
Biological Activity of Root Extract Decalepis hamiltonii (Wight & Arn) against Three Mosquito Vectors and Their Non-Toxicity against the Mosquito Predators

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Plant Material
2.2. Preparation of Extraction
2.3. GC-MSAnalysis
2.4. Mosquito Culture
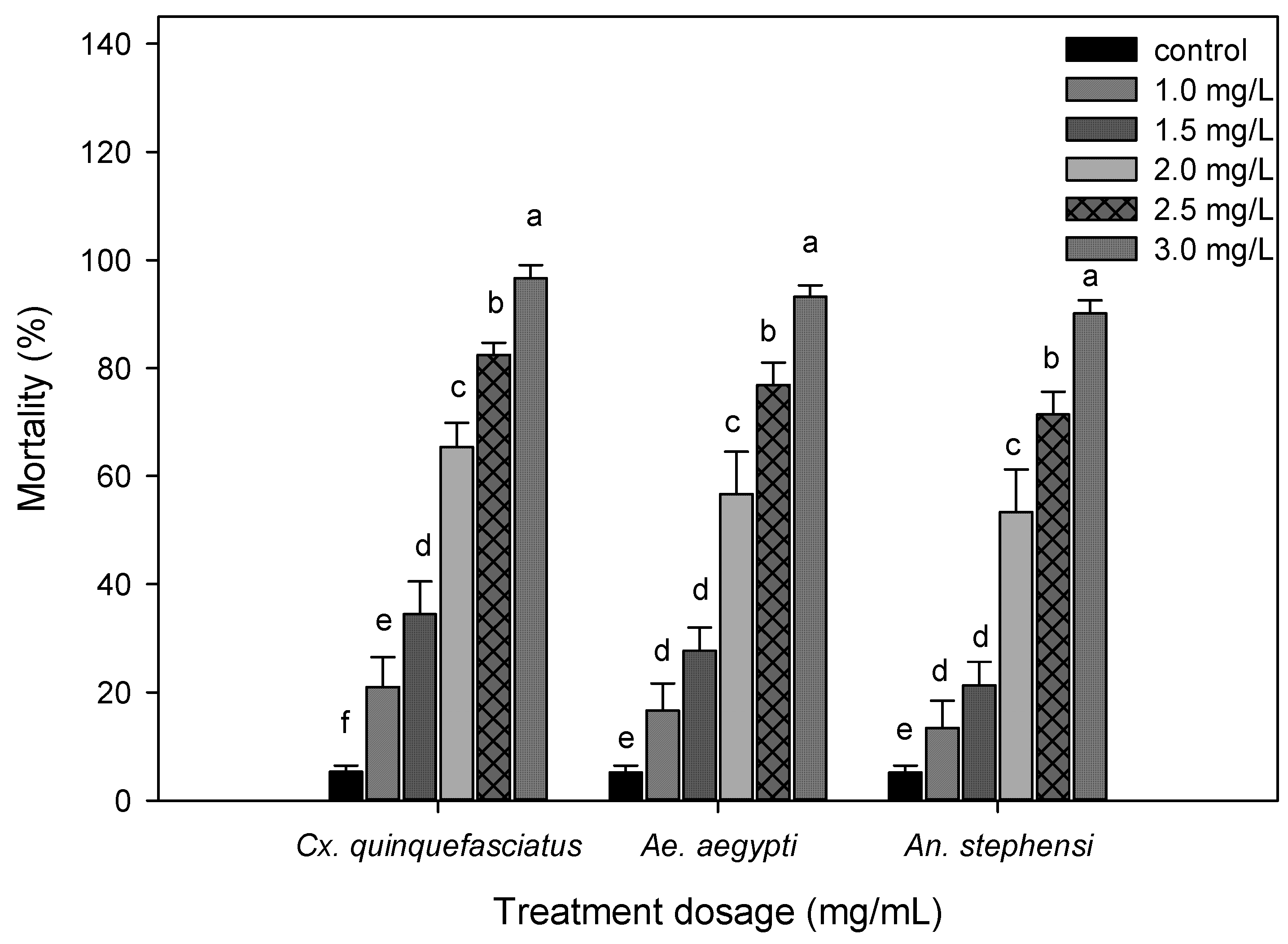
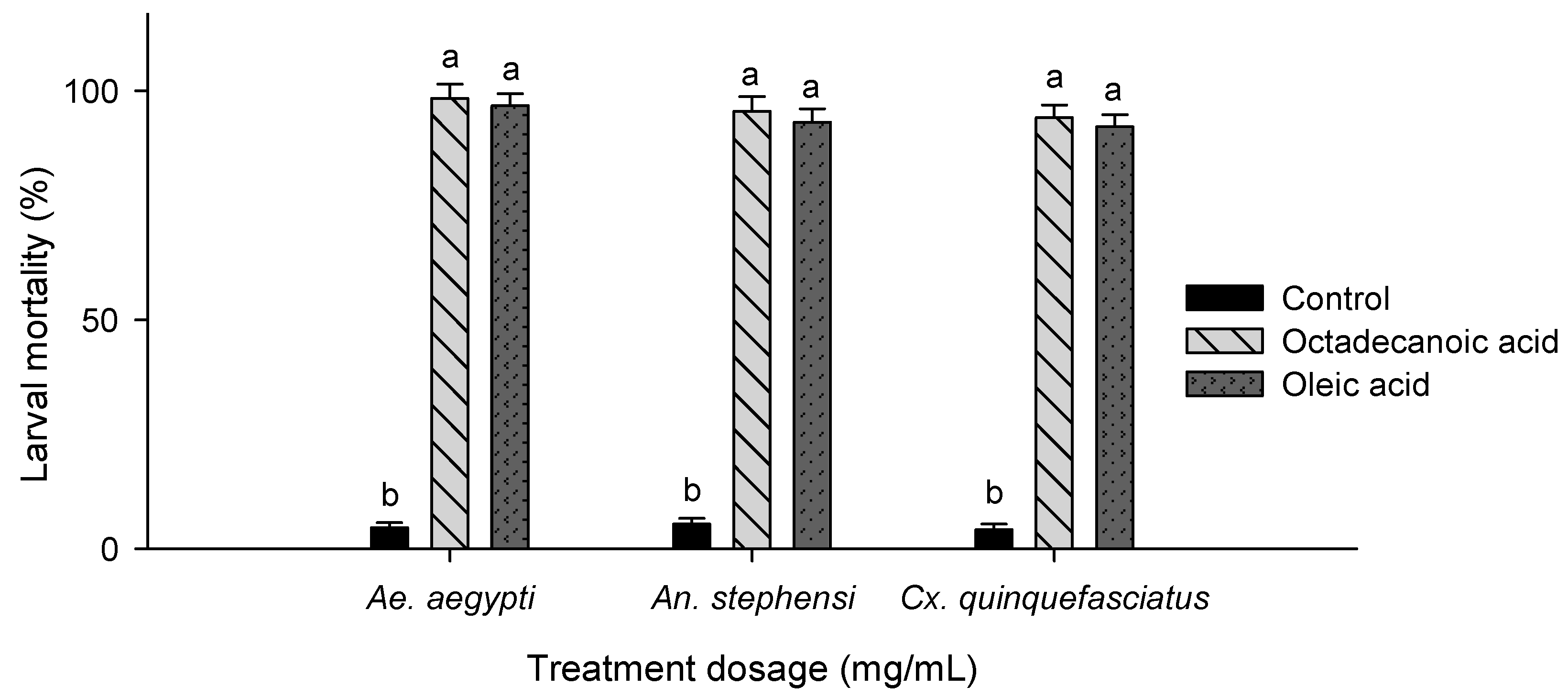
2.5. Larvicidal Bioassay
2.6. Chemicals
2.7. Enzyme Assay
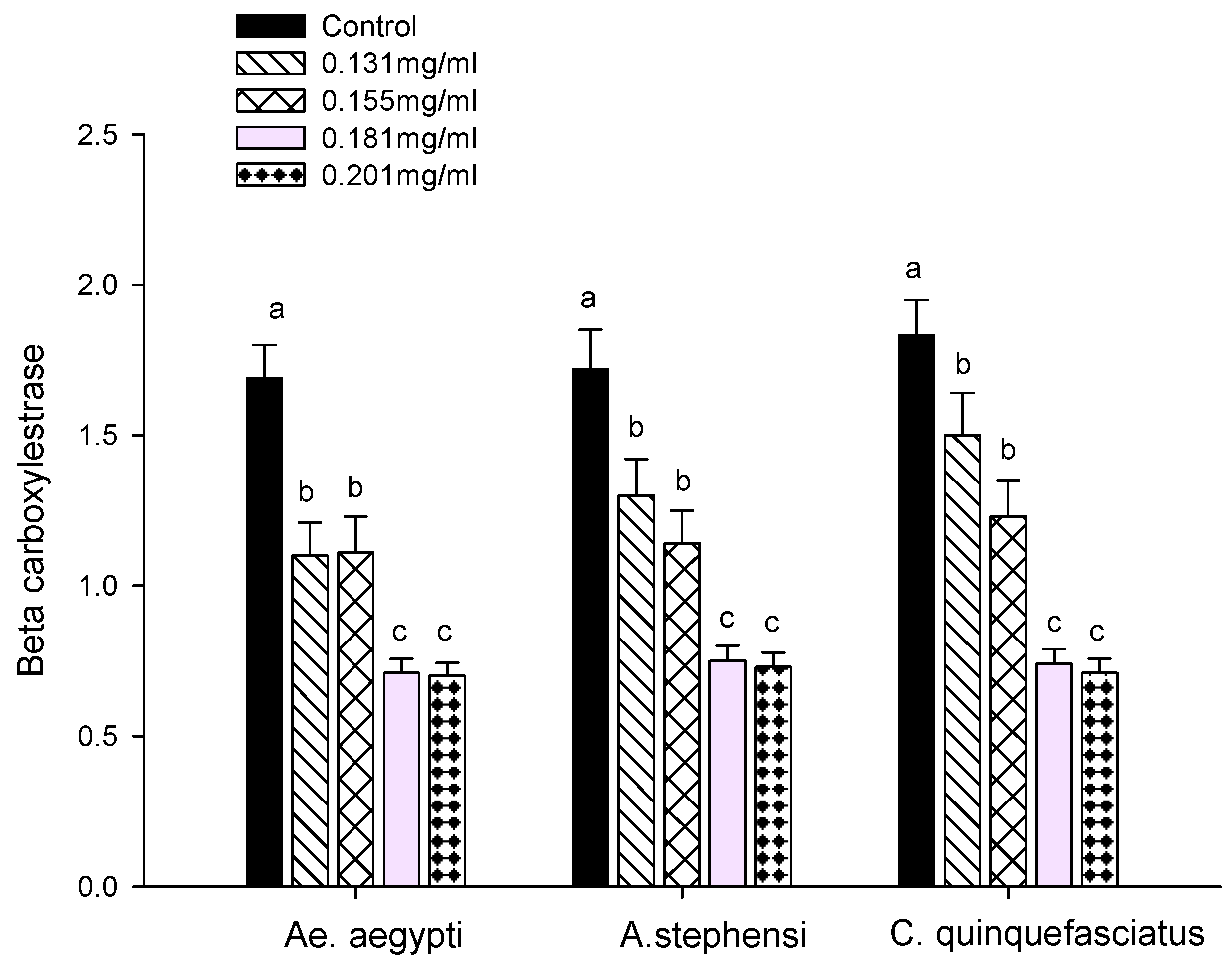
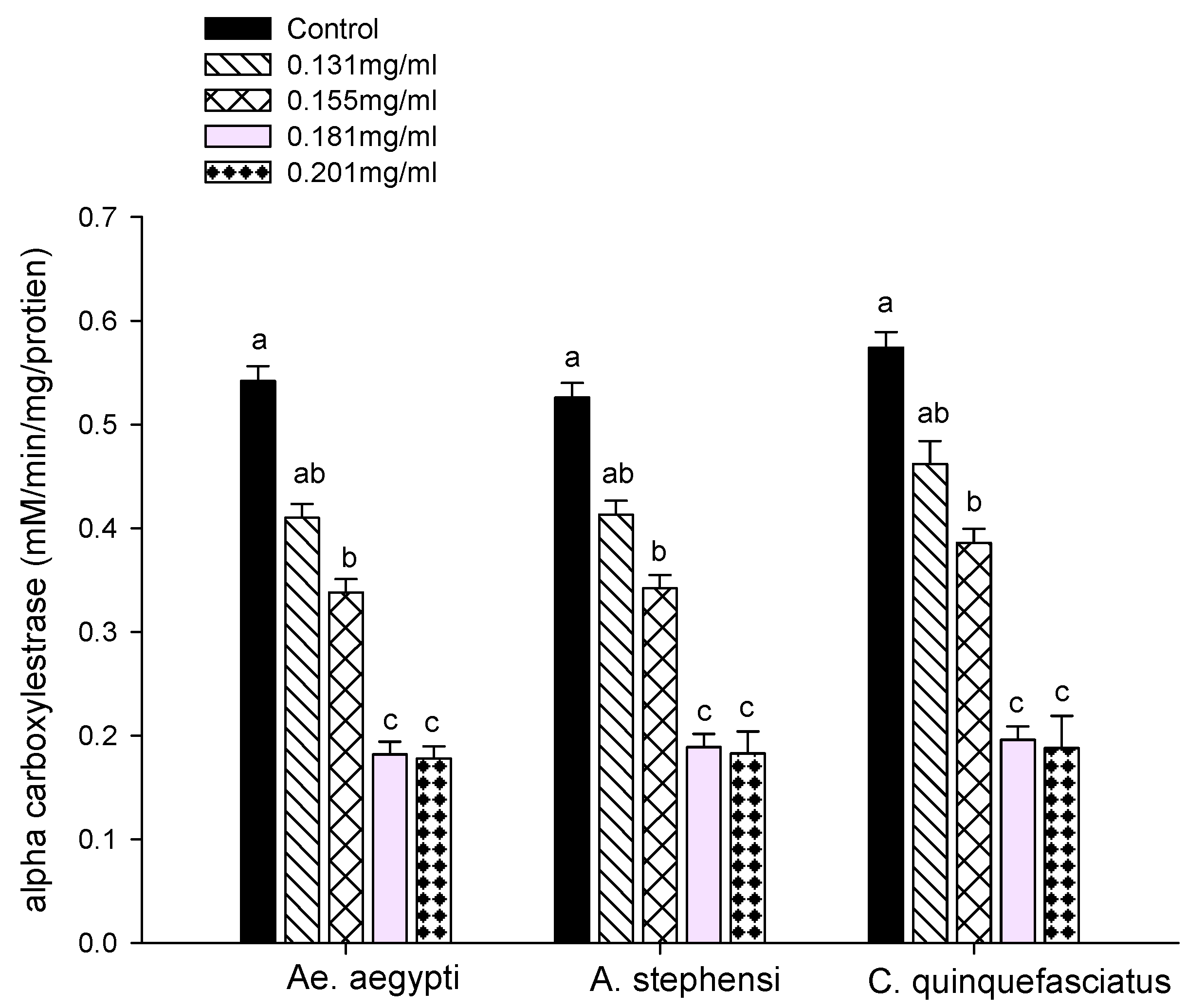
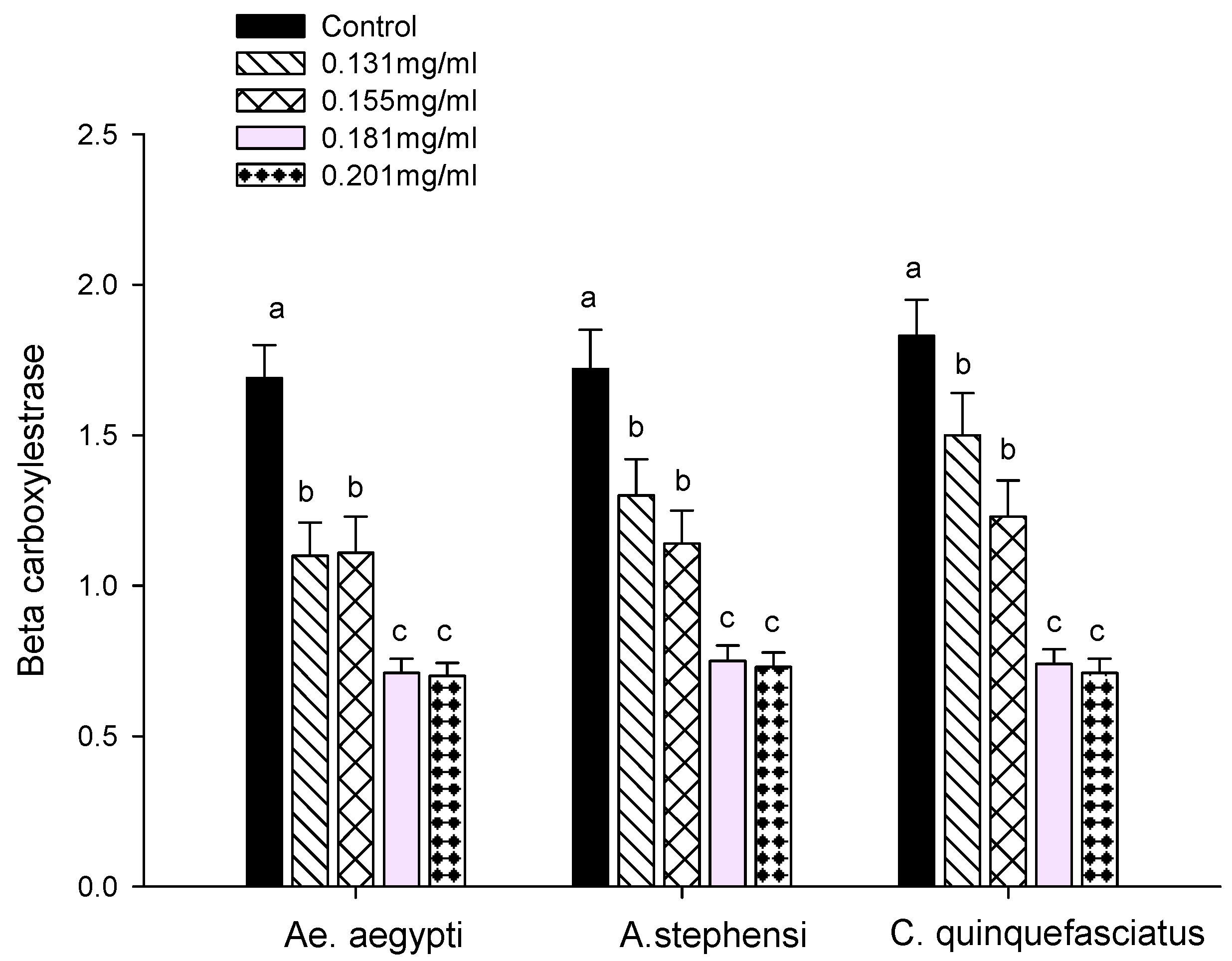
2.7.1. Carboxylesterase Assays
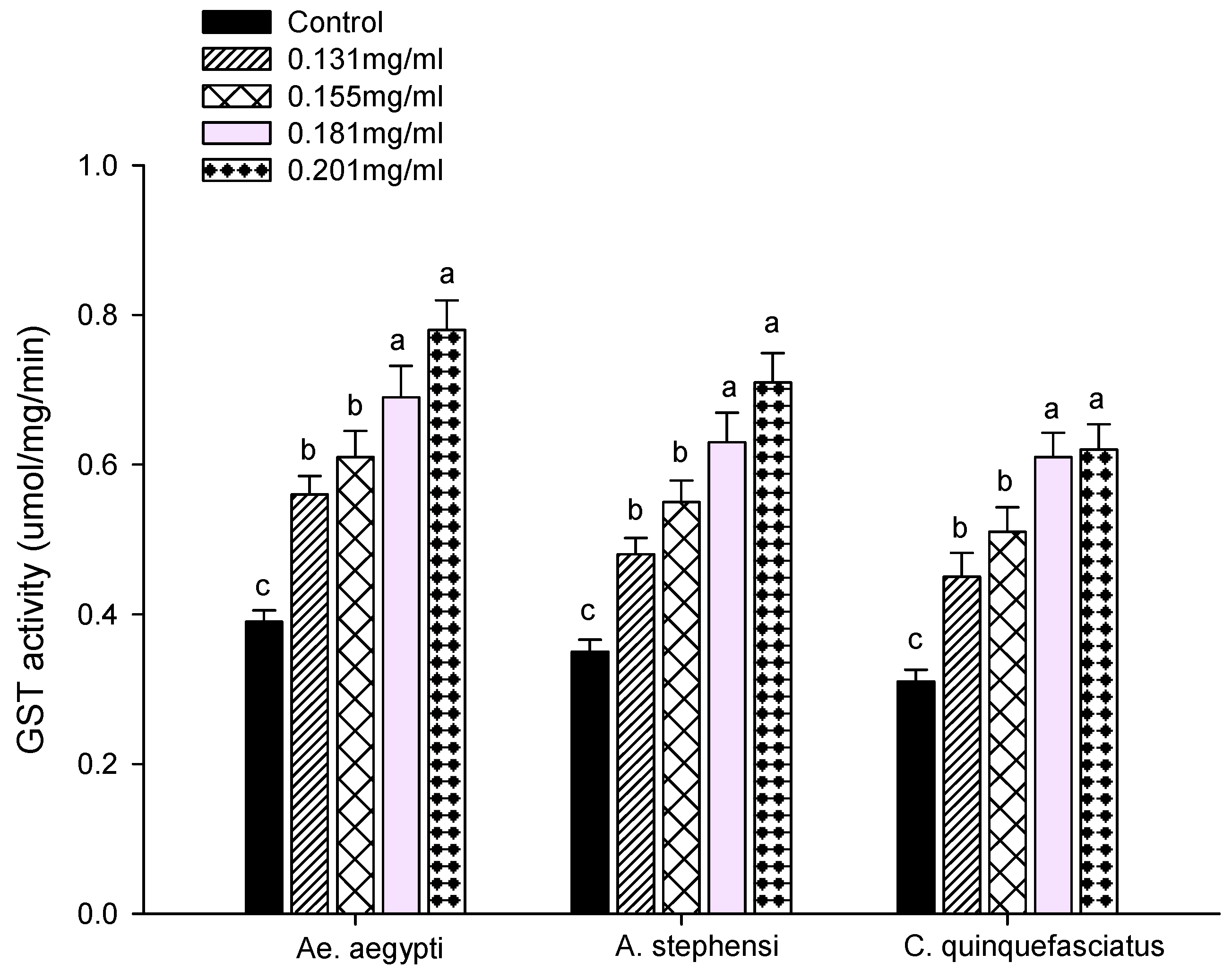
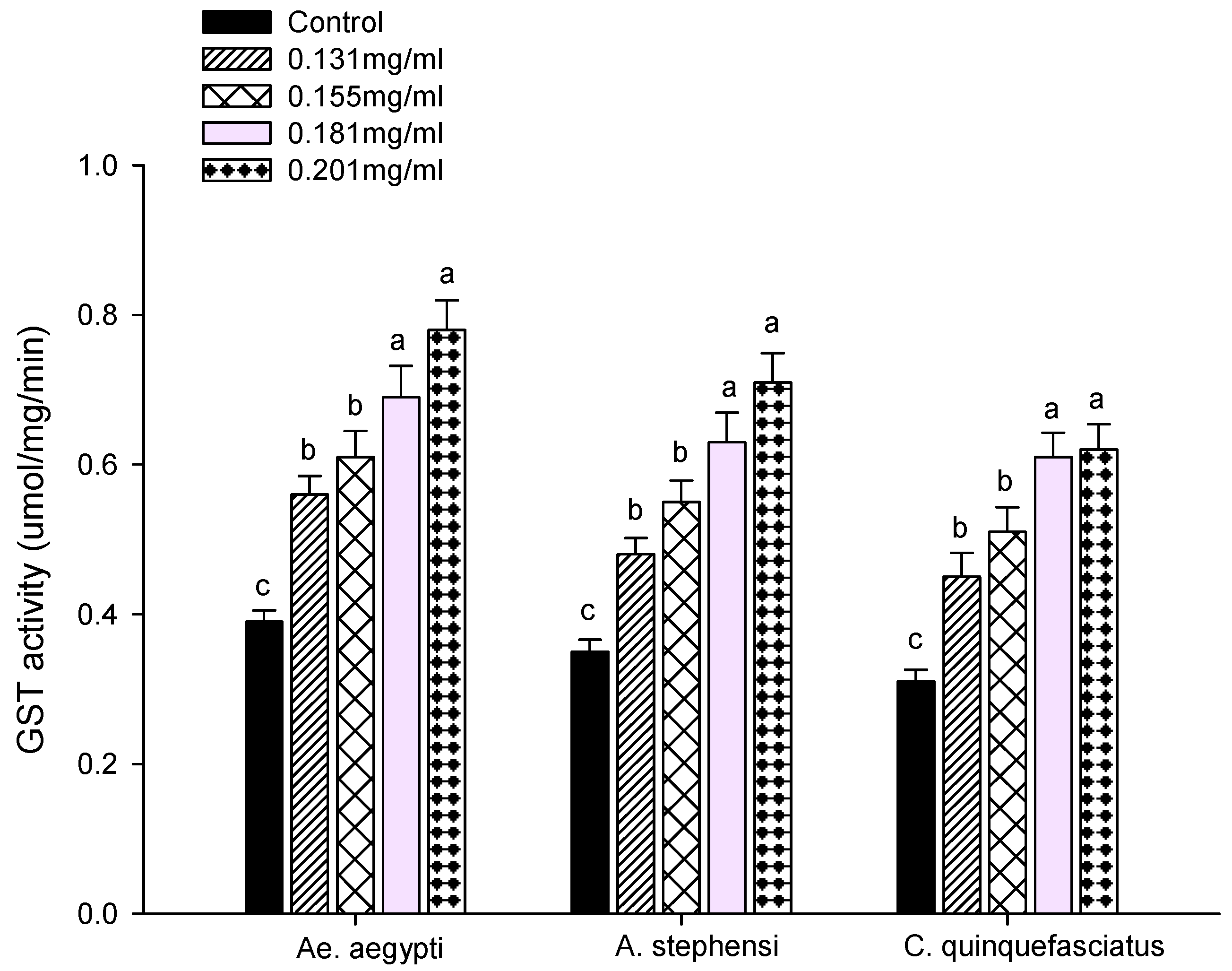
2.7.2. Glutathione S-Transferase Assay
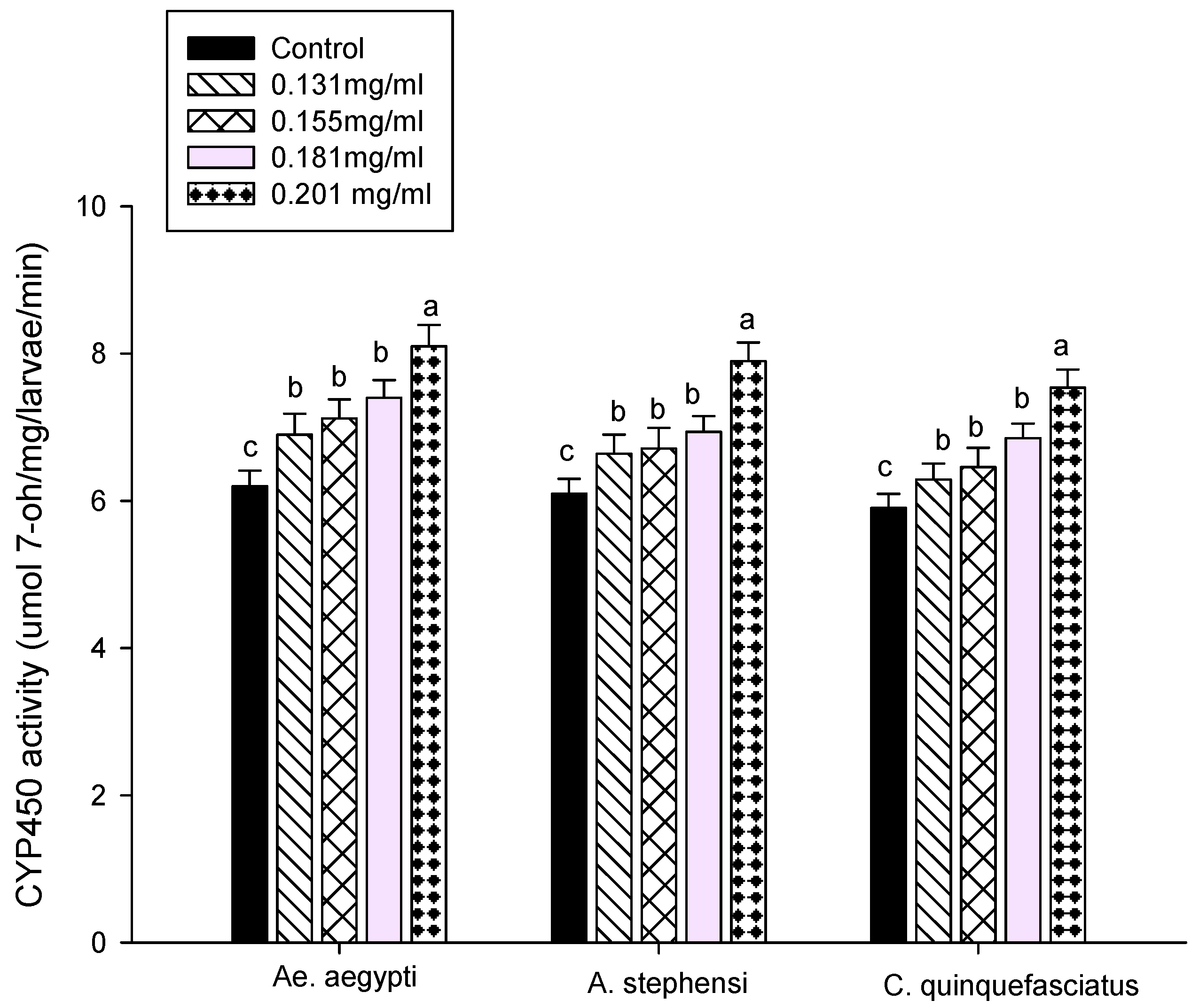
2.7.3. Cytochrome P450 Assay
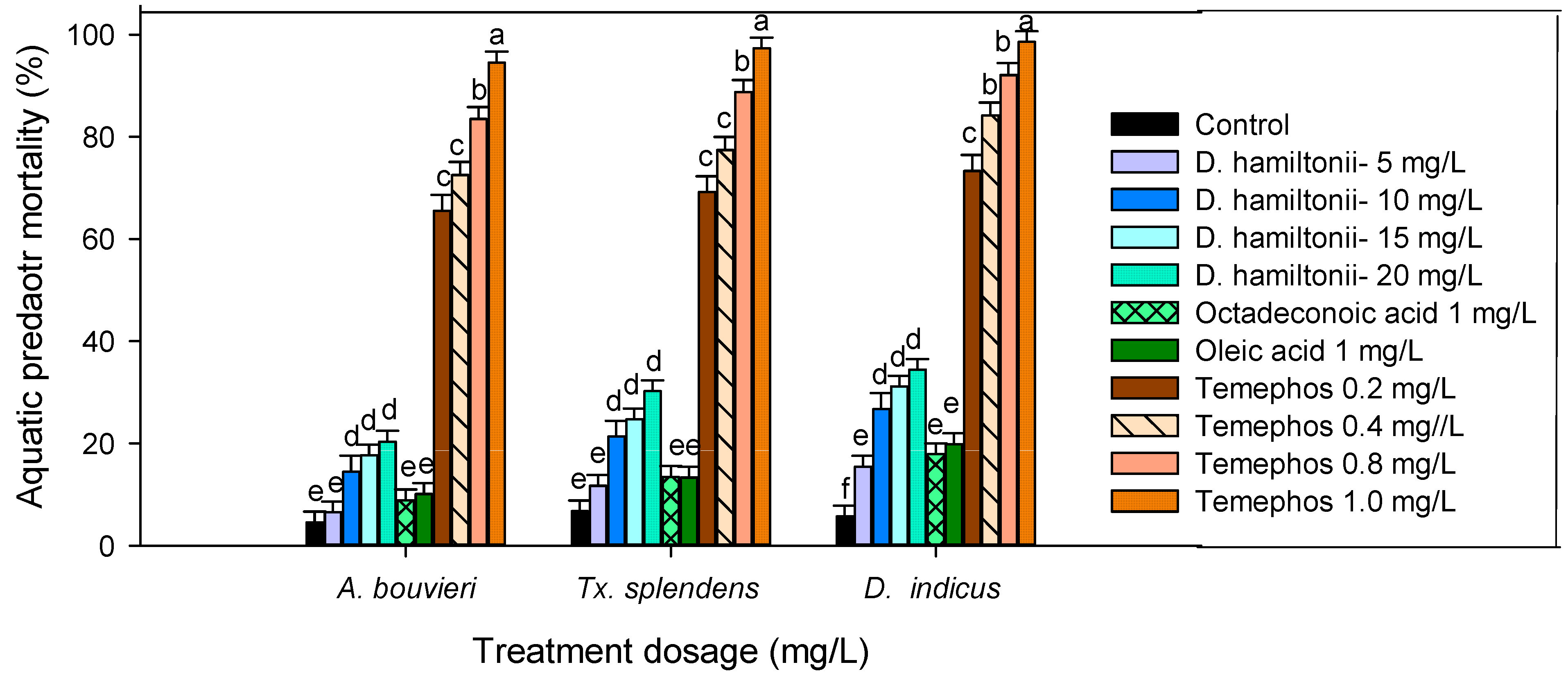
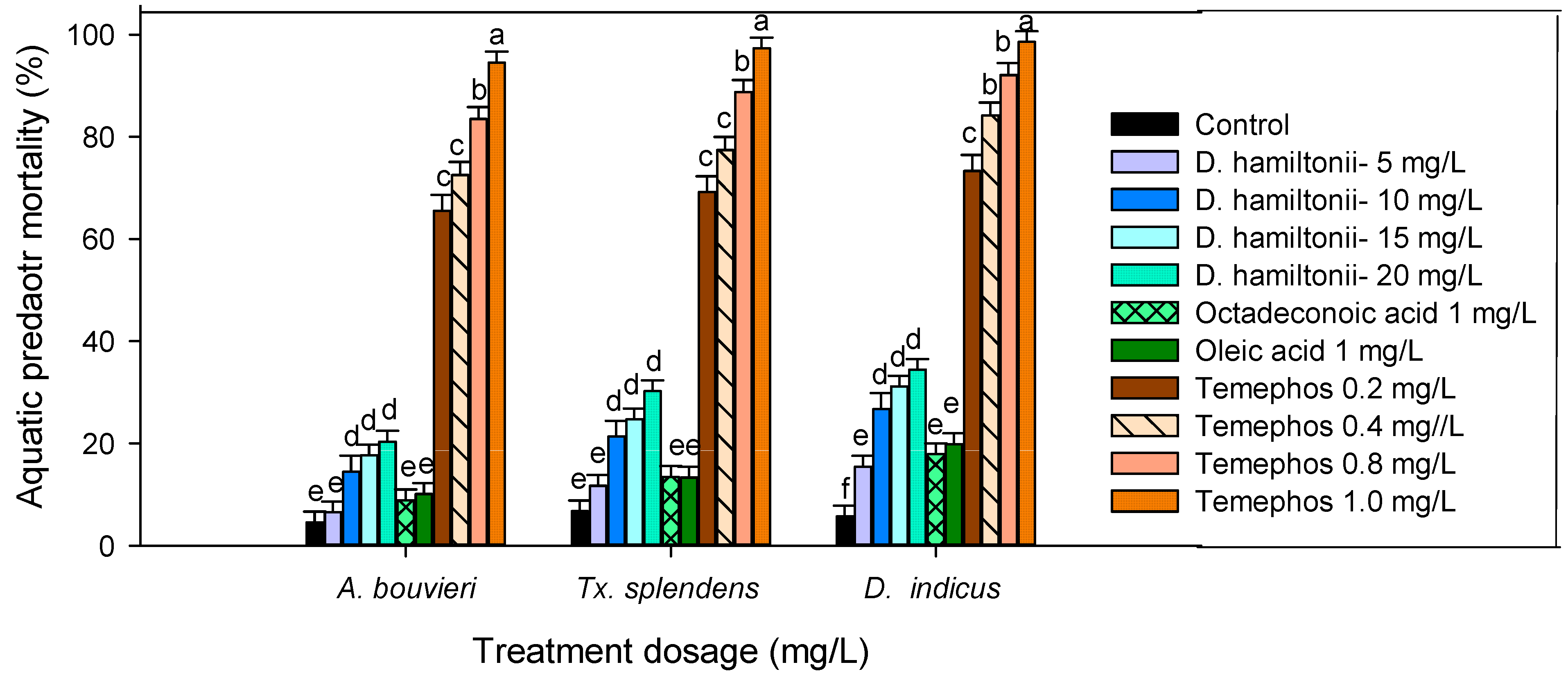
2.8. Non-Target Toxicity against Mosquito Predaotrs
2.9. Data Analysis
3. Results
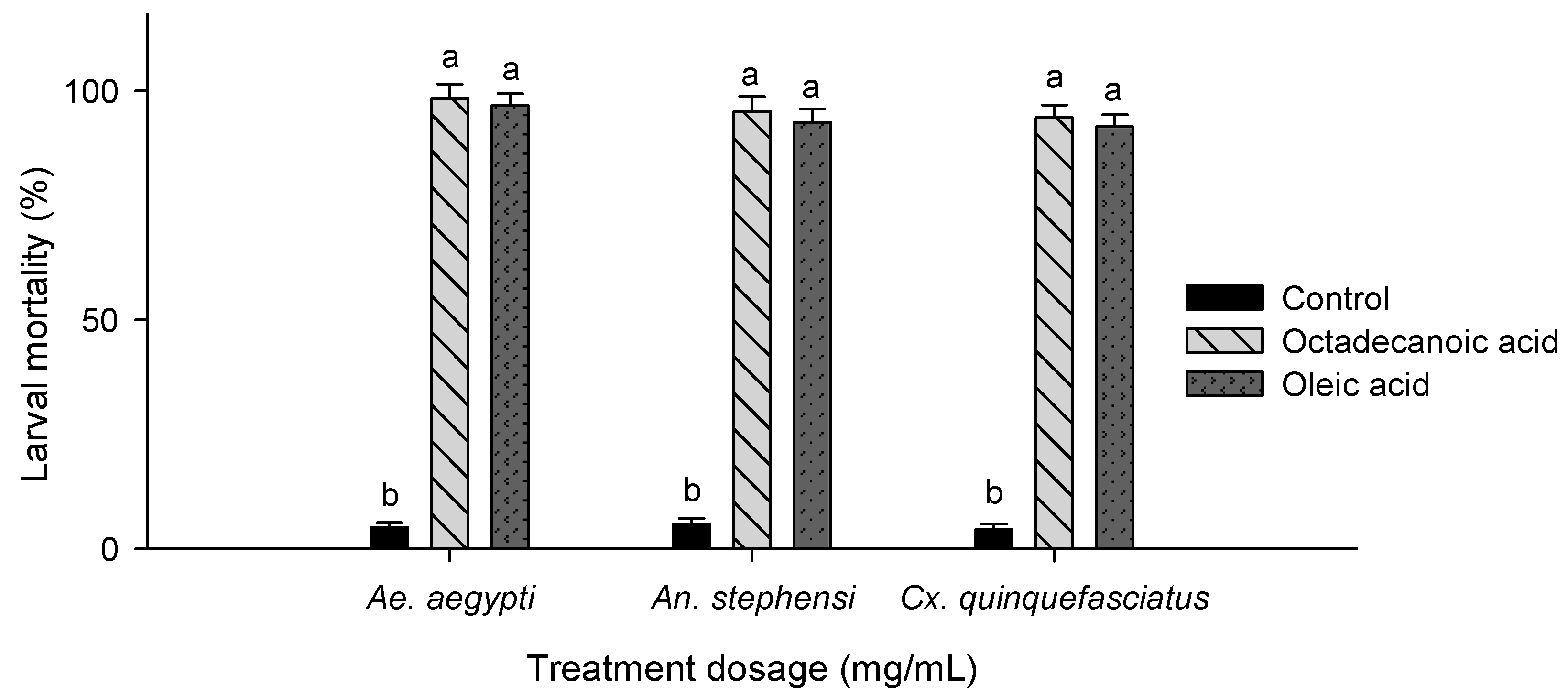
3.1. GC-MS Analysis
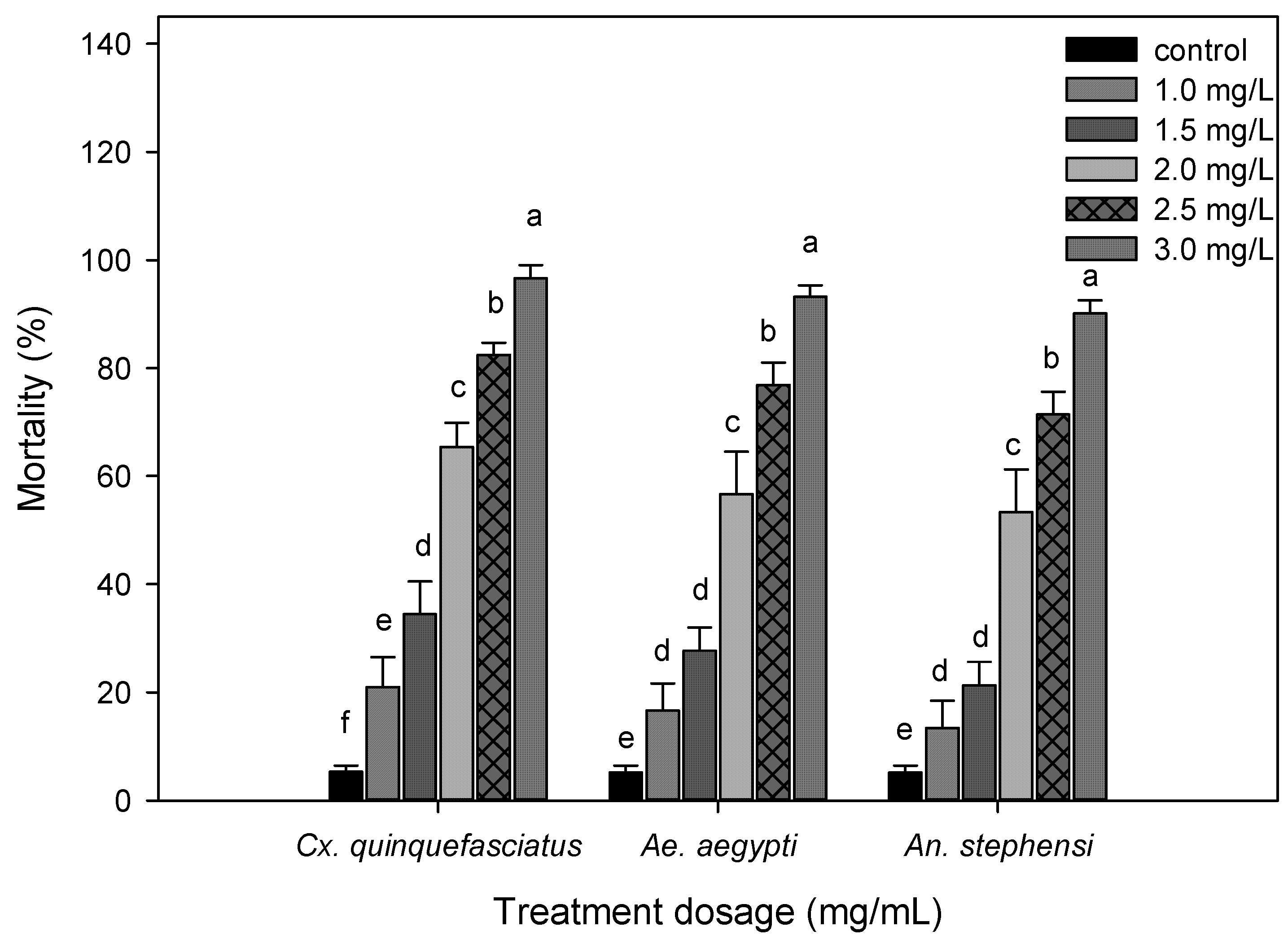
3.2. Larvicidal Activity
3.3. Enzyme Activity
3.4. Non-Target Activity of Mosquito Predators
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kovendan, K.; Murugan, K.; Shanthakumar, S.P.; Vincent, S. Evaluation of larvicidal and pupicidal activity of Morinda citrifolia L. (Noni) (Family: Rubiaceae) against three mosquito vectors. Asian Pac. J. Trop. Dis. 2012, 2, S362–S369. [Google Scholar] [CrossRef]
- James, A.A. Mosquito molecular genetics: The hands that feed bite back. Science 1992, 257, 37–38. [Google Scholar] [CrossRef]
- Gubler, D.J. Resurgent vector borne disease as a global health problem. Emerg. Infect. Dis. 1998, 4, 442–450. [Google Scholar] [CrossRef]
- Peng, Z.; Yang, J.; Wang, H. Simons FER Production and characterization of monoclonal antibodies to two new mosquitoes A. aegypti salivary proteins. Insect Biochem. Mol. Biol. 1999, 29, 909–914. [Google Scholar] [CrossRef]
- Hales, S.; De Wet, N.; Maindonald, J.; Woodward, A. Potential effect of population and climate changes on global distribution of dengue fever: An empirical model. Lancet 2002, 360, 830–834. [Google Scholar] [CrossRef]
- Maillard, M.; Marston, A.; Hostettman, K. Human Medicinal Agents from Plants; Balandrin, M., Ed.; American Chemical Society: Washington, DC, USA, 1993. [Google Scholar]
- Ali, M.S.; Ravikumar, S.; Beula, J.M.; Anuradha, V.; Yogananth, N. Insecticidal compounds from Rhizophoraceae mangrove plants for the management of dengue vector Aedes aegypti. J. Vector Borne Dis. 2014, 51, 106–114. [Google Scholar]
- World Health Organization. Lymphatic Filariasis. 2012. Available online: http://www.who.int/mediacentre/factsheets/fs102/en/ (accessed on 19 May 2021).
- Syedali, M.; Ravikumar, S.; Beula, J.M. Bioactivity of seagrass against the dengue fever mosquito A. aegypti larvae. Asian Pac. J. Trop. Biomed. 2012, 2, 570–573. [Google Scholar]
- Ezhumalai, P.; Arokiyaraj, C.; Ramanibai, R. Annona muricata: An alternate mosquito control agent with special reference to inhibition of detoxifying enzymes in Aedes aegypti. Ecotoxicol. Environ. Saf. 2020, 189, 110050. [Google Scholar]
- Rahuman, A.A.; Bagavan, A.; Kamaraj, C.; Saravanan, E.; Zahir, A.A.; Elango, G. Efficacy of larvicidal botanical extracts against C. quinquefasciatus Say (Diptera: Culicidae). Parasitol. Res. 2009, 104, 1365–1372. [Google Scholar] [CrossRef]
- Bagavan, A.; Kamaraj, C.; Rahuman, A.A.; Elango, G.; Zahir, A.A.; Pandiyan, G. Evaluation of larvicidal and nymphicidal potential of plant extracts against An. subpictus Grassi, Cx. tritaeniorhynchys Giles and Aphis gossypii Glover. Parasitol. Res. 2009, 104, 1109–1117. [Google Scholar] [CrossRef]
- Shahi, M.; Hanafi-Bojd, A.A.; Iranshahi, M.; Vatandoost, H.; Hanafi-Bojd, M.Y. Larvicidal efficacy of latex and extract of Calotropis procera (Gentianales: Asclepiadaceae) against C. quinquefasciatus and A. stephensi (Diptera: Culicidae). J. Vector Borne Dis. 2010, 47, 185–188. [Google Scholar]
- Isman, M.B. Neem and other botanical insecticides: Barriers to commercialization. Phytoparasitica 1999, 25, 339–344. [Google Scholar] [CrossRef]
- Shaalan, E.A.S.; Canyonb, D.; Younesc, M.W.F.; Abdel-Wahaba, H.; Mansoura, A.H. A review of botanical phytochemicals with mosquitocidal potential. Environ. Int. 2005, 3, 1149–1166. [Google Scholar] [CrossRef]
- Senthil-Nathan, S. A Review of Resistance Mechanisms of Synthetic Insecticides and Botanicals, Phytochemicals, and Essential Oils as Alternative Larvicidal Agents against Mosquitoes. Front. Physiol. 2020, 15, 1591. [Google Scholar] [CrossRef]
- World Health Organization. Report of the WHO Informal Consultation on the Evaluation and Testing of Insecticides; CTD/WHO PES/IC/96.1; WHO: Geneva, Switzerland, 1996. [Google Scholar]
- Maheswaran, R.; Ignacimuthu, S. A novel biopesticide PONNEEM to control human vector mosquitoes Anopheles stephensi L. and Culex quinquefasciatus Say. Environ.Sci. Pollut. Res. 2015, 22, 13153–13166. [Google Scholar] [CrossRef]
- Sadek, M.M. Antifeedant and toxic activity of Adhatoda vasica leaf extract against Spodoptera littoralis (Lep., Noctuidae). J. Appl. Entomol. 2003, 127, 396–404. [Google Scholar] [CrossRef]
- Selin-Rani, S.; Senthil-Nathan, S.; Revathi, K.; Chandrasekaran, R.; Thanigaivel, A.; Vasantha-Srinivasan, P.; Ponsankar, A.; Edwin, E.; Pradeepa, V. Toxicity of Alangium salvifolium Wang chemical constituents against the tobacco cutworm Spodoptera litura Fab. Pest. Biochem. Physiol. 2016, 126, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Oliveria, P.V.; Ferreira, J.C., Jr.; Moura, F.S.; Lima, G.S.; de Oliveira, F.M.; Oliveria, P.E.S.; Conserva, L.M.; Giulietti, A.M.; Lemos, R.L. Larvicidal activity of 94 extracts from ten plant species of northeastern of Brazil against Aedes aegypti L. (Diptera: Culicidae). Parasitol. Res. 2010, 107, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.S.; Lin, C.Y.; Chung, M.J.; Liu, Y.H.; Huang, C.G.; Chang, S.T. Larvicidal activities of wood and leaf essential oils and ethanolic extracts from Cunninghamia konishii Hayata against the dengue mosquitoes. Ind. Crop. Prod. 2013, 47, 310–315. [Google Scholar] [CrossRef]
- Rajashekar, Y.; Gunasekaran, N.; Shivanandappa, T. Insecticidal activity of the root extract of Decalepis hamiltonii against stored product insect pests and its application in grain protection. J. Food Sci. Technol. 2010, 47, 310–314. [Google Scholar] [CrossRef] [Green Version]
- Rajashekar, Y.; Rao, L.J.M.; Shivanandappa, T. Decaleside: A new class of natural insecticide targeting tarsal gustatory sites. Naturwissenschaften 2012, 99, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Amala, K.; Ganesan, R.; Karthi, S.; Senthil-Nathan, S.; Chellappanidian, M.; Krutmunag, P.; Radhakrishnan, N.; Moham-mad, F.; Ponsankar, A.; Vasantha-Srinivasan, P. Larval and gut enzyme toxicity of n-hexane extract Epaltes pygmaea DC. Against the arthropod vectors and its non-toxicity against aquatic predator. Toxin Rev. 2020. [Google Scholar] [CrossRef]
- Srinivasan, D.; Nathan, S.; Suresh, T.; Perumalsamy, P.L. Antimicrobial activity of certain Indian medicinal plants used in folkloric medicine. J. Ethnopharmacol. 2001, 74, 217–220. [Google Scholar] [CrossRef]
- World Health Organization. WHO Informal Consultation on the Evaluation on the Testing of Insecticides; CTD/WHO PES/IC/96.1; WHO: Geneva, Switzerland, 2020; p. 69. [Google Scholar]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Thanigaivel, A.; Senthil-Nathan, S.; Vasantha-Srinivasan, P.; Edwin, E.S.; Ponsankar, A.; Selin-Rani, S.; Pradeepa, V.; Chellappandian, M.; Kalaivani, K.; Abdel-Megeed, A.; et al. Chemicals isolated from Justicia adhatoda Linn reduce fitness of the mosquito. Arch. Insect Biochem. Physiol. 2017, 94, e21384. [Google Scholar] [CrossRef] [Green Version]
- Karthi, S.; Uthirarajan, K.; Manohar, V.; Venkatesan, M.; Chinnaperumal, K.; Vasantha-Srinivasan, P.; Krutmuang, P. Larvicidal enzyme inhibition and repellent activity of red mangrove Rhizophora mucronata (Lam.) leaf extracts and their biomolecules against three medically challenging arthropod vectors. Molecules 2020, 25, 3844. [Google Scholar] [CrossRef]
- Ponsakar, A.; Sahayaraj, K.; Senthil-Nathan, S.; Vasantha-Srinivasan, P.; Karthi, S.; Thanigaivel, A.; Petchidurai, G.; Madasamy, M.; Hunter, W.B. Toxicity and developmental effect of cucurbitacin E from Citrullus colocynthis L. (Cucurbitales: Cucurbitaceae) against Spodoptera litura Fab. and a non-target earthworm Eisenia fetida Savigny. Environ. Sci. Pollut. Res. 2020, 27, 23390–23401. [Google Scholar] [CrossRef]
- Thanigaivel, A.; Chandrasekaran, R.; Revathi, K.; Nisha, S.; Sathish-Narayanan, S.; Kirubakaran, S.A.; Senthil-Nathan, S. Larvicidal efficacy of Adhatoda vasica (L.) Nees against the bancroftian filariasis vector Culex quinquefasciatus Say and dengue vector Aedes aegypti L. in in vitro condition. Parasitol. Res. 2012, 110, 1993–1999. [Google Scholar] [CrossRef]
- Chellappandian, M.; Senthil-Nathan, S.; Vasantha-Srinivasan, P.; Karthi, S.; Thanigaivel, A.; Kalaivani, K.; Sivanesh, H.; Stanley-Raja, V.; Chanthini, K.M.; Shyam-Sundar, N. Target and non-target botanical pesticides effect of Trichodesma indicum(Linn) R. Br. and their chemical derivatives against the dengue vector, Aedes aegypti L. Environ. Sci. Pollut. Res. 2019, 26, 16303–16315. [Google Scholar] [CrossRef]
- Thanigaivel, A.; Vasantha-Srinivasan, P.; Senthil-Nathan, S.; Edwin, E.S.; Ponsankar, A.; Chellappandian, M.; Selin-Rani, S.; Lija-Escaline, J.; Kalaivani, K. Impact of Terminalia chebula Retz. against Aedes aegypti L. and non-target aquatic predatory in-sects. Ecotoxicol. Environ. Saf. 2016, 137, 210–217. [Google Scholar] [CrossRef]
- Senthil-Nathan, S. A review of bio pesticides and their mode of action against insect pests. In Environmental Sustainability—Role of Green Technologies; Springer: Berlin/Heidelberg, Germany, 2015; pp. 49–63. [Google Scholar]
- Senthil-Nathan, S. Physiological and biochemical effect of Neem and other Meliaceae plants secondary metabolites against Lepidopteran insects. Front. Physiol. 2013, 4, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Lija-Escaline, J.; Senthil-Nathan, S.; Thanigaivel, A.; Pradeepa, V.; Vasantha-Srinivasan, P.; Ponsankar, A.; Edwin, E.; Selin-Rani, S.; Abdel-Megeed, A. Physiological and biochemical effects of chemical constituents from Piper nigrum Linn (Piperaceae) against the dengue vector Aedes aegypti Liston (Diptera: Culicidae). Parasitol. Res. 2015, 114, 4239–4249. [Google Scholar] [CrossRef]
- Vasantha-Srinivasan, P.; Senthil-Nathan, S.; Thanigaivel, A.; Edwin, E.S.; Ponsankar, A.; Selin-Rani, S.; Pradeepa, V.; Sakthi-Bhagavathy, M.; Kalaivani, K.; Hunter, W.B.; et al. Developmental response of Spodoptera litura Fab. to treatments of crude volatile oil from Piper betle L. and evaluation of toxicity to earthworm, Eudrilus eugeniae Kinb. Chemosphere 2016, 155, 336–347. [Google Scholar] [CrossRef]
- Edwin, E.; Vasantha-Srinivasan, P.; Senthil-Nathan, S.; Thanigaivel, A.; Ponsankar, A.; Selin-Rani, S.; Kalaivani, K.; Hunter, W.B.; Duraipandiyan, V.; Al-Dhabi, N.A. Effect of andrographolide on phosphatases activity and cytotoxicity against Spodoptera litura. Invert. Surv. J. 2016, 13, 153–163. [Google Scholar]
- Vasantha-Srinivasan, P.; Chellappandian, M.; Senthil-Nathan, S.; Ponsankar, A.; Thanigaivel, A.; Karthi, S.; Edwin, E.-S.; Selin-Rani, S.; Kalaivani, K.; Maggi, F.; et al. A novel herbal product based on Piper betle and Sphaeranthus indicus essential oils: Toxicity, repellent activity and impact on detoxifying enzymes GST and CYP450 of Aedes aegypti Liston (Diptera: Culicidae). J. Asia Pac. Entomol. 2018, 21, 1466–1472. [Google Scholar] [CrossRef]
- Thanigaivel, A.; Chanthini, M.; Karthi, S.; Vasantha-Srinivasan, P.; Ponsakar, A.; Sivanesh, H.; Stanley-Raja, V.; Shyam-Sundar, N.; Narayanan, K.R.; Senthil-Nathan, S. Toxic effect of essential oil and its compounds isolated from Sphaeranthus amaranthoides Burm. f. against dengue mosquito vector Aedes aegypti Linn. Pest. Biochem. Physiol. 2019, 160, 163–170. [Google Scholar] [CrossRef]
- Chellappandian, M.; Vasantha-Srinivasan, P.; Senthil-Nathan, S.; Karthi, S.; Thanigaivel, A.; Ponsankar, A.; Kalaivani, K.; Hunter, W.B. Botanical essential oils and uses as mosquitocides and repellents against dengue. Environ. Int. 2018, 113, 214–230. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G. Plant-borne compounds and nanoparticles: Challenges for medicine, parasitology and entomology. Environ. Sci. Pollut. Res. 2018, 25, 10149–10150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurunathan, A.; Senguttuvan, J.; Paulsamy, S. Evaluation of mosquito repellent activity of isolated Oleic Acid, Eicosyl Ester from Thalictrum javanicum. Indian J. Pharm Sci. 2016, 78, 103–110. [Google Scholar] [PubMed] [Green Version]
- Song, X.; Chen, Z.; Jia, R.; Cao, M.; Zou, Y.; Li, L.; Liang, X.; Yin, L.; He, C.; Yue, G.; et al. Transcriptomics and proteomic studies reveal acaricidal mechanism of octadecanoic acid-3, 4-tetrahydrofuran diester against Sarcoptes scabiei var. cuniculi. Sci. Rep. 2017, 7, 45479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benelli, G.; Bedini, S.; Cosci, F.; Toniolo, C.; Conti, B.; Nicoletti, M. Larvicidal and ovideterrent properties of neem oil and fractions against the filariasis vector Aedes albopictus (Diptera: Culicidae): A bioactivity survey across production sites. Parasitol. Res. 2015, 114, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Murugan, K.; Jeyabalan, D. Effect of certain plant extracts against the mosquito, Anopheles stephensi Liston. Curr. Sci. 2001, 81, 667–672. [Google Scholar]
- Pushpalatha, E.; Muthukrishnan, J. Larvicidal activity of a few plant extracts against Culex quinquefasciatus and Anopheles stephens. Indian J. Malariol. 1995, 32, 14–23. [Google Scholar]
- Kamaraj, C.; Bagavan, A.; Elango, G.; Zahir, A.; Rajakumar, G.; Manimuthu, S. Larvicidal activity of medicinal plant extracts against Anopheles subpictus and Culex triaeniorhynchus. Indian J. Med. Res. 2011, 134, 101–106. [Google Scholar]
- Rattan, R.S. Mechanism of action of insecticidal secondary metabolites of plant origin. Crop. Prot. 2010, 29, 913–920. [Google Scholar] [CrossRef]
- Board, P.G.; Menon, D. Glutathione transferases, regulators of cellular metabolism and physiology. Biochem. Biophys. Acta 2011, 11, 3267–3288. [Google Scholar] [CrossRef] [PubMed]
- Rose, R.L.; Hodgson, E. A Textbook of Modern Toxicology; John Wiley & Sons: New York, NY, USA, 2014; 672p. [Google Scholar]
- Benelli, G. Plant-borne ovicides in the fight against mosquito vectors of medical and vertinary importance: A systematic review. Parasitol. Res. 2015, 114, 3201–3212. [Google Scholar] [CrossRef] [PubMed]
- Ponsankar, A.; Vasantha-Srinivasan, P.; Senthil-Nathan, S.; Edwin, E.S.; Thanigaivel, A. Target and non-target toxicity of botanical insecticide derived from Couroupita guianensis L. flower against generalist herbivore, Spodoptera litura Fab. and an earthworm, Eisenia foetida Savigny. Ecotoxicol. Environ. Saf. 2016, 133, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Edwin, E.; Vasantha-Srinivasan, P.; Senthil-Nathan, S.; Thanigaivel, A.; Ponsankar, A.; Pradeepa, V.; Selin-Rani, S.; Kalaivani, K.; Hunter, W.B.; Abdel-Megeed, A.; et al. Anti-dengue efficacy of bioactive andrographolide from Andrographis paniculata (Lamiales: Acanthaceae) against the primary dengue vector Aedes aegypti (Diptera: Culicidae). Acta Trop. 2016, 163, 167–178. [Google Scholar] [CrossRef]
- Abtew, A.; Subramanian, S.; Cheseto, X.; Kreiter, S.; Garzia, G.T.; Martin, T. Repellency of plant extracts against the legume flower thrips Megalurothrips sjostedti (Thysanoptera: Thripidae). Insects 2015, 6, 608–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandar, P.; Sudhan, M.; Manikanta Kumar, A.; Manjunath Sheety, K.S.; Hemanth, C. Antibacterial activity of various extracts roots of Decalepis hamiltonii Wight & Arn. Int. J. Pharmacol. Res. 2013, 3, 67–70. [Google Scholar]
- Devi, M.; Latha, P. GC-MS analysis of methanolic root extract of Decalepis hamiltonii (Wight & Arn). World J. Pharm. Pharm. Sci. 2014, 3, 983–989. [Google Scholar]
- Vasantha-Srinivasan, P.; Karthi, S.; Chellappandian, M.; Ponsankar, A.; Thanigaivel, A.; Senthil-Nathan, S.; Chandramohan, D.; Ganesan, R. Aspergillus flavus (Link) toxins reduces the fitness of dengue vector Aedes aegypti (Linn.) and their non-target toxicity against aquatic predator. Microb. Pathog. 2019, 128, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Yogarajalakshmi, P.; Poonguzhali, T.V.; Ganesan, R.; Karthi, S.; Senthil-Nathan, S.; Krutmuang, P.; Radhakrishnan, N.; Mo-hammad, F.; Kim, T.; Vasantha-Srinivsan, P. Toxicological screening of marine red algae Champia parvula (C. Agardh) against the dengue mosquito vector Aedes aegypti (Linn.) and its non-toxicity against three beneficial aquatic predators. Aquat. Toxicol. 2020, 222, 105474. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No | Name of the Compounds | Structure | R. Time | Peak Area (%) | M. Formula | M. Weight |
|---|---|---|---|---|---|---|
| 1 | Benzaldehyde, 2-hydroxy-4-methoxy- | 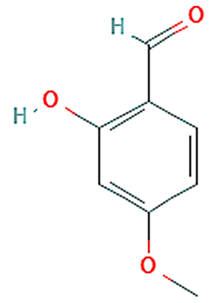 | 11.868 | 10.35 | C8H8O3 | 152.15 |
| 2 | Dodecanoic acid | 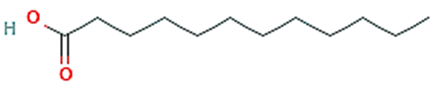 | 15.658 | 11.02 | C12H24O2 | 200.32 |
| 3 | Octadecanoic acid | 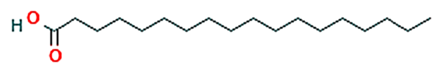 | 18.437 | 22.10 | C18H36O2 | 284.5 |
| 4 | n-Hexadecanoic acid |  | 21.058 | 21.05 | C16H32O2 | 256.42 |
| 5 | Linoleic acid methyl ester |  | 22.506 | 14.20 | C19H34O2 | 294.4721 |
| 6 | Oleic acid |  | 23.240 | 21.04 | C18H34O2 | 282.5 |
| Total Peak Area (%) | 99.76 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prakash, P.; Gayathiri, E.; Manivasagaperumal, R.; Krutmuang, P. Biological Activity of Root Extract Decalepis hamiltonii (Wight & Arn) against Three Mosquito Vectors and Their Non-Toxicity against the Mosquito Predators. Agronomy 2021, 11, 1267. https://doi.org/10.3390/agronomy11071267
Prakash P, Gayathiri E, Manivasagaperumal R, Krutmuang P. Biological Activity of Root Extract Decalepis hamiltonii (Wight & Arn) against Three Mosquito Vectors and Their Non-Toxicity against the Mosquito Predators. Agronomy. 2021; 11(7):1267. https://doi.org/10.3390/agronomy11071267
Chicago/Turabian StylePrakash, Palanisamy, Ekambaram Gayathiri, Rengarajan Manivasagaperumal, and Patcharin Krutmuang. 2021. "Biological Activity of Root Extract Decalepis hamiltonii (Wight & Arn) against Three Mosquito Vectors and Their Non-Toxicity against the Mosquito Predators" Agronomy 11, no. 7: 1267. https://doi.org/10.3390/agronomy11071267
APA StylePrakash, P., Gayathiri, E., Manivasagaperumal, R., & Krutmuang, P. (2021). Biological Activity of Root Extract Decalepis hamiltonii (Wight & Arn) against Three Mosquito Vectors and Their Non-Toxicity against the Mosquito Predators. Agronomy, 11(7), 1267. https://doi.org/10.3390/agronomy11071267