Feng, J.; Li, L.; Liu, J.; Sun, Y. Rapid Determination of Cellulose and Hemicellulose Contents in Corn Stover Using Near-Infrared Spectroscopy Combined with Wavelength Selection. Molecules 2022, 27, 3373. [Google Scholar] [CrossRef]
- Samadi, A.F.; Suzuki, H.; Ueda, T.; Yamamoto, T.; Adachi, S.; Ookawa, T. Identification of quantitative trait loci for breaking and bending types lodging resistance in rice, using recombinant inbred lines derived from Koshihikari and a strong culm variety, Leaf Star. Plant Growth Regul. 2019, 89, 83–98. [Google Scholar] [CrossRef]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and biological functions in plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef]
- Sorieul, M.; Dickson, A.; Hill, S.J.; Pearson, H. Plant fibre: Molecular structure and biomechanical properties, of a complex living material, influencing its deconstruction towards a biobased composite. Materials 2016, 9, 618. [Google Scholar] [CrossRef] [PubMed]
- Amaral, L.I.; Cortelazzo, Â.L.; Buckeridge, M.S.; Pereira, L.A.; Pereira, M.F. Anatomical and biochemical changes in the composition of developing seed coats of annatto (Bixa orellana L.). Trees 2009, 23, 287–293. [Google Scholar] [CrossRef]
- Lunin, V.V.; Wang, H.T.; Bharadwaj, V.S.; Alahuhta, M.; Peña, M.J.; Yang, J.Y.; Archer-Hartmann, S.A.; Azadi, P.; Himmel, M.E.; Moremen, K.W.; et al. Molecular mechanism of polysaccharide acetylation by the Arabidopsis xylan O-acetyltransferase XOAT1. Plant Cell 2020, 32, 2367–2382. [Google Scholar] [CrossRef]
- Ohtani, M.; Ramachandran, V.; Tokumoto, T.; Takebayashi, A.; Ihara, A.; Matsumoto, T.; Hiroyama, R.; Nishikubo, N.; Demura, T. Identification of novel factors that increase enzymatic saccharification efficiency in Arabidopsis wood cells. Plant Biotechnol. 2018, 34, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Zhong, R.; Richardson, E.A.; Himmelsbach, D.S.; McPhail, B.T.; Ye, Z.H. The PARVUS gene is expressed in cells undergoing secondary wall thickening and is essential for glucuronoxylan biosynthesis. Plant Cell Physiol. 2007, 48, 1659–1672. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Xiao, X.; Asjad, A.; Han, D.; Zheng, W.; Xiao, G.; Huang, Y.; Zhou, Q. Integration of GWAS and transcriptome analyses to identify SNPs and candidate genes for aluminum tolerance in rapeseed (Brassica napus L.). BMC Plant Biol. 2022, 22, 130. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Lee, J.; Miura, T.; Hasegawa, P.M. SIZ1 controls cell growth and plant development in Arabidopsis through salicylic acid. Plant Cell Physiol. 2010, 51, 103–113. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | Site | Min (%) | Max (%) | Amplitude (%) | Average (%) | Variance | Skewness | Kurtosis | G | E | G × E |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Stalk | Zunyi | 12.56 | 20.75 | 8.19 | 16.61 | 2.88 | 0.00 | −0.09 | 0.000 ** | 0.039 * | 0.758 |
| Yangluo | 12.02 | 18.68 | 6.66 | 15.53 | 1.87 | −0.18 | −0.32 | ||||
| Lu’an | 14.29 | 20.27 | 5.98 | 16.67 | 1.00 | 0.59 | 1.22 | ||||
| Taproot | Zunyi | 11.98 | 20.22 | 8.24 | 16.56 | 2.83 | −0.06 | −0.39 | 0.001 ** | 0.002 ** | 0.025 * |
| Yangluo | 11.11 | 18.93 | 7.83 | 15.21 | 2.86 | −0.07 | −0.59 | ||||
| Lu’an | 13.86 | 18.86 | 5.00 | 16.13 | 1.18 | 0.28 | −0.29 |
| Position | Correlation Interval | Originate | Terminate | PeakSNP | −log10(p) | Contribution Rate |
|---|---|---|---|---|---|---|
| Stalk | qHCs.C02 | 56,745,960 | 57,991,329 | Bn-scaff_22144_1-p207843 | 3.84 | 17.20% |
| qHCs.C05 | 1,242,999 | 9,322,094 | Bn-scaff_16045_1-p235411 | 4.01 | 10.62% | |
| qHCs.C08 | 46,627,013 | 46,843,716 | Bn-scaff_16197_1-p2621761 | 3.72 | 8.80% | |
| Taproot | qHCt.A09 | 6,620,400 | 7,024,760 | Bn-A09-p5255753 | 3.44 | 9.49% |
| qHCt.C05 | 1,242,999 | 1,243,397 | Bn-scaff_16414_1-p1114696 | 3.35 | 9.18% | |
| qHCt.C08 | 22,468,415 | 22,468,464 | Bn-scaff_16545_1-p80303 | 3.07 | 13.10% |
| Candidate Gene | Originate | Terminate | Arabidopsis Homologous Gene | Gene Name | Functional Annotation |
|---|---|---|---|---|---|
| BnaC02G0468200ZS | 57,411,375 | 57,412,944 | AT3G28150 | AXY4L | Involved in the synthesis and deposition of secondary wall cellulose |
| BnaC05G0014900ZS | 1,016,442 | 1,019,787 | AT1G02640 | BXL2 | Encodes a protein similar to a beta-xylosidase |
| BnaC05G0092200ZS | 5,255,987 | 5,257,234 | AT1G11545 | XTH8 | Xyloglucan endotransglucosylase/hydrolase 8 |
| BnaC05G0112100ZS | 6,929,662 | 6,931,370 | AT1G14080 | FUT6 | Encodes an alpha-(1,2)-fucosyltransferase |
| BnaC05G0112200ZS | 6,932,039 | 6,934,614 | AT1G14080 | FUT6 | Encodes an alpha-(1,2)-fucosyltransferase. |
| BnaC05G0112300ZS | 6,937,873 | 6,939,470 | AT1G14100 | FUT8 | Member of glycosyltransferase Family- 37. |
| BnaC05G0112400ZS | 6,944,599 | 6,946,246 | AT1G14100 | FUT8 | Member of glycosyltransferase Family- 37 |
| BnaC05G0156900ZS | 10,051,053 | 10,052,411 | AT1G18690 | GMA12 | Galactosyl transferase GMA12/MNN10 family protein |
| BnaC05G0161900ZS | 10,585,900 | 10,586,958 | AT1G19300 | GLZ | Encodes a putative family 8 glycosyl transferase that contributes to xylan biosynthesis |
| BnaC08G0415400ZS | 46,598,950 | 46,599,504 | AT3G62720 | XT1, XXT1 | Encodes a protein with xylosyltransferase activity |
| BnaC08G0116500ZS | 21,355,187 | 21,356,233 | AT4G14130 | XTH15, XTR7 | Xyloglucan endotransglycosylase-related protein |
| Candidate Genes | Homologs in Arabidopsis | Gene Annotation | Log2FC |
|---|---|---|---|
| BnaC02G0468300ZS | AT3G28180 | Encodes a gene similar to cellulose synthase | −4.71 |
| BnaC02G0470400ZS | AT3G28740 | Encodes a member of the cytochrome p450 family | 7.48 |
| BnaC05G0070100ZS | AT1G09310 | ABA responsive trichome formation regulator | −7.73 |
| BnaC05G0089100ZS | AT1G11080 | Serine carboxypeptidase-like 31 | 7.77 |
| BnaC05G0092200ZS | AT1G11545 | Xyloglucan endotransglucosylase/hydrolase 8 | −2.99 |
| BnaC05G0112400ZS | AT1G14100 | Member of Glycosyltransferase Family- 37 | −3.68 |
| BnaC05G0121900ZS | AT3G30739 | Hypothetical protein | 7.30 |
| BnaC05G0136700ZS | AT1G16410 | Member of CYP79F, Encodes cytochrome P450 | −10.74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Yang, Y.; Yu, W.; Liu, L.; Hu, Q.; Wei, W.; Liu, J. Dissecting the Genetic Mechanisms of Hemicellulose Content in Rapeseed Stalk. Agronomy 2022, 12, 2886. https://doi.org/10.3390/agronomy12112886
Xu Y, Yang Y, Yu W, Liu L, Hu Q, Wei W, Liu J. Dissecting the Genetic Mechanisms of Hemicellulose Content in Rapeseed Stalk. Agronomy. 2022; 12(11):2886. https://doi.org/10.3390/agronomy12112886
Chicago/Turabian StyleXu, Yinhai, Yuting Yang, Wenkai Yu, Liezhao Liu, Qiong Hu, Wenliang Wei, and Jia Liu. 2022. "Dissecting the Genetic Mechanisms of Hemicellulose Content in Rapeseed Stalk" Agronomy 12, no. 11: 2886. https://doi.org/10.3390/agronomy12112886
APA StyleXu, Y., Yang, Y., Yu, W., Liu, L., Hu, Q., Wei, W., & Liu, J. (2022). Dissecting the Genetic Mechanisms of Hemicellulose Content in Rapeseed Stalk. Agronomy, 12(11), 2886. https://doi.org/10.3390/agronomy12112886