
Silicon Modulates Molecular and Physiological Activities in Lsi1 Transgenic and Wild Lemont Rice Seedlings under Arsenic Stress


Abstract
:1. Introduction
2. Materials and Methods
2.1. Planting Conditions
2.2. As Concentration Measurement
2.3. Si Concentration and Dry Weight Measurement
2.4. Soluble Protein Contents Measurement
2.5. Transcriptomic Analyses
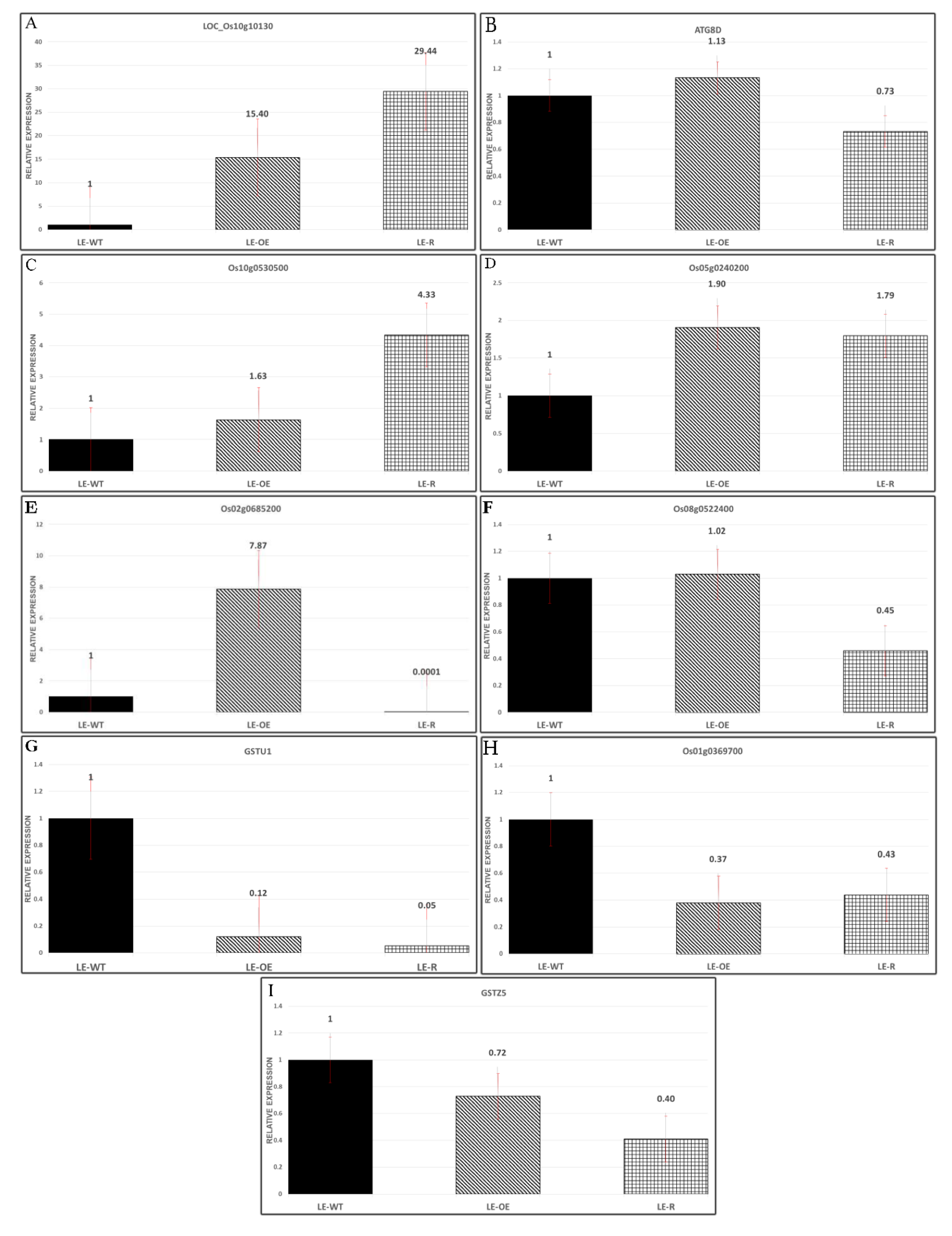
2.6. Quantitative RT-PCR
2.7. Statistical Analyses
3. Results
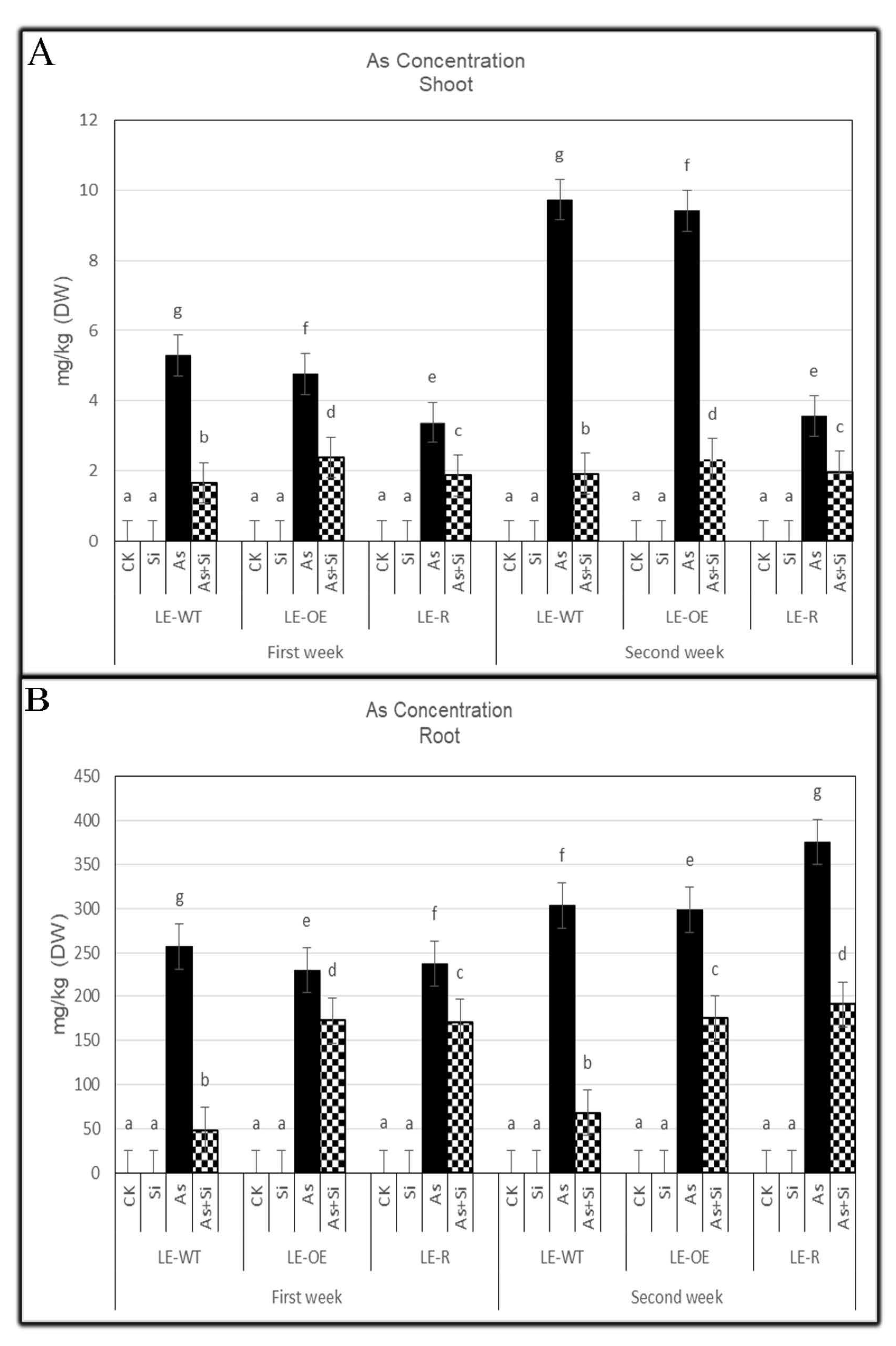
3.1. The Role of Si and As Treatments on As Concentration of Rice Shoots and Roots
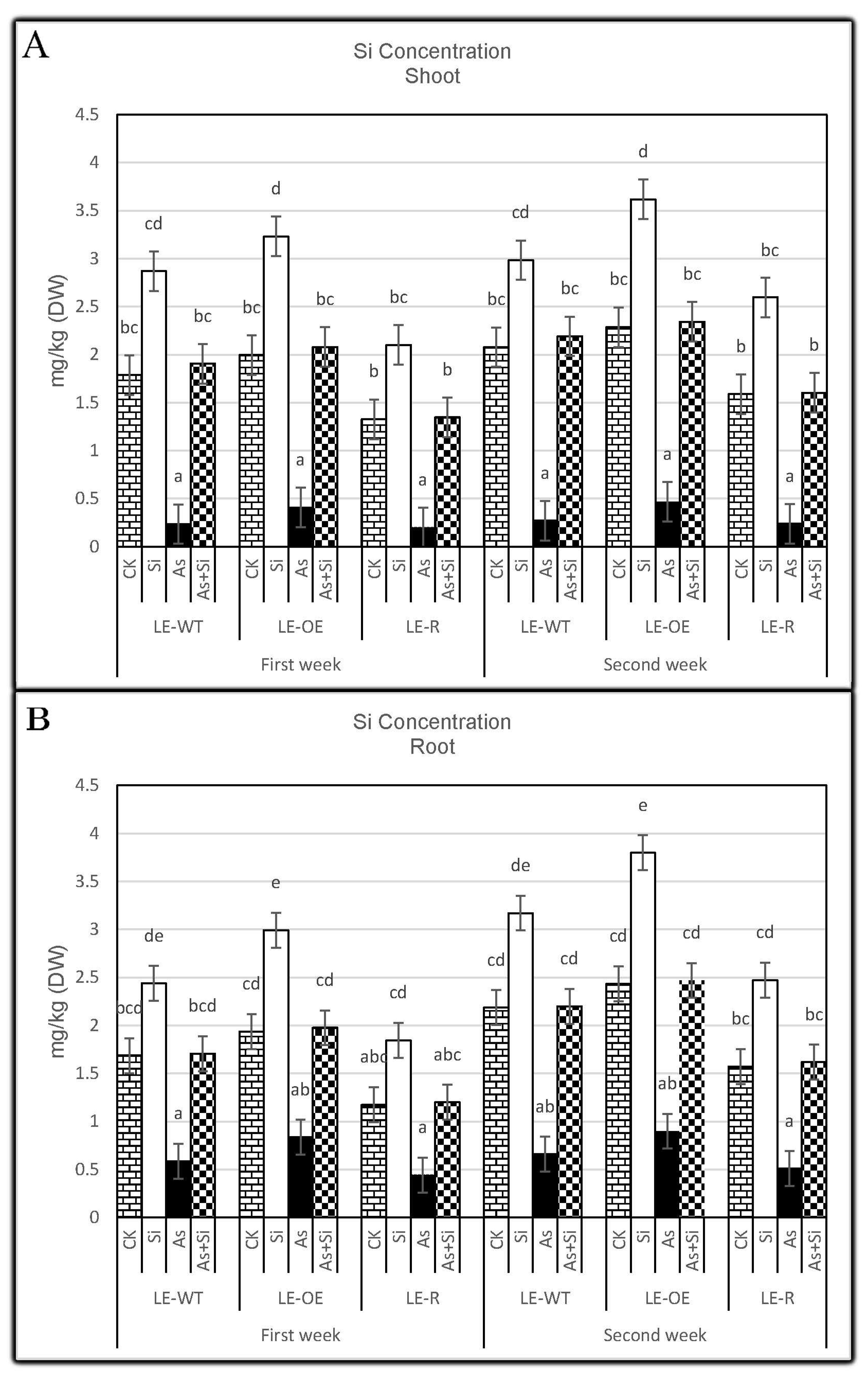
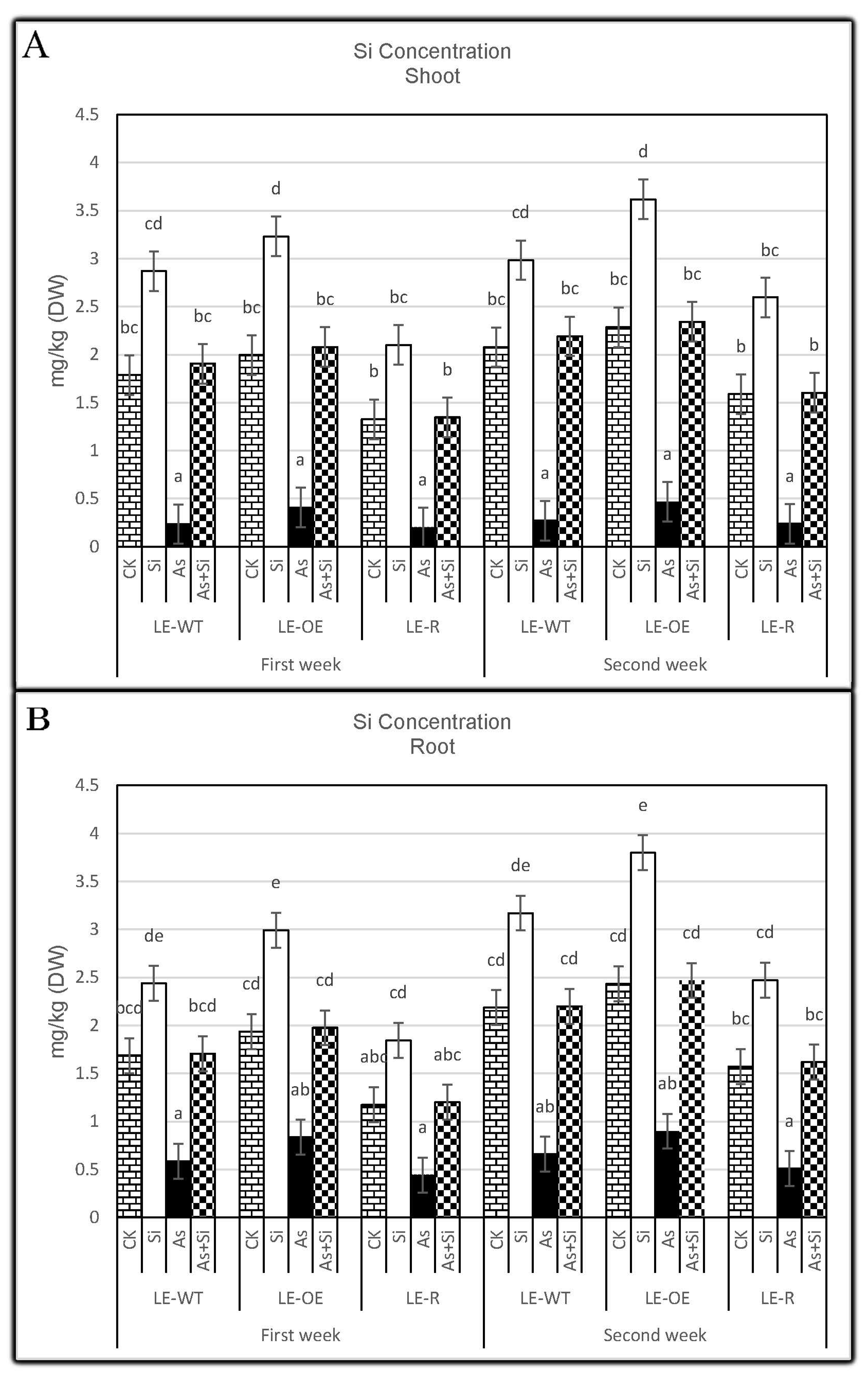
3.2. The Role of Si and As Treatments on Si Concentration of Rice Shoots and Roots
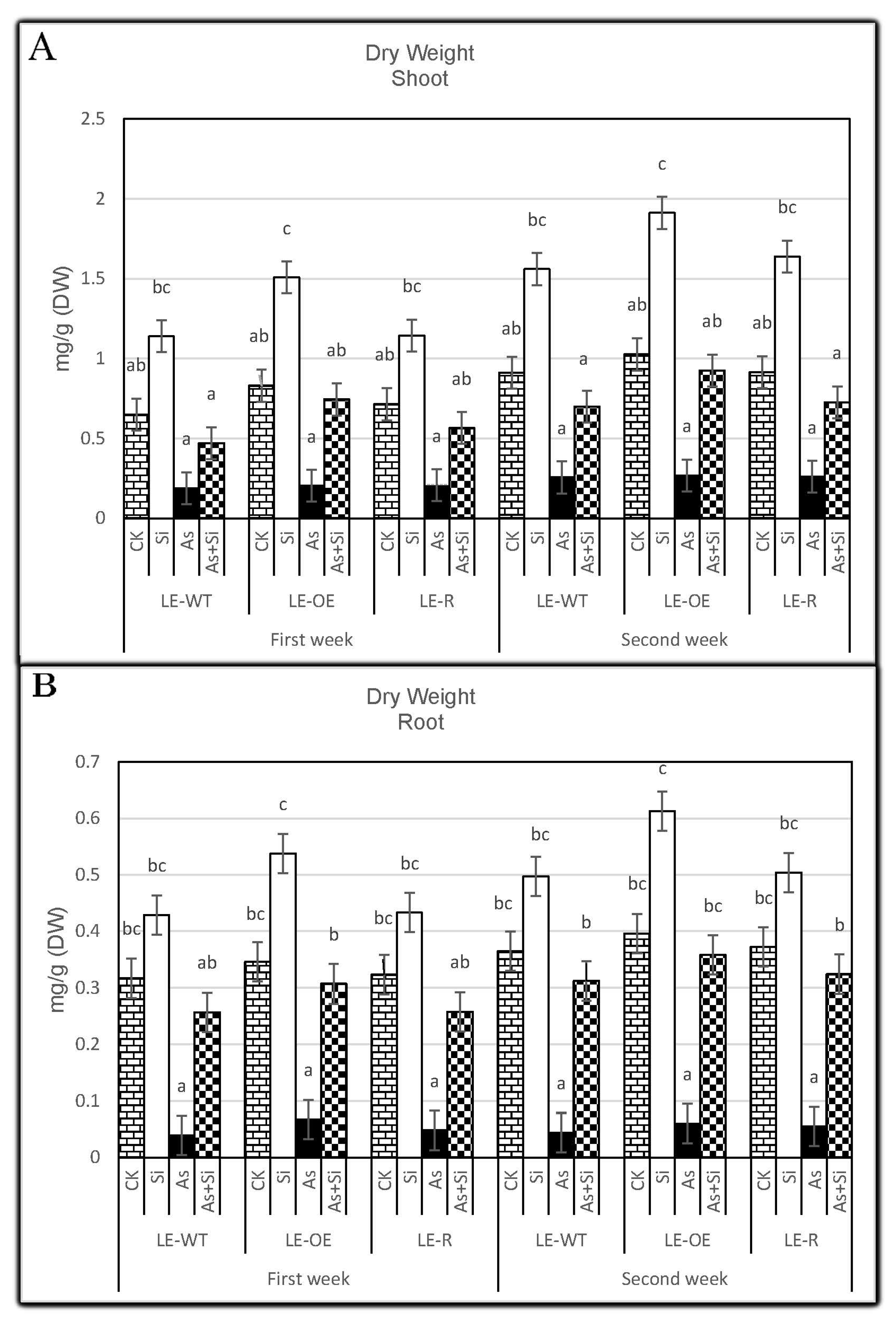
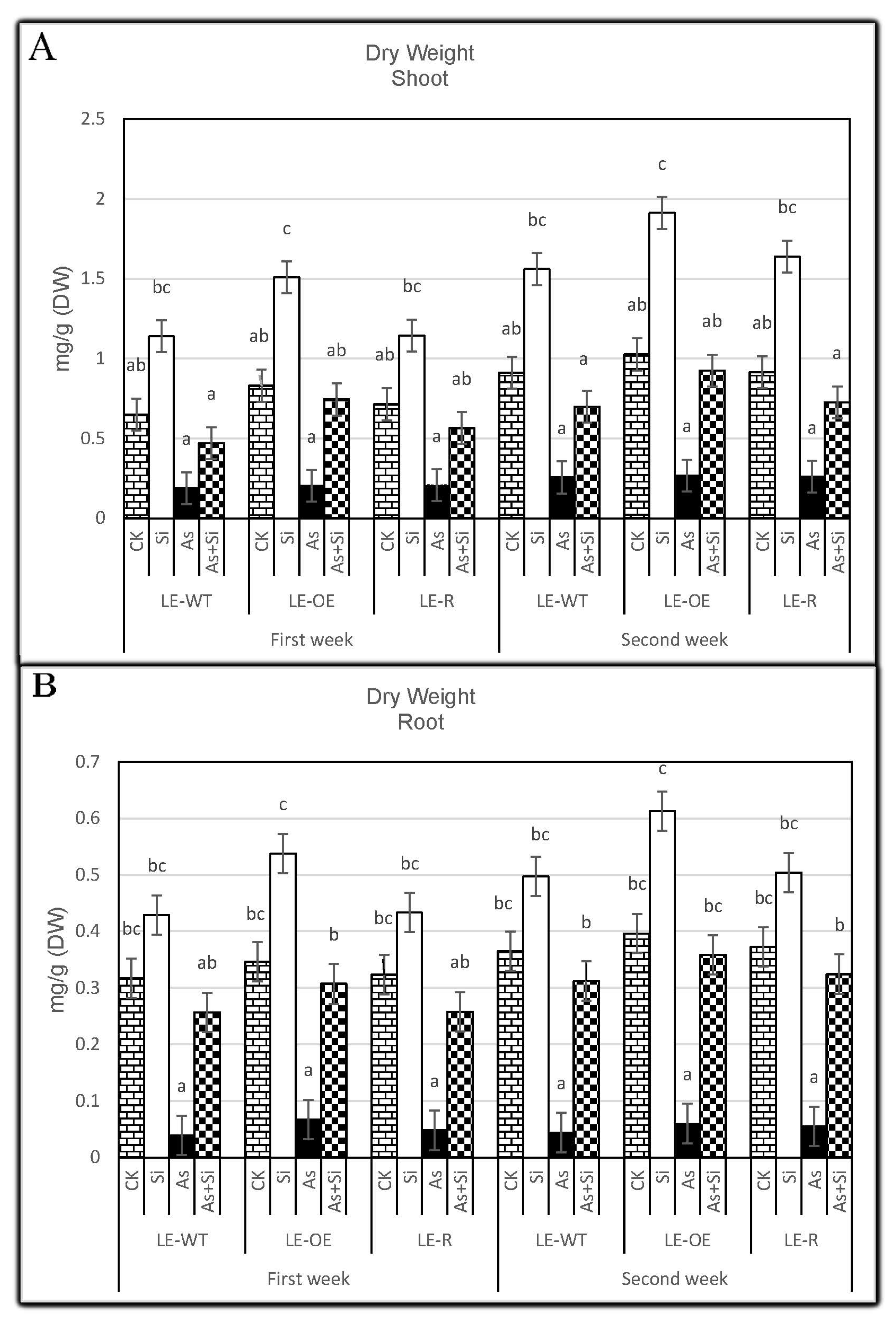
3.3. The Role of Si and As Treatments on Dry Weight of Rice Shoots and Roots
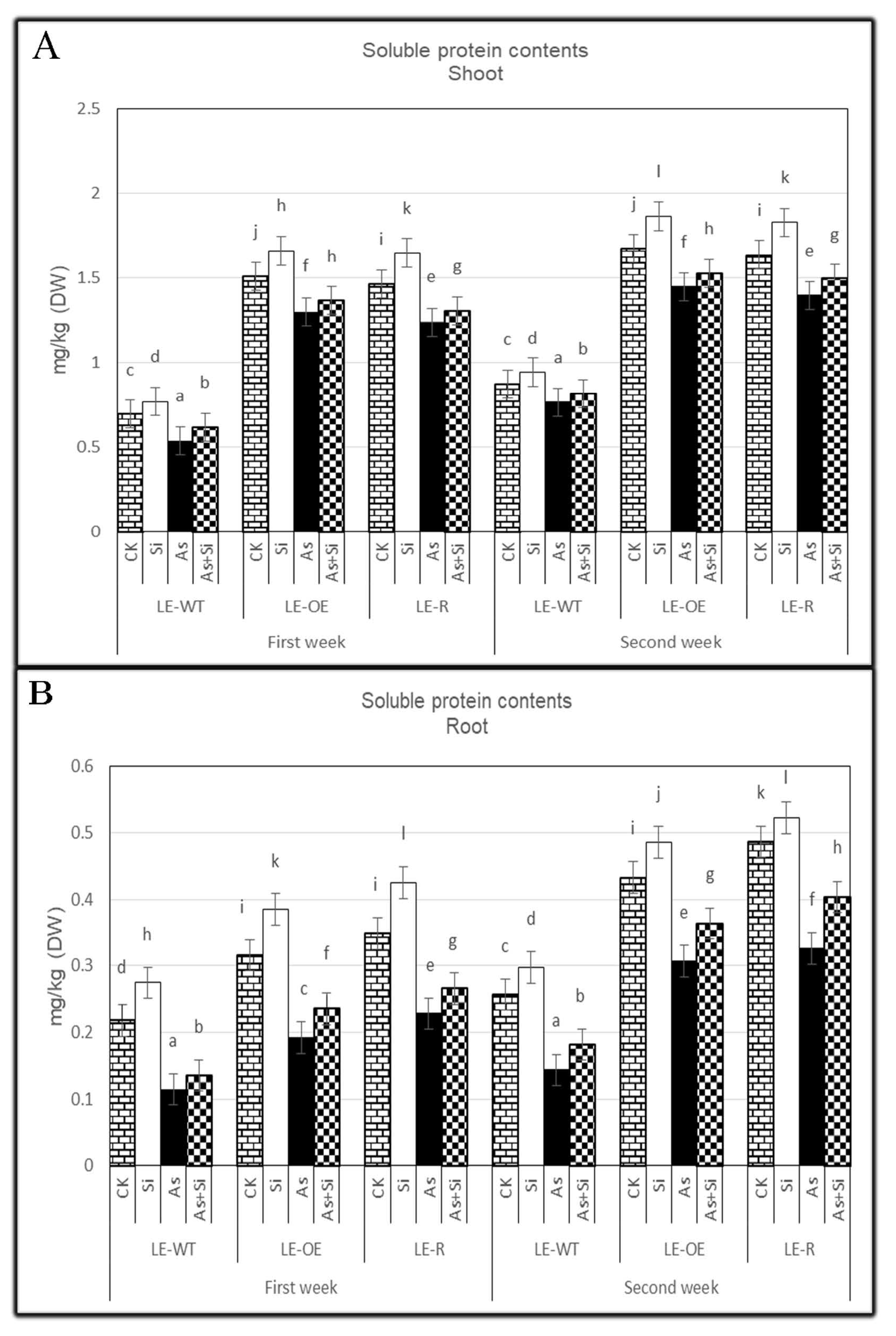
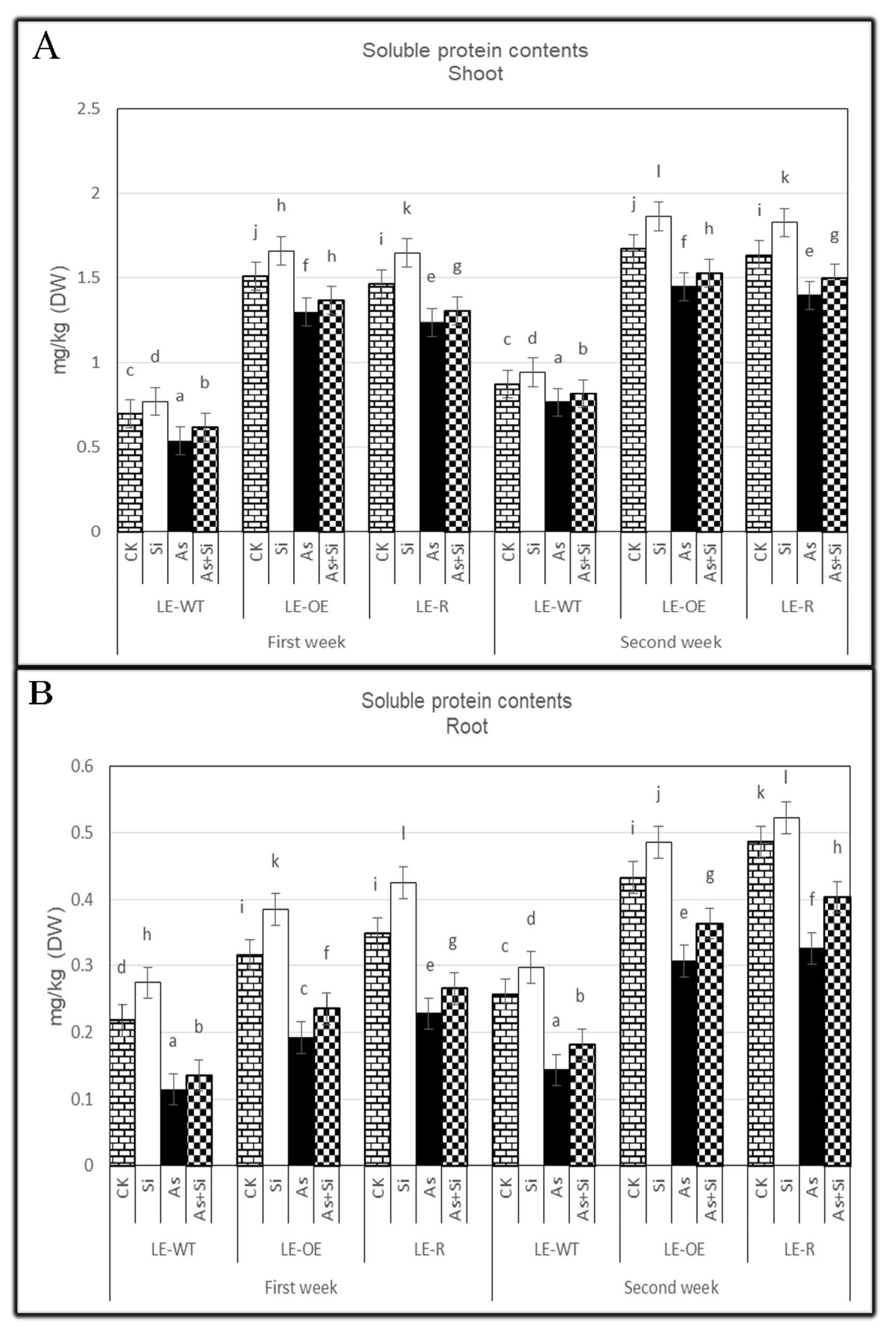
3.4. The Role of Si and As Treatments on Soluble Protein Contents of Rice Shoots and Roots
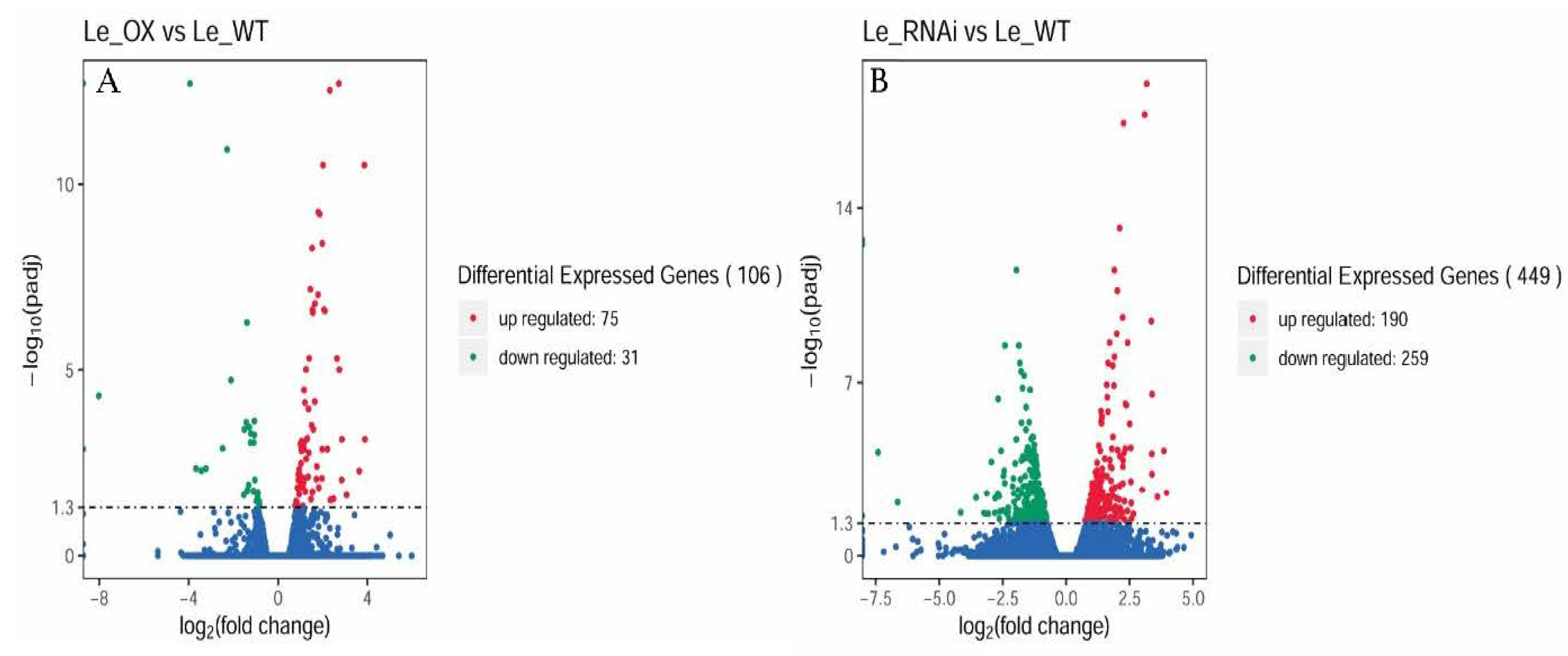
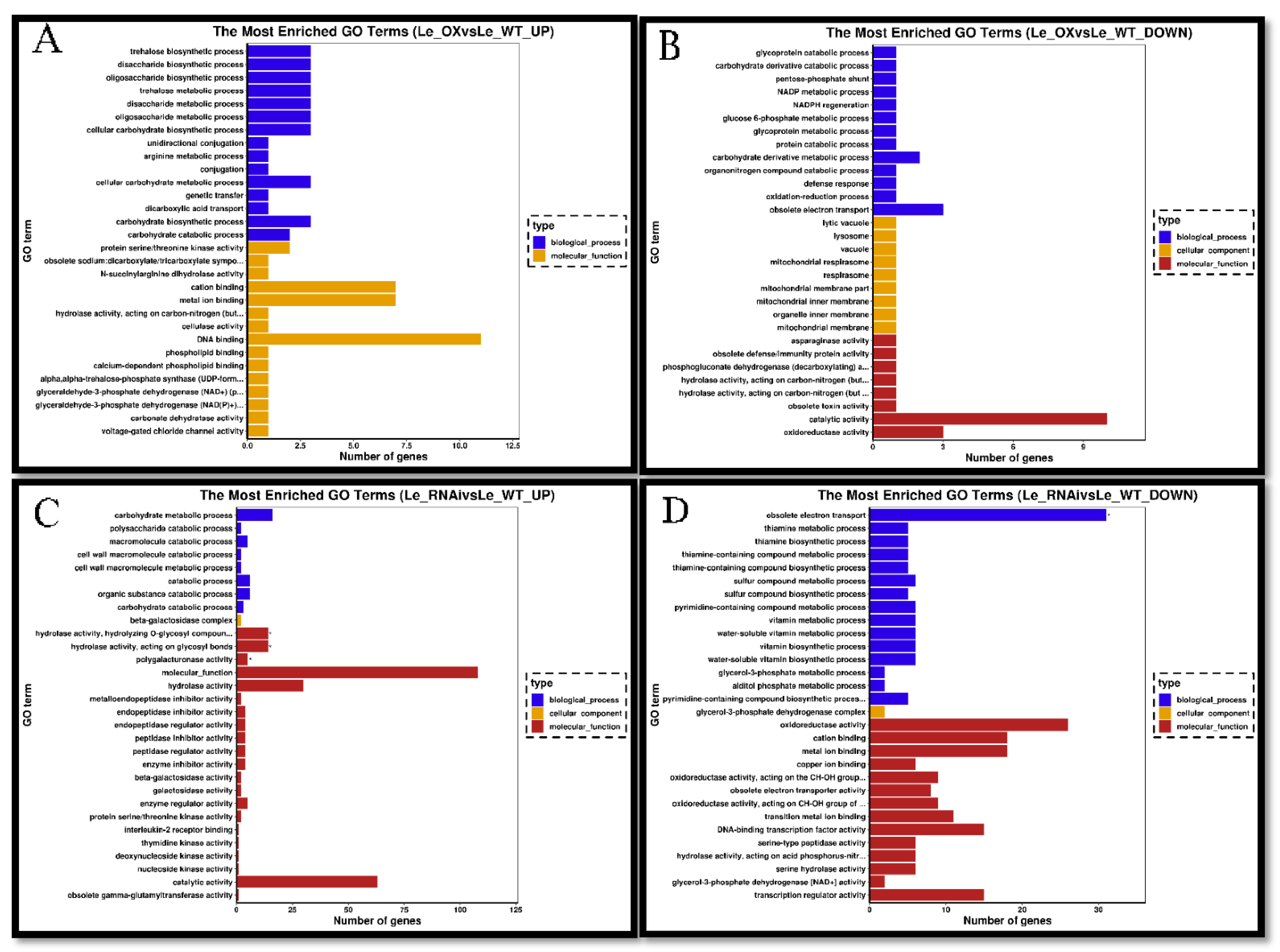
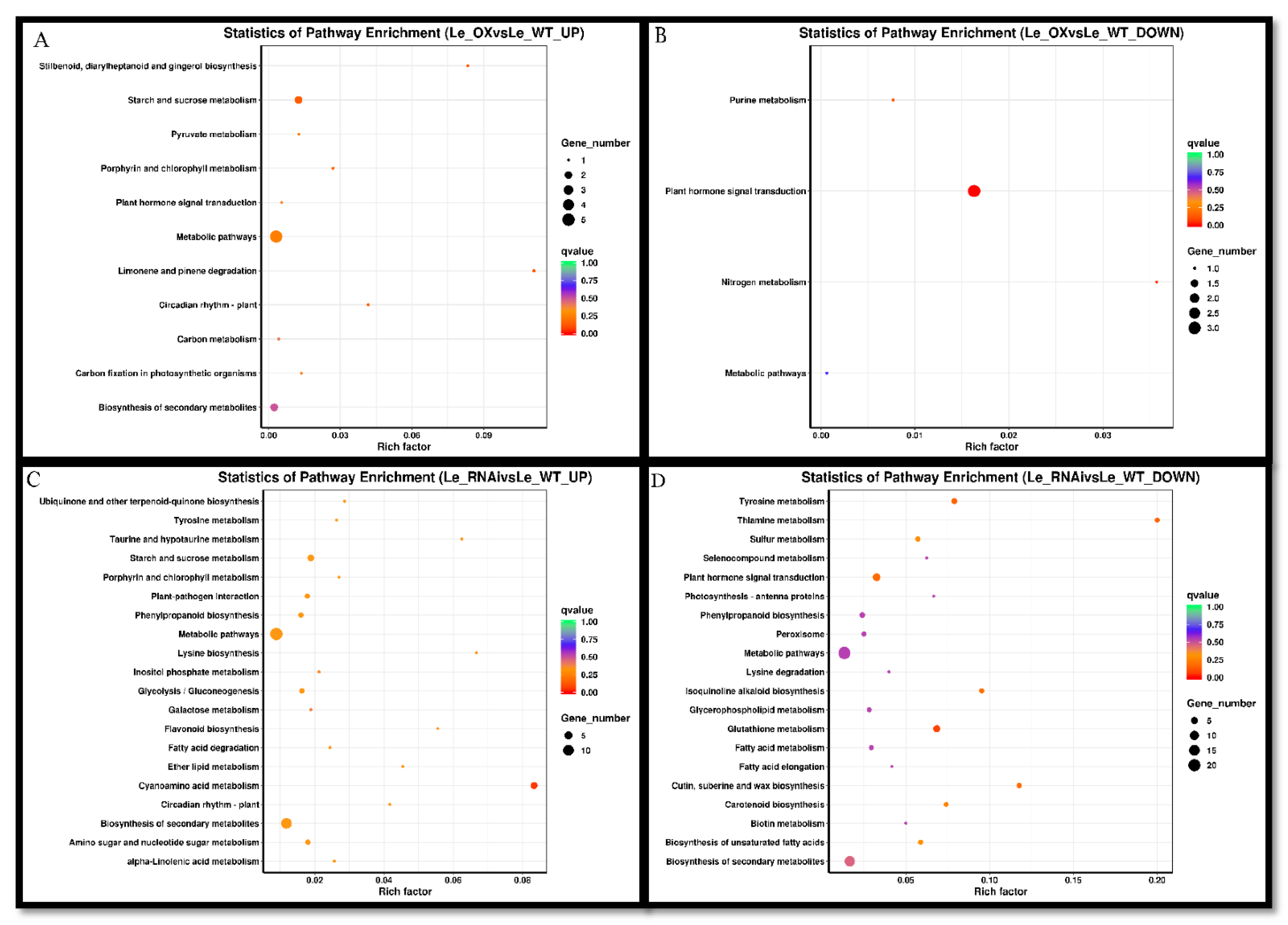
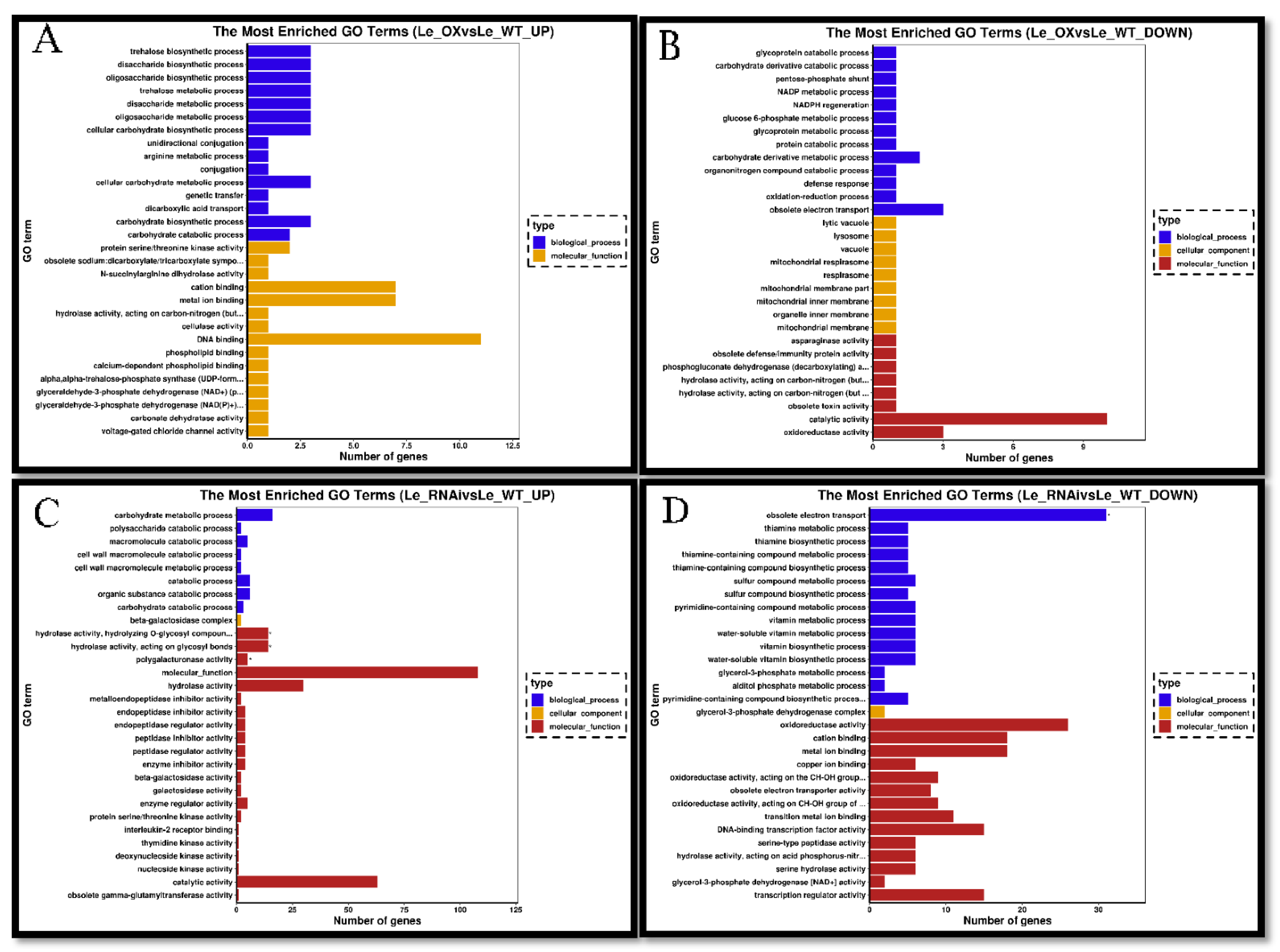
3.5. Transcriptomic Analyses
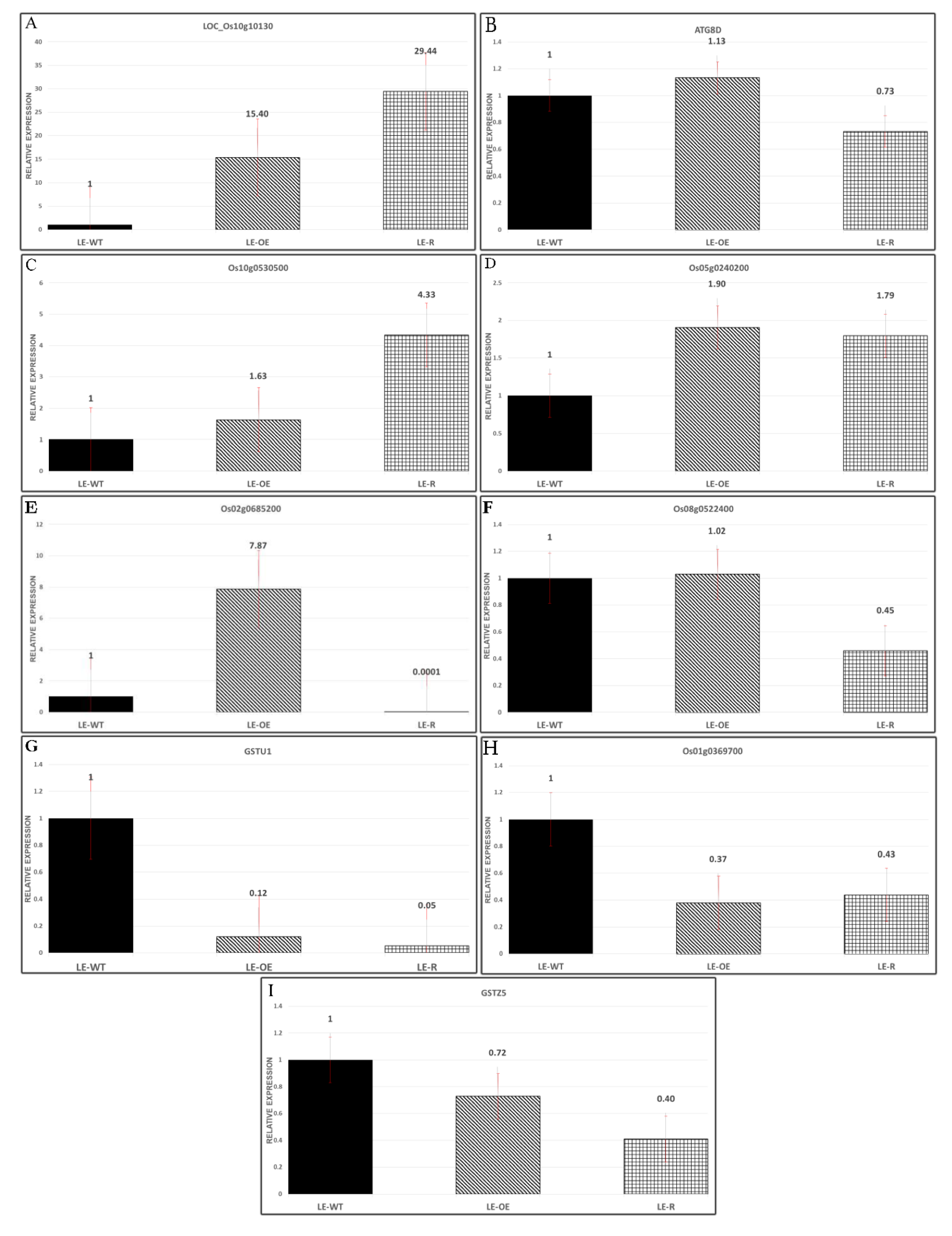
3.6. Expression of Some Genes in Different Varieties of Rice Exposed to Si and As Treatments
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhattacharya, S.; Gupta, K.; Debnath, S.; Ghosh, U.C.; Chattopadhyay, D.; Mukhopadhyay, A. Arsenic bioaccumulation in rice and edible plants and subsequent transmission through food chain in Bengal basin: A review of the perspectives for environmental health. Toxicol. Environ. Chem. 2012, 94, 429–441. [Google Scholar] [CrossRef]
- Bhattacharya, P.; Samal, A.C.; Majumdar, J.; Banerjee, S.; Santra, S.C. In Vitro assessment on the impact of soil arsenic in the eight rice varieties of West Bengal, India. J. Hazard. Mater. 2013, 262, 1091–1097. [Google Scholar] [CrossRef]
- Bhattacharya, P.; Samal, A.C.; Majumdar, J.; Santra, S.C. Uptake of arsenic in rice plant varieties cultivated with arsenic rich groundwater. Environ. Asia 2010, 3, 34–37. [Google Scholar]
- Chen, C.-L.; Chiou, H.-Y.; Hsu, L.-I.; Hsueh, Y.-M.; Wu, M.-M.; Chen, C.-J. Ingested arsenic, characteristics of well water consumption and risk of different histological types of lung cancer in northeastern Taiwan. Environ. Res. 2010, 110, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Sanaei, G. Industrial Toxicology; Tehran University Press: Tehran, Iran, 2004; Volume I. [Google Scholar]
- Richmond, K.E.; Sussman, M. Got silicon? The non-essential beneficial plant nutrient. Curr. Opin. Plant Biol. 2003, 6, 268–272. [Google Scholar] [CrossRef]
- Song, Z.; Wang, H.; Strong, P.J.; Shan, S. Increase of available soil silicon by Si-rich manure for sustainable rice production. Agron. Sustain. Dev. 2014, 34, 813–819. [Google Scholar] [CrossRef] [Green Version]
- Cuong, T.X.; Ullah, H.; Datta, A.; Hanh, T.C. Effects of silicon-based fertilizer on growth, yield and nutrient uptake of rice in tropical zone of Vietnam. Rice Sci. 2017, 24, 283–290. [Google Scholar] [CrossRef]
- He, C.; Ma, J.; Wang, L. A hemicellulose-bound form of silicon with potential to improve the mechanical properties and regeneration of the cell wall of rice. New Phytol. 2015, 206, 1051–1062. [Google Scholar] [CrossRef] [PubMed]
- You-Qiang, F.; Hong, S.; Dao-Ming, W.; Kun-Zheng, C. Silicon-mediated amelioration of Fe2+ toxicity in rice (Oryza sativa L.) roots. Pedosphere 2012, 22, 795–802. [Google Scholar]
- Hoogenkamp, H.; Kumagai, H.; Wanasundara, J. Rice protein and rice protein products. In Sustainable Protein Sources; Elsevier: Amsterdam, The Netherlands, 2017; pp. 47–65. [Google Scholar]
- Kim, Y.-H.; Khan, A.L.; Shinwari, Z.K.; Kim, D.-H.; Waqas, M.; Kamran, M.; Lee, I.-J. Silicon treatment to rice (Oryza sativa L. cv.‘Gopumbyeo’) plants during different growth periods and its effects on growth and grain yield. Pak. J. Bot. 2012, 44, 891–897. [Google Scholar]
- Seyfferth, A.L.; Fendorf, S. Silicate mineral impacts on the uptake and storage of arsenic and plant nutrients in rice (Oryza sativa L.). Environ. Sci. Technol. 2012, 46, 13176–13183. [Google Scholar] [CrossRef] [PubMed]
- Fleck, A.T.; Mattusch, J.; Schenk, M.K. Silicon decreases the arsenic level in rice grain by limiting arsenite transport. J. Plant. Nutrition. 2013, 176, 785–794. [Google Scholar] [CrossRef]
- Ali, W.; Isayenkov, S.V.; Zhao, F.-J.; Maathuis, F.J. Arsenite transport in plants. Cell. Mol. Life Sci. 2009, 66, 2329–2339. [Google Scholar] [CrossRef]
- Bhattacharya, P. Assessment of Arsenic Accumulation by Different Varieties of Rice (Oryza sativa L.) Irrigated with Arsenic-contaminated Groundwater in West Bengal (India). Environ. Pollut. Prot. 2017, 2, 92–99. [Google Scholar] [CrossRef]
- Pinson, S.R.; Tarpley, L.; Yan, W.; Yeater, K.; Lahner, B.; Yakubova, E.; Huang, X.-Y.; Zhang, M.; Guerinot, M.L.; Salt, D.E. Worldwide genetic diversity for mineral element concentrations in rice grain. Crop. Sci. 2015, 55, 294–311. [Google Scholar] [CrossRef]
- Rahman, M.A.; Hasegawa, H.; Rahman, M.M.; Rahman, M.A.; Miah, M. Accumulation of arsenic in tissues of rice plant (Oryza sativa L.) and its distribution in fractions of rice grain. Chemosphere 2007, 69, 942–948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.; Dubey, A.K.; Upadhyay, A.K.; Gautam, A.; Ranjan, R.; Srikishna, S.; Sahu, N.; Behera, S.K.; Mallick, S. GABA accretion reduces Lsi-1 and Lsi-2 gene expressions and modulates physiological responses in Oryza sativa to provide tolerance towards arsenic. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Xu, X.-Y.; Su, Y.-H.; McGrath, S.P.; Zhao, F.-J. Transporters of arsenite in rice and their role in arsenic accumulation in rice grain. Proc. Natl. Acad. Sci. USA 2008, 105, 9931–9935. [Google Scholar] [CrossRef] [Green Version]
- Ranjan, R.; Kumar, N.; Dubey, A.K.; Gautam, A.; Pandey, S.N.; Mallick, S. Diminution of arsenic accumulation in rice seedlings co-cultured with Anabaena sp.: Modulation in the expression of lower silicon transporters, two nitrogen dependent genes and lowering of antioxidants activity. Ecotoxicol. Environ. Saf. 2018, 151, 109–117. [Google Scholar] [CrossRef]
- Yu, L.j.; Luo, Y.f.; Liao, B.; Xie, L.j.; Chen, L.; Xiao, S.; Li, J.t.; Hu, S.n.; Shu, W.s. Comparative transcriptome analysis of transporters, phytohormone and lipid metabolism pathways in response to arsenic stress in rice (Oryza sativa). New Phytol. 2012, 195, 97–112. [Google Scholar] [CrossRef]
- Hartley-Whitaker, J.; Ainsworth, G.; Meharg, A.A. Copper-and arsenate-induced oxidative stress in Holcus lanatus L. clones with differential sensitivity. Plant Cell Environ. Res. 2001, 24, 713–722. [Google Scholar] [CrossRef]
- Fang, C.-X.; Wang, Q.-S.; Yu, Y.; Li, Q.-M.; Zhang, H.-L.; Wu, X.-C.; Chen, T.; Lin, W.-X. Suppression and overexpression of Lsi1 induce differential gene expression in rice under ultraviolet radiation. Plant Growth Regul. 2011, 65, 1–10. [Google Scholar] [CrossRef]
- Cock, J.; Yoshida, S.; Forno, D.A. Laboratory Manual for Physiological Studies of Rice. Int. Rice Res. Inst. 1976. [Google Scholar]
- Meharg, A.A.; Jardine, L. Arsenite transport into paddy rice (Oryza sativa) roots. New Phytol. 2003, 157, 39–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallmark, C.; Wilding, L.; Smeck, N. Silicon. Methods of Soil Analysis: Part. 2 Chemical Microbiological Properties; John Wiley & Sons: Hoboken, NJ, USA, 1983; Volume 9, pp. 263–273. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Chomczynski, P.; Mackey, K. Short technical reports. Modification of the TRI reagent procedure for isolation of RNA from polysaccharide-and proteoglycan-rich sources. Biotechniques 1995, 19, 942–945. [Google Scholar] [PubMed]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Sun, Z.; Li, Z.; Xue, R.; Cui, W.; Sun, S.; Liu, T.; Zeng, R.; Song, Y. Deficiency in silicon transporter Lsi1 compromises inducibility of anti-herbivore defense in rice plants. Front. Plant Sci. 2019, 10, 652. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Fang, C.; Li, Y.; Lin, W.; He, J.; Lin, R.; Lin, W. Cadmium-stress mitigation through gene expression of rice and silicon addition. Plant Growth Regul. 2017, 81, 91–101. [Google Scholar] [CrossRef]
- Liang, Y.; Wong, J.; Wei, L. Silicon-mediated enhancement of cadmium tolerance in maize (Zea mays L.) grown in cadmium contaminated soil. Chemosphere 2005, 58, 475–483. [Google Scholar] [CrossRef]
- Wang, H.-Y.; Wen, S.-L.; Chen, P.; Zhang, L.; Cen, K.; Sun, G.-X. Mitigation of cadmium and arsenic in rice grain by applying different silicon fertilizers in contaminated fields. Environ. Sci. Pollut. Res. 2016, 23, 3781–3788. [Google Scholar] [CrossRef]
- Sahebi, M.; Hanafi, M.M.; Rafii, M.; Azizi, P.; Abiri, R.; Kalhori, N.; Atabaki, N. Screening and expression of a silicon transporter gene (Lsi1) in wild-type indica rice cultivars. BioMed Res. Int. 2017, 2017. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, P.; Tripathi, R.D.; Singh, R.P.; Dwivedi, S.; Goutam, D.; Shri, M.; Trivedi, P.K.; Chakrabarty, D. Silicon mediates arsenic tolerance in rice (Oryza sativa L.) through lowering of arsenic uptake and improved antioxidant defence system. Ecol. Eng. 2013, 52, 96–103. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N.; Tamai, K.; Mitani, N. Genotypic difference in silicon uptake and expression of silicon transporter genes in rice. Plant Physiol. 2007, 145, 919–924. [Google Scholar] [CrossRef] [Green Version]
- Dubey, A.K.; Kumar, N.; Ranjan, R.; Gautam, A.; Pande, V.; Sanyal, I.; Mallick, S. Application of glycine reduces arsenic accumulation and toxicity in Oryza sativa L. by reducing the expression of silicon transporter genes. Ecotoxicol. Environ. Saf. 2018, 148, 410–417. [Google Scholar] [CrossRef]
- Ashfaque, F.; Inam, A.; Iqbal, S.; Sahay, S. Response of silicon on metal accumulation, photosynthetic inhibition and oxidative stress in chromium-induced mustard (Brassica juncea L.). South Afr. J. Bot. 2017, 111, 153–160. [Google Scholar] [CrossRef]
- Zhen, S.; van Iersel, M.W. Far-red light is needed for efficient photochemistry and photosynthesis. J. Plant Physiol. 2017, 209, 115–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, A.; Wang, X.; Wu, L.; Wang, F.; Wu, Z.; Yang, H.; Chen, Y.; Wen, D.; Liu, X. Silicon improves growth and alleviates oxidative stress in rice seedlings (Oryza sativa L.) by strengthening antioxidant defense and enhancing protein metabolism under arsanilic acid exposure. Ecotoxicol. Environ. Saf. 2018, 158, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Bharwana, S.; Ali, S.; Farooq, M.; Iqbal, N.; Abbas, F.; Ahmad, M. Alleviation of lead toxicity by silicon is related to elevated photosynthesis, antioxidant enzymes suppressed lead uptake and oxidative stress in cotton. Bioremed. Biodeg. 2013, 4, 187. [Google Scholar]
- Farooq, M.A.; Ali, S.; Hameed, A.; Ishaque, W.; Mahmood, K.; Iqbal, Z. Alleviation of cadmium toxicity by silicon is related to elevated photosynthesis, antioxidant enzymes; suppressed cadmium uptake and oxidative stress in cotton. Ecotoxicol. Environ. Saf. 2013, 96, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Dirr, H.; Reinemer, P.; Huber, R. X-ray crystal structures of cytosolic glutathione S-transferases: Implications for protein architecture, substrate recognition and catalytic function. Eur. J. Biochem. 1994, 220, 645–661. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Wu, S.; Yu, R.; Cang, T.; Chen, L.; Zhao, X.; Cai, L.; Wu, C. Identification and expression pattern of ag lutathione S-transferase in E chinochloa crus-galli. Weed Res. 2013, 53, 314–321. [Google Scholar] [CrossRef]
- Jain, M.; Ghanashyam, C.; Bhattacharjee, A. Comprehensive expression analysis suggests overlapping and specific roles of rice glutathione S-transferase genes during development and stress responses. BMC Genom. 2010, 11, 73. [Google Scholar] [CrossRef] [Green Version]
- Wagner, U.; Edwards, R.; Dixon, D.P.; Mauch, F. Probing the diversity of the Arabidopsis glutathione S-transferase gene family. Plant Mol. Biol. 2002, 49, 515–532. [Google Scholar] [CrossRef] [PubMed]
- Nounjan, N.; Nghia, P.T.; Theerakulpisut, P. Exogenous proline and trehalose promote recovery of rice seedlings from salt-stress and differentially modulate antioxidant enzymes and expression of related genes. J. Plant Physiol. 2012, 169, 596–604. [Google Scholar] [CrossRef]
- Radyukina, N.; Shashukova, A.; Makarova, S.; Kuznetsov, V.V. Exogenous proline modifies differential expression of superoxide dismutase genes in UV-B-irradiated Salvia officinalis plants. Russ. J. Plant Physiol. 2011, 58, 51–59. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Khan, A.L.; Kim, D.-H.; Lee, S.-Y.; Kim, K.-M.; Waqas, M.; Jung, H.-Y.; Shin, J.-H.; Kim, J.-G.; Lee, I.-J. Silicon mitigates heavy metal stress by regulating P-type heavy metal ATPases, Oryza sativa low silicon genes, and endogenous phytohormones. BMC Plant Biol. 2014, 14, 13. [Google Scholar] [CrossRef] [Green Version]
- Meharg, C.; Meharg, A.A. Silicon, the silver bullet for mitigating biotic and abiotic stress, and improving grain quality, in rice? Environ. Exp. Bot. 2015, 120, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Bouain, N.; Shahzad, Z.; Rouached, A.; Khan, G.A.; Berthomieu, P.; Abdelly, C.; Poirier, Y.; Rouached, H. Phosphate and zinc transport and signalling in plants: Toward a better understanding of their homeostasis interaction. J. Exp. Bot. 2014, 65, 5725–5741. [Google Scholar] [CrossRef] [Green Version]
- Khan, E.; Gupta, M. Arsenic–silicon priming of rice (Oryza sativa L.) seeds influence mineral nutrient uptake and biochemical responses through modulation of Lsi-1, Lsi-2, Lsi-6 and nutrient transporter genes. Sci. Rep. 2018, 8, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) | Primer Length (nt) | Tm (°C) | PCR Product Length (bp) |
|---|---|---|---|---|
| Os10g0530500-F | ACAACATGTTCCCTGGAATGG | 21 | 55.6 | 166 |
| Os10g0530500-R | TCGACGTACCCGATGGAGTC | 20 | 59.5 | |
| Os08g0522400-F | CAAAGACAAGCTTTCACCGTAA | 22 | 53.9 | 282 |
| Os08g0522400-R | CAGAAAAGAACGCTGCCTTTAA | 22 | 53.9 | |
| Os01g0369700-F | GTTCGGTGAGATTCCAGTACTG | 22 | 57.7 | 86 |
| Os01g0369700-R | TGTACTTGCGAAAGATGTACCT | 22 | 53.9 | |
| GSTU1-F | GTGAGTTTGTTGTTACCGTTGA | 22 | 53.9 | 84 |
| GSTU1-R | TGACAATCTCAGAGAATCGGAG | 22 | 55.8 | |
| GSTZ5-F | AAGATTGTCGCGATTGATCTTG | 22 | 53.9 | 95 |
| GSTZ5-R | TGATTGTTGTGCTCAAGTGAAG | 22 | 53.9 | |
| Os05g0240200-F | CTGAAGATGTTGGCTACTTTCG | 22 | 55.8 | 177 |
| Os05g0240200-R | CATCTTTCAGGAACCGCATATG | 22 | 55.8 | |
| ATG8D-F | TCTTCTGGAGTCTACACGTCTA | 22 | 55.8 | 111 |
| ATG8D-R | GTCTTCTTCCTTGATGCGAATC | 22 | 55.8 | |
| LOC_Os10g10130-F | TTTGGATGCTGTCTTGAAACTG | 22 | 53.9 | 87 |
| LOC_Os10g10130-R | ATCTGGATGAAGTAGTCCGAAC | 22 | 55.8 | |
| Os02g0685200-F | CGGTGGGTTCTCGAATAACTC | 21 | 57.6 | 193 |
| Os02g0685200-R | CGTGGTTGCAATTGACATCTTA | 22 | 53.9 | |
| actin1(Os03g0718100)-F | CTTCATAGGAATGGAAGCTGCGGGTA | 26 | 61.2 | 197 |
| actin1(Os03g0718100)-R | CGACCACCTTGATCTTCATGCTGCTA | 26 | 61.2 |
| Sampling Time | Parameter | Line | As | Si | Line × As | Line × Si | As × Si | Line × As × Si |
|---|---|---|---|---|---|---|---|---|
| 1st week | Shoot As concentration | 344.84 ** | 15,523.48 ** | 2192.96 ** | 236.03 ** | 85.79 ** | 848.92 ** | 151.63 ** |
| Root As concentration | 182.45 ** | 25,123.40 ** | 5047.21 ** | 122.71 ** | 270.55 ** | 1568.16 ** | 213.83 ** | |
| Shoot dry weight | 6.96 ** | 69.76 ** | 34.11 ** | 0.32 ns | 0.55 ns | 1.05 ns | 0.32 ns | |
| Root dry weight | 3.29 * | 62.42 ** | 35.89 ** | 0.37 ns | 0.33 ns | 1.90 ns | 0.09 ns | |
| 2nd week | Shoot As concentration | 987.26 ** | 14,477.16 ** | 3755.15 ** | 861.59 ** | 359.47 ** | 1547.55 ** | 299.44 ** |
| Root As concentration | 395.65 ** | 28,989.96 ** | 7140.00 ** | 244.32 ** | 273.65 ** | 1789.42 ** | 122.37 ** | |
| Shoot dry weight | 3.98 * | 77.01 ** | 41.90 ** | 0.17 ns | 0.29 ns | 2.22 ns | 0.16 ns | |
| Root dry weight | 1.69 ns | 74.41 ** | 47.70 ** | 0.12 ns | 0.28 ns | 2.23 ns | 0.11 ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boorboori, M.R.; Lin, W.; Jiao, Y.; Fang, C. Silicon Modulates Molecular and Physiological Activities in Lsi1 Transgenic and Wild Lemont Rice Seedlings under Arsenic Stress. Agronomy 2021, 11, 1532. https://doi.org/10.3390/agronomy11081532
Boorboori MR, Lin W, Jiao Y, Fang C. Silicon Modulates Molecular and Physiological Activities in Lsi1 Transgenic and Wild Lemont Rice Seedlings under Arsenic Stress. Agronomy. 2021; 11(8):1532. https://doi.org/10.3390/agronomy11081532
Chicago/Turabian StyleBoorboori, Mohammad Reza, Wenxiong Lin, Yanyang Jiao, and Changxun Fang. 2021. "Silicon Modulates Molecular and Physiological Activities in Lsi1 Transgenic and Wild Lemont Rice Seedlings under Arsenic Stress" Agronomy 11, no. 8: 1532. https://doi.org/10.3390/agronomy11081532
APA StyleBoorboori, M. R., Lin, W., Jiao, Y., & Fang, C. (2021). Silicon Modulates Molecular and Physiological Activities in Lsi1 Transgenic and Wild Lemont Rice Seedlings under Arsenic Stress. Agronomy, 11(8), 1532. https://doi.org/10.3390/agronomy11081532