3.1. Whole Genome Sequencing of the Strains under Investigation
The genomic features of the studied strains are presented in
Table 2.
The currently described symbiotic
fix, nif, nod, noe and
nol genes as well as genes that promote plant growth (
acdRS, gibberellin- and auxin-biosynthesis related) and genes of the T3SS, T4SS and T6SS secretion systems involved in the formation of symbiosis were searched throughout the whole genome sequences of three commercial rhizobial strains (
S. meliloti RCAM1750;
R. leguminosarum RCAM0626, RCAM1365) and four strains isolated from the relict legumes
O. popoviana (
M. japonicum Opo-235,
M. japonicum Opo-242,
Bradyrhizobium sp. Opo-243) and
A. chorinensis (
M. kowhaii Ach-343). Presence of symbiotic
fix, nif, nod, noe and
nol genes is shown in
Table 3.
The common
nodABC genes necessary for legume nodulation [
17] were found in all commercial strains and only in three isolates:
M. japonicum Opo-235,
M. japonicum Opo-242 and
M. kowhaii Ach-343. These strains possessed also
nif and
fix genes required for nitrogen fixation and forming clusters:
nifHDK and
nifENB genes encoding structural and catalytic components of the nitrogenase complex [
18] as well as
fixABCX genes participating in electron transfer to nitrogenase [
19]. At the same time, some of
fixNOPQ and
fixGHIS genes participating in symbiotically essential oxidase complex [
20] were not observed in the commercial strains: the strain
R. leguminosarum RCAM1365 lacked
fixQHS; the strains
R. leguminosarum RCAM0626 and
S. meliloti RCAM1750 lacked
fixS and
fixH, respectively (
Table 3). The isolate
Bradyrhizobium sp. Opo-243 had only one group of
fixKJLNOQPGHIS genes, which formed a single cluster. All the studied strains possessed some
nol and
noe genes, as well as additional
nod, nif and
fix genes. It should be noted that the set of symbiotic genes for each strain was unique.
Presence of the genes related to plant growth promotion (
acdRS, gibberellin- and auxin-biosynthesis related genes) is shown in
Table 4.
It was repeatedly shown that the
acdS gene of rhizobia encoding enzyme 1-aminocyclopropane-1-carboxylate (ACC) deaminase plays important role in nodulation process [
21]. Particularly, pea plants inoculated with ACC deaminase minus mutants of
Rhizobium leguminosarum bv.
viciae 128C53K [
22] and 1066S [
23] showed decreased nodule number, nitrogenase activity and shoot biomass as compared with a wild type. The commercial strains
R. leguminosarum RCAM0626 and RCAM1365 did not contain the
acdS gene. At the same time, the strain
S. meliloti RCAM1750, as well as the isolates
Bradyrhizobium sp. Opo-243 and
M. kowhaii Ach-343, had both
acdS and
acdR genes required for the synthesis of ACC deaminase and its regulation, respectively. The isolates
M. japonicum Opo-235 and
M. japonicum Opo-242 contained only
acdS gene.
All isolates had significantly more genes associated with the biosynthesis of gibberellins and auxins compared to the commercial strains (
Table 4). It should be noted that gibberellin- and auxin-biosynthesis related genes are commonly known as factors responsible for plant growth, the efficiency of plant–microbe interactions and symbiosis formation. For example, they are involved in the processes of nodule initiation and development [
24,
25,
26].
The genes of T3SS, T4SS and T6SS secretion systems presented in the studied strains are shown in
Table 5. The T3SS secretion system was scarcely present in either commercial or isolated strains by one
fli gene in the isolates
M. japonicum Opo-235 and
M. kowhaii Ach-343. In contrast, all strains, with the exception of the
R. leguminosarum RCAM0626, possessed multiple
vir genes belonging to theT4SS playing an important role in the translocation of a wide range of virulence factors into the host cell [
27,
28]. No genes of the T6SS secretion system were found in the commercial strains or in the isolate
Bradyrhizobium sp. Opo-243, while four remaining isolates had different sets of these genes (
Table 5). Most of them (
icmF, tssABCEGJKL, tagFH, vgrG, vasAEFK, clpV1, impA, vipAB, sciN, dotU, Vgr family, Hcp family, FHA domain) was presented in the isolate
M. kowhaii Ach-343. The T6SS secretion system, like T3SS and T4SS, is involved in bacterial virulence due to secretion of broad classes of effectors proteins: cytotoxins; lysozymes; lipoproteins; factors of adherence to epithelial cells and penetrating system, pilus formation, conjugation, protein translocation and so on [
29,
30,
31,
32]. It is known that these secretion systems in rhizobia can affect the development of plant–microbe interactions and the host specificity of strains, as well as determine the number of nodules formed [
33].
3.2. Plant Nodulation Assays
The plant nodulation tests were performed on alfalfa, common vetch and red clover plants with the participation of the corresponding commercial strains and the isolates
M. japonicum Opo-235,
M. japonicum Opo-242,
Bradyrhizobium sp. Opo-243 or
M. kowhaii Ach-343. Mono- and co-inoculation of alfalfa plants showed that three out of four isolates (except for
M. kowhaii Ach-343) led to a significant increase in the number of nodules in variants of joint inoculation with the commercial strain
S. meliloti RCAM1750 (
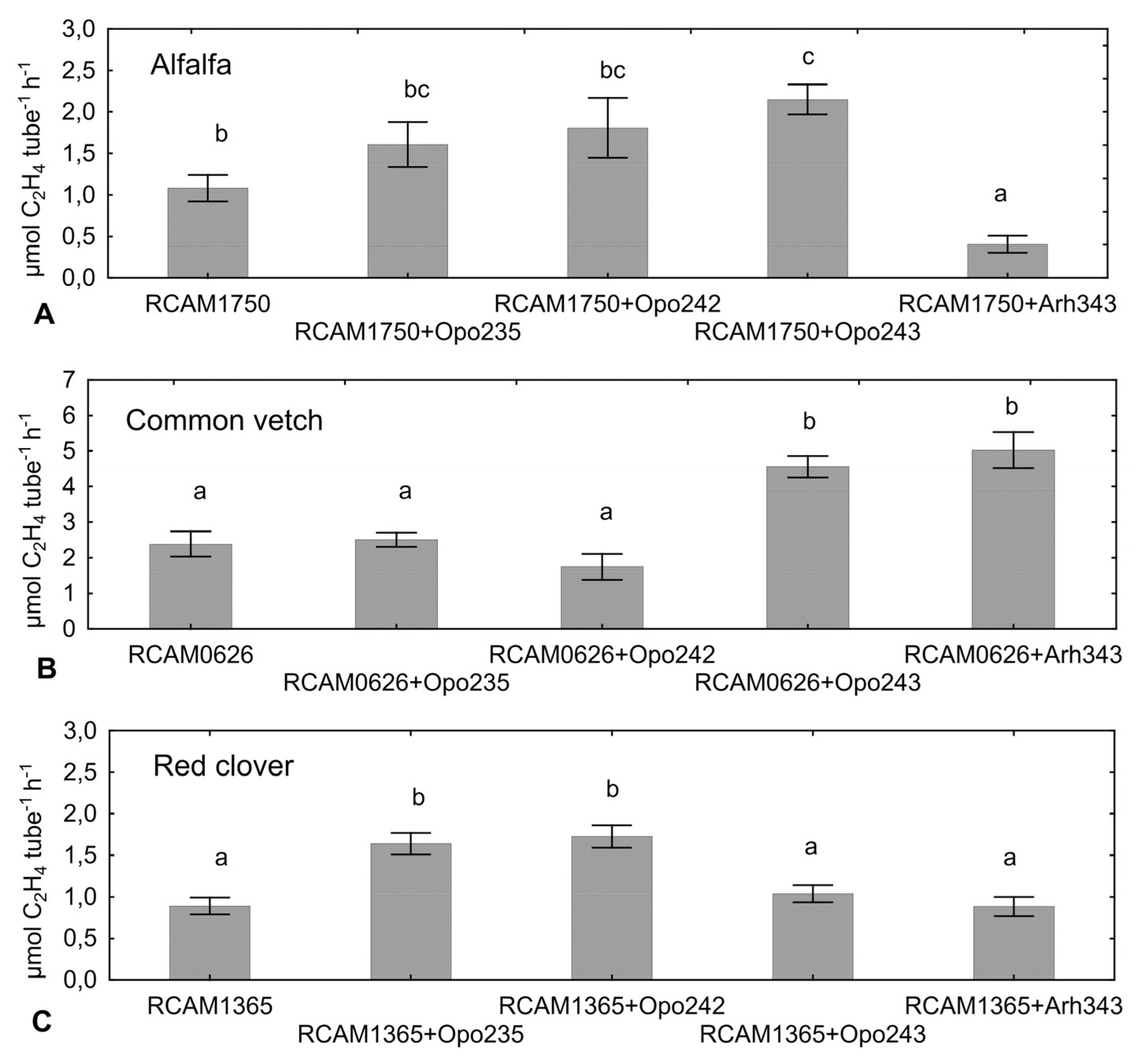
Table 6). In some cases, this was accompanied by an increase in the biomass of shoots and the total biomass of plants (roots and shoots). An enhanced total nitrogen-fixing activity was observed only when the strain
S. meliloti RCAM1750 was co-inoculated with
Bradyrhizobium sp. Opo-243 (
Figure 1), although the level of activity calculated per one nodule did not differ from the commercial strain (data not shown). After co-inoculation with
S. meliloti RCAM1750 and
M. kowhaii Ach-343, despite a significant decrease in nodule number and nitrogen-fixing activity, an increase in the biomass of roots, shoots and total plant was observed.
The isolate
M. japonicum Opo-235 co-inoculated with
S. meliloti RCAM1750 contributed to significant acceleration of the nodule formation in comparison (
Figure 2) and also led to the increase in plant biomass of alfalfa, although no increase in the total nitrogen-fixing activity was detected (
Table 6,
Figure 1).
The other two isolates
M. japonicum Opo-242 and
Bradyrhizobium sp. Opo-243 also contributed to the formation of more nodules. However,
M. japonicum Opo-242 had no effect on plant biomass, while
Bradyrhizobium sp. Opo-243 increased the biomass of alfalfa shoots. It was shown that the nodules in the variant
S. meliloti RCAM1750 +
M. japonicum Opo-242 were small, and among them there were many tumor-like structures (
Figure 3).
When the isolates
M. japonicum Opo-235 and
Bradyrhizobium sp. Opo-243 were used together with the strain
S. meliloti RCAM1750, almost all nodules had a typical elongated shape, although both isolates themselves formed a small number of abnormal structures on the alfalfa roots (
Figure 3). None of the isolates were detected in the alfalfa nodules and tumors either in mono-inoculation or in co-inoculation variants.
The data of plant nodulation experiment on mono- and co-inoculation of common vetch with the commercial strain
R. leguminosarum RCAM0626 and the isolates are shown in the
Table 7. Unlike alfalfa plants, the isolates themselves did not form nodules or tumors on the common vetch roots. Isolate
M. kowhaii Ach-343 increased nodule number when co-inoculated with
R. leguminosarum RCAM0626, while the isolate
M. japonicum Opo-242 had no effect on this parameter. The level of total nitrogen-fixing activity enhanced upon co-inoculation with
R. leguminosarum RCAM0626 and
Bradyrhizobium sp. Opo-243 or
M. kowhaii Ach-343 (
Figure 1). However, this was associated with the formation of a significantly larger number of nodules, but not with their activity per one nodule (data not shown). An increase in the nodule number and the level of nitrogen-fixing activity had practically no effect on the plant biomass (
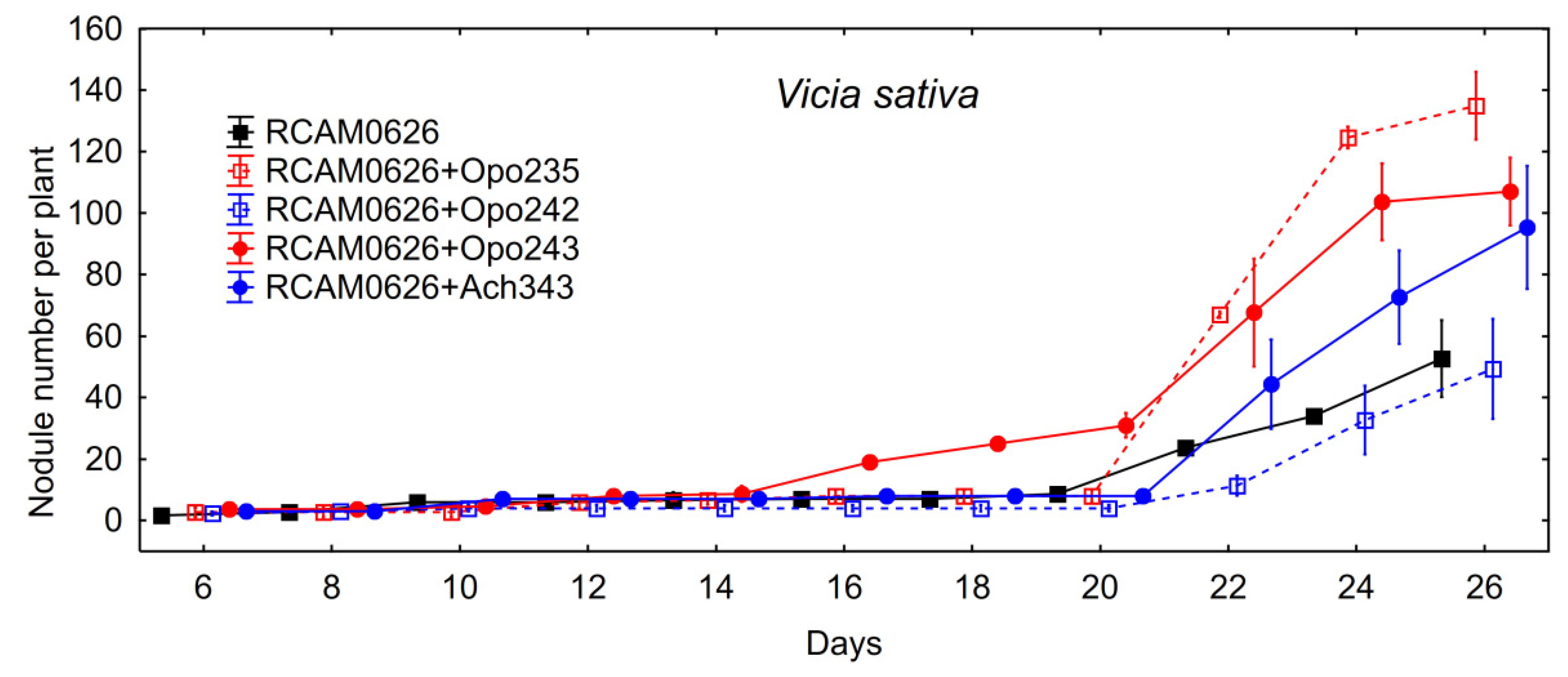
Table 7). It is possible that some unknown growth limiting factors were presented under model gnotobiotic conditions and a potential of such symbiosis for biomass production could be more pronounced under optimal conditions. The study of the nodulation rate on the common vetch roots showed that only
Bradyrhizobium sp. Opo-243 contributed to a significantly faster nodule formation compared to inoculation with
R. leguminosarum RCAM0626 (
Figure 4).
In the experiment on
T. pratense, the increase in the level of nitrogen-fixing activity was detected when
R. leguminosarum RCAM1365 was combined with
M. japonicum Opo-235 or
M. japonicum Opo-242 (
Figure 1). The level of activity calculated per one nodule in these two variants was also statistically significant increased (129.1 and 125.4 nmol C
2H
4 nodule
−1 h
−1, respectively), compared to mono-inoculation with the commercial strain RCAM1365 (66.2 nmol C
2H
4 nodule
−1 h
−1).
In this regard, it is important to note that in these two variants of co-inoculation the isolates were re-isolated from nodules (the data not presented). At the same time,
M. japonicum Opo-235 itself formed a small number of typical elongated nodules on the
T. pratense roots containing cells of this strain, whereas the isolate
M. japonicum Opo-242 formed tumors (
Figure 5) without bacteria inside them. Studying the nodulation rate on red clover did not reveal any variants of co-inoculation that differed from mono-inoculation with the commercial strain
R. leguminosarum RCAM1365 (data not shown).
No change in the nodule number was observed as compared to the commercial strain (
Table 8). It should be noted that the increase in the level of nitrogen-fixing activity did not lead to a significant increase in plant weight.
3.3. Comparison of the Genomic Data and Plant Nodulation Assays
We tried to explain the influence of the Baikal isolates on the efficiency of symbiosis with alfalfa, common vetch and red clover using a comparative genomic analysis of all strains that participated in plant nodulation experiments. Symbiotic and non-symbiotic genes that are absent in the commercial strains
S. meliloti RCAM1750,
R. leguminosarum RCAM0626 and
R. leguminosarum RCAM1365 but are present in the isolates are shown in
Table 9. Based on the analysis of the results obtained in the nodulation assay with alfalfa it can be assumed that the symbiotic genes
nodLZ, nolKL and
noeKL could act as the main accessory genes of the isolates, which contributed to an increase in the rate of nodulation and the efficiency of symbiosis with this plant species. The genes
nodL and
nodZ perform, respectively, 6-O-acetylation and fucosilation of the non-reducing end of the Nod factor (NF) and has been shown to be important for symbiosis of alfalfa and soybean with
Sinorhizobium strains [
34,
35,
36,
37]. The
noeK, noeL, and
nolK genes associated with the expression of the
nodZ gene are involved in the synthesis of the precursor of the fucosyl group from mannose [
37,
38,
39]. The gene
nolL carries out 4-O-acetylation of the fucose residue in NF, which arises as a result of the work of the gene
nodZ [
40]. Thus, all of the genes discussed above are responsible for the specificity of the plant–rhizobia interactions by NFs modification (acetylation and fucosilation). The
nodT gene, also found in the isolates but not in
S. meliloti RCAM1750 (
Table 9), encodes an outer membrane lipoprotein playing a role in the secretion of NFs [
41,
42].
However, a positive effect of the gene nodT on the formation of symbiosis upon joint inoculation with the commercial strain seems unlikely. We also do not focus on the nif and fix genes of isolates that are absent in the strain S. meliloti RCAM1750, since none of the isolates was detected in the alfalfa nodules and tumors either in mono-inoculation or in co-inoculation variants.
Genes of the isolates that were absent in the commercial strain
R. leguminosarum RCAM0626 are shown in
Table 9. Among them, the
nodPZ and
nolK genes could affect the rate of common vetch nodule formation, their total number and/or nitrogen-fixing activity. The
nodP gene is involved in the 6-O-sulfation of the reducing end of the NFs [
17]. It was known that NodP functioned in conjunction with NodQ, which synthesizes the donor of the sulfate group [
43]. It should be noted that the commercial strain
R. leguminosarum RCAM0626 has the
nodQ gene, although the importance of NFs sulfation for the common vetch has not been shown. The functions of
nodZ and
nolK genes have been described above. It is noteworthy that the
nolK gene is present only in the isolate
Bradyrhizobium sp. Opo-243, which alone contributed to the acceleration of nodulation on
V. sativa plants (
Figure 4). The
nodZ and
nolK genes were found by other authors in a number of
R. leguminosarum strains, e.g., in many pea nodulating rhizobia the NFs carry a fucosyl group, which may indicate the importance of this modification for the tribe Fabeae [
44]. Since we did not detect an increase in nitrogen-fixing activity per nodule in variants of co-inoculation of common vetch, we do not discuss the role of
nif and
fix genes that are present in isolates, although the strain
M. kowhaii Ach-343 was re-isolated from the nodules together with the commercial strain RCAM0626.
The nodulation experiment with red clover, unlike those with alfalfa and common vetch, showed an increase in the level of nitrogen-fixing activity per one nodule caused by co-inoculations. The additional
nif and
fix genes of
M. japonicum Opo-235 and
M. japonicum Opo-242 might contributed to this parameter when co-inoculated with
R. leguminosarum RCAM1365 (
Table 9). Among them,
nifZQXW genes associated with the synthesis of the Fe-S-cofactor, fixation of molybdenum and protection of nitrogenase from oxygen damage, as well as the
nifV gene, which is involved in the maturation of nitrogenase through the synthesis of homocitrate [
45,
46,
47,
48,
49]. The
fixHSQ genes are involved in the synthesis of components of the oxidase complex, which is associated with the membrane and allows cells to breathe at low oxygen concentrations [
20,
46]. The
fixKL genes, together with the
fixJT genes, determine the expression of genes necessary for growth under microaerophilic conditions [
20,
50].
It should be noted that, in our previous work devoted to the analysis of genetic complementation in strains isolated from the relict symbiotic systems, the genes indicated above have already come into view. Thus, the
nolK gene was mentioned in connection with the ability of the isolate
Bradyrhizobium sp. Opo-243 to increase the rate of nodulation on the relict legume
Oxytropis popoviana (host plant) when used with its paired strain
M. japonicum Opo-242 isolated from the same nodule [
3]. The presence of the complementary genes
nifQV and
fixJKL in
M. japonicum Opo-235 and
M. kowhaii Ach-343 was associated with an increase in the nitrogen-fixing activity of nodules on
Glycyrrhiza uralensis plants co-inoculated with both of these strains. Moreover, it was shown that isolates complementing each other for these genes were localized in the same nodule cells [
4].
The genes responsible for the synthesis of auxins and genes of the T6SS secretion system can also positively influence the process of nodule formation. The presence of these genes in
M. kowhaii Ach-343 apparently caused a significant increase in biomass of alfalfa plants with a decrease in the number of root nodules (
Table 6). The influence of secretion systems on the formation of plant–microbial relationships was previously shown on the example of two strains (
M. loti 582 and
M. huakuii 583) isolated from the endemic of Kamchatka
Oxytropis kamtschatica [
16]. These strains possessed the T3SS and T6SS genes and formed two types of nodules (typical elongated and abnormal rounded) on roots of the host plant after mono-inoculations. It was suggested that this phenomenon can be regulated by two different nodulation strategies: (1) NF dependent, by the infection thread formation, and (2) NF independent, by the direct penetration of bacteria through the host cell wall. However, it certainly cannot be ruled out that other genetic factors, which are not considered here, could influence the process of the symbiosis formation.
The mechanisms of the rhizobial synergy described above, which are associated with the genetic complementation of co-microsymbionts, can be based on interbacterial interactions involving membrane and secreted proteins. In addition, some affecting proteins can enter the environment after cell lysis. The phenomenon of bacterial synergy in the context of accelerating and increasing the efficiency of nodulation is most easily explained by the modification of NFs that occurs outside the cells or on their surface. At the same time, the cooperation between co-microsymbionts most likely depends on their spatial arrangement (rhizosphere, rhizoplane or nodule), and in the case of localization of bacteria in one nodule or in the same plant cells, the level of interaction should be higher.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}