
Cover Crops as Reservoirs for Young Vine Decline Pathogens


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Soil
2.2. Pathogen Inoculation and Plant Growing Conditions
2.3. Accessing Colonization by I. liriodendra
2.4. Data Analyses
3. Results
3.1. Abundance of Ilyonectria in Roots
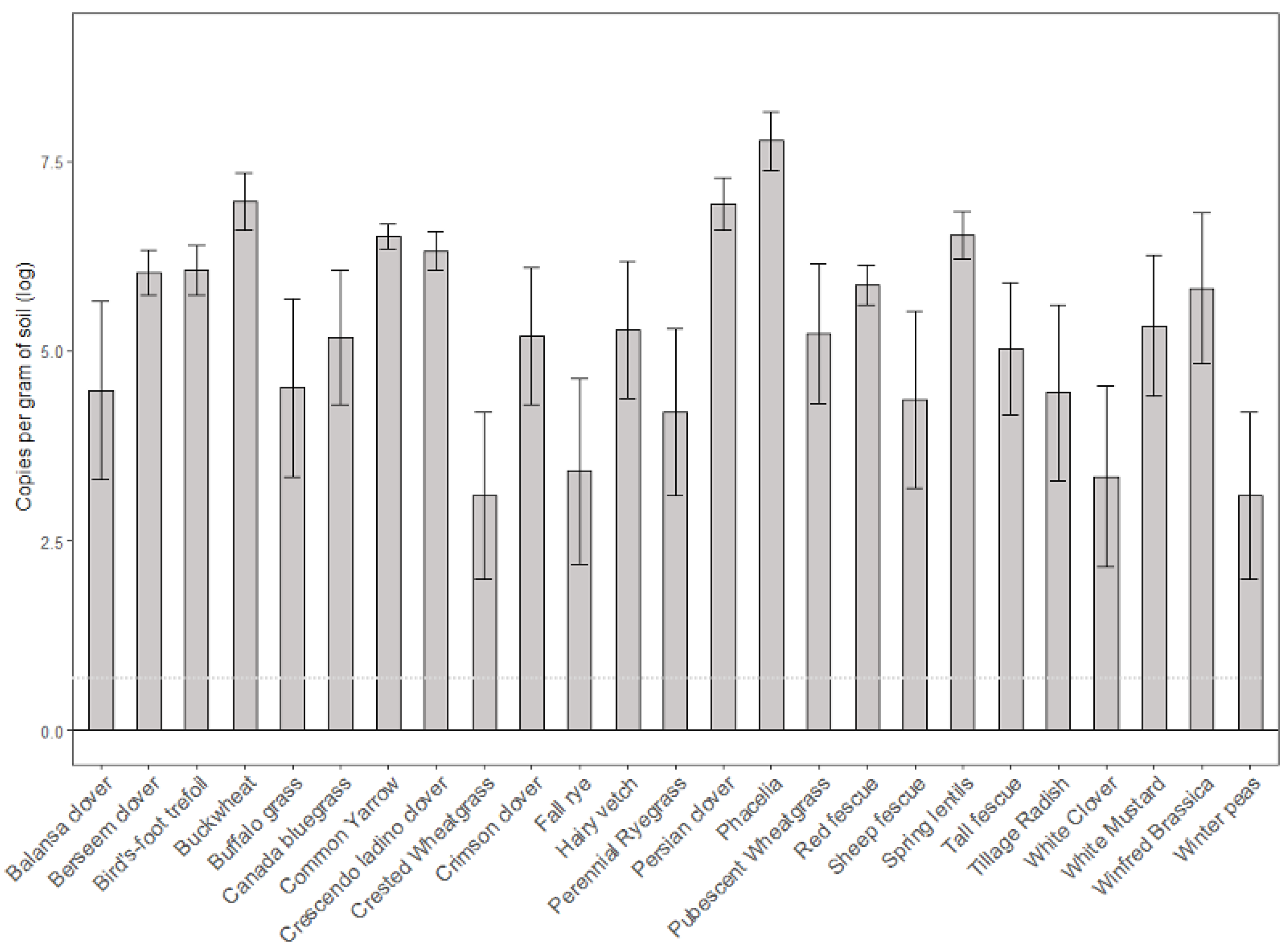
3.2. Abundance of Ilyonectria in Soil
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
| sample.ID | block | cover.crop | root.positives | soil.positives | root.per.gram | soil.per.gram |
| 1 | 1 | Balansa clover | 0 | 0 | 0 | 0 |
| 26 | 2 | Balansa clover | 0 | 2 | 0 | 370.37037 |
| 51 | 3 | Balansa clover | 0 | 1 | 0 | 172.413793 |
| 76 | 4 | Balansa clover | 0 | 6 | 0 | 1034.48276 |
| 101 | 5 | Balansa clover | 1 | 0 | 200 | 0 |
| 126 | 6 | Balansa clover | 0 | 6 | 0 | 1034.48276 |
| 151 | 7 | Balansa clover | 3 | 600 | ||
| 2 | 1 | Berseem clover | 0 | 4 | 0 | 714.285714 |
| 27 | 2 | Berseem clover | 0 | 1 | 0 | 172.413793 |
| 52 | 3 | Berseem clover | 2 | 1 | 400 | 178.571429 |
| 77 | 4 | Berseem clover | 1 | 6 | 192 | 1153.84615 |
| 102 | 5 | Berseem clover | 0 | 3 | 0 | 517.241379 |
| 127 | 6 | Berseem clover | 1 | 1 | 192 | 217.391304 |
| 152 | 7 | Berseem clover | 4 | 740.740741 | ||
| 3 | 1 | Bird's-foot trefoil | 1 | 1 | 200 | 178.571429 |
| 28 | 2 | Bird's-foot trefoil | 0 | 1 | 0 | 192.307692 |
| 53 | 3 | Bird's-foot trefoil | 0 | 11 | 0 | 1964.28571 |
| 78 | 4 | Bird's-foot trefoil | 0 | 3 | 0 | 576.923077 |
| 103 | 5 | Bird's-foot trefoil | 0 | 3 | 0 | 576.923077 |
| 128 | 6 | Bird's-foot trefoil | 0 | 1 | 0 | 192.307692 |
| 153 | 7 | Bird's-foot trefoil | 3 | 625 | ||
| 4 | 1 | Buckwheat | 29 | 8 | 6042 | 1428.57143 |
| 29 | 2 | Buckwheat | 82 | 14 | 15769 | 2800 |
| 54 | 3 | Buckwheat | 10 | 9 | 1923 | 1875 |
| 79 | 4 | Buckwheat | 13 | 4 | 2500 | 689.655172 |
| 104 | 5 | Buckwheat | 9 | 1 | 1667 | 178.571429 |
| 129 | 6 | Buckwheat | 5 | 3 | 1000 | 600 |
| 154 | 7 | Buckwheat | 14 | 2692.30769 | ||
| 5 | 1 | Buffalo grass | 1 | 6 | 1667 | 1111.11111 |
| 30 | 2 | Buffalo grass | 0 | 2 | 0 | 416.666667 |
| 55 | 3 | Buffalo grass | 2 | 0 | 3333 | 0 |
| 80 | 4 | Buffalo grass | 0 | 3 | 0 | 652.173913 |
| 105 | 5 | Buffalo grass | 0 | 2 | 0 | 434.782609 |
| 130 | 6 | Buffalo grass | 0 | 0 | 0 | 0 |
| 155 | 7 | Buffalo grass | 2 | 400 | ||
| 6 | 1 | Canada bluegrass | 0 | 2 | 0 | 416.666667 |
| 31 | 2 | Canada bluegrass | 0 | 1 | 0 | 227.272727 |
| 56 | 3 | Canada bluegrass | 0 | 0 | 0 | 0 |
| 81 | 4 | Canada bluegrass | 0 | 3 | 0 | 625 |
| 106 | 5 | Canada bluegrass | 2 | 4 | 400 | 714.285714 |
| 131 | 6 | Canada bluegrass | 1 | 1 | 185 | 185.185185 |
| 156 | 7 | Canada bluegrass | 4 | 714.285714 | ||
| 7 | 1 | Common yarrow | 0 | 4 | 0 | 714.285714 |
| 32 | 2 | Common yarrow | 2 | 6 | 400 | 1153.84615 |
| 57 | 3 | Common yarrow | 23 | 2 | 4600 | 370.37037 |
| 82 | 4 | Common yarrow | 5 | 2 | 1000 | 416.666667 |
| 107 | 5 | Common yarrow | 5 | 3 | 1000 | 625 |
| 132 | 6 | Common yarrow | 2 | 5 | 345 | 1086.95652 |
| 157 | 7 | Common yarrow | 4 | 689.655172 | ||
| 8 | 1 | Crescendo ladino | 0 | 1 | 0 | 178.571429 |
| 33 | 2 | Crescendo ladino | 0 | 2 | 0 | 384.615385 |
| 58 | 3 | Crescendo ladino | 0 | 3 | 0 | 576.923077 |
| 83 | 4 | Crescendo ladino | 0 | 3 | 0 | 500 |
| 108 | 5 | Crescendo ladino | 0 | 8 | 0 | 1379.31035 |
| 133 | 6 | Crescendo ladino | 0 | 5 | 0 | 1041.66667 |
| 158 | 7 | Crescendo ladino | 3 | 555.555556 | ||
| 9 | 1 | Crested wheatgrass | 1 | 1 | 208 | 200 |
| 34 | 2 | Crested wheatgrass | 1 | 1 | 714 | 172.413793 |
| 59 | 3 | Crested wheatgrass | 0 | 0 | 0 | 0 |
| 84 | 4 | Crested wheatgrass | 0 | 1 | 0 | 200 |
| 109 | 5 | Crested wheatgrass | 1 | 2 | 200 | 370.37037 |
| 134 | 6 | Crested wheatgrass | 1 | 0 | 200 | 0 |
| 159 | 7 | Crested wheatgrass | 0 | 0 | ||
| 10 | 1 | Crimson clover | 0 | 0 | 0 | 0 |
| 35 | 2 | Crimson clover | 0 | 3 | 0 | 600 |
| 60 | 3 | Crimson clover | 0 | 1 | 0 | 200 |
| 85 | 4 | Crimson clover | 2 | 7 | 400 | 1400 |
| 110 | 5 | Crimson clover | 0 | 3 | 0 | 517.241379 |
| 135 | 6 | Crimson clover | 1 | 2 | 172 | 384.615385 |
| 160 | 7 | Crimson clover | 1 | 185.185185 | ||
| 11 | 1 | Fall rye | 1 | 0 | 192 | 0 |
| 36 | 2 | Fall rye | 0 | 6 | 0 | 1071.42857 |
| 61 | 3 | Fall rye | 1 | 1 | 200 | 166.666667 |
| 86 | 4 | Fall rye | 0 | 0 | 0 | 0 |
| 111 | 5 | Fall rye | 1 | 2 | 208 | 370.37037 |
| 136 | 6 | Fall rye | 0 | 2 | 0 | 370.37037 |
| 161 | 7 | Fall rye | 0 | 0 | ||
| 12 | 1 | Hairy vetch | 2 | 4 | 357 | 689.655172 |
| 37 | 2 | Hairy vetch | 0 | 2 | 0 | 312.5 |
| 62 | 3 | Hairy vetch | 0 | 3 | 0 | 576.923077 |
| 87 | 4 | Hairy vetch | 2 | 1 | 400 | 217.391304 |
| 112 | 5 | Hairy vetch | 6 | 0 | 1034 | 0 |
| 137 | 6 | Hairy vetch | 1 | 2 | 200 | 416.666667 |
| 162 | 7 | Hairy vetch | 5 | 961.538462 | ||
| 13 | 1 | Perennial ryegrass | 1 | 1 | 185 | 192.307692 |
| 38 | 2 | Perennial ryegrass | 0 | 0 | 0 | 0 |
| 63 | 3 | Perennial ryegrass | 2 | 4 | 417 | 740.740741 |
| 88 | 4 | Perennial ryegrass | 0 | 0 | 0 | 0 |
| 113 | 5 | Perennial ryegrass | 0 | 2 | 0 | 434.782609 |
| 138 | 6 | Perennial ryegrass | 0 | 2 | 0 | 416.666667 |
| 163 | 7 | Perennial ryegrass | 1 | 217.391304 | ||
| 14 | 1 | Persian clover | 0 | 6 | 0 | 1200 |
| 39 | 2 | Persian clover | 0 | 2 | 0 | 384.615385 |
| 64 | 3 | Persian clover | 5 | 5 | 962 | 862.068966 |
| 89 | 4 | Persian clover | 1 | 10 | 192 | 1923.07692 |
| 114 | 5 | Persian clover | 0 | 33 | 0 | 5500 |
| 139 | 6 | Persian clover | 20 | 3 | 3846 | 576.923077 |
| 164 | 7 | Persian clover | 3 | 500 | ||
| 15 | 1 | Phacelia | 18 | 2 | 3750 | 370.37037 |
| 40 | 2 | Phacelia | 99 | 8 | 19038 | 1428.57143 |
| 65 | 3 | Phacelia | 2 | 14 | 385 | 2500 |
| 90 | 4 | Phacelia | 69 | 10 | 13269 | 2083.33333 |
| 115 | 5 | Phacelia | 15 | 26 | 3125 | 5200 |
| 140 | 6 | Phacelia | 124 | 18 | 23846 | 3103.44828 |
| 165 | 7 | Phacelia | 45 | 9000 | ||
| 16 | 1 | Pubescent wheatgrass | 0 | 0 | 0 | 0 |
| 41 | 2 | Pubescent wheatgrass | 0 | 1 | 0 | 185.185185 |
| 66 | 3 | Pubescent wheatgrass | 5 | 8800 | 833.333333 | |
| 91 | 4 | Pubescent wheatgrass | 0 | 1 | 0 | 200 |
| 116 | 5 | Pubescent wheatgrass | 0 | 2 | 0 | 333.333333 |
| 141 | 6 | Pubescent wheatgrass | 4 | 3 | 769 | 517.241379 |
| 166 | 7 | Pubescent wheatgrass | 9 | 1500 | ||
| 17 | 1 | Red fescue | 0 | 1 | 0 | 208.333333 |
| 42 | 2 | Red fescue | 3 | 3 | 600 | 555.555556 |
| 67 | 3 | Red fescue | 0 | 4 | 0 | 869.565217 |
| 92 | 4 | Red fescue | 0 | 1 | 0 | 156.25 |
| 117 | 5 | Red fescue | 0 | 2 | 0 | 370.37037 |
| 142 | 6 | Red fescue | 11 | 1 | 2115 | 178.571429 |
| 167 | 7 | Red fescue | 4 | 666.666667 | ||
| 18 | 1 | Sheep fescue | 0 | 3 | 0 | 600 |
| 43 | 2 | Sheep fescue | 1 | 1 | 185 | 185.185185 |
| 68 | 3 | Sheep fescue | 0 | 0 | 0 | 0 |
| 93 | 4 | Sheep fescue | 0 | 0 | 0 | 0 |
| 118 | 5 | Sheep fescue | 0 | 2 | 0 | 400 |
| 143 | 6 | Sheep fescue | 2 | 1 | 400 | 185.185185 |
| 168 | 7 | Sheep fescue | 11 | 2115.38462 | ||
| 19 | 1 | Spring lentils | 0 | 5 | 0 | 925.925926 |
| 44 | 2 | Spring lentils | 1 | 2 | 192 | 416.666667 |
| 69 | 3 | Spring lentils | 0 | 2 | 0 | 416.666667 |
| 94 | 4 | Spring lentils | 1 | 12 | 200 | 2222.22222 |
| 119 | 5 | Spring lentils | 0 | 3 | 0 | 652.173913 |
| 144 | 6 | Spring lentils | 5 | 7 | 1000 | 1521.73913 |
| 169 | 7 | Spring lentils | 1 | 192.307692 | ||
| 20 | 1 | Tall fescue | 0 | 4 | 0 | 869.565217 |
| 45 | 2 | Tall fescue | 0 | 1 | 0 | 200 |
| 70 | 3 | Tall fescue | 0 | 0 | 0 | 0 |
| 95 | 4 | Tall fescue | 0 | 1 | 0 | 208.333333 |
| 120 | 5 | Tall fescue | 0 | 2 | 0 | 416.666667 |
| 145 | 6 | Tall fescue | 1 | 3 | 192 | 750 |
| 170 | 7 | Tall fescue | 1 | 172.413793 | ||
| 21 | 1 | Tillage radish | 0 | 0 | 0 | 0 |
| 46 | 2 | Tillage radish | 1 | 0 | 192 | 0 |
| 71 | 3 | Tillage radish | 1 | 6 | 200 | 1034.48276 |
| 96 | 4 | Tillage radish | 0 | 3 | 0 | 500 |
| 121 | 5 | Tillage radish | 0 | 2 | 0 | 333.333333 |
| 146 | 6 | Tillage radish | 31 | 3 | 6200 | 535.714286 |
| 171 | 7 | Tillage radish | 2 | 357.142857 | ||
| 22 | 1 | White clover | 0 | 4 | 0 | 833.333333 |
| 47 | 2 | White clover | 3 | 0 | 600 | 0 |
| 72 | 3 | White clover | 0 | 2 | 0 | 370.37037 |
| 97 | 4 | White clover | 0 | 1 | 0 | 208.333333 |
| 122 | 5 | White clover | 1 | 1 | 179 | 227.272727 |
| 147 | 6 | White clover | 17 | 0 | 3269 | 0 |
| 172 | 7 | White clover | 0 | 0 | ||
| 23 | 1 | White mustard | 0 | 2 | 0 | 400 |
| 48 | 2 | White mustard | 4 | 1 | 741 | 217.391304 |
| 73 | 3 | White mustard | 7 | 10 | 1346 | 2000 |
| 98 | 4 | White mustard | 1 | 0 | 192 | 0 |
| 123 | 5 | White mustard | 5 | 2 | 1000 | 434.782609 |
| 148 | 6 | White mustard | 20 | 3 | 4000 | 535.714286 |
| 173 | 7 | White mustard | 2 | 400 | ||
| 24 | 1 | Winfred brassica | 0 | 2 | 0 | 384.615385 |
| 49 | 2 | Winfred brassica | 8 | 8 | 1667 | 1333.33333 |
| 74 | 3 | Winfred brassica | 0 | 3 | 0 | 652.173913 |
| 99 | 4 | Winfred brassica | 2 | 4 | 370 | 714.285714 |
| 124 | 5 | Winfred brassica | 0 | 11 | 0 | 2115.38462 |
| 149 | 6 | Winfred brassica | 40 | 6 | 7692 | 1000 |
| 174 | 7 | Winfred brassica | 0 | 0 | ||
| 25 | 1 | Winter peas | 2 | 1 | 400 | 200 |
| 50 | 2 | Winter peas | 1 | 0 | 192 | 0 |
| 75 | 3 | Winter peas | 0 | 0 | 0 | 0 |
| 100 | 4 | Winter peas | 0 | 0 | 0 | 0 |
| 125 | 5 | Winter peas | 1 | 1 | 185 | 227.272727 |
| 150 | 6 | Winter peas | 0 | 1 | 0 | 166.666667 |
| 175 | 7 | Winter peas | 2 | 322.580645 | ||
| 176 | 1 | Exp control background inoculant level | 55556 | 6.70804154 | ||
| 176 | 2 | Exp control background inoculant level | 55556 | 6.70804154 | ||
| 176 | 3 | Exp control background inoculant level | 55556 | 6.70804154 | ||
| 176 | 4 | Exp control background inoculant level | 55556 | 6.70804154 | ||
| 176 | 5 | Exp control background inoculant level | 55556 | 6.70804154 | ||
| 176 | 6 | Exp control background inoculant level | 55556 | 6.70804154 | ||
| 176 | 7 | Exp control background inoculant level | 55556 | 6.70804154 | ||
References
- Power, A.G.; Mitchell, C.E. Pathogen spillover in disease epidemics. Am. Nat. 2004, 164, S79–S89. [Google Scholar] [CrossRef] [PubMed]
- Beckstead, J.; Meyer, S.E.; Connolly, B.M.; Huck, M.B.; Street, L.E. Cheatgrass facilitates spillover of a seed bank pathogen onto native grass species. J. Ecol. 2010, 98, 168–177. [Google Scholar] [CrossRef]
- Akgül, D.S.; Ahioğlu, M. Fungal pathogens associated with young grapevine decline in the Southern Turkey vineyards. BIO Web Conf. 2019, 15, 01027. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Haag, P.; Bowen, P.; O’Gorman, D.T. Grapevine Trunk Diseases in British Columbia: Incidence and Characterization of the Fungal Pathogens Associated with Black Foot Disease of Grapevine. Plant Dis. 2014, 98, 456–468. [Google Scholar] [CrossRef]
- Manici, L.M.; Kelderer, M.; Franke-Whittle, I.H.; Rühmer, T.; Baab, G.; Nicoletti, F.; Caputo, F.; Topp, A.; Insam, H.; Naef, A. Relationship between root-endophytic microbial communities and replant disease in specialized apple growing areas in Europe. Appl. Soil Ecol. 2013, 72, 207–214. [Google Scholar] [CrossRef]
- Álvarez-Pérez, J.M.; González-García, S.; Cobos, R.; Olego, M.Á.; Ibañez, A.; Díez-Galán, A.; Garzón-Jimeno, E.; Coque, J.J. Use of endophytic and rhizosphere actinobacteria from grapevine plants to reduce nursery fungal graft infections that lead to young grapevine decline. Appl. Environ. Microbiol. 2017, 83, e01564-17. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, M.; Diniz, I.; Cabral, A.; Rego, C.; Oliveira, H. Unveiling inoculum sources of black foot pathogens in a commercial grapevine nursery. Phytopathol. Mediterr. 2013, 52, 298–312. [Google Scholar]
- Hrycan, J.; Hart, M.; Bowen, P.; Forge, T. Grapevine trunk disease fungi: Their roles as latent pathogens and stress factors that favour disease development and symptom expression. Phytopathol. Mediterr. 2020, 59, 395–424. [Google Scholar]
- Baumgartner, K.; Hillis, V.; Lubell, M.; Norton, M.; Kaplan, J. Managing grapevine trunk diseases in California’s southern san joaquin valley. Am. J. Enol. Vitic. 2019, 70, 267–276. [Google Scholar] [CrossRef]
- Claverie, M.; Notaro, M.; Fontaine, F.; Wery, J. Current knowledge on Grapevine Trunk Diseases with complex etiology: A systemic approach. Phytopathol. Mediterr. 2020, 59, 29–53. [Google Scholar] [CrossRef]
- Adaskaveg, J.; Gubler, D.; Michailides, T. Fungicides, Bactericides, and Biologicals for Deciduous Tree Fruit, Nut, Strawberry, and Vine Crops 2017. UC Davis: Department of Plant Pathology. Available online: https://www.vineyardteam.org/files/resources/2_Excerpt%20from%20fungicide%20efficacy%20timing.pdf (accessed on 17 May 2022).
- British Columbia Ministry of Agriculture. Pest Control Products Recommended for use on Grapes in British Columbia. Pest Manag. Available online: http://www.agf.gov.bc.ca/cropprot/grapeipm/grape_pesticides.pdf (accessed on 17 May 2022).
- Vallejo, A.; Millán, L.; Abrego, Z.; Sampedro, M.C.; Sánchez-Ortega, A.; Unceta, N.; Gómez-Caballero, A.; Goicolea, M.A.; Diez-Navajas, A.M.; Barrio, R.J. Fungicide distribution in vitiviniculture ecosystems according to different application strategies to reduce environmental impact. Sci. Total Environ. 2019, 687, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, N.; Perito, M.A. consumers’ willingness to consume sustainable and local wine in italy. Ital. J. Food Sci. 2020, 32, 222–233. [Google Scholar]
- Naher, L.; Yusuf, U.K.; Ismail, A.; Hossain, K. Trichoderma spp.: A biocontrol agent for sustainable management of plant diseases. Pak. J. Bot. 2014, 46, 1489–1493. [Google Scholar]
- Mounier, E.; Cortes, F.; Cadious, M.; Pajot, E. The benefits of Trichoderma atroviride I-1237 for the protection of grapevines against trunk diseases: From the nursery to the vineyard. Phytopathol. Mediterr. 2014, 53, 591–592. [Google Scholar]
- van Jaarsveld, W.J.; Halleen, F.; Bester, M.C.; Pierron, R.J.; Stempien, E.; Mostert, L. Investigation of Trichoderma species colonization of nursery grapevines for improved management of black foot disease. Pest Manag. Sci. 2020, 77, 397–405. [Google Scholar] [CrossRef]
- Clark, A. Managing Cover Crops Profitably, 3rd ed.; Sustainable Agriculture Research and Education: South Burlington, VT, USA, 2012; p. 3. [Google Scholar]
- Runno-Paurson, E.; Lääniste, P.; Eremeev, V.; Tähtjärv, T.; Kaurilind, E.; Tosens, T.; Niinemets, Ü.; Williams, I.H. Does winter oilseed rape as a winter cover crop influence potato late blight development in an organic crop rotation? Biol. Agric. Hortic. 2019, 36, 71–83. [Google Scholar] [CrossRef]
- Nivelle, E.; Verzeaux, J.; Habbib, H.; Kuzyakov, Y.; Decocq, G.; Roger, D.; Lacoux, J.; Duclercq, J.; Spicher, F.; Nava-Saucedo, J.E.; et al. Functional response of soil microbial communities to tillage, cover crops and nitrogen fertilization. Appl. Soil Ecol. 2016, 108, 147–155. [Google Scholar] [CrossRef]
- Eisenhauer, N. Aboveground-belowground interactions drive the relationship between plant diversity and ecosystem function. Res. Ideas Outcomes 2018, 4, e23688. [Google Scholar] [CrossRef]
- Dietrich, P.; Roscher, C.; Clark, A.T.; Eisenhauer, N.; Schmid, B.; Wagg, C. Diverse plant mixtures sustain a greater arbuscular mycorrhizal fungi spore viability than monocultures after 12 years. J. Plant Ecol. 2020, 13, 478–488. [Google Scholar] [CrossRef]
- Gattullo, C.E.; Mezzapesa, G.N.; Stellacci, A.M.; Ferrara, G.; Occhiogrosso, G.; Petrelli, G.; Castellini, M.; Spagnuolo, M. Cover crop for a sustainable viticulture: Effects on soil properties and table grape production. Agronomy 2020, 10, 1334. [Google Scholar] [CrossRef]
- Benitez, M.S.; Taheri, W.I.; Lehman, R.M. Selection of fungi by candidate cover crops. Appl. Soil Ecol. 2016, 103, 72–82. [Google Scholar] [CrossRef]
- Vukicevich, E.; Lowery, T.D.; Úrbez-Torres, J.R.; Bowen, P.; Hart, M. Groundcover management changes grapevine root fungal communities and plant-soil feedback. Plant Soil 2018, 424, 419–433. [Google Scholar] [CrossRef]
- Langenhoven, S.D.; Halleen, F.; Spies, C.F.J.; Stempien, E.; Mostert, L. Detection and quantification of black foot and crown and root rot pathogens in grapevine nursery soils in the Western Cape of South Africa. Phytopathol. Mediterr. 2018, 57, 519–537. [Google Scholar]
- Agustí-Brisach, C.; Gramaje, D.; León, M.; García-Jiménez, J.; Armengol, J. Evaluation of vineyard weeds as potential hosts of black-foot and petri disease pathogens. Plant Dis. 2011, 95, 803–810. [Google Scholar] [CrossRef]
- Van Niekerk, J.M.; Bester, W.; Halleen, F.; Crous, P.W.; Fourie, P.H. The distribution and symptomatology of grapevine trunk disease pathogens are influenced by climate. Phytopathol. Mediterr. 2011, 50, 98–111. [Google Scholar]
- Preece, C.; Peñuelas, J. Rhizodeposition under drought and consequences for soil communities and ecosystem resilience. Plant Soil 2016, 409, 1–17. [Google Scholar] [CrossRef]
- Hiscox, J.; Savoury, M.; Müller, C.T.; Lindahl, B.D.; Rogers, H.J.; Boddy, L. Priority effects during fungal community establishment in beech wood. ISME J. 2015, 9, 2246–2260. [Google Scholar] [CrossRef]
- Fukami, T.; Dickie, I.A.; Paula, W.J.; Paulus, B.C.; Park, D.; Roberts, A.; Buchanan, P.K.; Allen, R.B. Assembly history dictates ecosystem functioning: Evidence from wood decomposer communities. Ecol. Lett. 2010, 13, 675–684. [Google Scholar] [CrossRef]
- Holland, T.; Bowen, P.; Kokkoris, V.; Urbez-Torres, J.R.; Hart, M. Does inoculation with arbuscular mycorrhizal fungi reduce trunk disease in grapevine rootstocks? Horticulturae 2019, 5, 61. [Google Scholar] [CrossRef]
- Kokkoris, V.; Li, Y.; Hamel, C.; Hanson, K.; Hart, M. Site specificity in establishment of a commercial arbuscular mycorrhizal fungal inoculant. Sci. Total Environ. 2019, 660, 1135–1143. [Google Scholar] [CrossRef]
- Pathrose, B.; Jones, E.E.; Jaspers, M.V.; Ridgway, H.J. High genotypic and virulence diversity in Ilyonectria liriodendri isolates associated with black foot disease in New Zealand vineyards. Plant Pathol. 2014, 63, 613–624. [Google Scholar] [CrossRef]
- Martínez-Diz, M.P.; Andrés-Sodupe, M.; Berbegal, M.; Bujanda, R.; Díaz-Losada, E.; Gramaje, D. Droplet Digital PCR Technology for Detection of Ilyonectria liriodendri from Grapevine Environmental Samples. Plant Dis. 2020, 104, 1144–1150. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, D.P.; Nouri, M.T.; Trouillas, F.P. Taxonomy and multi-locus phylogeny of cylindrocarpon-like species associated with diseased roots of grapevine and other fruit and nut crops in California. Fungal Syst. Evol. 2018, 1, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Arafat, Y.; Tayyab, M.; Khan, M.U.; Chen, T.; Amjad, H.; Awais, S.; Lin, X.; Lin, W.; Lin, S. Long-term monoculture negatively regulates fungal community composition and abundance of tea orchards. Agronomy 2019, 9, 466. [Google Scholar] [CrossRef]
- Dann, E.K.; Cooke, A.W.; Forsberg, L.I.; Pegg, K.G.; Tan, Y.P.; Shivas, R.G. Pathogenicity studies in avocado with three nectriaceous fungi, Calonectria ilicicola, Gliocladiopsis sp. and Ilyonectria liriodendri. Plant Pathol. 2012, 61, 896–902. [Google Scholar] [CrossRef]
- Gilbert, C.; Dempcy, J.; Ganong, C.; Patterson, R.; Spicer, G.S. Phylogenetic relationships within Phacelia subgenus Phacelia (Hydrophyllaceae) inferred from nuclear rDNA ITS sequence data. Syst. Bot. 2005, 30, 627–634. [Google Scholar] [CrossRef]
- Tiryaki, I.; Keles, H. Reversal of the inhibitory effect of light and high temperature on germination of Phacelia tanacetifolia seeds by melatonin. J. Pineal Res. 2012, 52, 332–339. [Google Scholar] [CrossRef]
- Bacq-Labreuil, A.; Crawford, J.; Mooney, S.J.; Neal, A.L.; Ritz, K. Phacelia (Phacelia tanacetifolia Benth.) affects soil structure differently depending on soil texture. Plant Soil 2019, 441, 543–554. [Google Scholar] [CrossRef]
- Büchi, L.; Wendling, M.; Amossé, C.; Jeangros, B.; Charles, R. Cover crops to secure weed control strategies in a maize crop with reduced tillage. Field Crop. Res. 2020, 247. [Google Scholar] [CrossRef]
- Koike Salinas, S.T.; Smith, R.F.; Jackson, L.E.; Wyland, L.J.; Inman, J.I.; Chaney, W.E. Phacelia, Lana woollypod vetch, and Austrian winter pea: Three new cover crop hosts of Sclerotinia minor in California. Plant Dis. 1996, 80. [Google Scholar] [CrossRef]
- Kluth, C.; Buhre, C.; Varrelmann, M. Susceptibility of intercrops to infection with Rhizoctonia solani AG 2-2 IIIB and influence on subsequently cultivated sugar beet. Plant Pathol. 2010, 59, 1409–1412. [Google Scholar] [CrossRef]
- León, M.; Berbegal, M.; Abad-Campos, P.; Ramón-Albalat, A.; Caffi, T.; Rossi, V.; Hasanaliyeva, G.; Noceto, P.A.; Wipf, D.; Širca, S.; et al. Evaluation of sown cover crops and spontaneous weed flora as a potential reservoir of black-foot pathogens in organic viticulture. Biology 2021, 10, 498. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, O. Search for the wild ancestor of buckwheat III. The wild ancestor of cultivated common buckwheat, and of tatary buckwheat. Econ. Bot. 1998, 52, 123–133. [Google Scholar] [CrossRef]
- Morrall, R.A.A.; McKenzie, D.L. Diseases of specialty crops in Saskatchewan: 1. Notes on Buckwheatt and Sunflower 1972-73. Can. Plant Dis. Surv. 1975, 55, 69–72. Available online: https://phytopath.ca/wp-content/uploads/2014/10/cpds-archive/vol55/CPDS_Vol_55_No_2_(69-72)1975.pdf (accessed on 25 January 2022).
- Pathak, N.; Prajneshu, M.; Ahmad, S.; Kumar, L.; Bhaduri, A.; Dhandapani, A.; Sharma, O.P. Phytochemical Analysis and Antifungal Activity of Weed Extracts against Rhizoctonia Root Rot in Buckwheat ( Fagopyrum tataricum. Biopest. Int. 2021, 16, 125–131. [Google Scholar]
- Zini, P.B.; Poletto, T.; Fantinel, V.S.; Andrade, N.; Nunes, U.R.; Muniz, M.D.F.B.; Jacques, R.J.S. Buckwheat seed quality and pathogenicity of Fusarium spp. in plants. J. Seed Sci. 2022, 44. [Google Scholar] [CrossRef]
- Olivier, C.; Vaughn, S.F.; Mizubuti, E.S.G.; Loria, R. Variation in allyl isothiocyanate production within Brassica species and correlation with fungicidal activity. J. Chem. Ecol. 1999, 25, 2687–2701. [Google Scholar] [CrossRef]
- Wen, L.; Lee-Marzano, S.; Ortiz-Ribbing, L.M.; Gruver, J.; Hartman, G.L.; Eastburn, D.M. Suppression of soilborne diseases of soybean with cover crops. Plant Dis. 2017, 101, 1918–1928. [Google Scholar] [CrossRef]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| N. | Family | Binomial | Commom Name |
|---|---|---|---|
| 1 | Fabaceae | Trifolium michelianum | Balansa clover |
| 2 | Fabaceae | Trifolium alexandrinum | Berseem clover |
| 3 | Fabaceae | Lotus corniculatus | Bird’s-foot trefoil |
| 4 | Polygonaceae | Fagopyrum esculentum | Buckwheat |
| 5 | Poaceae | Bouteloua dactyloides | Buffalo grass |
| 6 | Poaceae | Poa compressa | Canada bluegrass |
| 7 | Asteraceae | Achillea millefolium | Common Yarrow |
| 8 | Fabaceae | Trifolium repens | Crescendo ladino clover |
| 9 | Poaceae | Agropyron cristatum | Crested Wheatgrass |
| 10 | Fabaceae | Trifolium incarnatum | Crimson clover |
| 11 | Poaceae | Secale cereale | Fall rye |
| 12 | Fabaceae | Vicia villosa | Hairy vetch |
| 13 | Poaceae | Lolium perenne | Perennial Ryegrass |
| 14 | Fabaceae | Trifolium resupinatum | Persian clover |
| 15 | Boraginaceae | Phacelia tanacetifolia | Phacelia |
| 16 | Poaceae | Thinopyrum intermedium | Pubescent Wheatgrass |
| 17 | Poaceae | Festuca rubra | Red fescue |
| 18 | Poaceae | Festuca ovina | Sheep fescue |
| 19 | Fabaceae | Lens culinaris | Spring lentils |
| 20 | Poaceae | Festuca arundinacea | Tall fescue |
| 21 | Brassicaceae | Raphanus sativus | Tillage Radish |
| 22 | Fabaceae | Trifolium repens | White Clover |
| 23 | Brassicaceae | Sinapis alba | White Mustard |
| 24 | Brassicaceae | Brassica rapa | Winfred Brassica |
| 25 | Fabaceae | Pisum sativum | Winter peas |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosa, D.; Sharifi, M.; Hart, M.M. Cover Crops as Reservoirs for Young Vine Decline Pathogens. Agronomy 2022, 12, 2422. https://doi.org/10.3390/agronomy12102422
Rosa D, Sharifi M, Hart MM. Cover Crops as Reservoirs for Young Vine Decline Pathogens. Agronomy. 2022; 12(10):2422. https://doi.org/10.3390/agronomy12102422
Chicago/Turabian StyleRosa, Daniel, Mehdi Sharifi, and Miranda M. Hart. 2022. "Cover Crops as Reservoirs for Young Vine Decline Pathogens" Agronomy 12, no. 10: 2422. https://doi.org/10.3390/agronomy12102422
APA StyleRosa, D., Sharifi, M., & Hart, M. M. (2022). Cover Crops as Reservoirs for Young Vine Decline Pathogens. Agronomy, 12(10), 2422. https://doi.org/10.3390/agronomy12102422