
Can Intensified Pasture Systems Reduce Enteric Methane Emissions from Beef Cattle in the Atlantic Forest Biome?

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Systems Establishment and Management
2.2. Animals Management
2.3. Slaughter and Carcass Yield
2.4. Forage Production and Quality
2.5. Pasture Intake
2.6. Enteric Methane Emissions
2.7. Statistical Analysis
3. Results
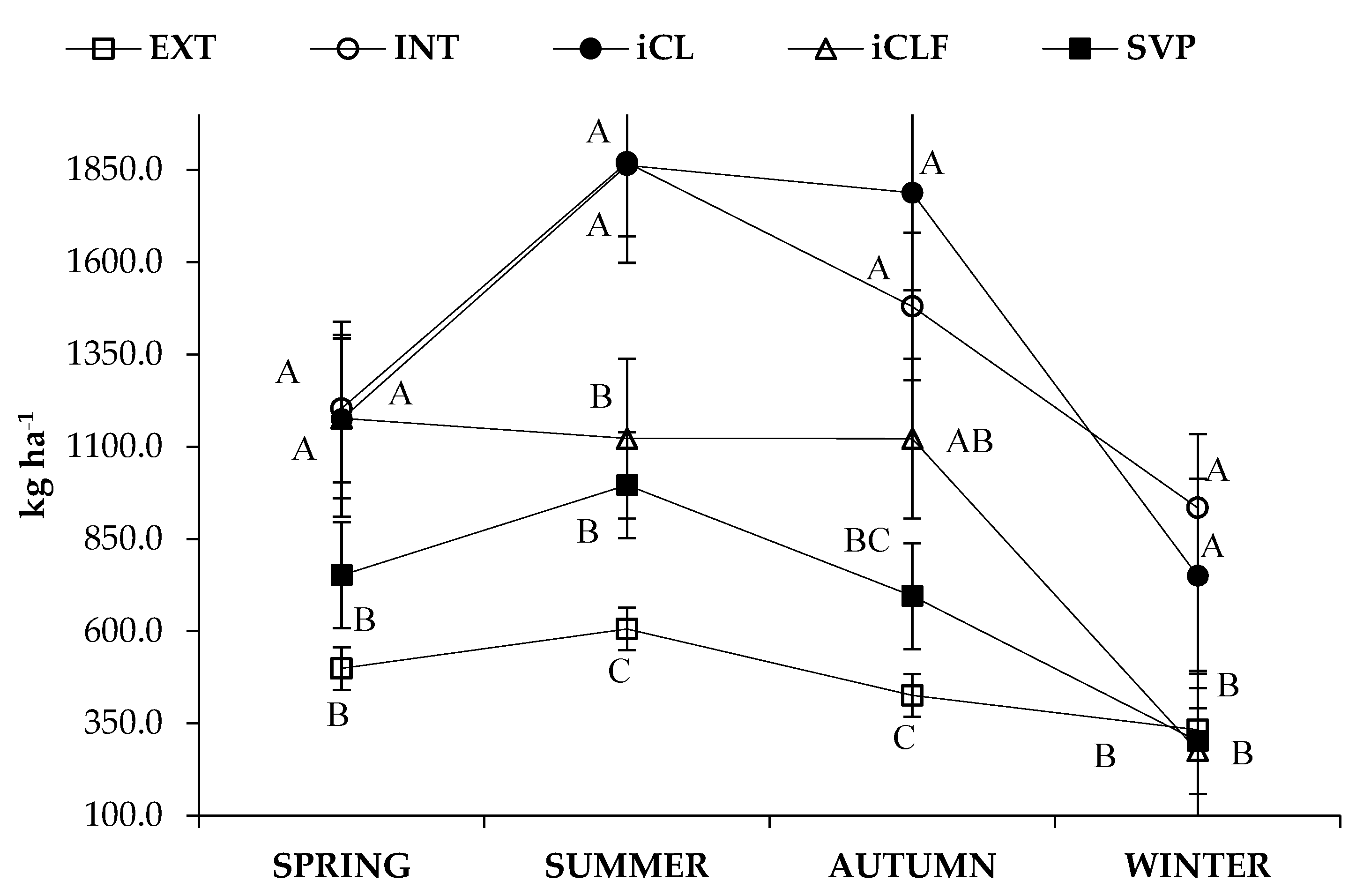
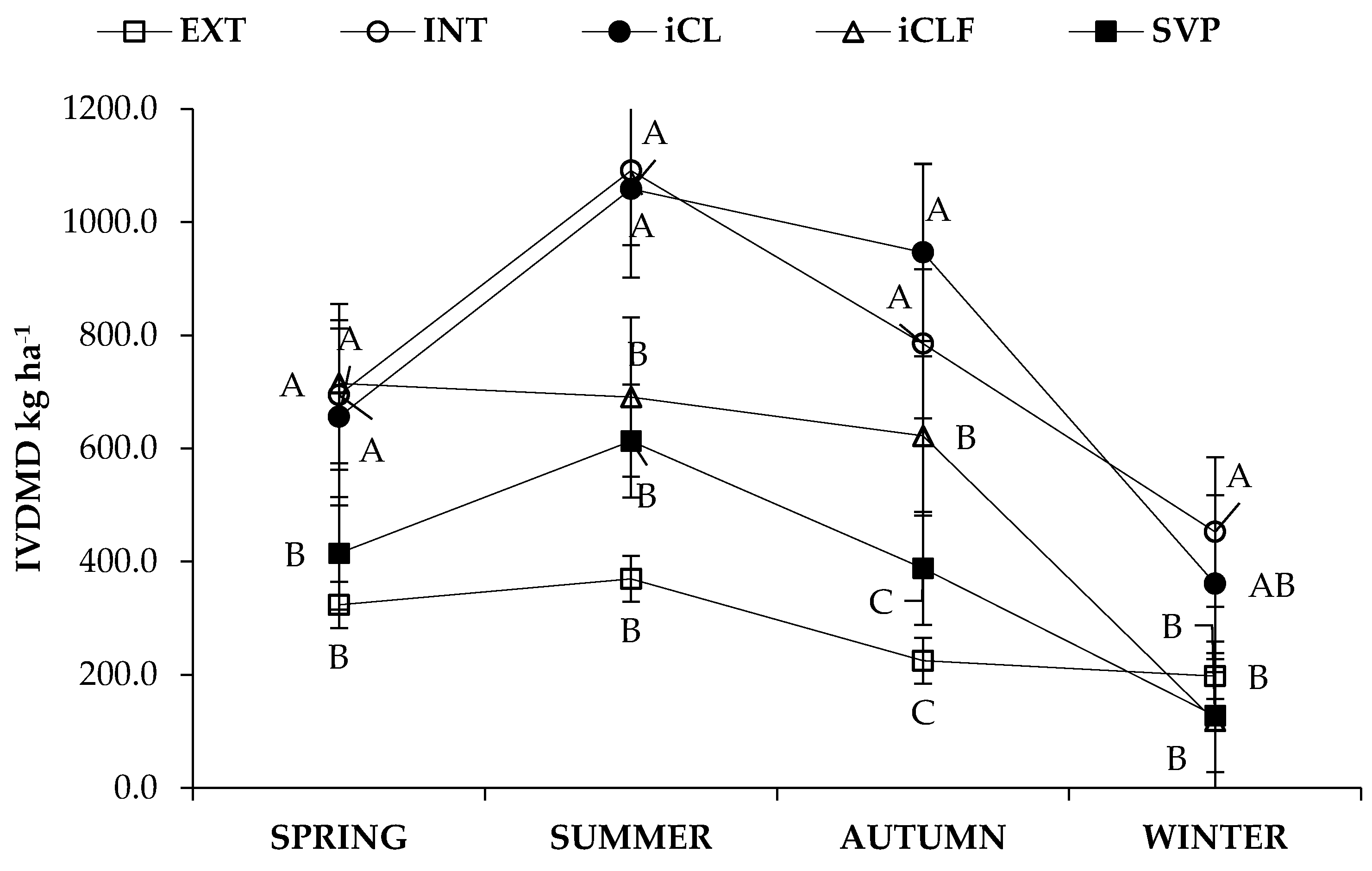
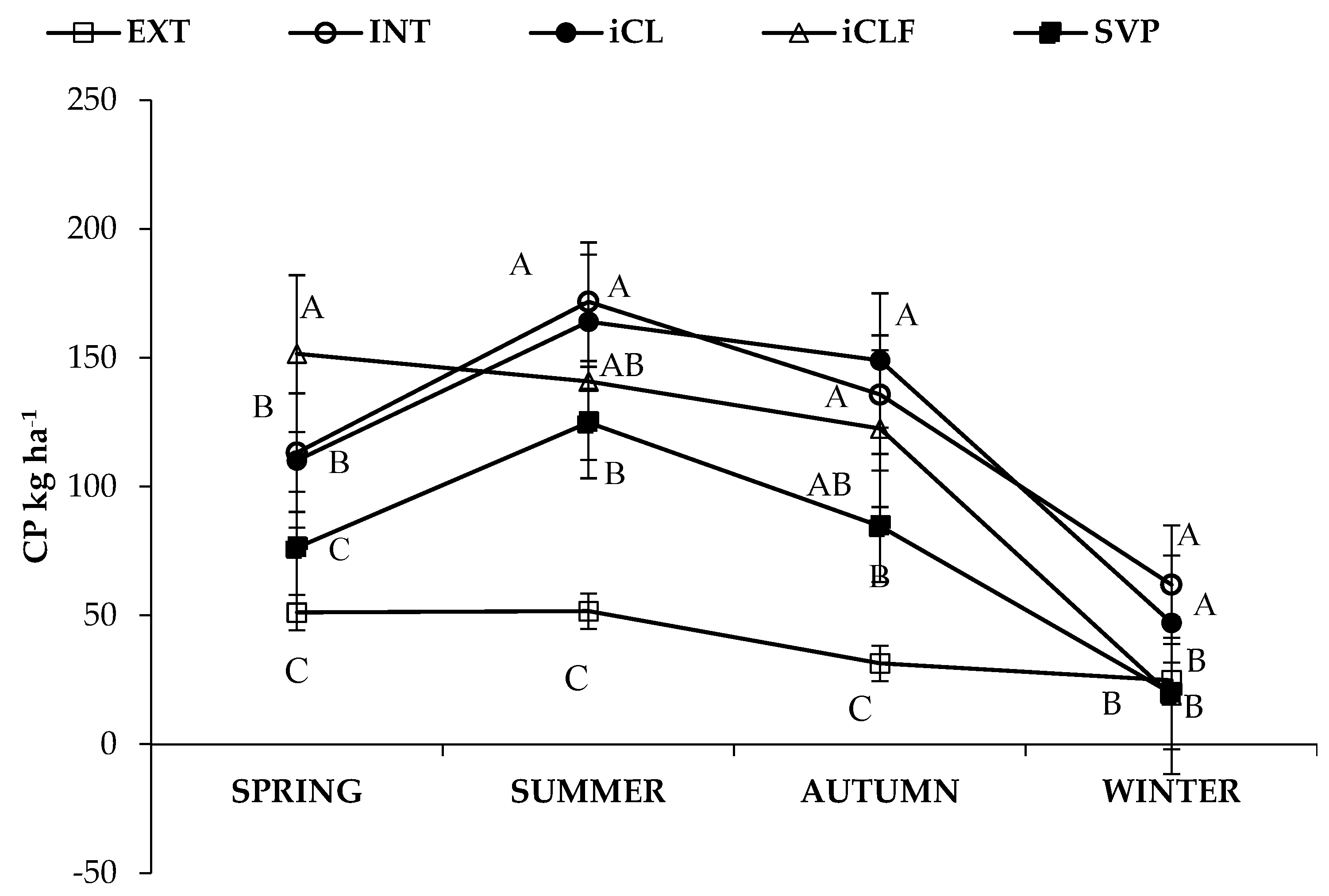
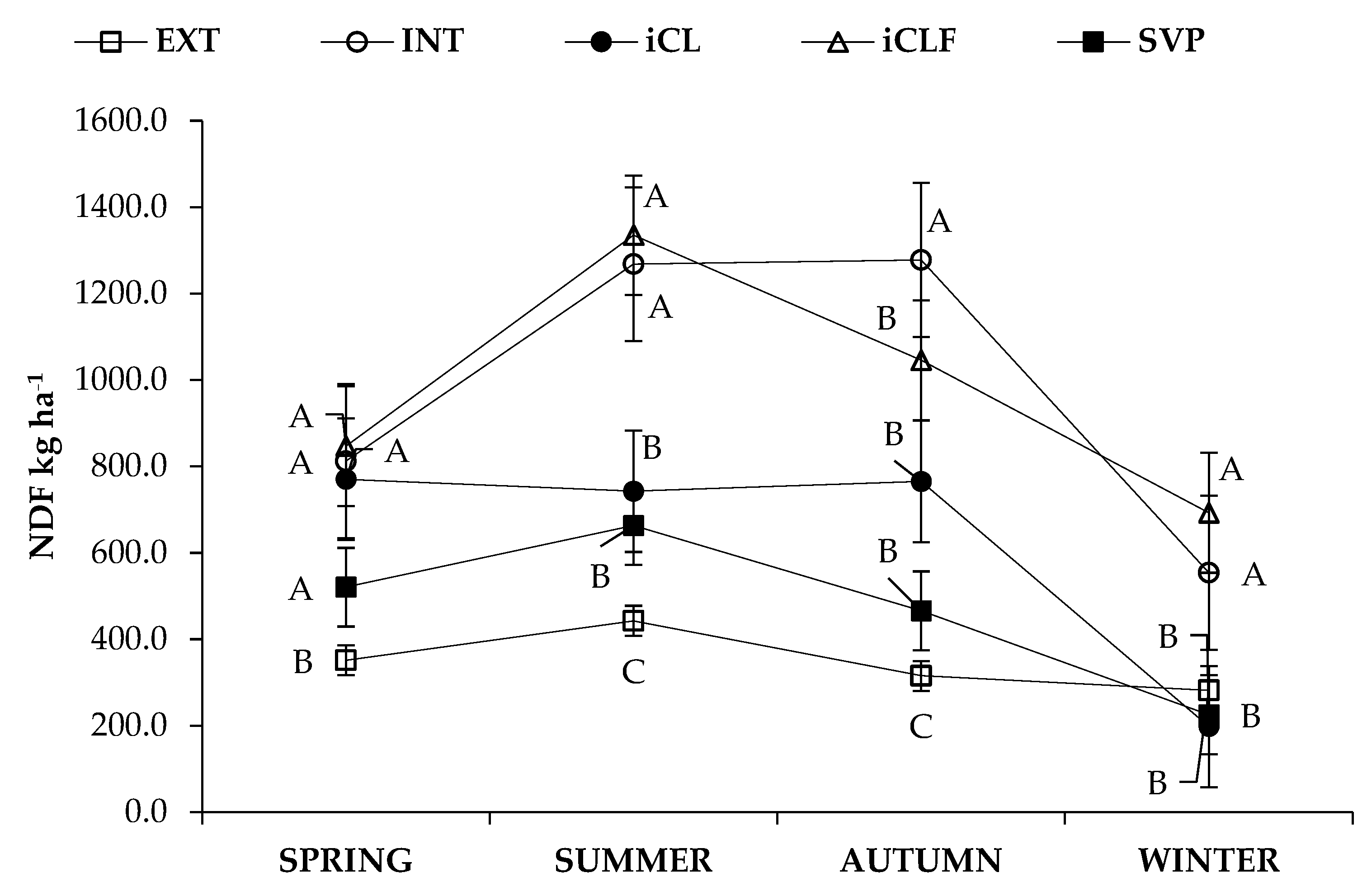
3.1. Forage Mass, In Vitro Digestibility and Nutrient Availability
3.2. Animal Performance and Methane Emissions
4. Discussion
4.1. Forage Mass, In Vitro Digestibility and Nutrient Availability
4.2. Animal Performance
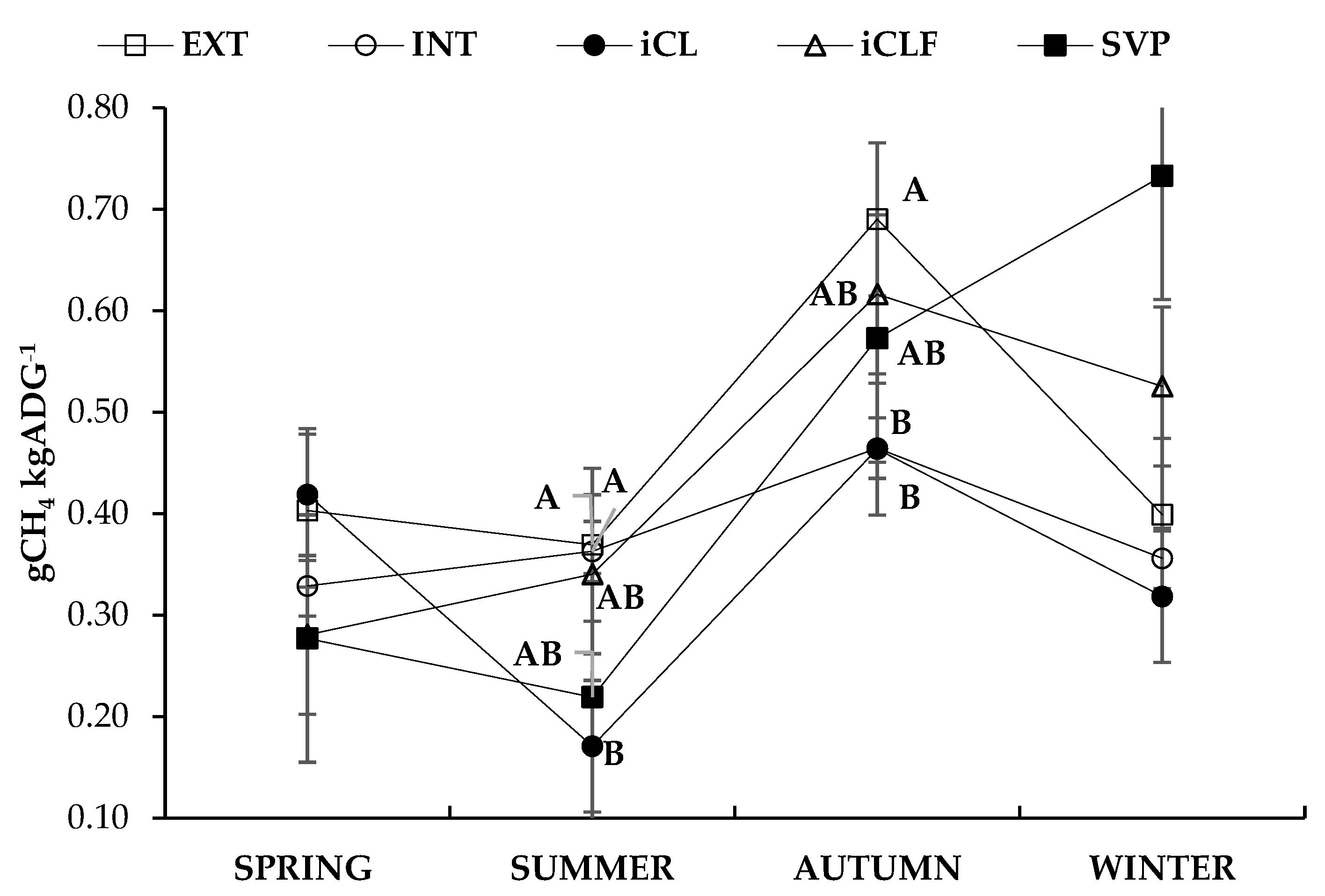
4.3. Emission of Enteric Methane
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A









References
- United Nations—U.N. World Population Prospects 2019. Department of Economic and Social Affairs—Population Dynamics. Available online: https://population.un.org/wpp/ (accessed on 1 July 2020).
- Prochnow, A.; Heiermann, M.; Plöchl, M.; Amon, T.; Hobbs, P.J. Bioenergy from permanent grassland–A review: 2. Combustion. Bioresour. Technol. 2009, 100, 4945–4954. [Google Scholar] [CrossRef] [PubMed]
- Gerssen-Gondelach, S.J.; Lauwerijssen, R.B.; Havlík, P.; Herrero, M.; Valin, H.; Faaij, A.P.; Wicke, B. Intensification pathways for beef and dairy cattle production systems: Impacts on GHG emissions, land occupation and land use change. Agric. Ecosyst. Environ. 2017, 240, 135–147. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Emissions due to agriculture. In Global, Regional and Country Trends 2000–2018; FAOSTAT Analytical Brief Series No 18; FAO: Rome, Italy, 2020. [Google Scholar]
- Werth, S.; CLEAR Center—Clarity and Leadership for Environmental Awareness and Research at UC Davis. The Biogenic Carbon Cycle and Cattle. 2020. Available online: https://clear.ucdavis.edu/explainers/biogenic-carbon-cycle-and-cattle (accessed on 5 July 2020).
- Soussana, J.F.; Tallec, T.; Blanfort, V. Mitigating the greenhouse gas balance of ruminant production systems through carbon sequestration in grasslands. Animal 2010, 4, 334–350. [Google Scholar] [CrossRef]
- Empresa Brasileira de Pesquisa Agropecuária—EMBRAPA. Integrated Crop-Livestock-Forestry Systems. Available online: https://www.embrapa.br/en/tema-integracao-lavoura-pecuaria-floresta-ilpf (accessed on 9 September 2022).
- Empresa Brasileira de Pesquisa Agropecuária—EMBRAPA. Integração Lavoura Pecuária Floresta—ILPF. Available online: https://www.embrapa.br/tema-integracao-lavoura-pecuaria-floresta-ilpf/nota-tecnica (accessed on 9 September 2018).
- Empresa Brasileira de Pesquisa Agropecuária—EMBRAPA. Integração Lavoura Pecuária—ILP. Available online: https://www.embrapa.br/busca-de-solucoes-tecnologicas/-/produto-servico/1055/sistema-integracao-lavoura-pecuaria (accessed on 9 September 2018).
- Costa, M.P.; Schoeneboom, J.C.; Oliveira, S.A.; Viñas, R.S.; de Medeiros, G.A. A socio-eco-efficiency analysis of integrated and non-integrated crop-livestock-forestry systems in the Brazilian Cerrado based on LCA. J. Clean. Prod. 2017, 171, 1460–1471. [Google Scholar] [CrossRef]
- De Moraes, A.; Carvalho, P.C.F.; Anghinoni, I.; Lustosa, S.B.C.; Andrade, S.E.V.G.; Kunrath, T.R. Integrated crop–livestock systems in the Brazilian subtropics. Eur. J. Agron. 2014, 57, 4–9. [Google Scholar] [CrossRef]
- Savian, J.V.; Barth Neto, A.; de David, D.B.; Bremm, C.; Schons, R.M.T.; Genro, T.C.M.; do Amaral, G.A.; Gere, J.; McManus, C.M.; Bayer, C.; et al. Grazing intensity and stocking methods on animal production and methane emission by grazing sheep: Implications for integrated crop–livestock system. Agric. Ecosyst. Environ. 2014, 190, 112–119. [Google Scholar] [CrossRef]
- Pontes, L.S.; Barro, R.S.; Savian, J.V.; Berndt, A.; Moletta, J.L.; Porifício-Da -Silva, V.; Bayer, C.; Carvalho, P.C.F. Performance and methane emissions by beef heifer grazing in temperate pastures and in integrated crop-livestock systems: The effect of shade and nitrogen fertilization. Agric. Ecosyst. Environ. 2018, 253, 90–97. [Google Scholar] [CrossRef]
- Raij, B.V.; Cantarella, H.; Quaggio, J.A.; Furlani, A.M.C. Recommendations for Fertilization and Liming for the State of Sao Paulo; Boletim Técnico; IAC: Campinas, Brazil, 1996; Volume 100. [Google Scholar]
- Mott, G.O.; Lucas, H.L. The design, conduct and interpretation of grazing trials on cultivated and improved pastures. In Proceedings of the International Grassland Congress, State College, PA, USA, 17–23 August 1952. [Google Scholar]
- RIISPOA. Regulamento da Inspeção Industrial e Sanitária de Produtos de Origem Animal (RIISPOA); Brasília-DF. anexos e sala de matança; RIISPOA: Brasília, Brazil, 1971. [Google Scholar]
- Pezzopane, J.R.M.; Bernardi, A.C.C.; Bosi, C.; Oliveira, P.P.A.; Marconato, M.H.; Pedroso, A.F.; Esteves, S.N. Forage productivity and nutritive value during pasture renovation in integrated systems. Agrofor. Syst. 2019, 93, 39–49. [Google Scholar] [CrossRef]
- Oliveira, P.P.A. Protocolo Recomendado para Avaliação do Consumo Voluntário de Animais em Pastejo; Documentos (INFOTECA-E); Embrapa Pecuária Sudeste: São Carlos, Brazil, 2014; p. 14. [Google Scholar] [CrossRef]
- Carvalho, P.C.F.; Kozloski, G.V.; Ribeiro Filho, H.M.N.; Reffatti, M.V.; Genro, T.C.M.; Euclides, V.P.B. Avanços metodológicos na determinação do consumo de ruminantes em pastejo. R. Bras. Zootec. 2007, 36, 151–170. [Google Scholar] [CrossRef]
- Myers, W.D.; Ludden, P.A.; Nayihigihugu, V.; Hess, B. Technical Note: A procedure for the preparation and quantitative analysis of samples for titanium dioxide. J. Anim. Sci. 2004, 82, 179–183. [Google Scholar] [CrossRef]
- Valente, T.N.P.; Detmann, E.; Valadares-Filho, S.C.; Cunha, M.; Queiroz, A.C.; Sampaio, C.B. In situ estimation of indigestible compounds contents in cattle feed and feces using bags made from different textiles. R. Bras. Zootec. 2011, 40, 666–675. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Cook, C.W. Symposium on nutrition of forages and pastures: Collecting forage samples representative of ingested material of grazing animals for nutritional studies. J. Anim. Sci. 1964, 23, 265–270. [Google Scholar] [CrossRef]
- Johnson, K.; Hurley, M.; Westberg, H.; Lamb, B.; Zimmerman, P. Measurement of methane emissions from ruminant livestock using a sulfur hexafluoride tracer technique. Environ. Sci. Technol. 1994, 28, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Primavesi, O.; Berndt, A.; Lima, M.A.; Frighetto, R.T.S.; Demarchi, J.J.A.A.; Pedreira, M.S.; Berchielli, T.T.; Oliveira, S.G. Greenhouse gas production in agricultural systems: Groundwork for an inventory of methane emissions by ruminants. In Carbon Stocks and Greenhouse Gas Emissions in Brazilian Agriculture; Boddey, R.M., Lima, M.A., Alves, B.J.R., Machado, P.L.O.A., Urquiaga, S., Eds.; Empresa Brasileira de Pesquisa Agropecuária (EMBRAPA): Brasília, Brazil, 2014; pp. 191–216. [Google Scholar]
- Berndt, A.; Boland, T.M.; Deighton, M.H.; Gere, J.I.; Grainger, C.; Hegarty, R.S.; Iwaasa, A.D.; Koolaard, J.P.; Lassey, K.R.; Luo, D.; et al. Guidelines for Use of Sulphur Hexafluoride (SF6) Tracer Technique to Measure Enteric Methane Emissions from Ruminants; Lambert, M.G., Ed.; New Zealand Agricultural Greenhouse Gas Research Centre: Wellington, New Zealand, 2014. [Google Scholar]
- Johnson, K.A.; Johnson, D.E. Methane emissions from cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Goonewardene, L.A. The Use of MIXED Models in the Analysis of Animal Experiments with Repeated Measures Data. Can. J. Anim. Sci. 2004, 84, 1–11. [Google Scholar] [CrossRef]
- Deiss, L.; Moraes, A.D.; Pelissari, A.; Skora Neto, F.; Silva, V.P.D.; Andreolla, V.R.M. Oat growth under different nitrogen doses in eucalyptus alley cropping system in subtropical Brazil. Cienc. Agron. 2014, 45, 1014–1023. [Google Scholar] [CrossRef][Green Version]
- Oliveira, C.C.D.; Villela, S.D.J.; Almeida, R.G.D.; Alves, F.V.; Behling-Neto, A.; Martins, P.G.M.D.A. Performance of Nellore heifers, forage mass, and structural and nutritional characteristics of Brachiaria brizantha grass in integrated production systems. Trop. Anim. Health Prod. 2014, 46, 167–172. [Google Scholar] [CrossRef]
- Santos, D.C.; Júnior, R.G.; Vilela, L.; Maciel, G.A.; França, A.F.S. Implementation of silvopastoral systems in Brazil with Eucalyptus urograndis and Brachiaria brizantha: Productivity of forage and anexploratory test of the animal response. Agric. Ecosyst. Environ. 2018, 266, 174–180. [Google Scholar] [CrossRef]
- Associação Brasileira das Indústrias Exportadoras de Carne—ABIEC. Beef Report: Perfil da Pecuária No Brasil. Available online: http://abiec.com.br/publicacoes/beef-report-2020/ (accessed on 9 February 2022).
- Sousa, L.F.; Mauricio, R.M.; Moreira, G.R.; Goncalves, L.C.; Borges, I.; Pereira, L.G.R. Nutritional evaluation of “Braquiarão" grass in association with "Aroeira" trees in a silvopastoral system. Agrofor. Syst. 2010, 79, 189–199. [Google Scholar] [CrossRef]
- Coêlho, J.J.; Mello, A.C.L.; Santos, M.V.F.; Dubeux Junior, J.C.B.; Cunha, M.V.; Lira, M.A. Prediction of the nutritional value of grass species in the semiarid region by repeatability analysis. Pesqui. Agropecu. Bras. 2018, 53, 378–385. [Google Scholar] [CrossRef]
- Paciullo, D.S.C.; Carvalho, C.A.B.; Aroeira, L.J.M.; Morenz, M.J.F.; Lopes, F.C.F.; Rossiello, R.O.P. Morfofisiologia e valor nutritivo do capim-braquiária sob sombreamento natural e a sol pleno. Pesqui. Agropecu. Bras. 2007, 42, 573–579. [Google Scholar] [CrossRef]
- Belesky, D.P.; Burner, D.M.; Ruckle, J.M. Tiller production in cocksfoot (Dactylis glomerata) and tall fescue (Festuca arundinacea) growing along a light gradient. Grass Forage Sci. 2011, 66, 370–380. [Google Scholar] [CrossRef]
- Pandey, C.B.; Verma, S.K.; Dagar, J.C.; Srivastava, R.C. Forage production and nitrogen nutrition in three grasses under coconut tree shades in the humid-tropics. Agrofor. Syst. 2011, 83, 1–12. [Google Scholar] [CrossRef]
- Paciullo, D.S.C.; Castro, C.R.T.; Gomide, C.A.M.; Maurício, R.M.; Pires, A.F.A.; Müler, M.D.; Xavier, D.F. Performance of dairy heifers in a silvopastoral system. J. Livest. Sci. 2011, 141, 166–172. [Google Scholar] [CrossRef]
- Azevêdo, J.A.G.; Valadares Filho, S.C.; Pina, D.S.; Valadares, R.F.D.; Detmann, E. Predição de consumo de matéria seca por bovinos de corte em confinamento. In Exigências Nutricionais de Zebuínos Puros e Cruzados: BR-Corte; Valadares, S.C., Marcondes, M.I., Chizzotti, M.L., Paulino, P.V.R., Eds.; UFV: Vicosa, Brasil, 2010; pp. 47–64. [Google Scholar]
- National Research Council (NRC). Nutrient Requirements of Beef Cattle, 8th ed.; The National Academy Press: Washington, DC, USA, 2016. [Google Scholar]
- Oliveira, P.P.A.; Corte, R.R.S.; Silva, S.L.; Rodriguez, P.H.M.; Sakamoto, L.S.; Pedroso, A.F.; Tullio, R.R.; Berndt, A. The effect of grazing system intensification on the growth and meat quality of beef cattle in the Brazilian Atlantic Forest biome. Meat. Sci. 2018, 139, 157–161. [Google Scholar] [CrossRef]
- Sakamoto, L.S. Intensidades de Emissão de Gás Metano de Bovinos Nelore Terminados a Pasto e Cruzados em Confinamento. Ph.D. Thesis, Faculdade de Zootecnia e Engenharia de Alimentos, Universidade de São Paulo, Pirassununga, Brazil, 2018. Available online: https://teses.usp.br/teses/disponiveis/74/74131/tde-23102018-124307/en.php (accessed on 5 February 2020). [CrossRef]
- Lopes, L.B.; Eckstein, C.; Pina, D.S.; Carnevalli, R.A. The influence of trees on the thermal environment and behaviour of grazing heifers in Brazilian Midwest. Trop. Anim. Health. Pro. 2016, 48, 755–761. [Google Scholar] [CrossRef]
- Salton, J.C.; Mercante, F.M.; Tomazi, M.; Zanatta, J.A.; Concenço, G.; Silva, W.M.; Retore, M. Integrated crop-livestock system in tropical Brazil: Toward a sustainable production system. Agric. Ecosyst. Environ. 2014, 152, 70–79. [Google Scholar] [CrossRef]
- IPCC. Guidelines for National Greenhouse Gas Inventories; Eggleston, H.S., Buendia, L., Miwa, K., Ngara, T., Tanabe, K., Eds.; Prepared by the National Greenhouse Gas Inventories Programme; Institute for Global Environmental Strategies: Hayama, Japan, 2006. [Google Scholar]
- Buddle, B.M.; Denis, M.; Attwood, G.T.; Alterman, E.; Janssen, P.H.; Ronimus, R.S.; Pinares-Patiño, C.S.; Muetzel, S.; Wedlock, D.N. Strategies to reduce methane emissions from farm ruminants grazing on pasture. Vet. J. 2011, 188, 11–17. [Google Scholar] [CrossRef]
- Clark, H.; Kelliher, F.; Pinares-Patiño, C. Reducing CH4 emissions from grazing ruminants in New Zealand: Challenges and opportunities. Asian-Australas. J. Anim. Sci. 2011, 24, 295–302. [Google Scholar] [CrossRef]
- Pinares-Patiño, C.S.; Franco, F.E.; Molano, G.; Kjestrup, H.; Sandoval, E.; Maclean, S.; Battistotti, M.; Koolaard, J.; Laubach, J. Feed intake and methane emissions from cattle grazing pasture sprayed with canola oil. J. Livest. Sci. 2016, 184, 7–12. [Google Scholar] [CrossRef]
- Mercadante, M.E.Z.; Caliman, A.P.M.; Canesin, R.C.; Bonilha, S.F.M.; Berndt, A.; Frighetto, R.T.S.; Magnani, E.; Branco, R.H. Relationship between residual feed intake and enteric methane emission in Nellore cattle. R. Bras. Zootec. 2015, 44, 255–262. [Google Scholar] [CrossRef]
- Oliveira Silva, R.; Barioni, L.G.; Hall, J.J.; Moretti, A.C.; Veloso, R.F.; Alexander, P.; Crespolini, M.; Moran, D. Sustainable intensification of Brazilian livestock production through optimized pasture restoration. Agric. Syst. 2017, 153, 201–211. [Google Scholar] [CrossRef]
- Cardoso, A.S.; Berndt, A.; Leytem, A.; Alves, B.J.R.; De Carvalho, I.N.O.; Soares, L.H.B.; Urquiaga, S.; Boddey, R.M. Impact of the intensification of beef production in Brazil on greenhouse gas emissions and land use. Agric. Syst. 2016, 143, 86–96. [Google Scholar] [CrossRef]
- Oliveira Silva, R.; Barioni, L.G.; Hall, J.A.J.; Folegatti, M.M.; Albertini, T.Z.; Fernandes, F.A.; Moran, D. Increasing beef production could lower greenhouse gas emissions in Brazil if decoupled from deforestation. Nat. Clim. Chang. 2016, 6, 493–497. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pasture System * | |||||
|---|---|---|---|---|---|
| iCLF | iCL | iLF | EXT | INT | |
| Pasture area: | 2.7 ha | 3.2 ha | 2.7 ha | 2.9 ha | 2.9 ha |
| Number of paddocks: | 6 | 6 | 6 | 1 | 6 |
| Pasture area during the maize cropping: | 1.8 ha | 2.1 ha | - | - | - |
| Grazing method: | Rotational | Rotational | Rotational | Continuous | Rotational |
| Stocking rate: | Variable | Variable | Variable | Variable | Variable |
| Soil management: | Yes | Yes | Yes | No | Yes |
| Integrated crops: | Eucalyptus + Maize | Maize | Eucalyptus | - | - |
| Spring–Summer | Autumn–Winter | ||
|---|---|---|---|
| Mineral | Content (%) 1 | Mineral | Content (%) 1 |
| Phosphor | 13.0 | Phosphor | 1.2 |
| Calcium | 21.4 | Calcium | 8.0 |
| Magnesium | 2.3 | Magnesium | 0.2 |
| Sulfur | 4.0 | Sulfur | 1.5 |
| Zinc | 1.0 | Zinc | 0.064 |
| Copper | 0.35 | Copper | 0.017 |
| Manganese | 0.17 | Manganese | 0.013 |
| Cobalt | 0.03 | Cobalt | 0.001 |
| Iodine | 0.03 | Iodine | 0.012 |
| Fluorine | 0.003 | Fluorine | 0.02 |
| Selenium | 0.13 | Selenium | 0.0003 |
| - | - | Crude protein | 40.0 |
| Fixed Effects | Variables | |||||
|---|---|---|---|---|---|---|
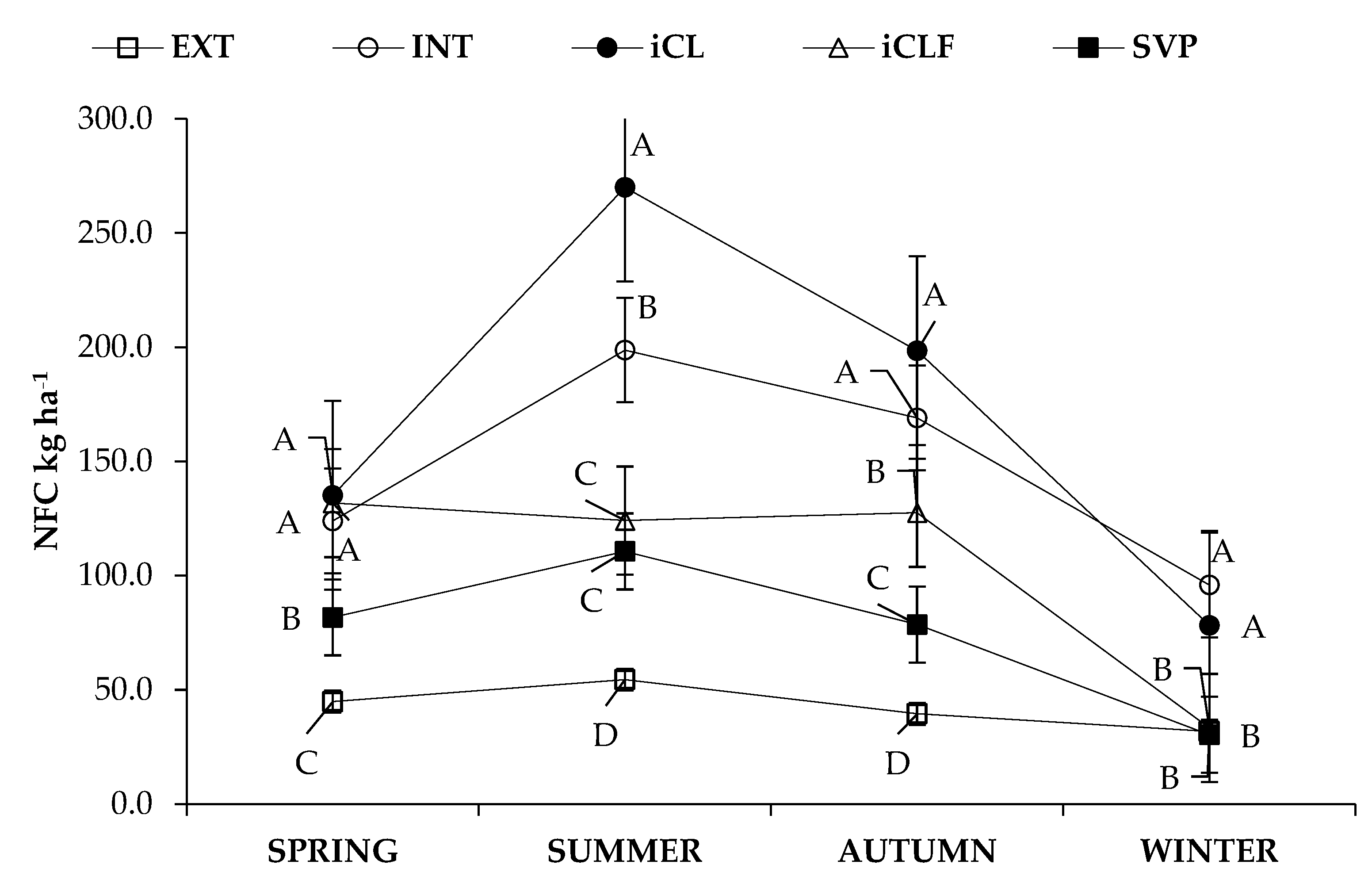
| System * | Season | Forage Mass | IVDDM | CP | NDF | NFC |
| kg ha−1 | ||||||
| Main Effects | ||||||
| EXT | 465.90 | 279.0 | 39.7 | 348.1 | 42.7 | |
| INT | 1372.07 | 756.0 | 120.6 | 980.1 | 147.0 | |
| iCL | 1393.83 | 755.7 | 117.6 | 978.1 | 170.5 | |
| iCLF | 924.17 | 536.7 | 108.5 | 619.1 | 104.2 | |
| iLF | 686.33 | 386.1 | 76.4 | 469.0 | 75.4 | |
| Spring | 961.09 | 560.9 | 100.5 | 660.3 | 103.5 | |
| Summer | 1291.49 | 764.7 | 130.6 | 890.4 | 151.7 | |
| Autumn | 1102.20 | 593.5 | 104.6 | 774.1 | 122.7 | |
| Winter | 519.06 | 251.7 | 34.5 | 390.7 | 54.0 | |
| Means | ||||||
| Mean | 968.5 | 542.7 | 92.6 | 678.9 | 108.0 | |
| SEM | 61.5 | 35.3 | 6.3 | 42.9 | 7.8 | |
| Statistical Probabilities | ||||||
| System | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| Season | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| System × Season | 0.0004 | <0.0001 | <0.0001 | 0.0007 | <0.0001 | |
| Variables | Systems * | |||||||
|---|---|---|---|---|---|---|---|---|
| n | EXT | INT | iCL | iCLF | iLF | SEM | p-Value | |
| ILW (kg) | 60 | 253 | 267 | 256 | 267 | 267 | 8.39 | 0.5940 |
| FLW (kg) | 60 | 429 b | 484 a | 466 a | 416 b | 414 b | 16.76 | <0.0001 |
| DMI (kg day−1) | 60 | 9.8 a | 8.7 ab | 7.5 b | 8.9 ab | 8.3 ab | 0.31 | <0.0001 |
| LWG (kg ha−1 year−1) | 60 | 290 c | 615 a | 487 ab | 385 bc | 497 ab | 53.98 | <0.0001 |
| CH4 (g day−1) | 60 | 199.7 | 226.1 | 209.8 | 180.9 | 196.7 | 7.3 | 0.1606 |
| CH4 (g kg LW−1) | 60 | 0.62 | 0.58 | 0.61 | 0.53 | 0.58 | 0.03 | 0.2047 |
| CH4 (kg kgDMI−1) | 60 | 0.028 a | 0.028 a | 0.029 a | 0.022 b | 0.025 ab | 0.001 | <0.0001 |
| gCH4 kgADG−1 LWG ha−1 year−1 | 60 | 1.6 a | 0.6 c | 0.8 bc | 1.1 ab | 0.7 bc | 0.09 | 0.0031 |
| kgCH4 kg Carcass eq.−1 | 60 | 0.496 a | 0.250 b | 0.297 b | 0.345 b | 0.286 b | 0.024 | 0.0047 |
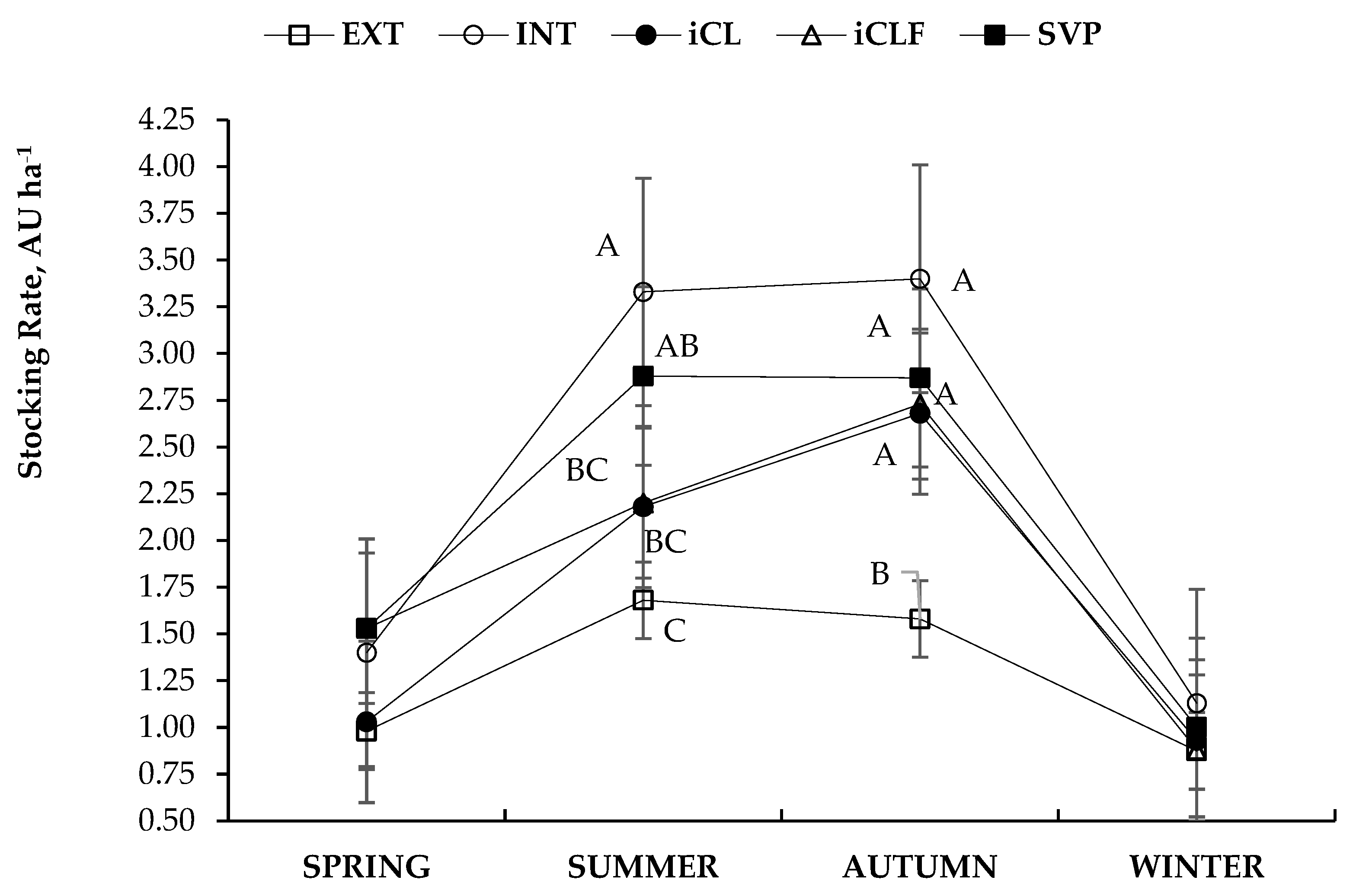
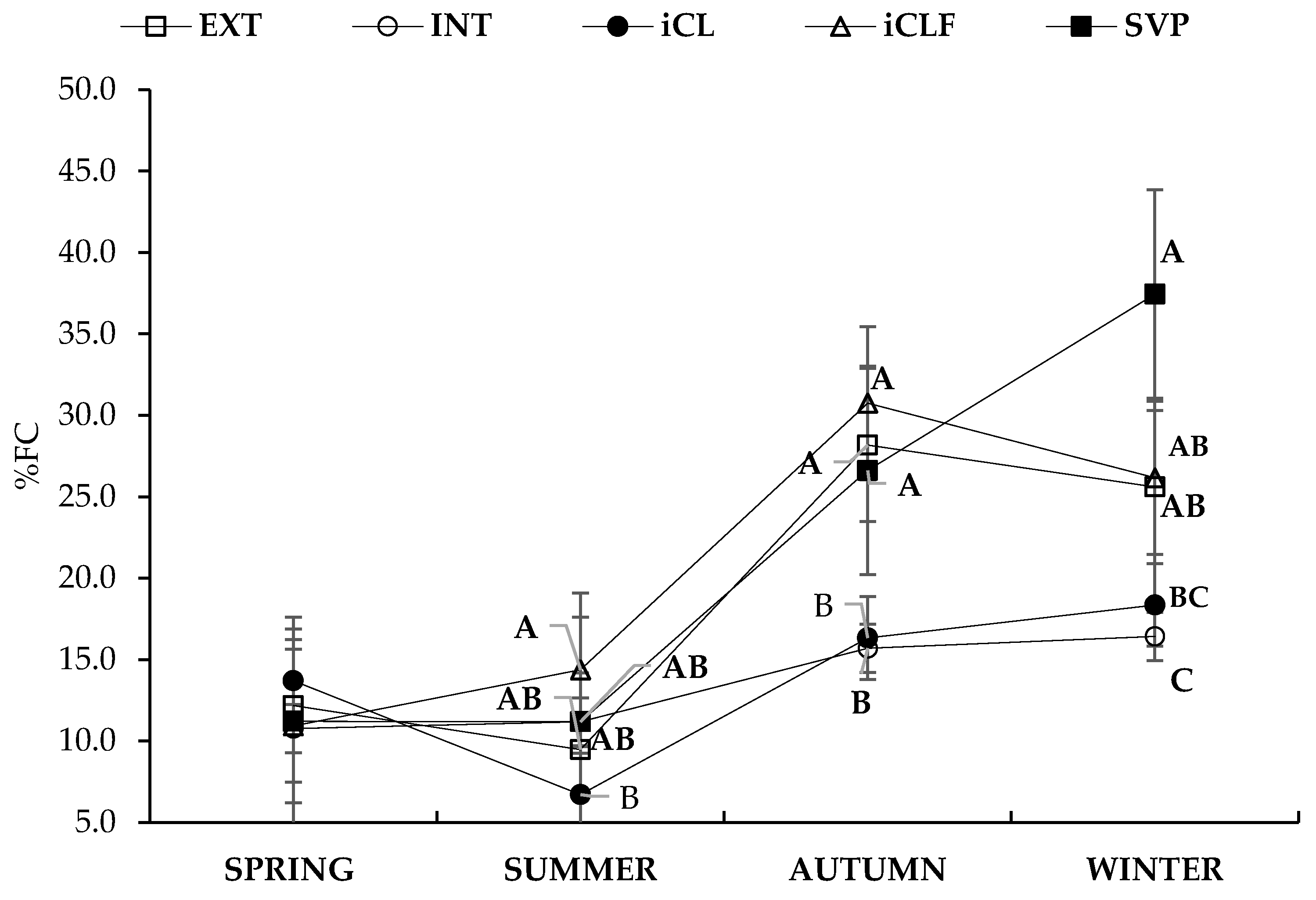
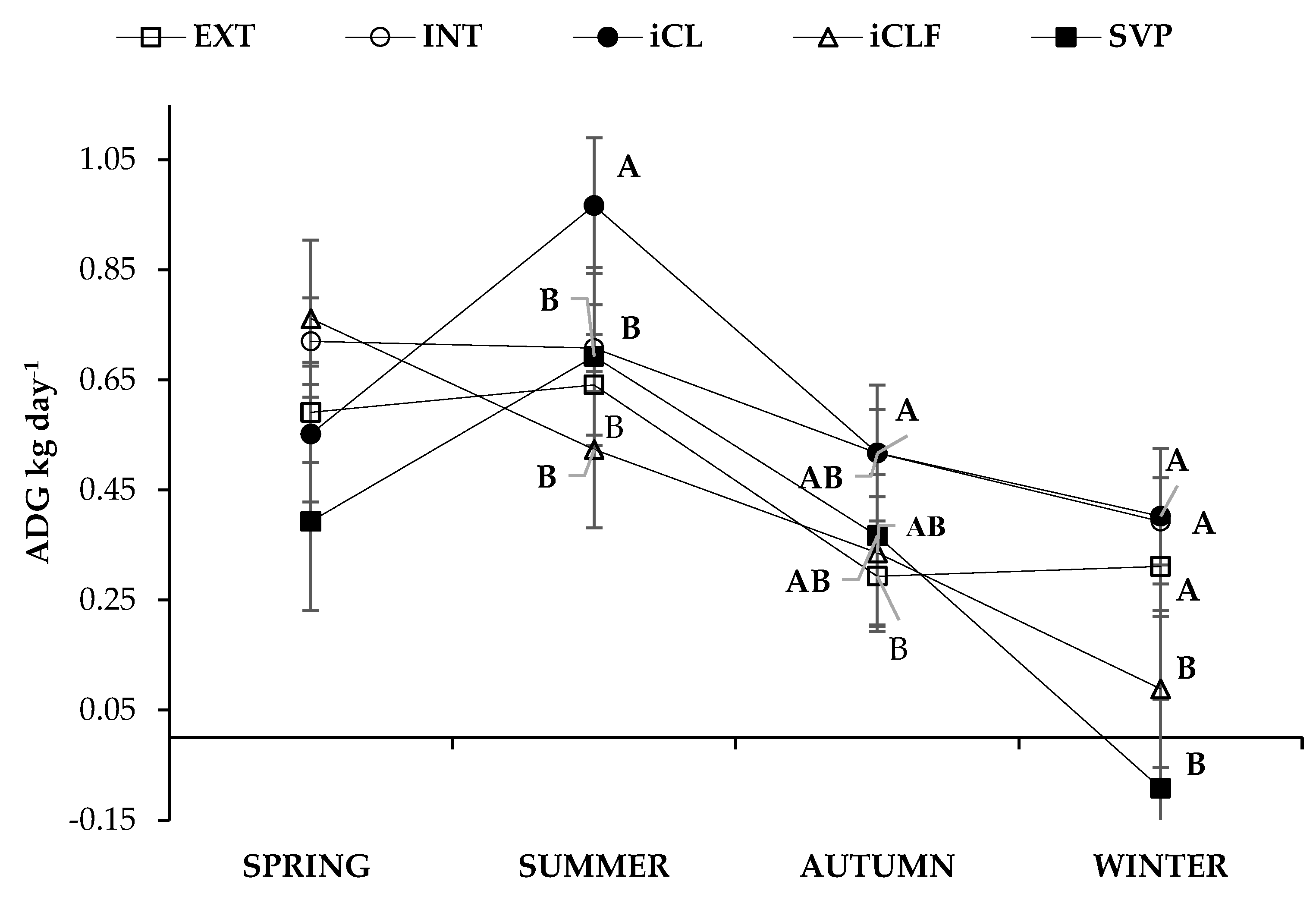
| Fixed Effects | Variables | ||||
|---|---|---|---|---|---|
| Systems * | Season | Stocking Rate (AU ha−1) | FC (kgDM kgLW−1) | ADG (kg day−1) | CH4 (kg kgADG−1) |
| EXT | 1.3 | 26.2 | 0.46 | 0.465 | |
| INT | 2.3 | 17.0 | 0.59 | 0.378 | |
| iCL | 1.7 | 14.9 | 0.58 | 0.343 | |
| iCLF | 1.8 | 20.6 | 0.43 | 0.441 | |
| iLF | 2.1 | 22.5 | 0.44 | 0.450 | |
| Spring | 1.3 | 12.7 | 0.67 | 0.342 | |
| Summer | 2.5 | 11.9 | 0.68 | 0.293 | |
| Autumn | 2.7 | 24.9 | 0.42 | 0.562 | |
| Winter | 0.96 | 31.5 | 0.23 | 0.466 | |
| Means | |||||
| Mean | 1.8 | 20.2 | 0.50 | 0.416 | |
| SEM | 0.10 | 3.3 | 0.02 | 0.053 | |
| p values | |||||
| System | <0.0001 | <0.0001 | <0.0001 | 0.0031 | |
| Season | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| System × Season | 0.0049 | 0.0201 | <0.0001 | 0.0007 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meo-Filho, P.; Berndt, A.; Pezzopane, J.R.M.; Pedroso, A.F.; Bernardi, A.C.C.; Rodrigues, P.H.M.; Bueno, I.C.S.; Corte, R.R.; Oliveira, P.P.A. Can Intensified Pasture Systems Reduce Enteric Methane Emissions from Beef Cattle in the Atlantic Forest Biome? Agronomy 2022, 12, 2738. https://doi.org/10.3390/agronomy12112738
Meo-Filho P, Berndt A, Pezzopane JRM, Pedroso AF, Bernardi ACC, Rodrigues PHM, Bueno ICS, Corte RR, Oliveira PPA. Can Intensified Pasture Systems Reduce Enteric Methane Emissions from Beef Cattle in the Atlantic Forest Biome? Agronomy. 2022; 12(11):2738. https://doi.org/10.3390/agronomy12112738
Chicago/Turabian StyleMeo-Filho, Paulo, Alexandre Berndt, José R. M. Pezzopane, André F. Pedroso, Alberto C. C. Bernardi, Paulo H. M. Rodrigues, Ives C. S. Bueno, Rosana R. Corte, and Patrícia P. A. Oliveira. 2022. "Can Intensified Pasture Systems Reduce Enteric Methane Emissions from Beef Cattle in the Atlantic Forest Biome?" Agronomy 12, no. 11: 2738. https://doi.org/10.3390/agronomy12112738
APA StyleMeo-Filho, P., Berndt, A., Pezzopane, J. R. M., Pedroso, A. F., Bernardi, A. C. C., Rodrigues, P. H. M., Bueno, I. C. S., Corte, R. R., & Oliveira, P. P. A. (2022). Can Intensified Pasture Systems Reduce Enteric Methane Emissions from Beef Cattle in the Atlantic Forest Biome? Agronomy, 12(11), 2738. https://doi.org/10.3390/agronomy12112738