
The Modulation of Auxin-Responsive Genes, Phytohormone Profile, and Metabolomic Signature in Leaves of Tomato Cuttings Is Specifically Modulated by Different Protein Hydrolysates

 ,
, 
 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Auxin-Like Activity Bioassays
2.2. Gene Expression Analysis
2.3. Metabolomic Analysis
2.4. Statistical Analyses
3. Results
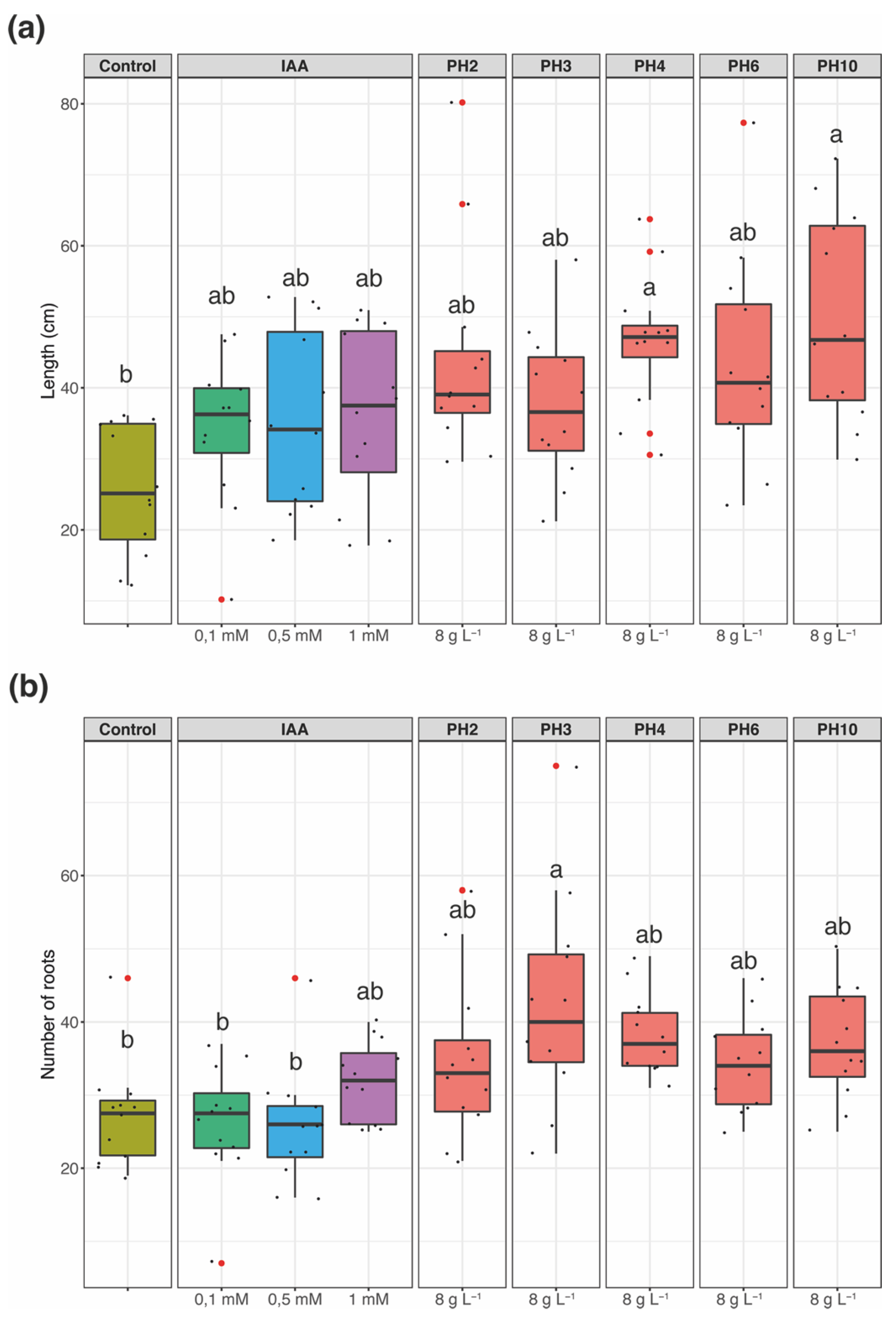
3.1. Rooting Bioassay
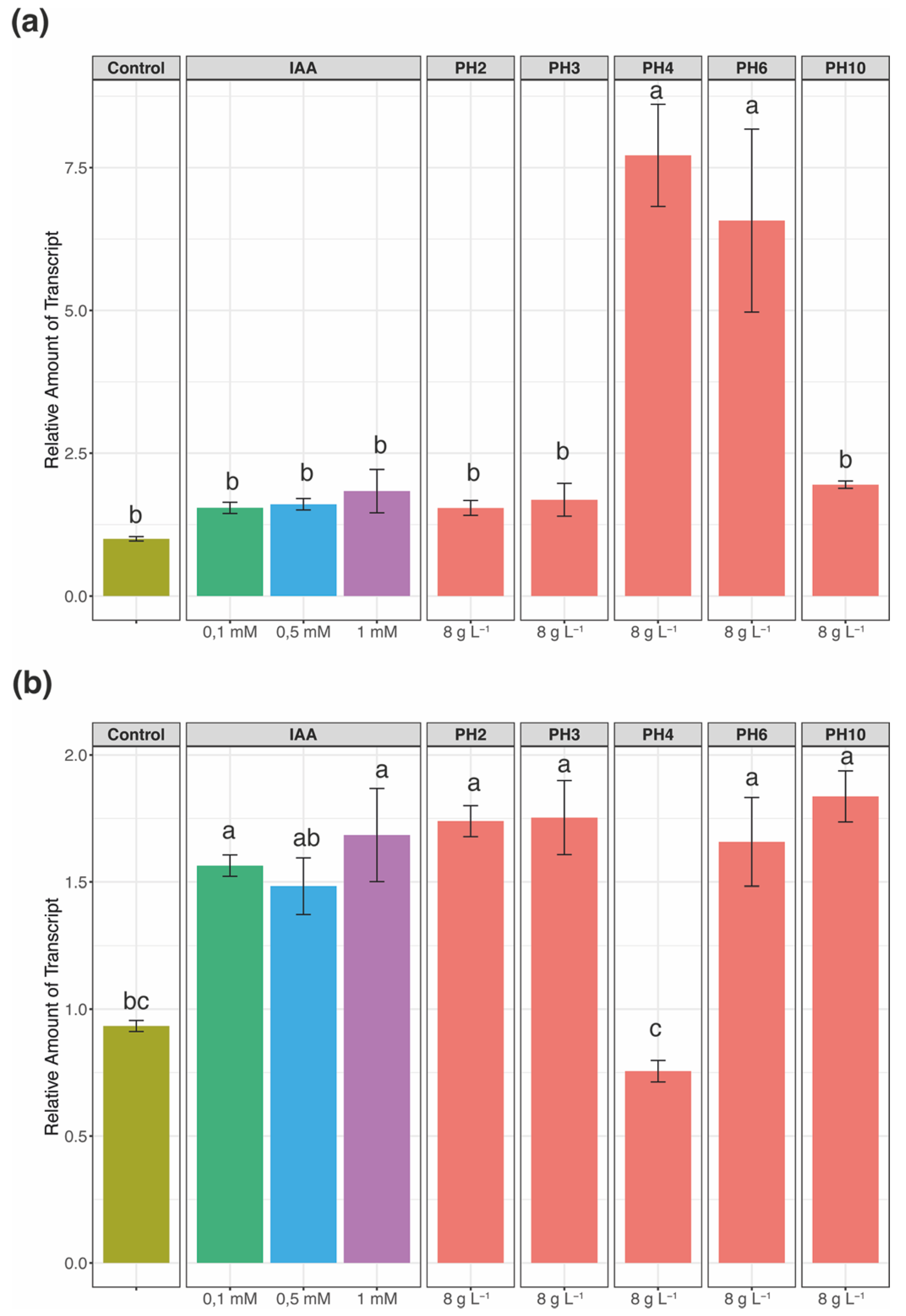
3.2. Effects on IAA Responsive Genes
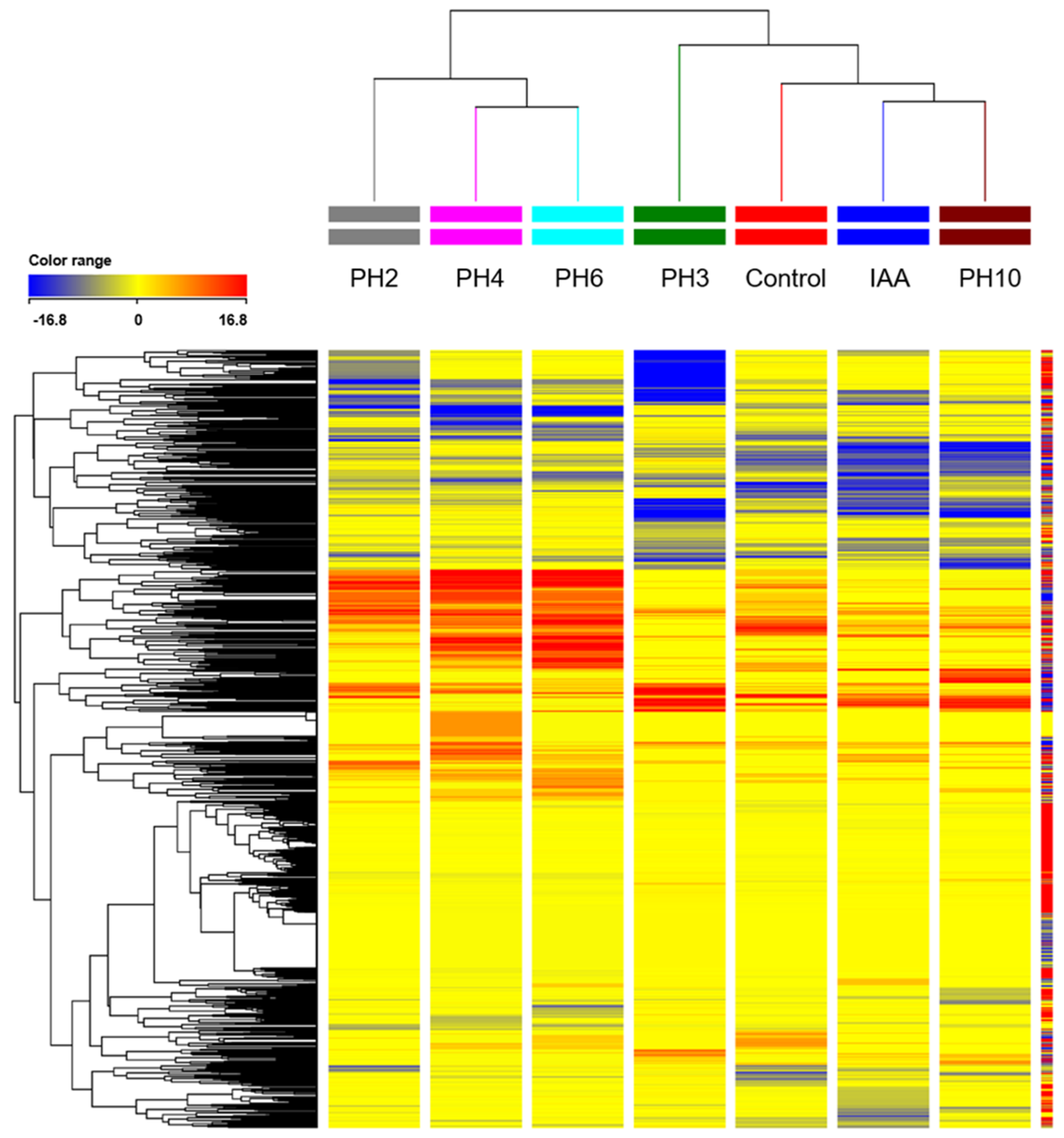
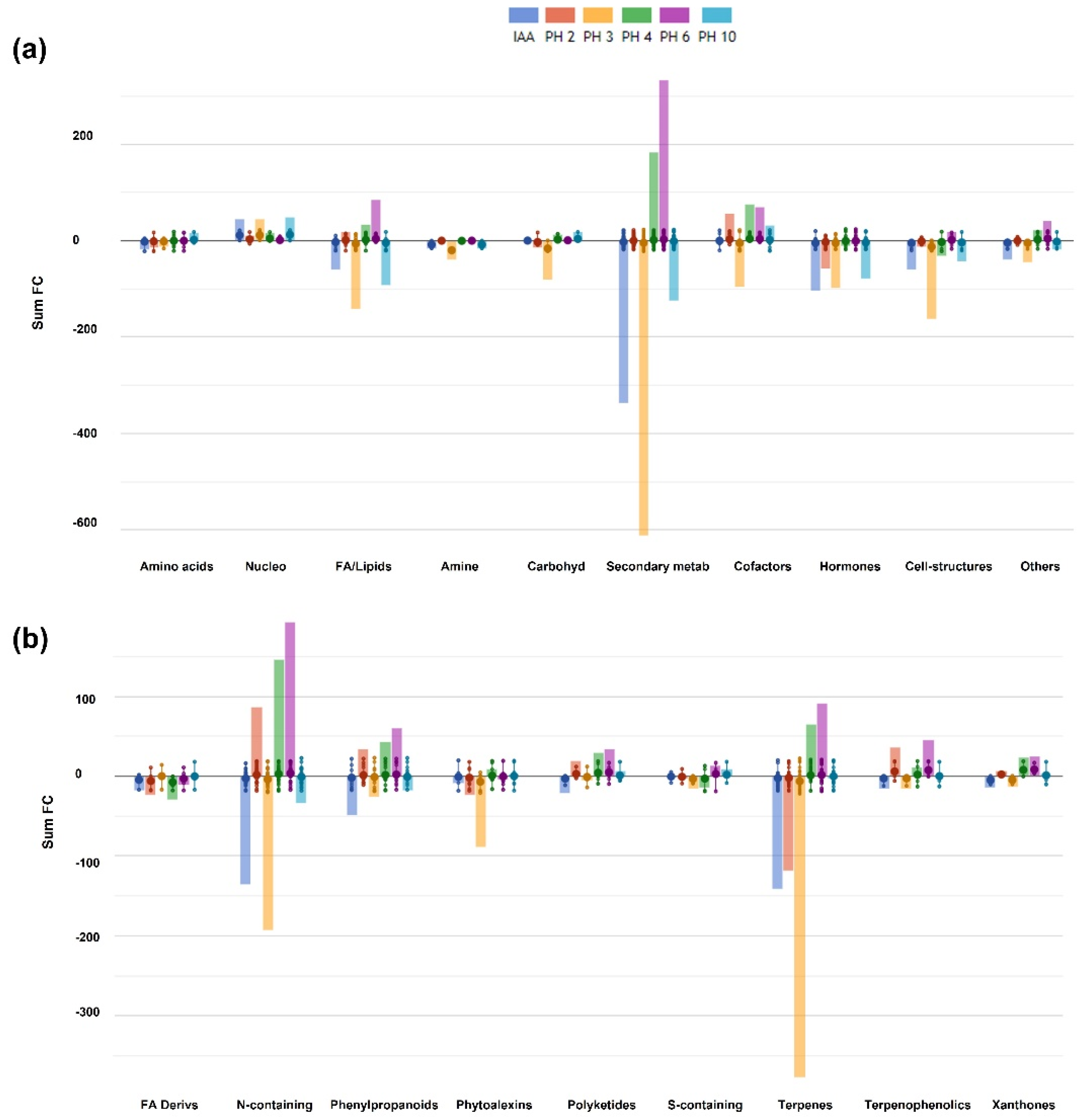
3.3. Untargeted Metabolomic Profiling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Calicioglu, O.; Flammini, A.; Bracco, S.; Bellù, L.; Sims, R. The future challenges of food and agriculture: An integrated analysis of trends and solutions. Sustainability 2019, 11, 222. [Google Scholar] [CrossRef] [Green Version]
- Taiz, L. Agriculture, plant physiology, and human population growth: Past, present, and future. Theor. Exp. Plant Physiol. 2013, 25, 167–181. [Google Scholar] [CrossRef] [Green Version]
- Koppelaar, R.H.E.M.; Weikard, H.P. Assessing phosphate rock depletion and phosphorus recycling options. Glob. Environ. Chang. 2013, 23, 1454–1466. [Google Scholar] [CrossRef]
- Prăvălie, R.; Patriche, C.; Borrelli, P.; Panagos, P.; Roșca, B.; Dumitraşcu, M.; Nita, I.A.; Săvulescu, I.; Birsan, M.V.; Bandoc, G. Arable lands under the pressure of multiple land degradation processes. A global perspective. Environ. Res. 2021, 194, 110697. [Google Scholar] [CrossRef]
- Caradonia, F.; Battaglia, V.; Righi, L.; Pascali, G.; La Torre, A. Plant biostimulant regulatory framework: Prospects in Europe and current situation at international level. J. Plant Growth Regul. 2019, 38, 438–448. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauffman, G.L.; Kneivel, D.P.; Watschke, T.L. Effects of a biostimulant on the heat tolerance associated with photosynthetic capacity, membrane thermostability, and polyphenol production of perennial ryegrass. Crop Sci. 2007, 47, 261–267. [Google Scholar] [CrossRef]
- Jardin, P.; Xu, L.; Geelen, D. Agricultural Functions and Action Mechanisms of Plant Biostimulants (PBs). In The Chemical Biology of Plant Biostimulants; Wiley: Hoboken, NJ, USA, 2020; pp. 1–30. [Google Scholar]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: A global perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Colantoni, A.; Recchia, L.; Bernabei, G.; Cardarelli, M.; Rouphael, Y.; Colla, G. Analyzing the environmental impact of chemically-produced protein hydrolysate from leather waste vs. Enzymatically-produced protein hydrolysate from legume grains. Agriculture 2017, 7, 62. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Rouphael, Y.; Canaguier, R.; Svecova, E.; Cardarelli, M. Biostimulant action of a plant-derived protein hydrolysate produced through enzymatic hydrolysis. Front. Plant Sci. 2014, 5, 448. [Google Scholar] [CrossRef] [Green Version]
- Ertani, A.; Pizzeghello, D.; Francioso, O.; Tinti, A.; Nardi, S. Biological activity of vegetal extracts containing phenols on plant metabolism. Molecules 2016, 21, 205. [Google Scholar] [CrossRef]
- Michniewicz, M.; Brewer, P.B.; Friml, J. Polar auxin transport and asymmetric auxin distribution. Arab. Book 2007, 5, e0108. [Google Scholar] [CrossRef] [Green Version]
- Casanova-Sáez, R.; Mateo-Bonmatí, E.; Ljung, K. Auxin Metabolism in Plants. Cold Spring Harb. Perspect. Biol. 2021, 13, a039867. [Google Scholar] [CrossRef]
- Petrozza, A.; Santaniello, A.; Summerer, S.; Di Tommaso, G.; Di Tommaso, D.; Paparelli, E.; Piaggesi, A.; Perata, P.; Cellini, F. Physiological responses to Megafol® treatments in tomato plants under drought stress: A phenomic and molecular approach. Sci. Hortic. 2014, 174, 185–192. [Google Scholar] [CrossRef]
- Lucini, L.; Miras-Moreno, B.; Rouphael, Y.; Cardarelli, M.; Colla, G. Combining molecular weight fractionation and metabolomics to elucidate the bioactivity of vegetal protein hydrolysates in tomato plants. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Evans, M.L.; Ishikawa, H.; Estelle, M.A. Responses of Arabidopsis roots to auxin studied with high temporal resolution: Comparison of wild type and auxin-response mutants. Planta 1994, 194, 215–222. [Google Scholar] [CrossRef]
- Muday, G.K.; Lomax, T.L.; Rayle, D.L. Characterization of the growth and auxin physiology of roots of the tomato mutant, diageotropica. Planta 1995, 195, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Ertani, A.; Cavani, L.; Pizzeghello, D.; Brandellero, E.; Altissimo, A.; Ciavatta, C.; Nardi, S. Biostimulant activity of two protein hydrolyzates in the growth and nitrogen metabolism of maize seedlings. J. Plant Nutr. Soil Sci. 2009, 172, 237–244. [Google Scholar] [CrossRef]
- Ceccarelli, A.V.; Miras-Moreno, B.; Buffagni, V.; Senizza, B.; Pii, Y.; Cardarelli, M.; Rouphael, Y.; Colla, G.; Lucini, L. Foliar application of different vegetal-derived protein hydrolysates distinctively modulates tomato root development and metabolism. Plants 2021, 10, 326. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Ku, K.M.; Choi, S.; Cardarelli, M. Vegetal-derived biostimulant enhances adventitious rooting in cuttings of basil, tomato, and chrysanthemum via brassinosteroid-mediated processes. Agronomy 2019, 9, 74. [Google Scholar] [CrossRef] [Green Version]
- Pii, Y.; Alessandrini, M.; Dall’Osto, L.; Guardini, K.; Prinsi, B.; Espen, L.; Zamboni, A.; Varanini, Z. Time-resolved investigation of molecular components involved in the induction of NO3− high affinity transport system in maize roots. Front. Plant Sci. 2016, 7, 1657. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST©) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Salehi, H.; Chehregani, A.; Lucini, L.; Majd, A.; Gholami, M. Morphological, proteomic and metabolomic insight into the effect of cerium dioxide nanoparticles to Phaseolus vulgaris L. under soil or foliar application. Sci. Total Environ. 2018, 616–617, 1540–1551. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Lucini, L.; Miras-Moreno, B.; Colla, G.; Bonini, P.; Cardarelli, M. Metabolomic responses of maize shoots and roots elicited by combinatorial seed treatments with microbial and non-microbial biostimulants. Front. Microbiol. 2020, 11, 664. [Google Scholar] [CrossRef]
- Miras-Moreno, B.; Corrado, G.; Zhang, L.; Senizza, B.; Righetti, L.; Bruni, R.; El-Nakhel, C.; Sifola, M.I.; Pannico, A.; De Pascale, S.; et al. The metabolic reprogramming induced by sub-optimal nutritional and light inputs in soilless cultivated green and red butterhead lettuce. Int. J. Mol. Sci. 2020, 21, 6381. [Google Scholar] [CrossRef]
- Salek, R.M.; Steinbeck, C.; Viant, M.R.; Goodacre, R.; Dunn, W.B. The role of reporting standards for metabolite annotation and identification in metabolomic studies. Gigascience 2013, 2, 13. [Google Scholar] [CrossRef]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; Vandergheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef]
- Tsugawa, H.; Kind, T.; Nakabayashi, R.; Yukihira, D.; Tanaka, W.; Cajka, T.; Saito, K.; Fiehn, O.; Arita, M. Hydrogen Rearrangement Rules: Computational MS/MS Fragmentation and Structure Elucidation Using MS-FINDER Software. Anal. Chem. 2016, 88, 7946–7958. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer, 2016; ISBN 3319242776. Available online: https://www.springer.com/jp/book/9783319242750 (accessed on 12 June 2020).
- de Mendiburu, F. Package ‘agricolae’. R Packag. Version 2019, 1–2. [Google Scholar]
- Tang, Y.; Horikoshi, M.; Li, W. Ggfortify: Unified interface to visualize statistical results of popular r packages. R J. 2016, 8, 478–489. [Google Scholar] [CrossRef] [Green Version]
- Caspi, R.; Dreher, K.; Karp, P.D. The challenge of constructing, classifying, and representing metabolic pathways. FEMS Microbiol. Lett. 2013, 345, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Karp, P.D.; Paley, S.M.; Krummenacker, M.; Latendresse, M.; Dale, J.M.; Lee, T.J.; Kaipa, P.; Gilham, F.; Spaulding, A.; Popescu, L.; et al. Pathway Tools version 13.0: Integrated software for pathway/genome informatics and systems biology. Brief. Bioinform. 2010, 11, 40–79. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Jones, B.; Li, Z.; Frasse, P.; Delalande, C.; Regad, F.; Chaabouni, S.; Latché, A.; Pech, J.-C.; Bouzayen, M. The Tomato Aux/IAA Transcription Factor IAA9 Is Involved in Fruit Development and Leaf Morphogenesis. Plant Cell 2005, 17, 2676 LP–2692. [Google Scholar] [CrossRef] [Green Version]
- Celletti, S.; Astolfi, S.; Guglielmo, N.; Colla, G.; Cesco, S.; Mimmo, T. Evaluation of a legume-derived protein hydrolysate to mitigate iron deficiency in plants. Agronomy 2020, 10, 1942. [Google Scholar] [CrossRef]
- Luo, J.; Zhou, J.-J.; Zhang, J.-Z. Aux/IAA gene family in plants: Molecular structure, regulation, and function. Int. J. Mol. Sci. 2018, 19, 259. [Google Scholar] [CrossRef] [Green Version]
- Abel, S.; Nguyen, M.D.; Theologis, A. The PS-IAA4/5-like family of early auxin-inducible mRNAs in Arabidopsis thaliana. J. Mol. Biol. 1995, 251, 533–549. [Google Scholar] [CrossRef] [PubMed]
- Rogg, L.E.; Lasswell, J.; Bartel, B. A gain-of-function mutation in IAA28 suppresses lateral root development. Plant Cell 2001, 13, 465–480. [Google Scholar] [CrossRef] [Green Version]
- Druege, U.; Hilo, A.; Pérez-Pérez, J.M.; Klopotek, Y.; Acosta, M.; Shahinnia, F.; Zerche, S.; Franken, P.; Hajirezaei, M.R. Molecular and physiological control of adventitious rooting in cuttings: Phytohormone action meets resource allocation. Ann. Bot. 2019, 123, 929–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.H.; Irving, H.R. Developing a model of plant hormone interactions. Plant Signal. Behav. 2011, 6, 494–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Z.; Li, J. Brassinosteroids regulate root growth, development, and symbiosis. Mol. Plant 2016, 9, 86–100. [Google Scholar] [CrossRef] [Green Version]
- González-García, M.P.; Vilarrasa-Blasi, J.; Zhiponova, M.; Divol, F.; Mora-García, S.; Russinova, E.; Caño-Delgado, A.I. Brassinosteroids control meristem size by promoting cell cycle progression in Arabidopsis roots. Development 2011, 138, 849–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouchel, C.F.; Osmont, K.S.; Hardtke, C.S. BRX mediates feedback between brassinosteroid levels and auxin signalling in root growth. Nature 2006, 443, 458–461. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, M.; Brütting, C.; Meza-Canales, I.D.; Großkinsky, D.K.; Vankova, R.; Baldwin, I.T.; Meldau, S. The role of cis-zeatin-type cytokinins in plant growth regulation and mediating responses to environmental interactions. J. Exp. Bot. 2015, 66, 4873–4884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, L.; Tayengwa, R.; Cheng, Z.M.; Peer, W.A.; Murphy, A.S.; Zhao, M. Auxin regulates adventitious root formation in tomato cuttings. BMC Plant Biol. 2019, 19, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Zhao, P.X.; Cai, X.T.; Mao, J.L.; Miao, Z.Q.; Xiang, C. Bin Integration of Jasmonic Acid and Ethylene Into auxin signaling in root development. Front. Plant Sci. 2020, 11, 271. [Google Scholar] [CrossRef]
- Gutierrez, L.; Mongelard, G.; Floková, K.; Pǎcurar, D.I.; Novák, O.; Staswick, P.; Kowalczyk, M.; Pǎcurar, M.; Demailly, H.; Geiss, G.; et al. Auxin controls Arabidopsis adventitious root initiation by regulating jasmonic acid homeostasis. Plant Cell 2012, 24, 2515–2527. [Google Scholar] [CrossRef] [Green Version]
- Lakehal, A.; Chaabouni, S.; Cavel, E.; Le Hir, R.; Ranjan, A.; Raneshan, Z.; Novák, O.; Păcurar, D.I.; Perrone, I.; Jobert, F.; et al. A molecular framework for the control of adventitious rooting by TIR1/AFB2-Aux/IAA-dependent auxin signaling in Arabidopsis. Mol. Plant 2019, 12, 1499–1514. [Google Scholar] [CrossRef]
- Ruiz Rosquete, M.; Barbez, E.; Kleine-Vehn, J. Cellular auxin homeostasis: Gatekeeping is housekeeping. Mol. Plant 2012, 5, 772–786. [Google Scholar] [CrossRef] [Green Version]
- Ludwig-Müller, J. Auxin conjugates: Their role for plant development and in the evolution of land plants. J. Exp. Bot. 2011, 62, 1757–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Peer, W.A. Auxin homeostasis: The DAO of catabolism. J. Exp. Bot. 2017, 68, 3145–3154. [Google Scholar] [CrossRef] [Green Version]
- Porco, S.; Pěnčík, A.; Rasheda, A.; Vo, U.; Casanova-Sáez, R.; Bishopp, A.; Golebiowska, A.; Bhosale, R.; Swarupa, R.; Swarup, K.; et al. Dioxygenase-encoding AtDAO1 gene controls IAA oxidation and homeostasis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2016, 113, 11016–11021. [Google Scholar] [CrossRef] [Green Version]
- Olatunji, D.; Geelen, D.; Verstraeten, I. Control of endogenous auxin levels in plant root development. Int. J. Mol. Sci. 2017, 18, 2587. [Google Scholar] [CrossRef] [Green Version]
- Gee, M.A.; Hagen, G.; Guilfoyle, T.J. Tissue-specific and organ-specific expression of soybean auxin-responsive transcripts GH3 and SAURs. Plant Cell 1991, 3, 419–430. [Google Scholar] [PubMed] [Green Version]
- Kelley, K.B.; Lambert, K.N.; Hager, A.G.; Riechers, D.E. Quantitative expression analysis of GH3, a gene induced by plant growth regulator herbicides in soybean. J. Agric. Food Chem. 2004, 52, 474–478. [Google Scholar] [CrossRef]
- LeClere, S.; Tellez, R.; Rampey, R.A.; Matsuda, S.P.T.; Bartel, B. Characterization of a family of IAA-amino acid conjugate hydrolases from Arabidopsis. J. Biol. Chem. 2002, 277, 20446–20452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rampey, R.A.; LeClere, S.; Kowalczyk, M.; Ljung, K.; Sandberg, G.; Bartel, B. A family of auxin-conjugate hydrolases that contributes to free indole-3-acetic acid levels during Arabidopsis germination. Plant Physiol. 2004, 135, 978–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staswick, P.E.; Serban, B.; Rowe, M.; Tiryaki, I.; Maldonado, M.T.; Maldonado, M.C.; Suza, W. Characterization of an Arabidopsis enzyme family that conjugates amino acids to indole-3-acetic acid. Plant Cell 2005, 17, 616–627. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Xu, R.; Ma, C.J.; Vlot, A.C.; Klessig, D.F.; Pichersky, E. Inactive methyl indole-3-acetic acid ester can be hydrolyzed and activated by several esterases belonging to the AtMES esterase family of Arabidopsis. Plant Physiol. 2008, 147, 1034–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant action of protein hydrolysates: Unraveling their effects on plant physiology and microbiome. Front. Plant Sci. 2017, 8, 2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böttger, A.; Vothknecht, U.; Bolle, C.; Wolf, A. Plant secondary metabolites and their general function in plants. In Lessons on Caffeine, Cannabis and Co; Springer: Cham, Switzerland, 2018; pp. 3–17. [Google Scholar]
- Kurepa, J.; Shull, T.E.; Karunadasa, S.S.; Smalle, J.A. Modulation of auxin and cytokinin responses by early steps of the phenylpropanoid pathway. BMC Plant Biol. 2018, 18, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Baffi, C.; Colla, G. A vegetal biopolymer-based biostimulant promoted root growth in melon while triggering brassinosteroids and stress-related compounds. Front. Plant Sci. 2018, 9, 472. [Google Scholar] [CrossRef] [Green Version]
- Bonini, P.; Rouphael, Y.; Miras-Moreno, B.; Lee, B.; Cardarelli, M.; Erice, G.; Cirino, V.; Lucini, L.; Colla, G. A microbial-based biostimulant enhances sweet pepper performance by metabolic reprogramming of phytohormone profile and secondary netabolism. Front. Plant Sci. 2020, 11, 1727. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Log FC | |||||||
|---|---|---|---|---|---|---|---|
| Class | Compound | IAA | PH 2 | PH 3 | PH 4 | PH 6 | PH 10 |
| Abscisic Acid | (3S,5R,6R)-3,5-Dihydroxy-6,7-Didehydro-5,6-Dihydro-12′-Apo-Β-Caroten-12′-Al | −8.76 | −0.36 | −8.64 | −0.97 | −8.98 | −8.83 |
| Violaxanthin | 0.03 | 0.49 | 0.68 | −17.83 | −18.68 | −0.60 | |
| Dihydroxyphaseic Acid | −8.88 | 8.30 | −8.72 | 8.71 | 8.89 | −8.96 | |
| Auxin | (Indol-3-Yl)Acetamide | 3.38 | 3.65 | 3.34 | 3.52 | 3.65 | −14.26 |
| N2-Hydroxy-L-Tryptophan | 3.95 | −16.42 | 4.75 | −16.38 | −16.72 | 4.82 | |
| Methyl (Indol-3-Yl)Acetate | −10.34 | 10.53 | −10.23 | 10.38 | 8.49 | −10.47 | |
| 2-oxoindole-3-acetyl-β-D-glucose (OxIAA-Glc) | −8.69 | −8.84 | 9.79 | −9.09 | −9.12 | 9.57 | |
| (indol-3-yl)acetyl-L-phenylalanine (IAA-Phe) | 14.97 | −4.81 | 15.06 | −5.08 | −5.11 | 14.76 | |
| N-β-D-glucosyl-(indol-3-yl)acetate (IAA-N-Glc) | −3.28 | −3.62 | −3.15 | −3.8 | −3.84 | 0.18 | |
| (indol-3-yl)acetyl-L-glutamine (IAA-Gln) | 0.22 | 0.03 | 0.36 | −3.26 | −2.81 | 0.04 | |
| (indol-3-yl)acetyl-L-valine (IAA-Val) | 0.34 | 21.24 | 0.47 | 16.84 | 16.94 | 0.15 | |
| Brassinosteroid | (22α)-Hydroxy-Campesterol | 8.52 | 0.04 | 24.25 | 23.80 | ||
| (22S,24R)-22-Hydroxy-5α-Ergostan-3-One | 0.34 | 0.04 | 0.47 | 20.89 | 20.92 | 0.15 | |
| 6-Deoxocathasterone | −10.97 | −11.07 | −10.64 | −8.04 | −8.00 | −10.97 | |
| Cytokinin | Dihydrozeatin-9-N-Glucoside | −7.20 | 0.09 | −1.75 | 0.11 | 0.33 | −2.90 |
| Cis-Zeatin-9-N-Glucoside | −8.18 | −18.28 | −5.49 | 0.06 | −18.61 | −10.50 | |
| Lupinate | −5.43 | 2.88 | 12.20 | −5.98 | 2.67 | −5.55 | |
| Ethylene | 1-Aminocyclopropane-1-Carboxylate | 1.73 | −0.18 | 2.54 | −0.20 | −0.07 | 1.68 |
| Gibberellin and Gibberellin Precursor | Gibberellin A38 | −0.15 | −2.76 | −16.85 | 0.94 | 1.10 | −17.23 |
| Gibberellin A24 | −9.55 | −0.95 | −0.21 | −0.36 | −1.30 | −6.49 | |
| Gibberellin A6 | −10.41 | −4.52 | −17.27 | −17.96 | −17.98 | −0.46 | |
| Gibberellin A12-Aldehyde | 19.96 | −0.02 | 0.36 | −0.36 | 19.75 | 19.74 | |
| Ent-7α-Hydroxykaur-16-En-19-Oate | −12.36 | −12.56 | −12.19 | 6.96 | 2.10 | 5.93 | |
| gibberellin A8-catabolite | 1.31 | −11.52 | −2.44 | −11.45 | −11.72 | 2.60 | |
| gibberellin A44 (closed lactone form) (GA44) | 0.48 | 0.12 | 0.55 | 5.95 | −0.16 | −0.11 | |
| methyl gibberellin A4 (GA4) | −3.17 | 3.36 | −3.04 | 13.92 | 13.95 | −3.36 | |
| 16,17-dihydro-16α,17-dihydroxy gibberellin A12 | 0.65 | 1.05 | 2.50 | 0.11 | 0.18 | −0.37 | |
| 16,17-dihydro-16α, 17-dihydroxy gibberellin A4 | −18.42 | −18.50 | 1.15 | −18.42 | −18.42 | ||
| Ent-7α-Hydroxykaur-16-En-19-Oate | −12.36 | −12.56 | −12.19 | 6.96 | 2.1 | 5.93 | |
| Jasmonate | 1-Aminocyclopropane-1-Carboxylate | 1.73 | −0.18 | 2.54 | −0.2 | −0.07 | 1.68 |
| L-Phenylalanine | −4.40 | 4.58 | −4.27 | 18.52 | 16.52 | −4.60 | |
| (9Z,13S,15Z)-12,13-Epoxyoctadeca-9,11,15-Trienoate | −16.76 | −17.05 | −16.64 | −11.57 | −8.7 | −16.82 | |
| Coenzyme A | 0.34 | 0.04 | 0.47 | −0.21 | 3.38 | 18.28 | |
| (+)-7-Epi-Jasmonate | −3.21 | 10.71 | 14.41 | −0.43 | 10.99 | −3.45 | |
| Juvenile hormone III | (2E,6E)-Farnesoate | −17.69 | −17.94 | −17.56 | −18.22 | −18.27 | −17.89 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buffagni, V.; Ceccarelli, A.V.; Pii, Y.; Miras-Moreno, B.; Rouphael, Y.; Cardarelli, M.; Colla, G.; Lucini, L. The Modulation of Auxin-Responsive Genes, Phytohormone Profile, and Metabolomic Signature in Leaves of Tomato Cuttings Is Specifically Modulated by Different Protein Hydrolysates. Agronomy 2021, 11, 1524. https://doi.org/10.3390/agronomy11081524
Buffagni V, Ceccarelli AV, Pii Y, Miras-Moreno B, Rouphael Y, Cardarelli M, Colla G, Lucini L. The Modulation of Auxin-Responsive Genes, Phytohormone Profile, and Metabolomic Signature in Leaves of Tomato Cuttings Is Specifically Modulated by Different Protein Hydrolysates. Agronomy. 2021; 11(8):1524. https://doi.org/10.3390/agronomy11081524
Chicago/Turabian StyleBuffagni, Valentina, Angela Valentina Ceccarelli, Youry Pii, Begoña Miras-Moreno, Youssef Rouphael, Mariateresa Cardarelli, Giuseppe Colla, and Luigi Lucini. 2021. "The Modulation of Auxin-Responsive Genes, Phytohormone Profile, and Metabolomic Signature in Leaves of Tomato Cuttings Is Specifically Modulated by Different Protein Hydrolysates" Agronomy 11, no. 8: 1524. https://doi.org/10.3390/agronomy11081524
APA StyleBuffagni, V., Ceccarelli, A. V., Pii, Y., Miras-Moreno, B., Rouphael, Y., Cardarelli, M., Colla, G., & Lucini, L. (2021). The Modulation of Auxin-Responsive Genes, Phytohormone Profile, and Metabolomic Signature in Leaves of Tomato Cuttings Is Specifically Modulated by Different Protein Hydrolysates. Agronomy, 11(8), 1524. https://doi.org/10.3390/agronomy11081524