
Seasonal Dynamics of Dry Matter Accumulation and Nutrients in a Mature Miscanthus × giganteus Stand in the Lower Silesia Region of Poland

 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
- to determine height of plants and the accumulation of the dry matter for stems, leaves and rhizomes,
- to measure variations in N, P, K, Ca and Mg concentrations, and differentiate accumulation of these elements in different parts of the plant; aerial parts (stems and leaves) and the underground rhizome,
- to measure harvestable dry matter yield and composition, and, to determine how the flux of nutrients throughout seasonal growth impacts the quality of harvested biomass across three harvest years with differing meteorological conditions and accumulation of biomass,
- to compare measured and modelled yield to examine how well standard model assumptions predict yield in diverse environments.
2. Materials and Methods
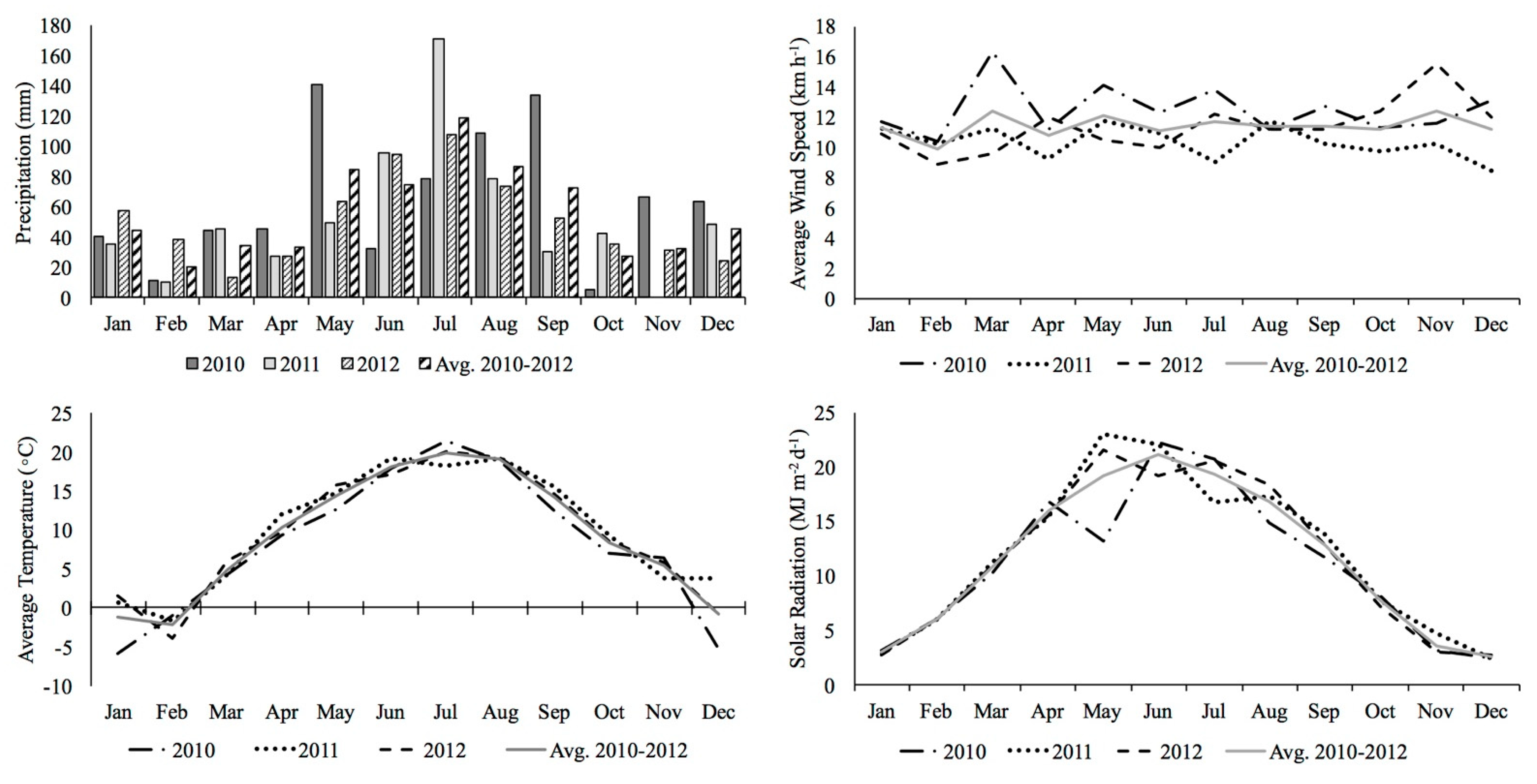
2.1. Experimental Site and Climatic Conditions
2.2. Agro-Technical and Soil Conditions
- -
- humus horizon 0–26 (30) cm deep: subangular blocky structures, consistence soft, firm; slightly loamy sand;
- -
- transitional soil horizon 30–38 (40) cm deep: subangular blocky structures, consistence soft, firm; slightly loamy sand;
- -
- parent material 40–66 (70) cm deep: subangular blocky structures, consistence soft, firm; loose sand;
- -
- parent material 70–88 cm deep: subangular blocky structures, consistence soft, firm; loose sand;
- -
- glial horizon +88 cm deep: single-grained structure, consistence loose; loose gravel sand.
2.3. Research Methods
2.4. Chemical Analysis
- -
- total N (Nitrogen), using Kieldahl’s method [26]
- -
- K, Ca—by flame photometry using Flapho 4, (Carl Zeiss Jena);
- -
- Mg, P—colorimetric method, using Spekol 10 (Carl Zeiss Jena).
2.5. Statistical Analysis
2.6. Analysis of the Impact of Environmental Conditions on Seasonal Growth
3. Results
3.1. Meteorological Data and Growing Season
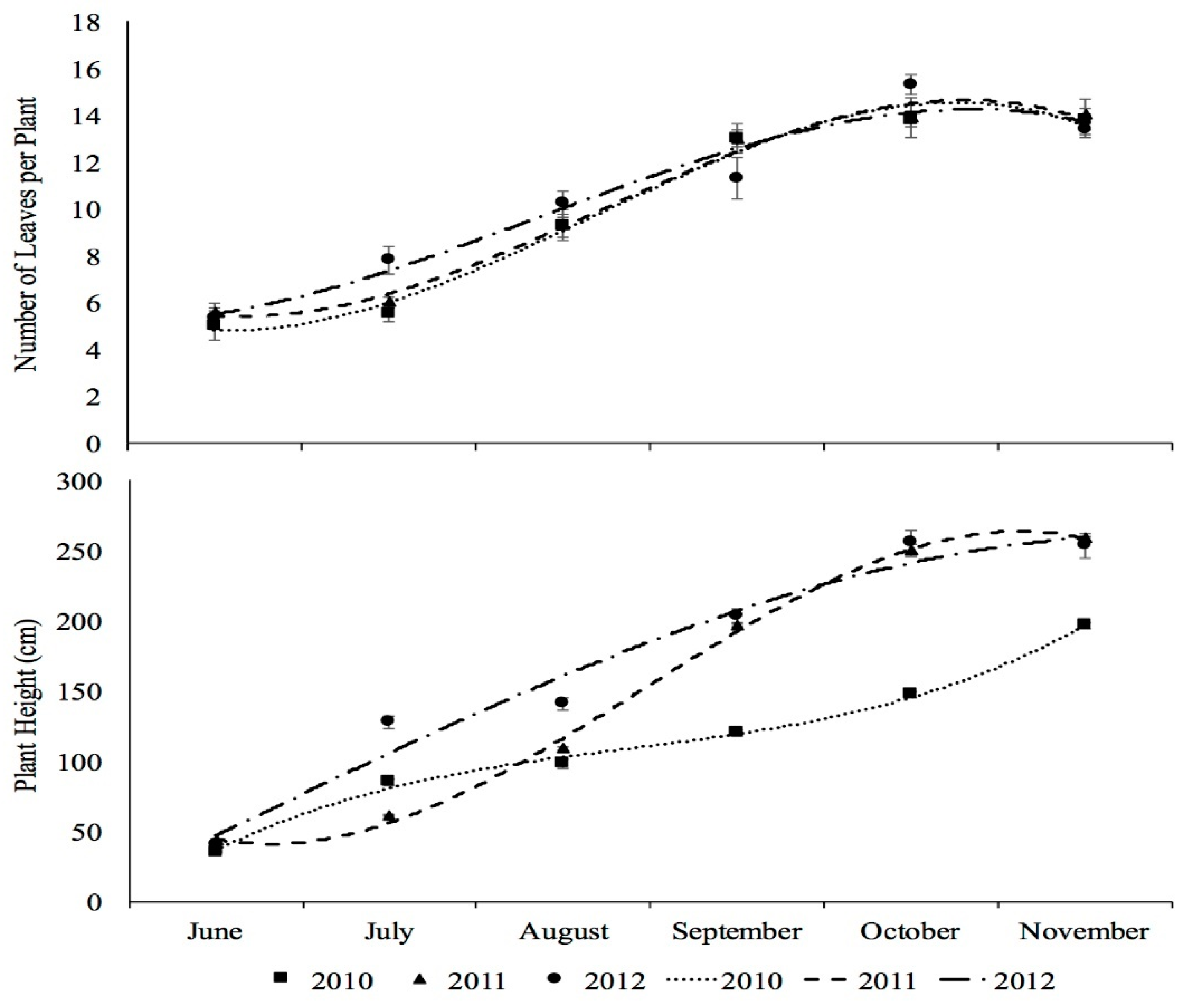
3.2. Plant Development
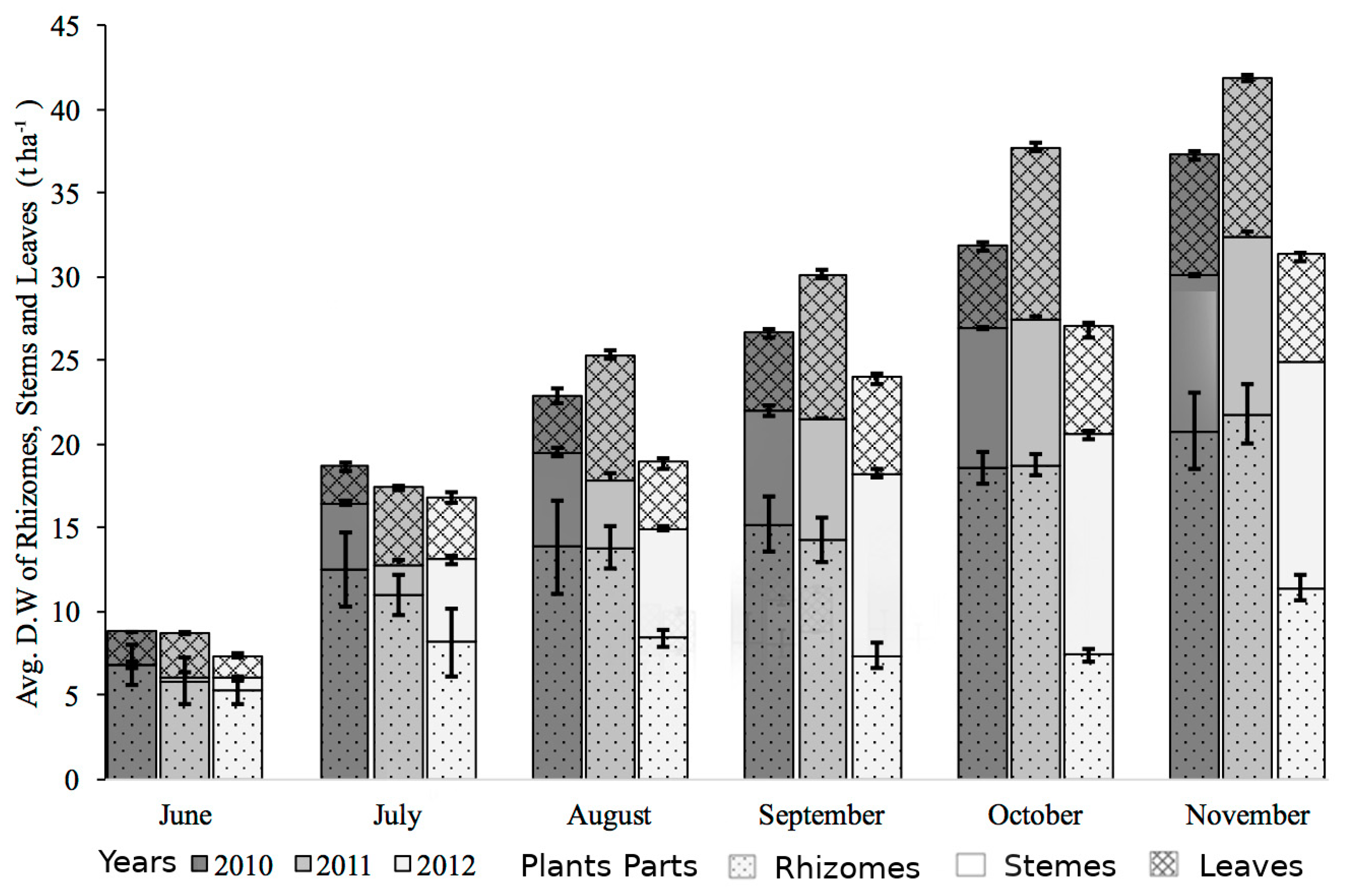
3.3. Biomass Accumulation
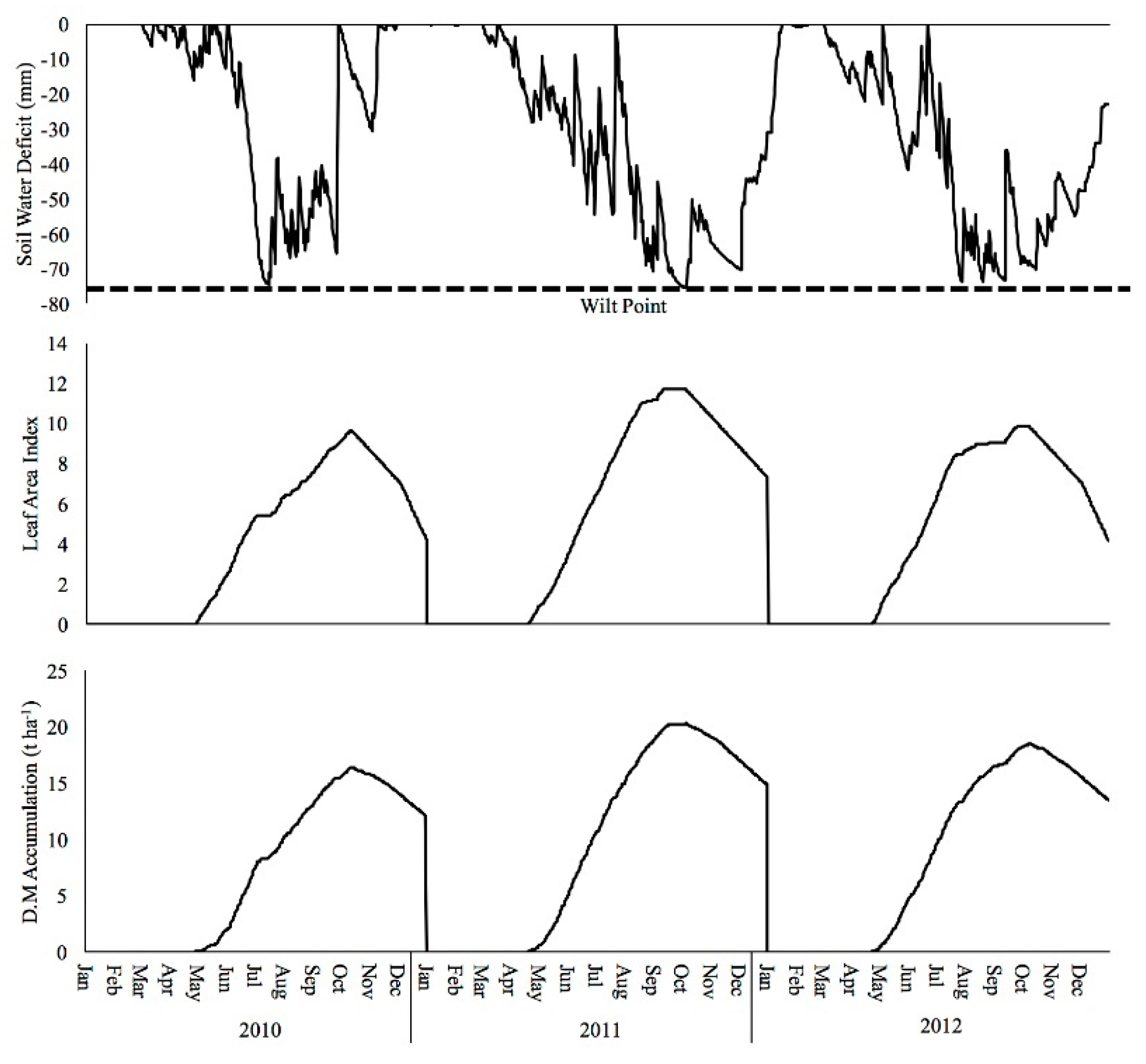
3.4. Soil Water, Drought Stress and Modelled Yields
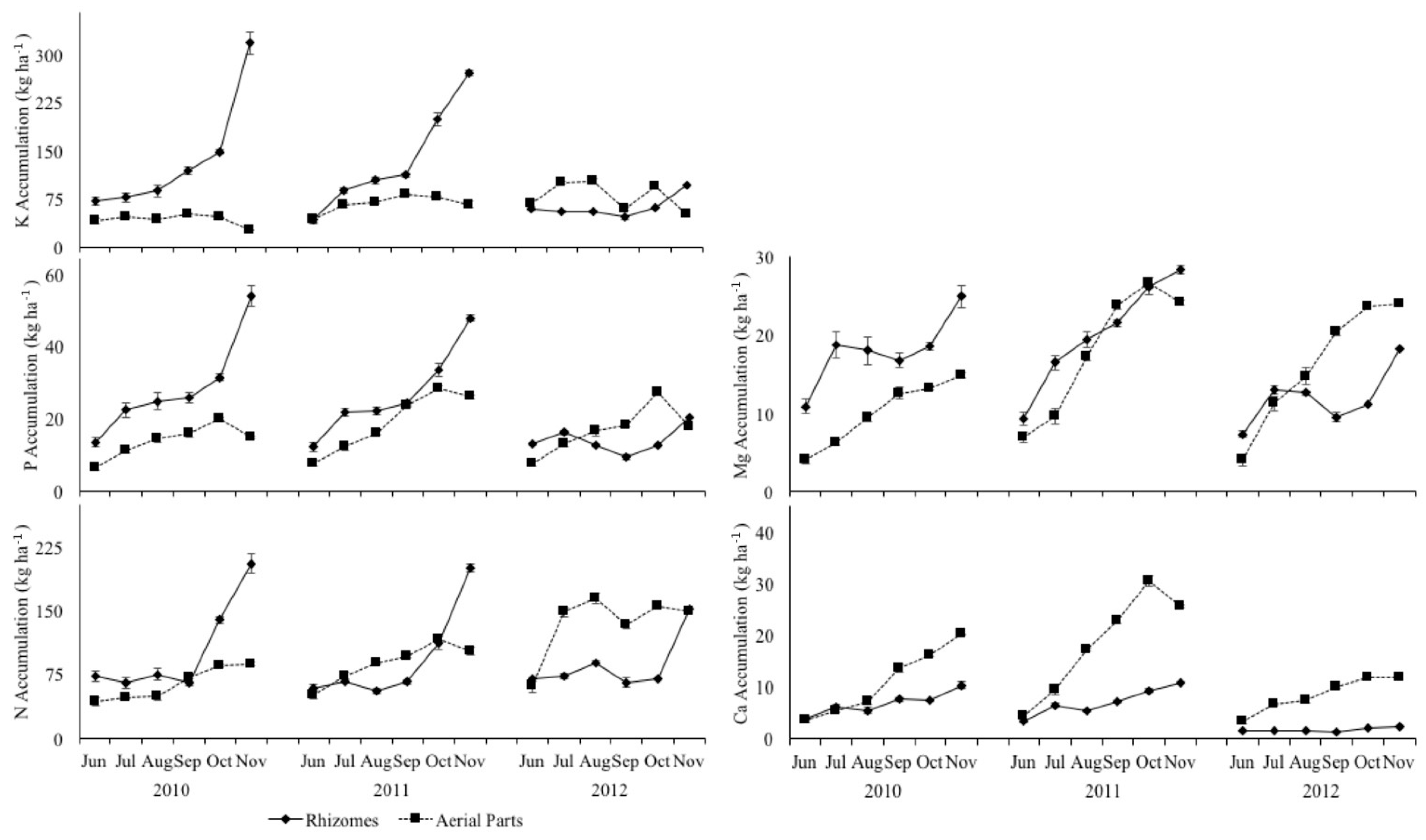
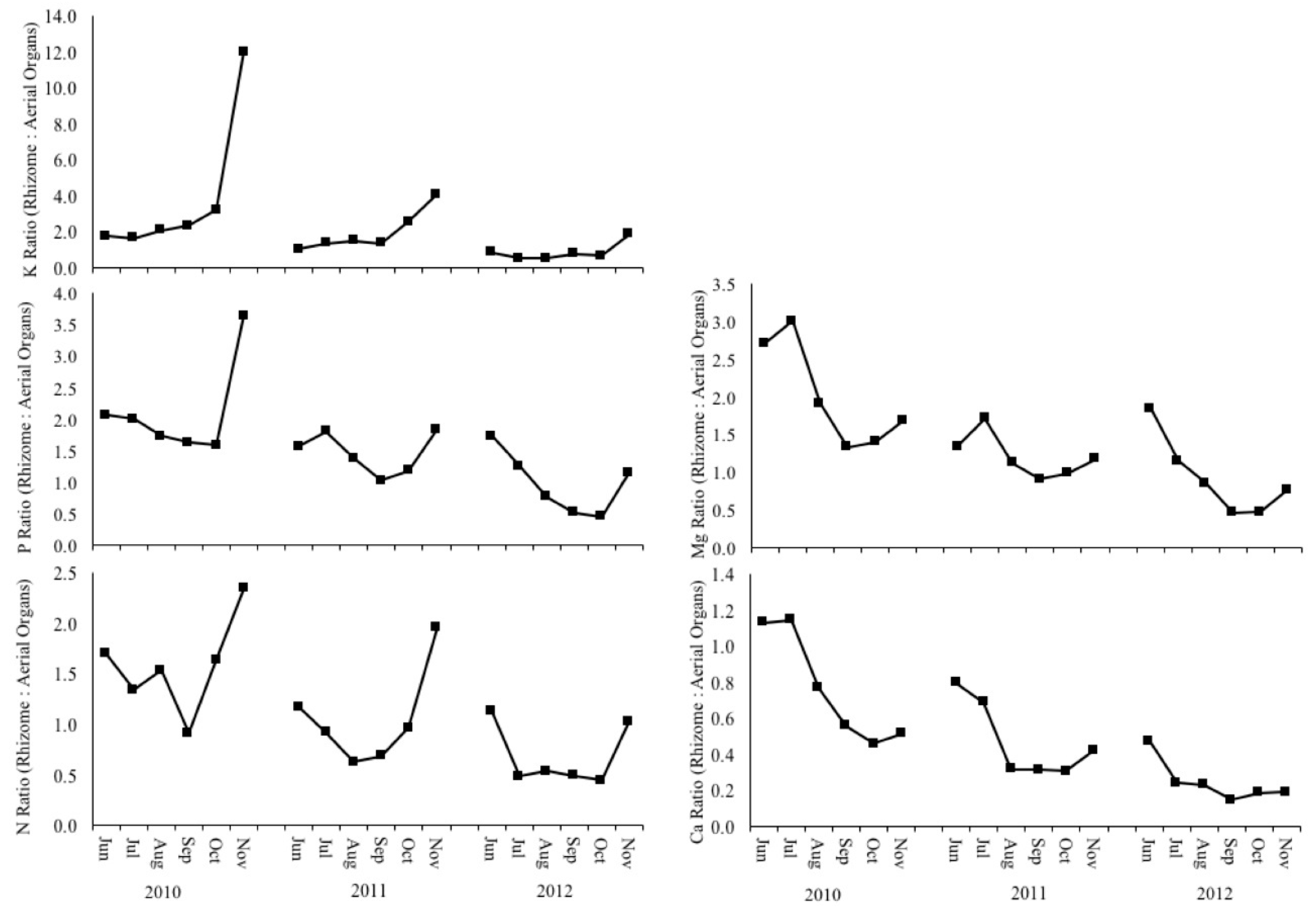
3.5. Biomass Quality and Macronutrient Management
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heaton, E.A.; Long, S.P.; Voigt, T.P.; Jones, M.B.; Clifton-Brown, J. Miscanthus for Renewable Energy Generation European Union Experience and Projections for Illinois. Rev. Environ. Sci. Bio. Tech. 2004, 9, 433–451. [Google Scholar] [CrossRef]
- Magenau, E.; Kiesel, A.; Clifton-Brown, J.; Lewandowski, I. Influence of cutting height on biomass yield and quality of miscanthus genotypes. GCB Bioenergy 2021, 8, 187. [Google Scholar] [CrossRef]
- Nsanganwimana, F.; Al Souki, K.S.; Waterlot, C.; Douay, F.; Pelfrêne, A.; Ridošková, A.; Louvel, B.; Pourrut, B. Potentials of Miscanthus × giganteus for phytostabilization of trace element-contaminated soils: Ex situ experiment. Ecotoxicol. Environ. Saf. 2021, 214, 112125. [Google Scholar] [CrossRef]
- Parrish, D.J.; Fike, J.H. The Biology and Agronomy of Switchgrass for Biofuels. Crit. Rev. Plant. Sci. 2005, 24, 423–459. [Google Scholar] [CrossRef]
- Kiesel, A.; Lewandowski, I. Miscanthus as biogas substrate—cutting tolerance and potential for anaerobic digestion. GCB Bioenergy 2017, 9, 153–167. [Google Scholar] [CrossRef]
- Dohleman, F.G.; Heaton, E.A.; Arundale, R.A.; Long, S.P. Seasonal dynamics of above- and below-ground biomass and nitrogen partitioning in Miscanthus × gigante us and Panicum virgatum across three growing seasons. GCB Bioenergy 2012, 4, 534–544. [Google Scholar] [CrossRef] [Green Version]
- Ouattara, M.S.; Laurent, A.; Ferchaud, F.; Berthou, M.; Borujerdi, E.; Butier, A.; Malvoisin, P.; Romelot, D.; Loyce, C. Evolution of soil carbon stocks under Miscanthus × giganteus and Miscanthus sinensis across contrasting environmental conditions. GCB Bioenergy 2021, 13, 161–174. [Google Scholar] [CrossRef]
- Beale, C.V.; Long, S.P. Seasonal Dynamics of Nutrient Accumulation and Partitioning in the Perrenial C4-Grasses Miscanthus × giganteus and Spartina Cynosurodies. Biomass Bioenergy 1997, 12, 419–428. [Google Scholar] [CrossRef]
- Christian, D.G.; Poulton, P.R.; Riche, A.B.; Yates, N.E.; Todd, A.D. The recovery over several seasons of 15 N-labelled fertilizer applied to Miscanthus × giganteus ranging from 1 to 3 years old. Biomass Bioenergy 2006, 30, 125–133. [Google Scholar] [CrossRef]
- Lewandowski, I.; Clifton-Brown, J.C.; Scurlock, J.M.O.; Huisman, W. Miscanthus: European experience with a novel energy crop. Biomass Bioenergy 2000, 19, 209–227. [Google Scholar] [CrossRef]
- Himken, M.; Lammel, J.; Neukirchen, D.; Czypionka-Krause, U.; Olfs, H.W. Cultivation of Miscanthus under West European conditions: Seasonal changes in dry matter production, nutrient uptake and remobilization. Plant Soil 1997, 189, 117–126. [Google Scholar] [CrossRef]
- Clifton-Brown, J.C.; Breuer, J.; Jones, M.B. Carbon mitigation by the energy crop, Miscanthus. Glob. Chang. Biol. 2007, 13, 2296–2307. [Google Scholar] [CrossRef]
- Borkowska, H.; Lipiński, W. Content of selected elements in biomass of several species of energy plants. Acta Agrophys. 2007, 10, 287–292. [Google Scholar]
- Jørgensen, U. Genotypic variation in dry matter accumulation and content of N, K and Cl in Miscanthus in Denmark. Biomass Bioenergy 1997, 12, 155–169. [Google Scholar] [CrossRef]
- Kahle, P.; Beuch, S.; Boelcke, B.; Leinweber, P.; Schulten, H.R. Cropping of Miscanthus in Central Europe: Biomass production and influence on nutrients and soil organic matter. Eur. J. Agron. 2001, 15, 171–184. [Google Scholar] [CrossRef]
- Wyżgolik, B.; Surowiec, E.; Brzakalik, K. Mineral balance in Miscanthus × giganteus. In Alternative Plants for Sustainable Agriculture; PAS: Poznań, Poland, 2006; pp. 115–120. [Google Scholar]
- Clifton-Brown, J.C.; Lewandowski, I. Screening Miscanthus genotypes in field trials to optimize biomass yield and quality in Southern Germany. Eur. J. Agron. 2002, 16, 97–110. [Google Scholar] [CrossRef]
- Atienza, S.G.; Satovic, Z.; Petersen, K.K.; Dolstra, O.; Martín, A. Identification of QTLs influencing combustion quality in Miscanthus sinensis Anderss. II. Chlorine and potassium content. Theor. Appl. Genet. 2003, 107, 857–863. [Google Scholar] [CrossRef]
- Kalembasa, D.; Malinowska, E. Zawartość potasu w różnych klonach trawy Miscanthus w zależności od nawożenia mineralnego. Nawozy i Nawożenie. Nawozy Nawożenie 2005, 3, 359–364. [Google Scholar]
- Kalembasa, D.; Malinowska, E.; Jaremko, D.; Jezowski, S. Zawartość potasu w różnych klonach trawy Miscanthus w zależności od nawożenia mineralnego. Zesz. Probl. Postęp. Nauk Rol. 2001, 480, 77–84. [Google Scholar]
- Buzar, M.; Galek, R.; Góra, J.; Grzyś, E.; Hurej, M.; Kotecki, A.; Kozak, M.; Piszcz, U.; Pląskowska, E.; Pusz, W.; et al. Uprawa Miskanta Olbrzymiego: Energetyczne i Pozaenergetyczne Możliwości Wykorzystania Słomy; Kotecki, A., Ed.; Wydawnictwo Uniwersytetu Przyrodniczego We Wrocławiu: Wrocław, Poland, 2010; p. 186. (In Polish) [Google Scholar]
- Clifton-Brown, J.; Hastings, A.; Mos, M.; McCalmont, J.P.; Ashman, C.; Awty-Carroll, D.; Cerazy, J.; Chiang, Y.-C.; Cosentino, S.; Cracroft-Eley, W.; et al. Progress in upscaling Miscanthus biomass production for the European bio-economy with seed-based hybrids. GCB Bioenergy 2017, 9, 6–17. [Google Scholar] [CrossRef] [Green Version]
- De Wit, M.; Faaij, A. European biomass resource potential and costs. Biomass Bioenergy 2010, 34, 188–202. [Google Scholar] [CrossRef]
- Hastings, A.; Clifton-Brown, J.; Wattenbach, M.; Stampl, P.; Mitchell, C.P.; Smith, P. Potential of Miscanthus grasses to provide energy and hence reduce greenhouse gas emissions. Agron. Sustain. Dev. 2008, 28, 465–472. [Google Scholar] [CrossRef] [Green Version]
- IUNG-PIB. Available online: http://www.iung.pulawy.pl/ (accessed on 13 September 2020).
- Kjeldahl, J. Neue Methode zur Bestimmung des Stickstoffs in organischen Körpern. Z. Für Anal. Chemie. 1883, 22, 366–382. [Google Scholar] [CrossRef] [Green Version]
- Campbell, G.S. Soil physics with Basic: Transport models for soil-plant systems. Dev. Soil Sci. 1985, 14, 6–134. [Google Scholar]
- Hastings, A.; Clifton-Brown, J.; Wattenbach, M.; Mitchell, C.P.; Smith, P. The development of MISCANFOR, a new Miscanthus crop growth model: Towards more robust yield predictions under different climatic and soil conditions. GCB Bioenergy 2009, 1, 154–170. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspration-Guidelines for computing crop water requirements—FAO Irrigation and drainage paper 56. FAO Rome 1998, 300, D05109. [Google Scholar]
- Christian, D.G.; Riche, A.B.; Yates, N.E. Growth, yield and mineral content of Miscanthus × giganteus grown as a biofuel for 14 successive harvests. Ind. Crops Prod. 2008, 28, 320–327. [Google Scholar] [CrossRef]
- Lewandowski, I. Propagation method as an important factor in the growth and development of Miscanthus × giganteus. Ind. Crops Prod. 1998, 8, 229–245. [Google Scholar] [CrossRef]
- Yazaki, Y.; Mariko, S.; Koizumi, H. Carbon dynamics and budget in a Miscanthus sinensis grassland in Japan. Ecol. Res. 2004, 19, 511–520. [Google Scholar] [CrossRef]
- Voća, N.; Leto, J.; Karažija, T.; Bilandžija, N.; Peter, A.; Kutnjak, H.; Šurić, J.; Poljak, M. Energy Properties and Biomass Yield of Miscanthus × giganteus Fertilized by Municipal Sewage Sludge. Molecules 2021, 26, 4371. [Google Scholar] [CrossRef] [PubMed]
- Baxter, X.C.; Darvell, L.I.; Jones, J.M.; Barraclough, T.; Yates, N.E.; Shield, I. Miscanthus combustion properties and variations with Miscanthus agronomy. Fuel 2014, 117, 851–869. [Google Scholar] [CrossRef] [Green Version]
- Kalembasa, D.; Janinhoff, A.; Malinowska, E.; Jaremko, D.; Jezowski, S. Zawartość siarki w wybranych klonach trawy Miscanthus. J. Elem. 2005, 10, 309–314. [Google Scholar]
- Richter, G.M.; Riche, A.B.; Dailey, A.G.; Gezan, S.A.; Powlson, D.S. Is UK biofuel supply from Miscanthus water-limited? Soil Use Manag. 2008, 24, 235–245. [Google Scholar] [CrossRef]
- Dohleman, F.G.; Long, S.P. More productive than maize in the Midwest: How does Miscanthus do it? Plant Physiol. 2009, 150, 2104–2115. [Google Scholar] [CrossRef] [Green Version]
- Peixoto, M.d.M.; Friesen, P.C.; Sage, R.F. Winter cold-tolerance thresholds in field-grown Miscanthus hybrid rhizomes. J. Exp. Bot. 2015, 66, 4415–4425. [Google Scholar] [CrossRef] [Green Version]
- Collison, R.F.; Raven, E.C.; Pignon, C.P.; Long, S.P. Light, Not Age, Underlies the Maladaptation of Maize and Miscanthus Photosynthesis to Self-Shading. Front. Plant Sci. 2020, 11, 783. [Google Scholar] [CrossRef]
- Strullu, L.; Cadoux, S.; Preudhomme, M.; Jeuffroy, M.H.; Beaudoin, N. Biomass production and nitrogen accumulation and remobilization by Miscanthus × giganteus as influenced by nitrogen stocks in belowground organs. Field Crops Res. 2011, 121, 381–391. [Google Scholar] [CrossRef]
- Cadoux, S.; Riche, A.B.; Yates, N.E. Machet, J.-M. Nutrient requirements of Miscanthus × giganteus: Conclusions from a review of published studies. Biomass Bioenergy 2012, 38, 14–22. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Average Fresh Weight of Rhizomes from 0.25 m2 (g) | Water Content of Rhizomes (g kg−1) | Average Number of Stems Per Plant | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 2010 | 2011 | 2012 | 2010 | 2011 | 2012 | 2010 | 2011 | 2012 | |
| June | 692.5 ± 55.8 | 522.5 ± 61.2 | 603.5 ± 42.3 | 755.8 ± 2.5 | 722.5 ± 2.8 | 748.2 ± 2.1 | 15.8 ± 1.7 | 11.3 ± 1.5 | 10.5 ± 1.0 |
| July | 976.0 ± 83.5 | 964.3 ± 60.0 | 701.0 ± 81.7 | 681.7 ± 2.2 | 715.0 ± 2.2 | 710.8 ± 4.2 | 19.0 ± 0.8 | 12.5 ± 1.0 | 11.3 ± 0.9 |
| August | 950.5 ± 105.3 | 990.0 ± 40.7 | 801.5 ± 21.2 | 634.2 ± 6.3 | 651.7 ± 2.9 | 738.3 ± 2.9 | 16.0 ± 1.0 | 12.3 ± 0.5 | 9.8 ± 0.5 |
| September | 951.0 ± 37.8 | 955.3 ± 38.9 | 556.0 ± 25.1 | 600.8 ± 8.2 | 625.8 ± 2.1 | 671.7 ± 2.9 | 15.8 ± 0.9 | 14.5 ± 0.6 | 9.3 ± 0.5 |
| October | 1129.0 ± 40.0 | 1377.3 ± 41.0 | 659.8 ± 25.9 | 588.3 ± 4.0 | 660.0 ± 7.1 | 720.0 ± 4.1 | 16.0 ± 0.0 | 15.3 ± 0.6 | 9.8 ± 0.5 |
| November | 1904.8 ± 112.2 | 1741.8 ± 59.7 | 821.3 ± 34.3 | 727.5 ± 1.6 | 687.5 ± 4.8 | 652.5 ± 2.8 | 16.8 ± 0.5 | 14.8 ± 0.5 | 9.8 ± 0.3 |
| 2010 | 2011 | 2012 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | P | K | Ca | Mg | N | P | K | Ca | Mg | N | P | K | Ca | Mg | |
| R | 0.89 | 0.80 | 0.26 | 0.86 | 0.91 | 0.86 | 0.88 | 0.89 | 0.90 | 0.82 | 0.59 | 0.40 | −0.33 | 0.00 | 0.66 |
| ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mos, M.; Robson, P.R.H.; Buckby, S.; Hastings, A.F.; Helios, W.; Jama-Rodzeńska, A.; Kotecki, A.; Kalembasa, D.; Kalembasa, S.; Kozak, M.; et al. Seasonal Dynamics of Dry Matter Accumulation and Nutrients in a Mature Miscanthus × giganteus Stand in the Lower Silesia Region of Poland. Agronomy 2021, 11, 1679. https://doi.org/10.3390/agronomy11081679
Mos M, Robson PRH, Buckby S, Hastings AF, Helios W, Jama-Rodzeńska A, Kotecki A, Kalembasa D, Kalembasa S, Kozak M, et al. Seasonal Dynamics of Dry Matter Accumulation and Nutrients in a Mature Miscanthus × giganteus Stand in the Lower Silesia Region of Poland. Agronomy. 2021; 11(8):1679. https://doi.org/10.3390/agronomy11081679
Chicago/Turabian StyleMos, Michal, Paul R. H. Robson, Sam Buckby, Astley F. Hastings, Waldemar Helios, Anna Jama-Rodzeńska, Andrzej Kotecki, Dorota Kalembasa, Stanisław Kalembasa, Marcin Kozak, and et al. 2021. "Seasonal Dynamics of Dry Matter Accumulation and Nutrients in a Mature Miscanthus × giganteus Stand in the Lower Silesia Region of Poland" Agronomy 11, no. 8: 1679. https://doi.org/10.3390/agronomy11081679
APA StyleMos, M., Robson, P. R. H., Buckby, S., Hastings, A. F., Helios, W., Jama-Rodzeńska, A., Kotecki, A., Kalembasa, D., Kalembasa, S., Kozak, M., Chmura, K., Serafin-Andrzejewska, M., & Clifton-Brown, J. (2021). Seasonal Dynamics of Dry Matter Accumulation and Nutrients in a Mature Miscanthus × giganteus Stand in the Lower Silesia Region of Poland. Agronomy, 11(8), 1679. https://doi.org/10.3390/agronomy11081679