
Anatomical and Biochemical Traits Associated with Field Resistance of Onion Cultivars to Onion Thrips and the Effect of Mechanical Injury on the Level of Biochemical Compounds in Onion Leaves

 ,
,  , , ,
, , ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Setup
2.2. Field Resistance Experiment
2.3. Biochemical Analyses of Leaves
2.3.1. Total Soluble Sugar Content
2.3.2. Reducing Sugar Content
2.3.3. Sucrose Content
2.3.4. Total Phenolic Content
2.3.5. Chlorophyll and Total Carotenoid Contents
2.4. Anatomical Studies of Leaves
2.5. Studies on the Effect of Mechanical Injury on the Level of Biochemical Compounds in Onion Leaves
2.6. Statistical Analysis
3. Results
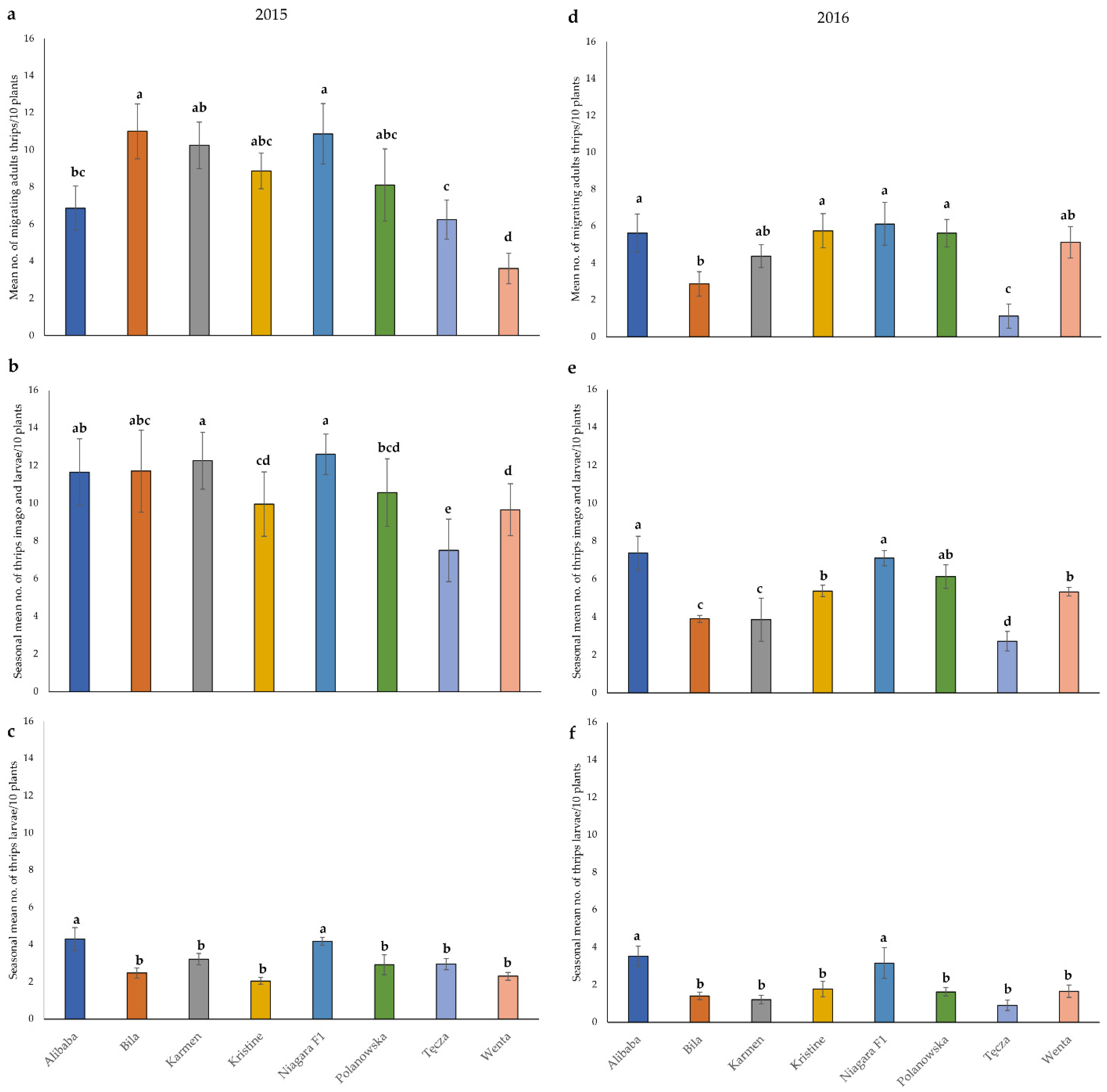
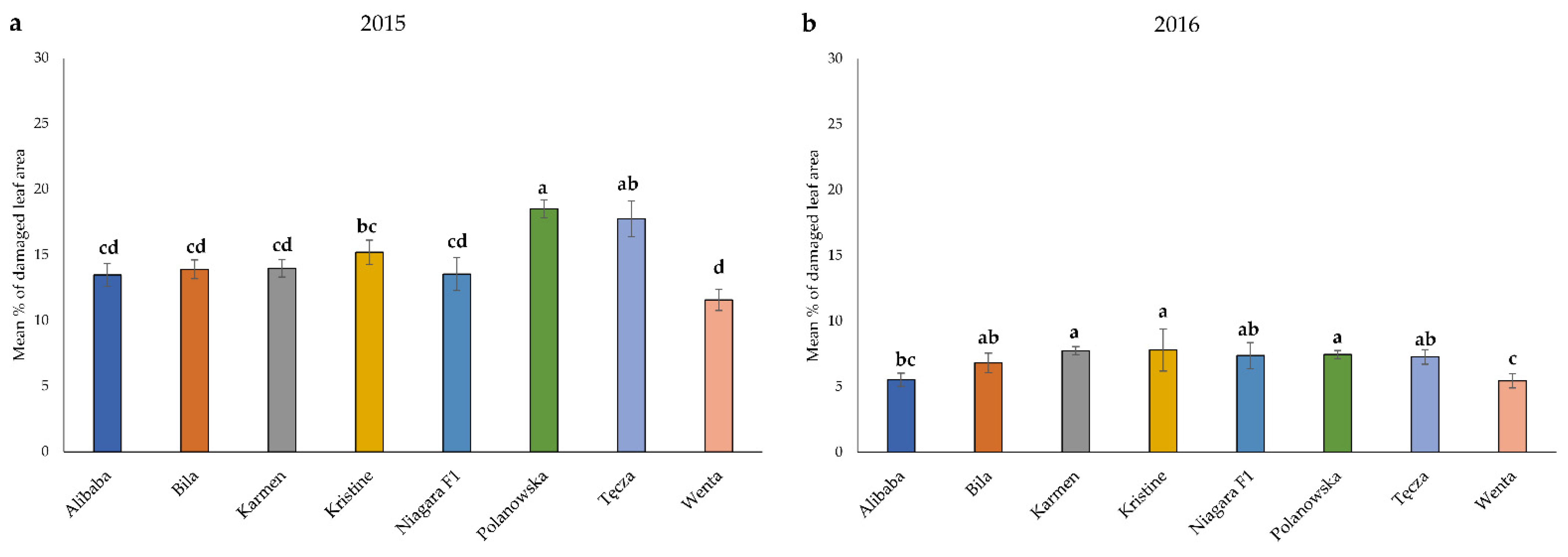
3.1. Field Resistance Assessment
3.2. Biochemical Bases of Resistance
3.3. Anatomical Characters of Leaves
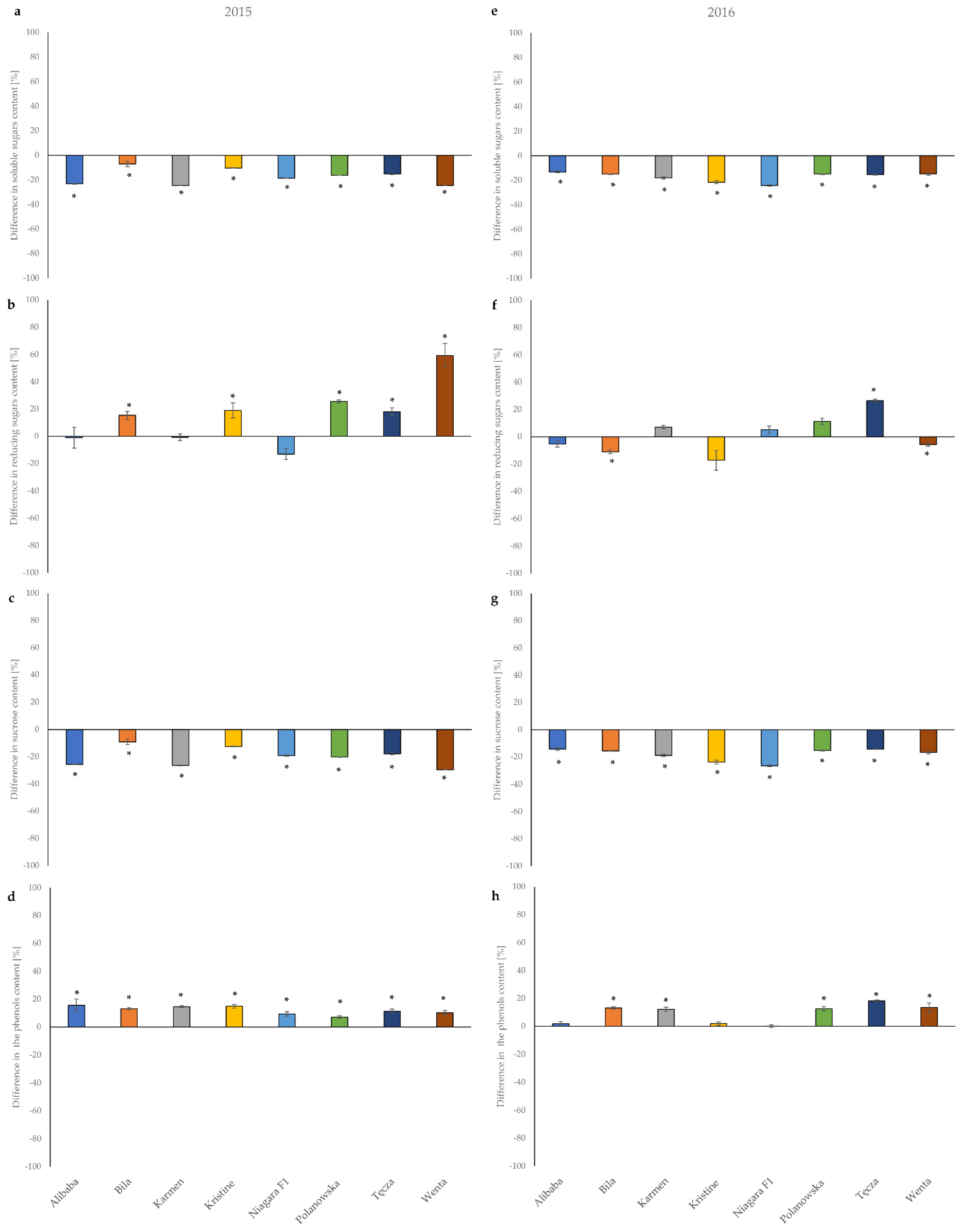
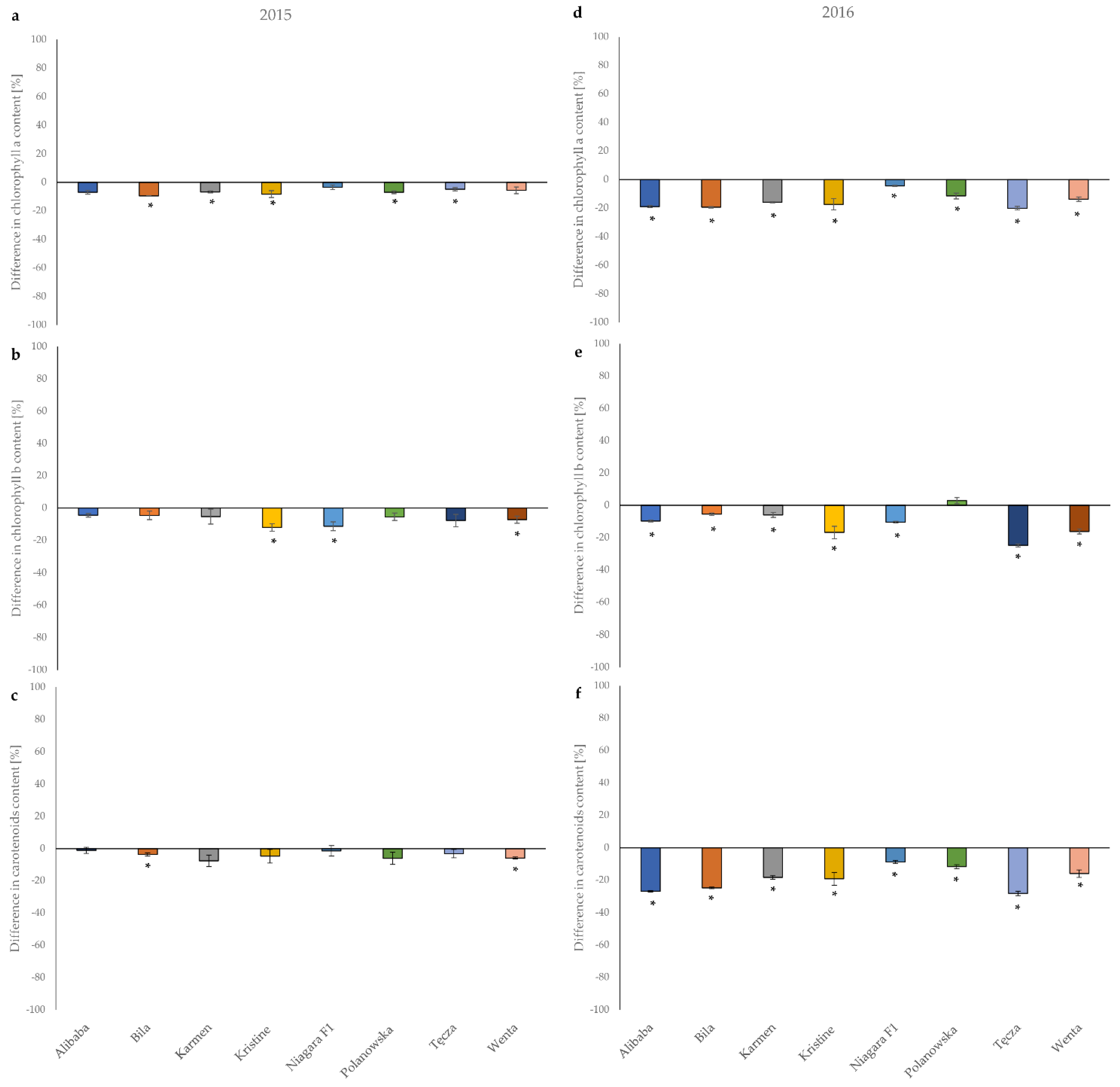
3.4. Effect of Mechanical Injury Caused by Feeding Onion Thrips on the Concentration of Carbohydrates, Total Phenolics, and Leaf Pigments Contents in Onion Leaves
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Onion Production by Country. 2020. Available online: https://www.Atlasbig.Com/En-Ca/Countries-By-Onion-Production (accessed on 2 August 2021).
- Eurostat 2020. Available online: https://ec.europa.eu/Info/Departments/Eurostat-European-Statistics_Pl (accessed on 26 February 2021).
- Diaz-Montano, J.; Fuchs, M.; Nault, B.A.; Fail, J.; Shelton, A.M. Onion thrips (Thysanoptera: Thripidae): A global pest of increasing concern in onion. J. Econ. Entomol. 2011, 104, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lewis, T. Pest thrips in perspective. In Thrips as Crop Pests; Lewis, T., Ed.; CAB International: New York, NY, USA, 1997; pp. 1–13. [Google Scholar]
- Szwejda, J.; Wrzodak, R. Phytophagous entomofauna occurring on onion plantations in Poland in years 1919–2007. J. Fruit Ornam. Plant Res. 2009, 71, 5–14. [Google Scholar] [CrossRef]
- Chisholm, I.F.; Lewis, T. A new look at thrips (Thysanoptera) mouthparts, their action and effects of feeding on plant tissue. Bull. Entomol. Res. 1984, 74, 663–675. [Google Scholar] [CrossRef]
- Kindt, F.; Joosten, N.N.; Tjallingii, W.F. Electrical penetration graphs of thrips revised: Combining DC-and AC-EPG signals. J. Insect Physiol. 2006, 52, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Boateng, C.O.; Schwartz, H.F.; Havey, M.J.; Otto, K. Evaluation of onion germplasm for resistance to Iris yellow spot (Iris yellow spot virus) and onion thrips, Thrips tabaci. Southwest Entomol. 2014, 39, 237–260. [Google Scholar] [CrossRef]
- Ghosheh, H.Z.; Al-Shannag, H.K. Influence of weeds and onion thrips, Thrips tabaci (Thysanoptera: Thripidae), on Onion Bulb Yield in Jordan. Crop Prot. 2000, 19, 175–179. [Google Scholar] [CrossRef]
- Rueda, A.; Badenes-Perez, F.R.; Shelton, A.M. Developing economic thresholds for onion thrips in Honduras. Crop Prot. 2007, 26, 1099–1107. [Google Scholar] [CrossRef]
- Fournier, F.; Boivin, G.; Stewart, R.K. Effect of Thrips tabaci (Thysanoptera: Thripidae) on yellow onion yields and economic thresholds for its management. J. Econ. Entomol. 1995, 88, 1401–1407. [Google Scholar] [CrossRef]
- Waiganjo, M.M.; Gitonga, L.M.; Mueke, J.M. Effects of weather on thrips population dynamics and its implications on the thrips pest management. J. Hortic. Sci. 2008, 1, 82–90. [Google Scholar]
- Diaz-Montano, J.; Fuchs, M.; Nault, B.A.; Fail, J.; Shelton, A.M. Evaluation of onion cultivars for resistance to onion thrips (Thysanoptera: Thripidae) and Iris yellow spot virus. J. Econ. Entomol. 2010, 103, 925–937. [Google Scholar] [CrossRef]
- Dutta, B.; Barman, A.K.; Srinivasan, R.; Avci, U.; Ullman, D.E.; Langston, D.B.; Gitaitis, R.D. Transmission of Pantoea ananatis and P. agglomerans, causal agents of center rot of onion (Allium cepa), by onion thrips (Thrips tabaci) through feces. Phytopathology 2014, 104, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Thind, T.S.; Jhooty, J.S. Association of thrips with purple blotch infection on onion plants caused by Alternaria porri. Indian Phytopathol. 1982, 35, 696–698. [Google Scholar]
- Alston, D.G.; Drost, D. Onion thrips (Thrips tabaci). ENT-117-08PR. Utah Pests Fact Sheet, Utah State University Extension. Utah State University Extension and Utah Plant Pest Diagnostic Laboratory, Logan, UT. 2008. Available online: http://extension.usu.edu/Files/Publications/Factsheet/Ent-117-08pr.Pdf (accessed on 25 February 2021).
- Allen, J.K.M.; Scott-Dupree, C.D.; Tolman, J.H.; Harris, C.R. Resistance of Thrips tabaci to pyrethroid and organophosphorus insecticides in Ontario, Canada. Pest Manag. Sci. 2005, 61, 809–815. [Google Scholar] [CrossRef]
- Shelton, A.M.; Zhao, J.Z.; Nault, B.A.; Plate, J.; Musser, F.R.; Larentzaki, E. Patterns of Insecticide Resistance in Onion Thrips (Thysanoptera: Thripidae) in Onion Fields in New York. J. Econ. Entomol. 2006, 99, 1798–1804. [Google Scholar] [CrossRef] [PubMed]
- Adesanya, A.W.; Waters, T.D.; Lavine, M.D.; Walsh, D.B.; Lavine, L.C.; Zhu, F. Multiple insecticide resistance in onion thrips populations from Western USA. Pestic. Biochem. Physiol. 2020, 165, 104553. [Google Scholar] [CrossRef]
- Moretti, E.A.; Harding, R.S.; Scott, J.G.; Nault, B.A. Monitoring onion thrips (Thysanoptera: Thripidae) susceptibility to spinetoram in New York onion fields. J. Econ. Entomol. 2019, 112, 1493–1497. [Google Scholar] [CrossRef]
- Barzman, M.; Barberi, P.; Birch, N.; Boonekamp, P.; Dachbrodt-Saaydeh, S.; Graf, B.; Hommel, B.; Jensen, J.E.; Kiss, J.; Kudsk, P.; et al. Eight principles of Integrated Pest Management. Agron. Sustain. Dev. 2015, 35, 1199–1215. [Google Scholar] [CrossRef]
- Smith, C.M. Plant Resistance to Arthropods: Molecular and Conventional Approaches, 1st ed.; Springer Science & Business Media: Dordrecht, The Netherlands, 2005. [Google Scholar]
- Capinera, J.L. Integrated pest management: Current concepts and ecological perspective. Fla. Entomol. 2014, 97, 1272. [Google Scholar] [CrossRef]
- Kogan, M.; Ortman, E.F. Antixenosis–A new term proposed to define Painter’s “nonpreference” modality of resistance. Bull. Entmol. Soc. Am. 1978, 24, 175–176. [Google Scholar] [CrossRef]
- Painter, R.H. Insect Resistance in Crop Plants; Macmillan: New York, NY, USA, 1951. [Google Scholar] [CrossRef]
- Diaz-Montano, J.; Fail, J.; Deutschlander, M.; Nault, B.A.; Shelton, A.M. Characterization of resistance, evaluation of the attractiveness of plant odors, and effect of leaf colour on different onion cultivars to onion thrips (Thysanoptera: Thripidae). J. Econ. Entomol. 2012, 105, 632–641. [Google Scholar] [CrossRef]
- Fail, J.; Deutschlander, M.E.; Shelton, A.M. Antixenotic resistance of cabbage to onion thrips (Thysanoptera: Thripidae). I. Light reflectance. J. Econ. Entomol. 2013, 106, 2602–2612. [Google Scholar] [CrossRef]
- Koschier, E.H.; Sedy, K.A.; Novak, J. Influence of plant volatiles on feeding damage caused by the onion thrips Thrips tabaci. Crop Prot. 2002, 21, 419–425. [Google Scholar] [CrossRef]
- Brown, A.S.S.; Simmonds, M.S.; Blaney, W.M. Relationship between nutritional composition of plant species and infestation levels of thrips. J. Chem. Ecol. 2002, 28, 2399–2409. [Google Scholar] [CrossRef] [PubMed]
- Pobożniak, M.; Koschier, E.H. Effects of pea (Pisum sativum L.) cultivars on Thrips tabaci Lindeman preference and performance. J. Agric. Sci. 2014, 152, 885–893. [Google Scholar] [CrossRef]
- Leiss, K.A.; Cristofori, G.; van Steenis, R.; Verpoorte, R.; Klinkhamer, P.G. An eco-metabolomic study of host plant resistance to western flower thrips in cultivated, biofortified and wild carrots. Phytochemistry 2013, 93, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Steenbergen, M.; Abd-el-Haliem, A.; Bleeker, P.; Dicke, M.; Escobar-Bravo, R.E.R.; Cheng, G.; Haring, M.A.; Kant, M.R.; Kappers, I.; Klinkhamer, P.G.L.; et al. Thrips advisor: Exploiting thrips-induced defenses to combat pests on crops. J. Exp. Bot. 2018, 69, 1837–1848. [Google Scholar] [CrossRef]
- Morsello, S.C.; Groves, R.L.; Nault, B.A.; Kennedy, G.G. Temperature and precipitation affect seasonal patterns of dispersing tobacco thrips, Frankliniella fusca, and onion thrips, Thrips tabaci (Thysanoptera: Thripidae) caught on sticky traps. Environ. Entomol. 2008, 37, 79–86. [Google Scholar] [CrossRef]
- Fok, E.J.; Petersen, J.D.; Nault, B.A. Relationships between insect predator populations and their prey, Thrips tabaci, in onion fields grown in large-scale and small-scale cropping systems. BioControl 2014, 59, 739–748. [Google Scholar] [CrossRef][Green Version]
- Mala, B.; Nighot, P. Natural enemy complex of Thrips tabaci Lindeman in onion and garlic. Pest Manag. Hort. Ecosyst. 2013, 19, 121–122. [Google Scholar]
- Damon, S.J.; Groves, R.L.; Havey, M.J. Variation for epicuticular waxes on onion foliage and impacts on numbers of onion thrips. J. Am. Soc. Hortic. Sci. 2014, 139, 495–501. [Google Scholar] [CrossRef]
- Munaiz, E.D.; Groves, R.L.; Havey, M.J. Amounts and types of epicuticular leaf waxes among onion accessions selected for reduced damage by onion thrips. J. Am. Soc. Hortic. Sci. 2020, 145, 30–35. [Google Scholar] [CrossRef]
- da Silva, V.C.P.; Bettoni, M.M.; Bona, C.; Foerster, L.A. Morphological and chemical characteristics of onion plants (Allium cepa L.) associated with resistance to onion thrips. Acta Sci. Agron. 2014, 37, 85–92. [Google Scholar] [CrossRef]
- Alimousavi, S.A.; Hassandokht, M.R.; Moharramipour, S.A.E.I.D. Evaluation of Iranian onion germplasms for resistance to thrips. Int. J. Agric. Biol. 2007, 9, 897–900. [Google Scholar]
- Njau, G.M.; Nyomora, A.M.; Dinssa, F.F.; Chang, J.C.; Malini, P.; Subramanian, S.; Srinivasan, R. Evaluation of onion (Allium cepa) germplasm entries for resistance to onion thrips, Thrips tabaci (Lindeman) in Tanzania. Int. J. Trop. Insect Sci. 2017, 37, 98–113. [Google Scholar] [CrossRef]
- Pobożniak, M.; Leśniak, M.; Chuda, A.; Adamus, A. Field assessment of the susceptibility of onion cultivars to thrips attack–preliminary results. Pol. J. Entomol. 2016, 85, 121–133. [Google Scholar] [CrossRef][Green Version]
- Pobożniak, M.; Olczyk, M.; Wójtowicz, T. Relationship between colonization by onion thrips (Thrips tabaci Lind.) and leaf colour measures across eight onion cultivars (Allium cepa L.). Agronomy 2021, 11, 963. [Google Scholar] [CrossRef]
- Zawirska, I. Thrips (Thysanoptera). In Diagnostics of Plant Pests and Their Natural Enemies; Kozłowski, M.W., Boczek, J., Eds.; SGGW: Warsaw, Poland, 1994; pp. 145–174. [Google Scholar]
- Strassen, R.Z. Die terebranten thysanopteren Europas und des Mittelmeer-Gebietes. Tierwelt Dtschl. 2003, 74, 1–277. [Google Scholar]
- Kucharczyk, H. Comparative Morphology of the Second Larval Instar of the Thrips Genus Species (Thysanoptera: Thripidae) Occurring in Poland; Mantis: Olsztyn, Poland, 2010; ISBN 978-83-929997-7-5. [Google Scholar]
- Edelson, J.V.; Magaro, J.J. Development of onion thrips, Thrips tabaci Lindeman, as a function of temperature. Southwest Entomol. 1988, 13, 171–176. [Google Scholar]
- Murai, T. Effect of temperature on development and reproduction of the onion thrips, Thrips tabaci Lindeman (Thysanoptera: Thripidae), on pollen and honey solution. Appl. Entomol. Zool. 2000, 35, 499–504. [Google Scholar] [CrossRef]
- Samotus, B.; Tuz, J.; Doerre, E. Evaluation of Blue Value in different plant materials as a tool for rapid starch determination. Acta Soc. Bot. Pol. 1993, 62, 137–141. [Google Scholar] [CrossRef]
- Nath, N.; Singh, M.P. Mechanism of the oxidation of reducing sugars (hexoses) by hexacyanoferrate (III) in alkaline medium and Lobry de Bruyn transformation. J. Phys. Chem. 1965, 69, 2038–2043. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. Soc. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Knolhoff, L.; Heckel, D.G. Behavioral assays for studies of host plant choice and adaptation in herbivorous insects. Ann. Rev. Entomol. 2014, 59, 263–278. [Google Scholar] [CrossRef]
- Bálint, J.; Burghardt, N.; Höhn, M.M.; Pénzes, B.; Fail, J. Does epidermal thickness influence white cabbage resistance against onion thrips (Thrips tabaci)? Not. Bot. Horti Agrobot. Cluj-Napoca 2013, 41, 444–449. [Google Scholar] [CrossRef][Green Version]
- Terry, L.I. Host Selection, Communication and Reproductive Behaviour. In Thrips as Crop Pests; Lewis, T., Ed.; CAB International: Wallingford, UK, 1997; pp. 65–118. [Google Scholar]
- Cramer, C.S.; Singh, N.; Kamal, N.; Pappu, H.R. Screening onion plant introduction accessions for tolerance to onion thrips and iris yellow spot. Hortic. Sci. 2014, 49, 1253–1261. [Google Scholar] [CrossRef]
- Eigenbrode, S.D.; Castagnola, T.; Roux, M.B.; Steljes, L. Mobility of three generalist predators is greater on cabbage with glossy leaf wax than on cabbage with a wax bloom. Entomol. Exp. Appl. 1996, 81, 335–343. [Google Scholar] [CrossRef]
- Trdan, S.; Znidarcic, D.; Zlatic, E.; Jerman, J. Correlation between epicuticular wax content in the leaves of early white cabbage (Brassica oleracea L. var. capitata) and damage caused by Thrips tabaci Lindeman (Thysanoptera: Thripidae). Acta Phytopathol. Entomol. Hung. 2004, 39, 173–185. [Google Scholar] [CrossRef]
- Voorrips, R.E.; Steenhuis-Broers, G.; Tiemens-Hulscher, M.; Lammerts van Bueren, E.T. Earliness, leaf surface wax and sugar content predict varietal differences for thrips damage in cabbage. In Proceedings of the 5th International Symposium on Brassicas and 16th Crucifer Genetics Work Shop, Lillehammer, Norway, 8–12 September 2008; Acta Hort: Lillehammer, Norway, 2010; Volume 867, pp. 127–132. [Google Scholar] [CrossRef]
- Lewis, T. Thrips, Their Biology, Ecology and Economic Importance, 1st ed.; Academic Press: London, UK, 1973; p. 349. [Google Scholar]
- Mollema, C.; Steehuis, G.; Ingammer, H. Genotypic effect of cucumber responses to infestation by western flower thrips. In Thrips Biology and Management; Parker, B.L., Skinner, M., Lewis, T., Eds.; Plenum Press: New York, NY, USA, 1995; pp. 397–401. [Google Scholar] [CrossRef]
- Miyazaki, J.; Stiller, W.N.; Wilson, L.J. Sources of plant resistance to thrips: A potential core component in cotton IPM. Entomol. Exp. Appl. 2016, 162, 30–40. [Google Scholar] [CrossRef]
- Ananthakrishnan, T.N.; Gopichandran, R. Chemical Ecology in Thrips-Host Plant Interactions; Oxford & IBH Publishers: New Delhi, India, 1993. [Google Scholar]
- Agbahoungba, S.; Karungi, J.; Badji, A.; Sadik, K.; Gibson, P.; Edema, R.; Assogbadjo, A.E.; Rubaihayo, P.R. Inheritance of cowpea resistance to flower thrips in Uganda germplasm. J. Plant Breed. Crop Sci. 2018, 10, 21–32. [Google Scholar] [CrossRef]
- Kandakoor, S.B.; Khan, H.K.; Chakravarthy, A.K.; Kumar, C.A.; Venkataravana, P. Biochemical constituents influencing thrips resistance in groundnut germplasm. J. Environ. Biol. 2014, 35, 675–681. [Google Scholar] [PubMed]
- Chandrayudu, E.; Vemena, K.; Naik, B.S.; Prathyusha, C. Biophysical and biochemical constituents influencing thrips and jassid resistance in groundnut germplasm. In Natural Resource Management: Ecological Perspectives, Proceedings of the Indian Ecological Society International Conference, Jammu, India, 18–20 February 2016; Peshin, R., Dhawan, A.K., Bano, F., Risam, K.S., Eds.; The Indian Ecological Society: Jammu, India, 2016; Volume 2, pp. 780–782. [Google Scholar]
- Bhonde, B.N.; Pawar, D.B.; Nikam, D.P. Role of biochemicals in imparting resistance in onion against onion thrips Thrips tabaci Lindeman. Adv. Life Sci. 2016, 5, 8798–8800. [Google Scholar]
- Žnidarčič, D.; Vidrih, R.; Germ, D.; Ban, D.; Trdan, S. Relationship between water-soluble carbohydrate composition of cabbage (Brassica oleracea L. var. capitata) and damage levels of onion thrips. Acta Agric. Slov. 2007, 89, 25–33. [Google Scholar] [CrossRef]
- Srinivas, P.S.; Aziz Qureshi, A.; Lawande, K.E. Growth stage susceptibility of onion (Allium cepa) and its role in thrips management. Indian J. Agric. Sci. 2008, 78, 98–101. [Google Scholar]
- Bennett, R.N.; Wallsgrove, R.M. Secondary metabolites in plant defense mechanisms. New Phytol. 1994, 127, 617–633. [Google Scholar] [CrossRef] [PubMed]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett. 2006, 4, 147–157. [Google Scholar] [CrossRef]
- Ahmed, A.; Radi, A.; Tolba, E. Efficiency of phenolic contents and some field factors on the population abundance of aphid species and onion thrips infesting bread wheat cultivars under irrigation conditions. J. Plant Prot. Pathol. 2019, 10, 69–75. [Google Scholar] [CrossRef]
- Akhtari, M.; Dashti, F.; Madadi, H.; Rondon, S. Evaluation of resistance to onion thrips (Thrips tabaci Lind.) in several Tareh Irani (Persian leek: Allium ampeloprasum Tareh group) landraces. Arch. Phytopathol. Plant Prot. 2014, 47, 29–41. [Google Scholar] [CrossRef]
- Miles, P.W. Aphid saliva. Biol. Rev. Camb. Philos. Soc. 1999, 74, 41–85. [Google Scholar] [CrossRef]
- Felton, G.W.; Donato, K.K.; Broadway, R.M.; Duffey, S.S. Impact of oxidized plant phenolics on the nutritional quality of dietary protein to a noctuid herbivore, Spodoptera exigua. J. Insect Physiol. 1992, 38, 277–285. [Google Scholar] [CrossRef]
- Leiss, K.A.; Choi, Y.H.; Abdel-Farid, I.B.; Verpoorte, R.; Klinkhamer, P.G. NMR metabolomics of thrips (Frankliniella occidentalis) resistance in Senecio hybrids. J. Chem. Ecol. 2009, 35, 219–229. [Google Scholar] [CrossRef]
- Miles, P.W.; Oertli, J.J. The significance of antioxidants in the aphid-plant interaction: The redox hypothesis. Entomol. Exp. Appl. 1993, 67, 275–283. [Google Scholar] [CrossRef]
- Havlíčková, H.; Cvikrová, M.; Eder, J. Phenolic acids in wheat cultivars in relation to plant suitability for and response to cereal aphids. J. Plant Dis. Prot. 1996, 103, 535–542. [Google Scholar]
- Ananthakrishnan, T.N.; Gopichandran, R.; Gurusubramanian, G. Influence of chemical profiles of host plants on the infestation diversity of Retithrips syriacus. J. Biosci. 1992, 17, 483–489. [Google Scholar] [CrossRef]
- Khalil, H.; Raza, A.B.M.; Afzal, M.; Aqueel, M.A.; Khalil, M.S.; Mansoor, M.M. Effects of plant morphology on the incidence of sucking insect pests complex in few genotypes of cotton. J. Saudi Soc. Agric. Sci. 2017, 16, 344–349. [Google Scholar] [CrossRef]
- Raza, A.B.M.; Afzal, M.; Sarfraz, M.; Bashir, M.H.; Gogi, M.D.; Sarwar, M.S. Physico-morphic plant characters in relation to resistance against sucking insect pests in some new cotton genotypes. Pak. Entomol. 2000, 22, 73–77. [Google Scholar]
- Bowman, D.T.; McCarty, J.C., Jr. Thrips (Thysanoptera: Thripidae) tolerance in cotton: Sources and heritability. J. Entomol. Sci. 1997, 32, 460–471. [Google Scholar] [CrossRef]
- Hanafy, A.R.I.; Tahany, R.A.; Nowar, E.E.; Hasan, S.M. Effect of anatomical and phytochemical diversity of two onion cultivars on the infestation with onion thrips (Thysanoptera: Thripidae). Middle East J. Appl. Sci. 2016, 6, 941–948. [Google Scholar]
- Abdel-Gawaad, A.A.W.; El-Gayar, F.H.; Soliman, A.S.; Zaghlool, O.A. Studies on Thrips tabaci Lindman: X. Mechanism of resistance to Thrips tabaci L. in cotton varieties. J. Appl. Entomol. 1973, 73, 251–255. [Google Scholar] [CrossRef]
- Childers, C.C.; Achor, D.S. Feeding and oviposition injury to flowers and developing floral buds of ‘Navel’ orange by Frankliniella bispinosa (Thysanoptera: Thripidae) in Florida. Ann. Entomol. Soc. Am. 1991, 84, 272–282. [Google Scholar] [CrossRef]
- Harrewijn, P.; Tjallingii, W.F.; Mollema, C. Electrical recording of plant penetration by western flower thrips. Entomol. Exp. Appl. 1996, 79, 345–353. [Google Scholar] [CrossRef]
- Kucharczyk, H.; Kucharczyk, M.; Winiarczyk, K.; Lubiarz, M.; Tchórzewska, D. Effects of temperature on the development of Thrips nigropilosus Uzel (Thysanoptera: Thripidae) on Mentha × piperita L. and the impact of pest on the host plant. Acta Sci. Pol. Hortoru. Cultus 2019, 18, 219–233. [Google Scholar] [CrossRef]
- Peterson, R.K.; Varella, A.C.; Higley, L.G. Tolerance: The forgotten child of plant resistance. Peer J. 2017, 5, e3934. [Google Scholar] [CrossRef]
- Johari, A.; Herlinda, S.; Irsan, C.; Pujiastuti, Y. Phenomenon of thrips (Thysanoptera) attack on chili plant (Capsicum annuum L.). Am. J. Agric. Biol. Sci. 2016, 11, 103–109. [Google Scholar] [CrossRef]
- Yu, D.; Huang, P.; Chen, Y.; Lin, Y.; Akutse, K.S.; Lan, Y.; Wei, H. Effects of flower thrips (Thysanoptera: Thripidae) on nutritional quality of banana (Zingiberales: Musaceae) buds. PLoS ONE 2018, 13, e0202199. [Google Scholar] [CrossRef]
- Strauss, S.Y.; Agrawal, A.A. The ecology and evolution of plant tolerance to herbivory. Trends Ecol. Evol. 1999, 14, 179–185. [Google Scholar] [CrossRef]
- Tiffin, P. Mechanisms of tolerance to herbivore damage: What do we know? Evol. Ecol. 2000, 14, 523–536. [Google Scholar] [CrossRef]
- Dai, Y.; Shao, M.; Hannaway, D.; Wang, L.; Liang, J.; Hu, L.; Lu, H. Effect of Thrips tabaci on anatomical features, photosynthetic characteristics and chlorophyll fluorescence of Hypericum sampsonii leaves. Crop Prot. 2009, 28, 327–332. [Google Scholar] [CrossRef]
- Naidu, R.A.; Krishnan, M.; Nayudu, M.V.; Gnanam, A. Studies on peanut green mosaic virus infected peanut (Arachis hypogaea L.) leaves. II. Chlorophyll-protein complexes and polypeptide composition of thylakoid membranes. Physiol. Plant Pathol. 1984, 25, 191–198. [Google Scholar] [CrossRef]
- Sestak, Z. Limitations for finding linear relationship between chlorophyll content and photosynthetic activity. Biol. Plant. 1996, 8, 336–346. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicators | Year | Scale | |||
|---|---|---|---|---|---|
| 4 Points | 3 Points | 2 Points | 1 Point | ||
| Mean number of migrating adult thrips per 10 plants | 2015 | 0.00–3.75 | 3.76–6.75 | 6.76–10.75 | >10.75 |
| 2016 | 0.00–1.50 | 1.51–3.50 | 3.51–5.50 | >5.50 | |
| Seasonal mean number of adults and larvae of thrips per 10 plants | 2015 | 0.00–7.50 | 7.51–10.00 | 10.01–12.00 | >12.00 |
| 2016 | 0.00–2.75 | 2.76–4.75 | 4.76–7.00 | >7.00 | |
| Mean percentage of damaged leaf area by feeding thrips | 2015 | 0.00–10.00 | 10.01–13.00 | 13.01–17.50 | >17.50 |
| 2016 | 0.00–5.00 | 5.01–6.50 | 6.51–7.25 | >7.25 | |
| Cultivars | Resistance to Plant Colonization by Migrating Adult Thrips | Field Resistance throughout the Growing Season | ||||
|---|---|---|---|---|---|---|
| Resistance to Thrips Abundance | Resistance to Thrips Damage | |||||
| No. Points | Level of Resistance | No. Points | Level of Resistance | No. Points | Level of Resistance | |
| Alibaba | 1.5 | susceptible | 1.5 | susceptible | 2.0 | susceptible |
| Bila | 2.0 | susceptible | 2.5 | susceptible | 2.0 | susceptible |
| Karmen | 1.5 | susceptible | 2.0 | susceptible | 1.5 | susceptible |
| Kristine | 1.5 | susceptible | 2.5 | susceptible | 1.5 | susceptible |
| Niagara F1 | 1.0 | high susceptible | 1.0 | high susceptible | 1.5 | susceptible |
| Polanowska | 1.5 | susceptible | 2.0 | susceptible | 1.0 | high susceptible |
| Tęcza | 3.5 | resistant | 4.0 | resistant | 1.5 | susceptible |
| Wenta | 3.0 | Moderately resistant | 2.5 | Moderately resistant | 4.0 | resistant |
| Cultivar | Mean Quantity (±SE) (mg/100 g FW 1) | |||
|---|---|---|---|---|
| Soluble Sugars | Reducing Sugars | Sucrose | Total Phenols | |
| Alibaba | 1.25 ± 0.012 b 2 | 0.125 ± 0.012 a | 1.13 ± 0.004 b | 97.23 ± 1.24 b |
| Bila | 0.91 ± 0.024 e | 0.077 ± 0.003 bc | 0.84 ± 0.021 e | 72.63 ± 0.33 d |
| Karmen | 1.13 ± 0.002 c | 0.084 ± 0.001 b | 1.04 ± 0.002 c | 74.85 ± 0.15 d |
| Kristine | 0.91 ± 0.002 e | 0.067 ± 0.007 c | 0.85 ± 0.002 e | 64.75 ± 0.88 d |
| Niagara F1 | 1.15 ± 0.002 c | 0.124 ± 0.008 a | 1.02 ± 0.006 c | 81.50 ± 0.62 c |
| Polanowska | 0.86 ± 0.007 f | 0.074 ± 0.001 bc | 0.78 ± 0.006 f | 73.24 ± 0.43 d |
| Tęcza | 1.05 ± 0.007 d | 0.079 ± 0.002 b | 0.97 ± 0.005 d | 68.08 ± 0.85 d |
| Wenta | 1.35 ± 0.008 a | 0.077 ± 0.006 bc | 1.27 ± 0.004 a | 99.93 ± 1.37 a |
| p cultivar | <0.001 | <0.001 | <0.001 | <0.001 |
| F blocks | 8.6 | 11.657 | 1.9 | 0.58 |
| p blocks | <0.001 | <0.001 | 0.156 | 0.636 |
| Cultivar | Mean Quantity (±SE) (mg/100 g FW 1 ) | |||
|---|---|---|---|---|
| Soluble Sugars | Reducing Sugars | Sucrose | Total Phenols | |
| Alibaba | 1.17 ± 0.011 b 2 | 0.13 ± 0.001 a | 1.04 ± 0.010 c | 74.82 ± 1.23 a |
| Bila | 0.96 ± 0.009 f | 0.10 ± 0.001 c | 0.87 ± 0.008 f | 54.83 ± 0.27 d |
| Karmen | 1.19 ± 0.016 b | 0.09 ± 0.003 d | 1.10 ± 0.014 b | 57.92 ± 0.30 c |
| Kristine | 0.94 ± 0.028 f | 0.05 ± 0.004 f | 0.88 ± 0.024 f | 47.91 ± 0.54 d |
| Niagara F1 | 1.09 ± 0.010 c | 0.11 ± 0.001 b | 0.97 ± 0.009 d | 67.55 ± 0.68 b |
| Polanowska | 1.37 ± 0.014 a | 0.12 ± 0.002 b | 1.25 ± 0.013 a | 49.69 ± 0.70 d |
| Tęcza | 1.03 ± 0.08 d | 0.10 ± 0.001 c | 0.93 ± 0.007 e | 76.26 ± 0.69 a |
| Wenta | 1.09 ± 0.019 e | 0.08 ± 0.003 e | 0.92 ± 0.016 e | 75.86 ± 1.87 a |
| p cultivar | <0.001 | <0.001 | <0.001 | <0.001 |
| F blocks | 16.3 | 9.12 | 15.5 | 0.77 |
| p blocks | <0.001 | <0.001 | 0.005 | 0.523 |
| Parameters | No. of Thrips (Imago + Larvae) | Damaged Leaf Area | ||||||
|---|---|---|---|---|---|---|---|---|
| Mean No. of Thrips on 28 July | Seasonal Mean No. of Thrips | Mean Percentage of Damaged Leaf Area on 28 July | Seasonal Mean Percentage of Damaged Leaf Area | |||||
| r | p | r | p | r | p | r | p | |
| Mean quantity of total soluble sugars (g/100−1 g FW) | 0.026 | 0.146 | 0.115 | 0.530 | −0.305 | 0.089 | −0.589 * | 0.000 |
| Mean quantity of reducing sugars (g/100−1 g FW) | 0.579* | 0.001 | 0.489 * | 0.004 | −0.216 | 0.234 | −0.281 | 0.120 |
| Mean quantity of sucrose (g/100−1 g FW) | 0.194 | 0.287 | 0.051 | 0.785 | −0.293 | 0.104 | −0.586 * | 0.000 |
| Mean total phenols contents (mg/100−1 g FW) | 0.313 | 0.081 | 0.082 | 0.655 | −0.253 | 0.162 | −0.534 * | 0.002 |
| Parameters | No. of Thrips (Imago + Larvae) | Damaged Leaf Area | ||||||
|---|---|---|---|---|---|---|---|---|
| Mean No. of Thrips on 8 August | Seasonal Mean No. of Thrips | Mean Percentage of Damaged Leaf Area on 8 August | Seasonal Mean Percentage of Damaged Leaf Area | |||||
| r | p | r | p | r | p | r | p | |
| Mean quantity of total sugars (g/100−1 g FW) | 0.523 * | 0.002 | 0.319 | 0.075 | 0.081 | 0.655 | 0.598 | 0.745 |
| Mean quantity of reducing sugars (g/100−1 g FW) | 0.395 * | 0.025 | 0.397 * | 0.024 | −0.120 | 0.513 | −0.246 | 0.175 |
| Mean quantity of sucrose (g/100−1 g FW) | 0.508 * | 0.003 | 0.282 | 0.118 | 0.112 | 0.543 | 0.110 | 0.548 |
| Mean total phenols contents (mg/100−1 g FW) | −0.124 | 0.500 | 0.411 | 0.823 | −0.390 * | 0.027 | −0.421 * | 0.016 |
| Cultivar | Max. Leaf Blade Thickness (Mean ± SE) (µm) | Min. Leaf Blade Thickness (Mean ± SE) (µm) | Mean Leaf Blade Thickness (Mean ± SE) (µm) |
|---|---|---|---|
| Alibaba | 528.6 ± 6.84 c 1 | 520.6 ± 6.86 a | 524.6 ± 6.74 a |
| Bila | 495.4 ± 2.73 d | 347.0 ± 5.63 e | 421.2 ± 2.73 d |
| Karmen | 598.4 ± 5.71 a | 432.0 ± 3.66 b | 515.2 ± 2.72 a |
| Kristine | 566.0 ± 7.44 b | 409.6 ± 27.51 bc | 487.8 ± 17.24 b |
| Niagara F1 | 451.6 ± 3.53 e | 376.4 ± 5.45 cd | 414.0 ± 2.24 d |
| Polanowska | 385.0 ± 4.37 g | 367.2 ± 2.17 de | 376.1 ± 2.35 f |
| Tęcza | 503.8 ± 3.55 d | 403.6 ± 4.74 bc | 453.7 ± 2.78 c |
| Wenta | 435.2 ± 3.68 f | 357.8 ± 4.29 de | 396.5 ± 2.21 e |
| F cultivar | 217.00 | 25.3 | 77.0 |
| p cultivar | <0.001 | <0.001 | <0.001 |
| Cultivar | Epidermis | Mesophyll | ||||
|---|---|---|---|---|---|---|
| Mean (±SE) | Cell Layers (No) | Mean (±SE) | ||||
| Thickness (µm) | Perimeter (µm) | Area (±SE) (µm2 ) | Perimeter (µm) | Area (µm2 ) | ||
| Alibaba | 25.65 ± 0.96 b 1 | 84.23 ± 9.70 c | 482.06 ± 26.26 c | 7 | 115.98 ± 10.51 e | 734.04 ± 131.19 d |
| Bila | 27.38 ± 0.79 ab | 100.33 ± 15.01 ab | 654.24± 35.99 ab | 8 | 208.85 ± 12.39 a | 1579.52 ± 151.44 abc |
| Karmen | 27.61 ± 1.07 ab | 96.50 ± 18.58 b | 615.49 ± 43.88 bc | 7 | 194.89 ± 9.19 ab | 1652.59 ±116.56 a |
| Kristine | 29.95 ± 0.65 a | 107.78 ± 9.22 a | 775.38 ± 26.19 a | 6 | 192.20 ± 14.23 abc | 1780.38 ± 198.65 ab |
| Niagara F1 | 26.28 0.97 b | 93.77 ± 11.26 b | 568.00 ± 31.80 bc | 6 | 152.49 ± 15.42 cde | 1121.54 ± 168.21 bc |
| Polanowska | 26.08 ± 1.56 b | 108.15 ± 18.40 a | 789.77 ± 68.38 a | 7 | 140.02 ±15.94 cd | 1084.48 ± 171.63 cd |
| Tęcza | 25.38 ± 1.01 b | 99.88 ± 18.50 ab | 687.90 ± 52.03 ab | 6 | 166.41 ± 11.31 abcd | 1480.72 ±156.18 abc |
| Wenta | 24.28 ± 1.10 b | 95.23 ± 17.20 b | 594.28 ± 46.91 bc | 8 | 153.72 ± 11.10 bcd | 1142.13 ±140.64 abc |
| F cultivar | 3.17 | 4.70 | 4.71 | 4.88 | 4.34 | |
| p cultivar | 0.004 | <0.001 | <0.001 | <0.001 | <0.001 | |
| Cultivar | Mean Diameter (±SE) (µm) | Mean Perimeter (±SE) (µm) | Mean Cross-Section Area (±SE) (µm2) | Mean Distance Between VB (±SE) (µm) |
|---|---|---|---|---|
| Alibaba | 156.67 ± 9.95 | 478.00 ± 12.16 | 13,077.33 ± 418.41 | >1000.00 a 1 |
| Bila | 118.33 ± 19.01 | 594.67 ± 101.13 | 17,144.00 ± 5637.38 | 831.68 ± 88.52 ab |
| Karmen | 170.00 ± 57.41 | 504.00 ± 153.13 | 16,817.67 ± 7145.38 | 744.33 ±128.19 abc |
| Kristine | 154.67 ± 40.68 | 470.00 ± 103.92 | 12,752.00 ± 3995.18 | 438.00 ± 22.19 bcd |
| Niagara F1 | 53.00 ±3.51 | 289.00 ± 46.36 | 4836.67 ± 1573.59 | 347.67 ± 10.82 cd |
| Polanowska | 76.67 ± 12.17 | 241.00 ± 30.80 | 3936.00 ± 1000.16 | 264.33 ± 65.46 d |
| Tęcza | 116.67 ± 52.44 | 343.66 ± 134.55 | 7752.33 ± 5078.374 | 496.00 ± 252.68 cd |
| Wenta | 60.00 ± 19.03 | 212.67 ± 68.91 | 3059.33 ± 1945.752 | 454.00 ± 112.01 bcd |
| F cultivar | 2.191 | 2.146 | 2.164 | 4.587 |
| p cultivar | 0.092 | 0.098 | 0.095 | 0.005 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pobożniak, M.; Olczyk, M.; Wójtowicz, T.; Kamińska, I.; Hanus-Fajerska, E.; Kostecka-Gugała, A.; Kruczek, M. Anatomical and Biochemical Traits Associated with Field Resistance of Onion Cultivars to Onion Thrips and the Effect of Mechanical Injury on the Level of Biochemical Compounds in Onion Leaves. Agronomy 2022, 12, 147. https://doi.org/10.3390/agronomy12010147
Pobożniak M, Olczyk M, Wójtowicz T, Kamińska I, Hanus-Fajerska E, Kostecka-Gugała A, Kruczek M. Anatomical and Biochemical Traits Associated with Field Resistance of Onion Cultivars to Onion Thrips and the Effect of Mechanical Injury on the Level of Biochemical Compounds in Onion Leaves. Agronomy. 2022; 12(1):147. https://doi.org/10.3390/agronomy12010147
Chicago/Turabian StylePobożniak, Maria, Marta Olczyk, Tomasz Wójtowicz, Iwona Kamińska, Ewa Hanus-Fajerska, Anna Kostecka-Gugała, and Michał Kruczek. 2022. "Anatomical and Biochemical Traits Associated with Field Resistance of Onion Cultivars to Onion Thrips and the Effect of Mechanical Injury on the Level of Biochemical Compounds in Onion Leaves" Agronomy 12, no. 1: 147. https://doi.org/10.3390/agronomy12010147
APA StylePobożniak, M., Olczyk, M., Wójtowicz, T., Kamińska, I., Hanus-Fajerska, E., Kostecka-Gugała, A., & Kruczek, M. (2022). Anatomical and Biochemical Traits Associated with Field Resistance of Onion Cultivars to Onion Thrips and the Effect of Mechanical Injury on the Level of Biochemical Compounds in Onion Leaves. Agronomy, 12(1), 147. https://doi.org/10.3390/agronomy12010147