
Effect of 50 Years of No-Tillage, Stubble Retention, and Nitrogen Fertilization on Soil Respiration, Easily Extractable Glomalin, and Nitrogen Mineralization


 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Details and Soil Analysis
2.2. Measurement of Soil Microbial Respiration
2.3. Determination of EEGRSP Concentration
2.4. Determination of N Mineralization Rate
2.5. Statistical Analysis
3. Results
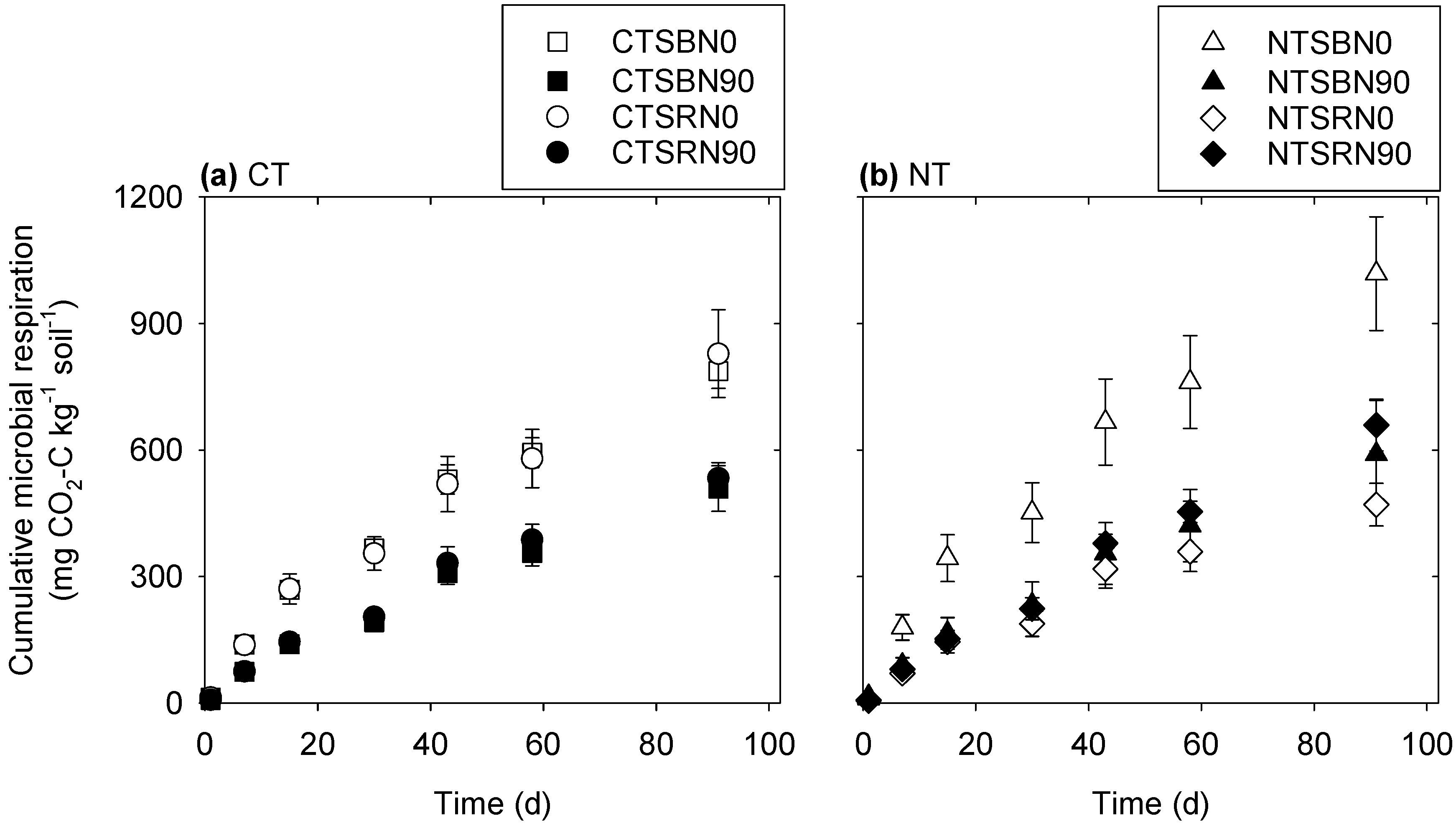
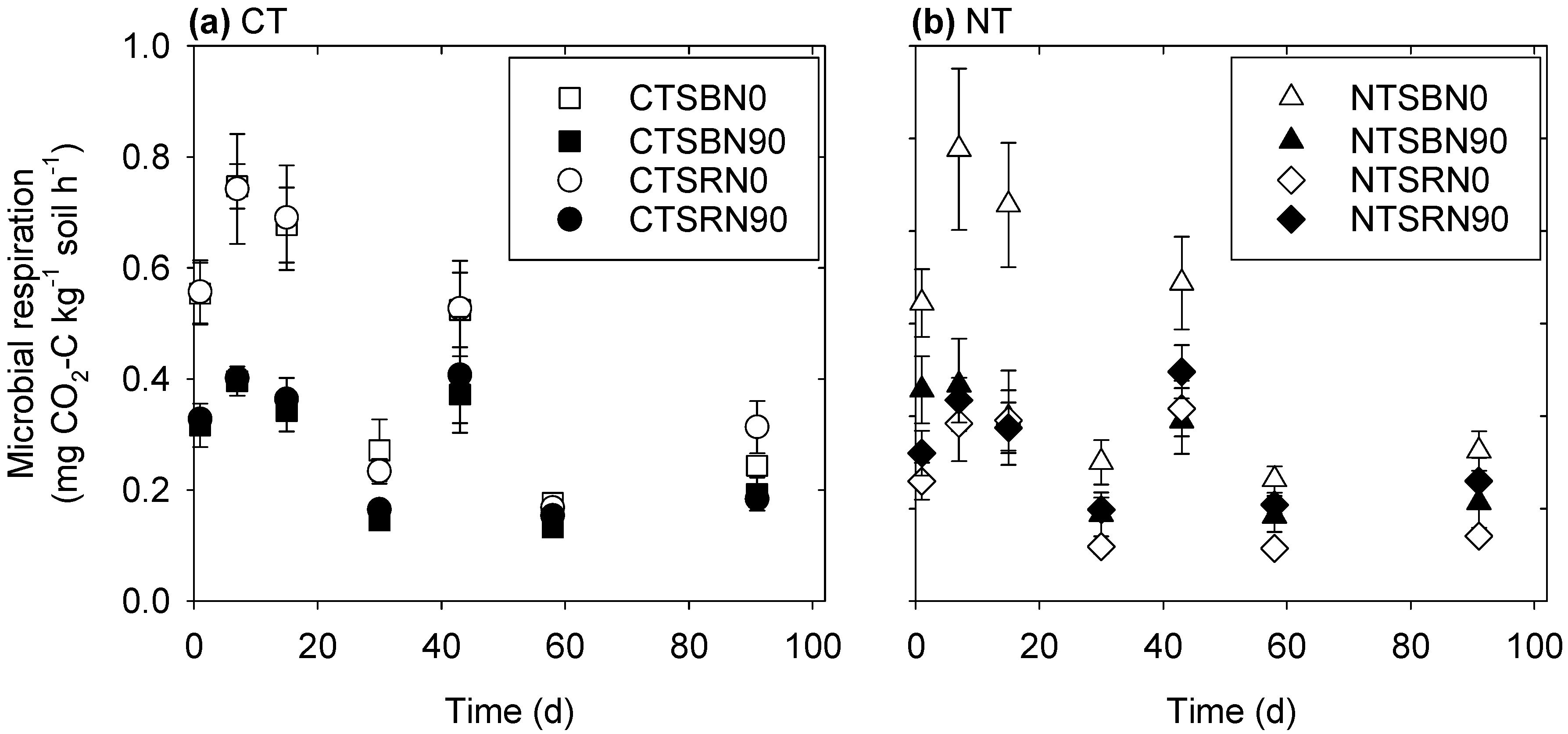
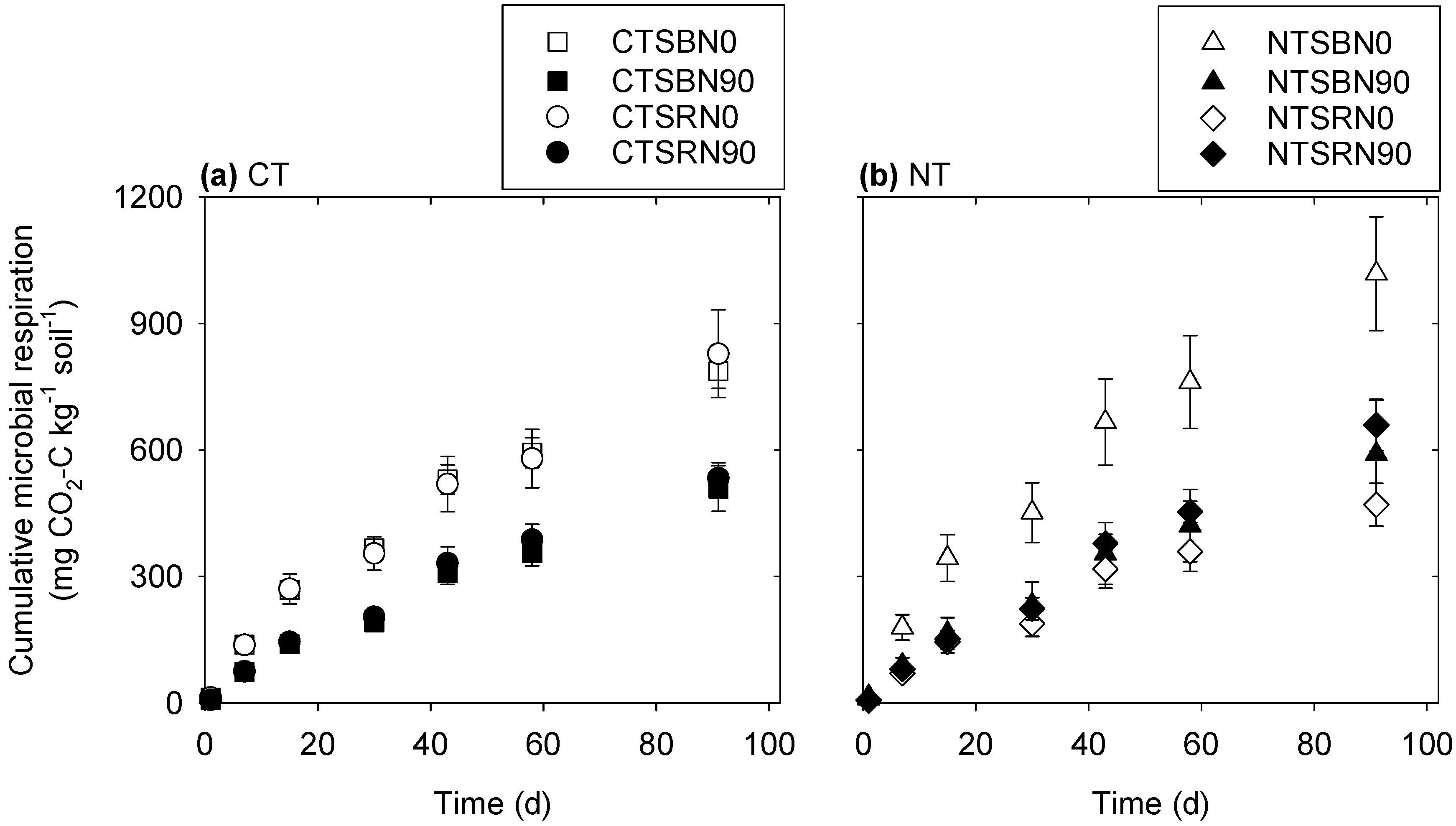
3.1. Microbial Respiration
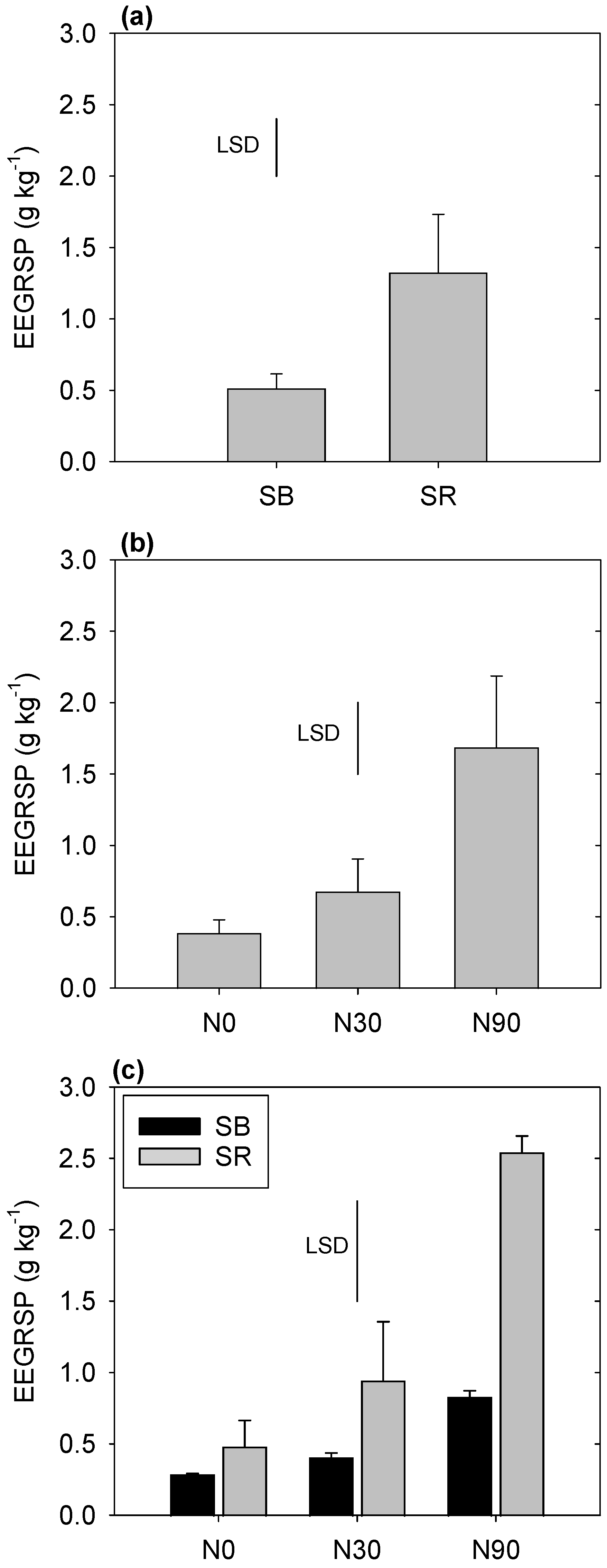
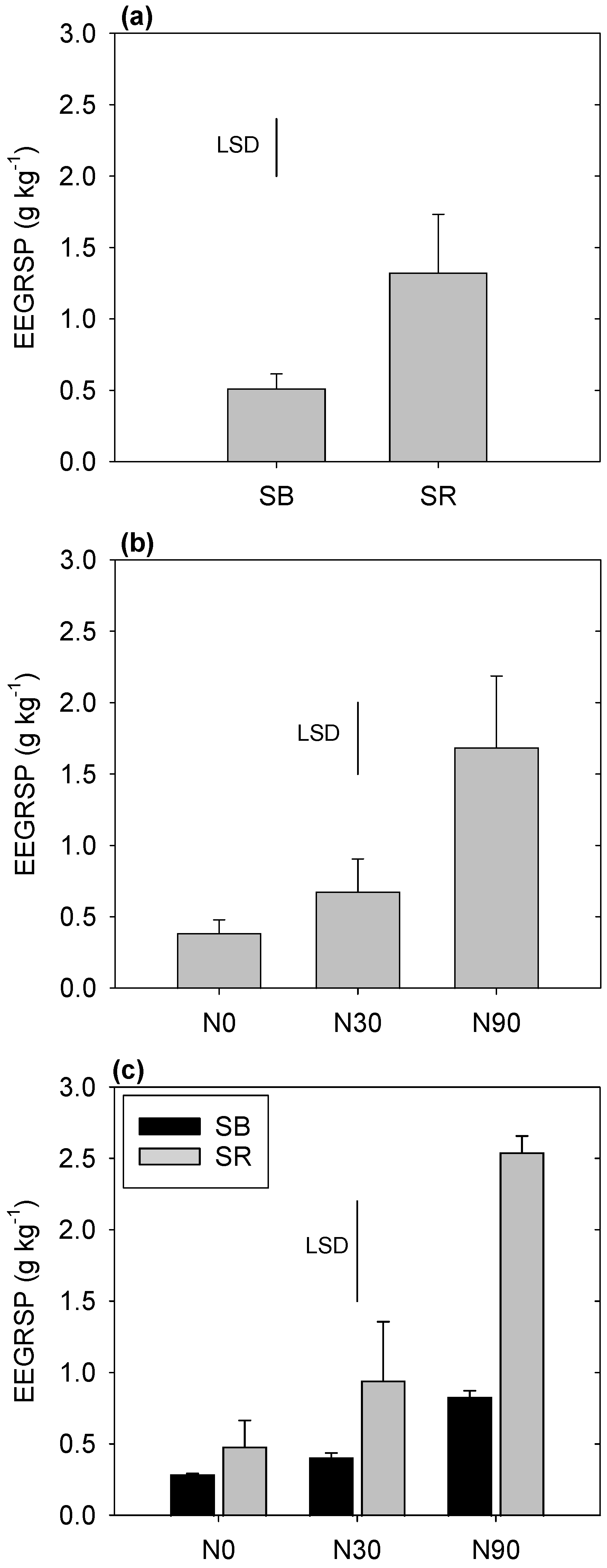
3.2. Easily Extractable Glomalin-Related Soil Protein (EEGRSP)
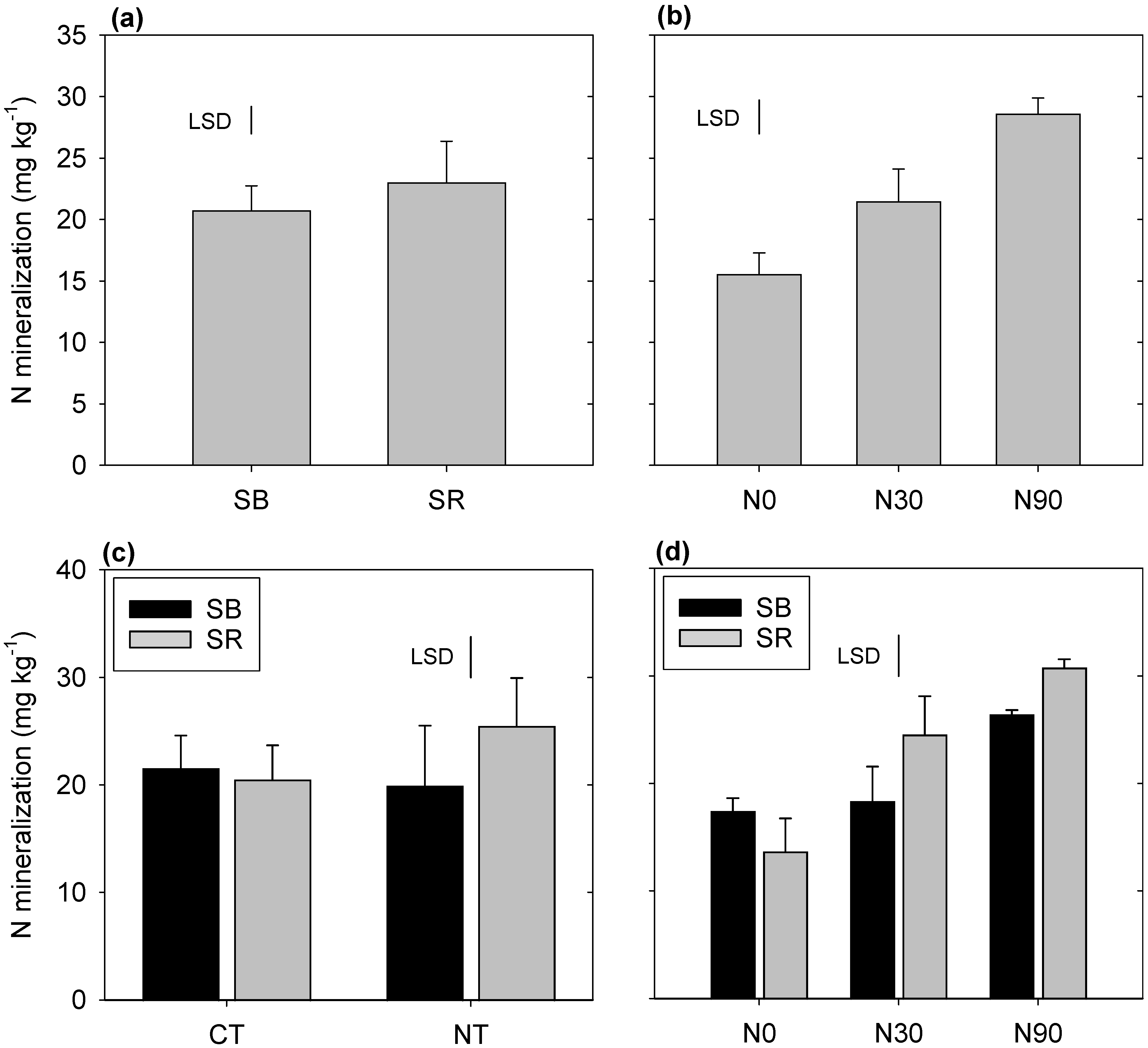
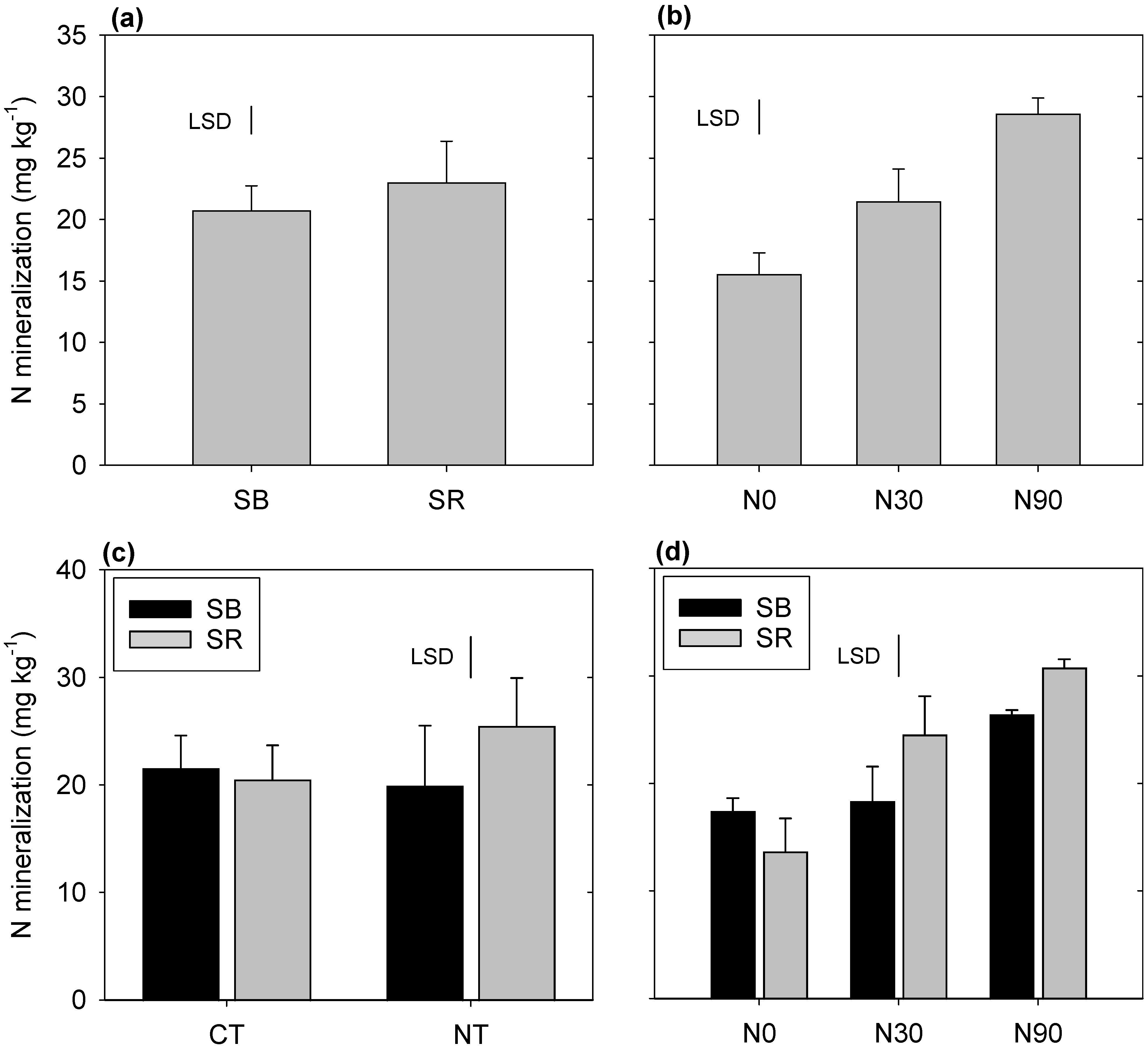
3.3. Net N Mineralization
4. Discussion
4.1. Microbial Respiration
4.2. EEGRSP
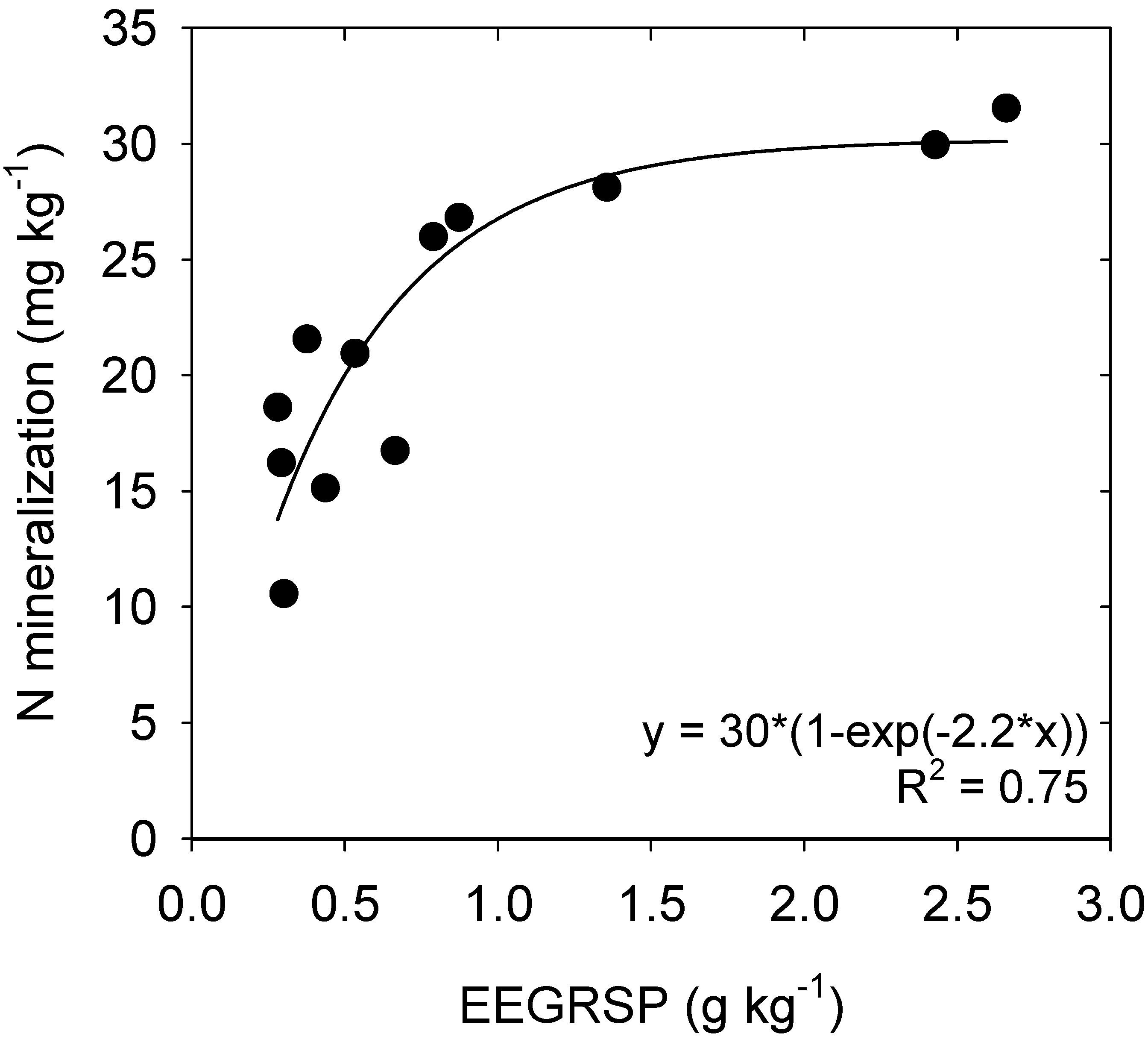
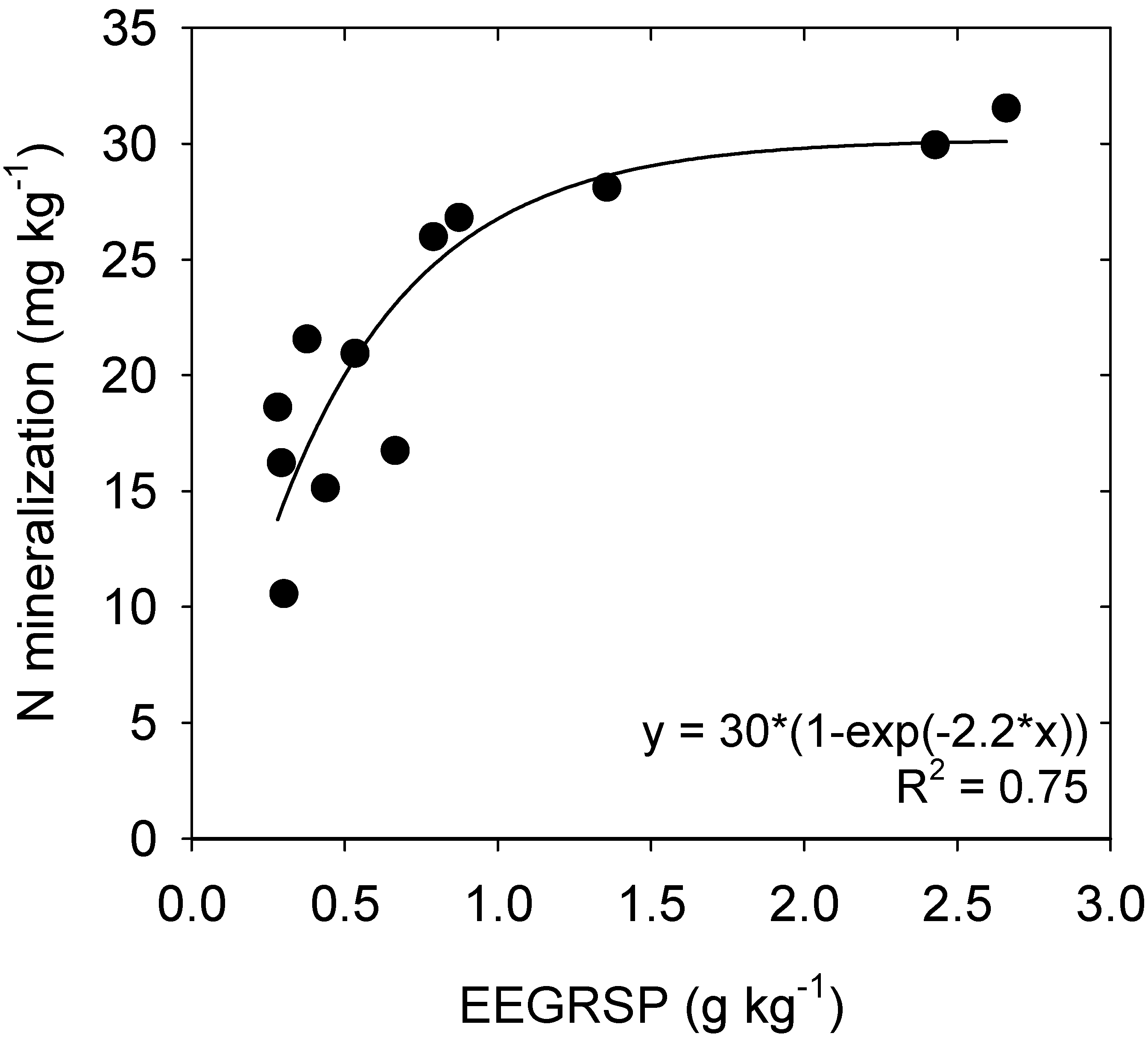
4.3. Nitrogen Mineralization
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dang, Y.P.; Dalal, R.C.; Menzies, N. No-Till Farming Systems for Sustainable Agriculture: Challenges and Opportunities; Springer: Cham, Switzerland, 2020. [Google Scholar]
- Pittelkow, C.M.; Liang, X.; Linquist, B.A.; van Groenigen, K.J.; Lee, J.; Lundy, M.E.; van Gestel, N.; Six, J.; Venterea, R.T.; van Kessel, C. Productivity limits and potentials of the principles of conservation agriculture. Nature 2015, 517, 365–368. [Google Scholar] [CrossRef]
- Lal, R. Sequestering carbon in soils of agro-ecosystems. Food Policy 2011, 36, S33–S39. [Google Scholar] [CrossRef]
- Mäder, P.; Edenhofer, S.; Boller, T.; Wiemken, A.; Niggli, U. Arbuscular mycorrhizae in a long-term field trial comparing low-input (organic, biological) and high-input (conventional) farming systems in a crop rotation. Biol. Fert. Soils 2000, 31, 150–156. [Google Scholar] [CrossRef]
- McGonigle, T.P.; Miller, M.H.; Young, D. Mycorrhizae, crop growth, and crop phosphorus nutrition in maize-soybean rotations given various tillage treatments. Plant Soil 1999, 210, 33–42. [Google Scholar] [CrossRef]
- Treseder, K.K.; Turner, K.M. Glomalin in Ecosystems. Soil Sci. Soc. Am. J. 2007, 71, 1257–1266. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.F.; Starr, J.L.; Paltineanu, I.C. Changes in Aggregate Stability and Concentration of Glomalin during Tillage Management Transition. Soil Sci. Soc. Am. J. 1999, 63, 1825–1828. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Wright, S.F.; Clark, D.A.; Ruess, R.W. Soil Stocks of Glomalin Produced by Arbuscular Mycorrhizal Fungi across a Tropical Rain Forest Landscape. J. Ecol. 2004, 92, 278–287. [Google Scholar] [CrossRef]
- Hurisso, T.T.; Culman, S.W.; Zhao, K. Repeatability and Spatiotemporal Variability of Emerging Soil Health Indicators Relative to Routine Soil Nutrient Tests. Soil Sci. Soc. Am. J. 2018, 82, 939–948. [Google Scholar] [CrossRef]
- Dai, J.; Hu, J.; Zhu, A.; Lin, X.; Nicholson, F. No-tillage with half-amount residue retention enhances microbial functional diversity, enzyme activity and glomalin-related soil protein content within soil aggregates. Soil Use Manag. Manag. 2017, 33, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Nie, J.; Zhou, J.-M.; Wang, H.-Y.; Chen, X.-Q.; Du, C.-W. Effect of Long-Term Rice Straw Return on Soil Glomalin, Carbon and Nitrogen. Pedosphere 2007, 17, 295–302. [Google Scholar] [CrossRef]
- Dai, J.; Hu, J.; Zhu, A.; Bai, J.; Wang, J.; Lin, X. No tillage enhances arbuscular mycorrhizal fungal population, glomalin-related soil protein content, and organic carbon accumulation in soil macroaggregates. J. Soils Sediments 2015, 15, 1055–1062. [Google Scholar] [CrossRef]
- Zhang, J.; Tang, X.; He, X.; Liu, J. Glomalin-related soil protein responses to elevated CO2 and nitrogen addition in a subtropical forest: Potential consequences for soil carbon accumulation. Soil Biol. Biochem. 2015, 83, 142–149. [Google Scholar] [CrossRef]
- Garcia, M.O.; Ovasapyan, T.; Greas, M.; Treseder, K.K. Mycorrhizal dynamics under elevated CO2 and nitrogen fertilization in a warm temperate forest. Plant Soil 2008, 303, 301–310. [Google Scholar] [CrossRef]
- Wuest, S.B.; Caesar-TonThat, T.C.; Wright, S.F.; Williams, J.D. Organic matter addition, N, and residue burning effects on infiltration, biological, and physical properties of an intensively tilled silt-loam soil. Soil Tillage Res. 2005, 84, 154–167. [Google Scholar] [CrossRef]
- Cordeiro, C.F.d.S.; Rodrigues, D.R.; Rocha, C.H.; Araujo, F.F.; Echer, F.R. Glomalin and microbial activity affected by cover crops and nitrogen management in sandy soil with cotton cultivation. App. Soil Ecol. 2021, 167, 104026. [Google Scholar] [CrossRef]
- Treseder, K.K. A Meta-Analysis of Mycorrhizal Responses to Nitrogen, Phosphorus, and Atmospheric CO2 in Field Studies. New Phytol. 2004, 164, 347–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlesinger, W.H.; Andrews, J.A.; Rustad, L.E.; Huntington, T.G.; Boone, R.D. Soil respiration and the global carbon cycle. Biogeochemistry 2000, 48, 7–20. [Google Scholar] [CrossRef]
- Bernhardt, E.S.; Barber, J.J.; Pippen, J.S.; Taneva, L.; Andrews, J.A.; Schlesinger, W.H. Long-Term Effects of Free Air CO2 Enrichment (FACE) on Soil Respiration. Biogeochemistry 2006, 77, 91–116. [Google Scholar] [CrossRef]
- Rochette, P.; Flanagan, L.B. Quantifying rhizosphere respiration in a corn crop under field conditions. Soil Sci. Soc. Am. J. 1997, 61, 466–474. [Google Scholar] [CrossRef]
- Thornton, P.E.; Doney, S.C.; Lindsay, K.; Moore, J.K.; Mahowald, N.; Randerson, J.T.; Fung, I.; Lamarque, J.F.; Feddema, J.J.; Lee, Y.H. Carbon-nitrogen interactions regulate climate-carbon cycle feedbacks: Results from an atmosphere-ocean general circulation model. Biogeosciences 2009, 6, 2099–2120. [Google Scholar] [CrossRef] [Green Version]
- Baillie, I.C. Soil Survey Staff 1999, Soil Taxonomy: A basic system of soil classification for making and interpreting soil surveys, 2nd edition. Agricultural Handbook 436, Natural Resources Conservation. Soil Use Manag. 2001, 17, 57–60. [Google Scholar] [CrossRef]
- Marley, J.M.; Littler, J.W. Winter cereal production on the Darling Downs dash an 11 year study of fallowing practices. Aust. J. Exp. Agric 1989, 29, 807. [Google Scholar] [CrossRef]
- Jones, A.R.; Gupta, V.V.S.R.; Buckley, S.; Brackin, R.; Schmidt, S.; Dalal, R.C. Drying and rewetting effects on organic matter mineralisation of contrasting soils after 36 years of storage. Geoderma 2019, 342, 12–19. [Google Scholar] [CrossRef]
- Wright, S.F.; Upadhyaya, A. Extraction of an abundant and unusual protein from soil and comparison with hyphal protein of arbuscular mycorrhizal fungi. Soil Sci. 1996, 161, 575–586. [Google Scholar] [CrossRef]
- Baird, R.; Bridgewater, L. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 2017. [Google Scholar]
- Mfilinge, P.; Atta, N.; Tsuchiya, M. Nutrient dynamics and leaf litter decomposition in a subtropical mangrove forest at Oura Bay, Okinawa, Japan. Trees 2002, 16, 172–180. [Google Scholar] [CrossRef]
- Finn, D.; Page, K.; Catton, K.; Strounina, E.; Kienzle, M.; Robertson, F.; Armstrong, R.; Dalal, R. Effect of added nitrogen on plant litter decomposition depends on initial soil carbon and nitrogen stoichiometry. Soil Biol. Biochem. 2015, 91, 160–168. [Google Scholar] [CrossRef]
- Reeves, S.H.; Somasundaram, J.; Wang, W.J.; Heenan, M.A.; Finn, D.; Dalal, R.C. Effect of soil aggregate size and long-term contrasting tillage, stubble and nitrogen management regimes on CO2 fluxes from a Vertisol. Geoderma 2019, 337, 1086–1096. [Google Scholar] [CrossRef]
- Six, J.; Paustian, K.; Elliott, E.T.; Combrink, C. Soil structure and organic matter. I, Distribution of aggregate-size classes and aggregate-associated carbon. Soil Sci. Soc. Am. J. 2000, 64, 681–689. [Google Scholar] [CrossRef]
- Jastrow, J.D. Soil aggregate formation and the accrual of particulate and mineral-associated organic matter. Soil Biol. Biochem. 1996, 28, 665–676. [Google Scholar] [CrossRef]
- Beare, M.H.; Hendrix, P.F.; Coleman, D.C. Water-Stable Aggregates and Organic Matter Fractions in Conventional- and No-Tillage Soils. Soil Sci. Soc. Am. J. 1994, 58, 777–786. [Google Scholar] [CrossRef]
- Jacinthe, P.A.; Lal, R. Labile carbon and methane uptake as affected by tillage intensity in a Mollisol. Soil Tillage Res. 2005, 80, 35–45. [Google Scholar] [CrossRef]
- Alluvione, F.; Halvorson, A.D.; Del Grosso, S.J. Nitrogen, Tillage, and Crop Rotation Effects on Carbon Dioxide and Methane Fluxes from Irrigated Cropping Systems. J. Environ. Qual. 2009, 38, 2023–2033. [Google Scholar] [CrossRef] [PubMed]
- Curtin, D.; Wang, H.; Selles, F.; McConkey, B.G.; Campbell, C.A. Tillage Effects on Carbon Fluxes in Continuous Wheat and Fallow–Wheat Rotations. Soil Sci. Soc. Am. J. 2000, 64, 2080–2086. [Google Scholar] [CrossRef]
- Thompson, J.P. Soil biotic and biochemical factors in a long-term tillage and stubble management experiment on a vertisol. 2. Nitrogen deficiency with zero tillage and stubble retention. Soil Tillage Res. 1992, 22, 339–361. [Google Scholar] [CrossRef]
- Jha, P.; Hati, K.M.; Dalal, R.C.; Dang, Y.P.; Kopittke, P.M.; Menzies, N.W. Soil carbon and nitrogen dynamics in a Vertisol following 50 years of no-tillage, crop stubble retention and nitrogen fertilization. Geoderma 2020, 358, 113996. [Google Scholar] [CrossRef]
- Wüthrich, C.; Schaub, D.; Weber, M.; Marxer, P.; Conedera, M. Soil respiration and soil microbial biomass after fire in a sweet chestnut forest in southern Switzerland. Catena 2002, 48, 201–215. [Google Scholar] [CrossRef]
- Almendros, G.; Leal, J.A. An evaluation of some oxidative degradation methods of humic substances applied to carbohydrate-derived humic-like polymers. J. Soil Sci. 1990, 41, 51–59. [Google Scholar] [CrossRef]
- Rashid, G.H. Effects of fire on soil carbon and nitrogen in a Mediterranean oak forest of Algeria. Plant Soil 1987, 103, 89–93. [Google Scholar] [CrossRef]
- Fernández, I.; Cabaneiro, A.; Carballas, T. Organic matter changes immediately after a wildfire in an atlantic forest soil and comparison with laboratory soil heating. Soil Biol. Biochem. 1997, 29, 1–11. [Google Scholar] [CrossRef]
- Lupwayi, N.Z.; Rice, W.A.; Clayton, G.W. Soil microbial diversity and community structure under wheat as influenced by tillage and crop rotation. Soil Biol. Biochem. 1998, 30, 1733–1741. [Google Scholar] [CrossRef]
- Ramirez, K.S.; Craine, J.M.; Fierer, N. Nitrogen fertilization inhibits soil microbial respiration regardless of the form of nitrogen applied. Soil Biol. Biochem. 2010, 42, 2336–2338. [Google Scholar] [CrossRef]
- Fierer, N.; Lauber, C.L.; Ramirez, K.S.; Zaneveld, J.; Bradford, M.A.; Knight, R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef] [Green Version]
- Swanston, C.; Homann, P.S.; Caldwell, B.A.; Myrold, D.D.; Ganio, L.; Sollins, P. Long-Term Effects Of Elevated Nitrogen On Forest Soil Organic Matter Stability. Biogeochemistry 2004, 70, 227–250. [Google Scholar] [CrossRef]
- Li, J.; Xie, R.Z.; Wang, K.R.; Ming, B.; Guo, Y.Q.; Zhang, G.Q.; Li, S.K. Variations in Maize Dry Matter, Harvest Index, and Grain Yield with Plant Density. Agron. J. 2015, 107, 829–834. [Google Scholar] [CrossRef]
- Manzoni, S.; Porporato, A. Soil carbon and nitrogen mineralization: Theory and models across scales. Soil Biol. Biochem. 2009, 41, 1355–1379. [Google Scholar] [CrossRef]
- Maaroufi, N.I.; Nordin, A.; Hasselquist, N.J.; Bach, L.H.; Palmqvist, K.; Gundale, M.J. Anthropogenic nitrogen deposition enhances carbon sequestration in boreal soils. Glob. Chang. Biol. 2015, 21, 3169–3180. [Google Scholar] [CrossRef] [PubMed]
- Frey, S.D.; Ollinger, S.; Nadelhoffer, K.; Bowden, R.; Brzostek, E.; Burton, A.; Caldwell, B.A.; Crow, S.; Goodale, C.L.; Grandy, A.S.; et al. Chronic nitrogen additions suppress decomposition and sequester soil carbon in temperate forests. Biogeochemistry 2014, 121, 305–316. [Google Scholar] [CrossRef]
- Bedini, S.; Pellegrino, E.; Avio, L.; Pellegrini, S.; Bazzoffi, P.; Argese, E.; Giovannetti, M. Changes in soil aggregation and glomalin-related soil protein content as affected by the arbuscular mycorrhizal fungal species Glomus mosseae and Glomus intraradices. Soil Biol. Biochem. 2009, 41, 1491–1496. [Google Scholar] [CrossRef]
- Silgram, M.; Chambers, B.J. Effects of long-term straw management and fertilizer nitrogen additions on soil nitrogen supply and crop yields at two sites in eastern England. J. Agric. Sci 2002, 139, 115–127. [Google Scholar] [CrossRef]
- Govaerts, B.; Sayre, K.D.; Ceballos-Ramirez, J.M.; Luna-Guido, M.L.; Limon-Ortega, A.; Deckers, J.; Dendooven, L. Conventionally tilled and permanent raised beds with different crop residue management: Effects on soil C and N dynamics. Plant Soil 2006, 280, 143–155. [Google Scholar] [CrossRef]
- Raiesi, F. Carbon and N mineralization as affected by soil cultivation and crop residue in a calcareous wetland ecosystem in Central Iran. Agric. Ecosyst. Environ. 2006, 112, 13–20. [Google Scholar] [CrossRef]
- Sarker, J.R.; Singh, B.P.; Dougherty, W.J.; Fang, Y.; Badgery, W.; Hoyle, F.C.; Dalal, R.C.; Cowie, A.L. Impact of agricultural management practices on the nutrient supply potential of soil organic matter under long-term farming systems. Soil Tillage Res. 2018, 175, 71–81. [Google Scholar] [CrossRef]
- Nannipieri, P.; Eldor, P. The chemical and functional characterization of soil N and its biotic components. Soil Biol. Biochem. 2009, 41, 2357–2369. [Google Scholar] [CrossRef]
- Fine, A.K.; Es, H.M.; Schindelbeck, R.R. Statistics, Scoring Functions, and Regional Analysis of a Comprehensive Soil Health Database. Soil Sci. Soc. Am. J. 2017, 81, 589–601. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Time (d) | |||||||
|---|---|---|---|---|---|---|---|---|
| df | 1 | 7 | 15 | 30 | 43 | 58 | 91 | |
| Tillage (T) | 1 | 0.617 | 0.923 | 0.810 | 0.897 | 0.475 | 0.017 | 0.968 |
| Stubble (S) | 1 | 0.002 | 0.019 | 0.059 | 0.059 | 0.565 | 0.048 | 0.472 |
| Nitrogen (N) | 1 | 0.001 | 0.000 | 0.000 | 0.026 | 0.034 | 0.433 | 0.113 |
| T × S | 1 | 0.002 | 0.019 | 0.031 | 0.123 | 0.362 | 0.016 | 0.069 |
| T × N | 1 | 0.033 | 0.357 | 0.422 | 0.101 | 0.809 | 0.180 | 0.085 |
| S × N | 1 | 0.100 | 0.031 | 0.059 | 0.015 | 0.071 | 0.001 | 0.160 |
| T × S × N | 1 | 0.122 | 0.036 | 0.068 | 0.170 | 0.126 | 0.013 | 0.007 |
| Source | Time (d) | |||||||
|---|---|---|---|---|---|---|---|---|
| df | 1 | 7 | 15 | 30 | 43 | 58 | 91 | |
| Tillage (T) | 1 | 0.622 | 0.894 | 0.849 | 0.848 | 0.863 | 0.660 | 0.738 |
| Stubble (S) | 1 | 0.003 | 0.014 | 0.027 | 0.020 | 0.065 | 0.055 | 0.098 |
| Nitrogen (N) | 1 | 0.001 | 0.000 | 0.000 | 0.000 | 0.000 | 0.001 | 0.002 |
| T × S | 1 | 0.002 | 0.014 | 0.018 | 0.019 | 0.048 | 0.037 | 0.033 |
| T × N | 1 | 0.034 | 0.296 | 0.346 | 0.206 | 0.337 | 0.301 | 0.177 |
| S × N | 1 | 0.101 | 0.030 | 0.039 | 0.017 | 0.019 | 0.011 | 0.020 |
| T × S × N | 1 | 0.123 | 0.036 | 0.046 | 0.045 | 0.047 | 0.035 | 0.014 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jha, P.; Hati, K.M.; Dalal, R.C.; Dang, Y.P.; Kopittke, P.M.; McKenna, B.A.; Menzies, N.W. Effect of 50 Years of No-Tillage, Stubble Retention, and Nitrogen Fertilization on Soil Respiration, Easily Extractable Glomalin, and Nitrogen Mineralization. Agronomy 2022, 12, 151. https://doi.org/10.3390/agronomy12010151
Jha P, Hati KM, Dalal RC, Dang YP, Kopittke PM, McKenna BA, Menzies NW. Effect of 50 Years of No-Tillage, Stubble Retention, and Nitrogen Fertilization on Soil Respiration, Easily Extractable Glomalin, and Nitrogen Mineralization. Agronomy. 2022; 12(1):151. https://doi.org/10.3390/agronomy12010151
Chicago/Turabian StyleJha, Pramod, Kuntal M. Hati, Ram C. Dalal, Yash P. Dang, Peter M. Kopittke, Brigid A. McKenna, and Neal W. Menzies. 2022. "Effect of 50 Years of No-Tillage, Stubble Retention, and Nitrogen Fertilization on Soil Respiration, Easily Extractable Glomalin, and Nitrogen Mineralization" Agronomy 12, no. 1: 151. https://doi.org/10.3390/agronomy12010151
APA StyleJha, P., Hati, K. M., Dalal, R. C., Dang, Y. P., Kopittke, P. M., McKenna, B. A., & Menzies, N. W. (2022). Effect of 50 Years of No-Tillage, Stubble Retention, and Nitrogen Fertilization on Soil Respiration, Easily Extractable Glomalin, and Nitrogen Mineralization. Agronomy, 12(1), 151. https://doi.org/10.3390/agronomy12010151