
Effect of Nutrient Solution pH on the Growth, Yield and Quality of Taraxacum officinale and Reichardia picroides in a Floating Hydroponic System


 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Treatments
2.2. Plant Growth Parameters
2.3. Leaves and Roots Macro-and Micro-Elements Concentrations
2.4. Chemical Composition Analyses in Leaves
2.4.1. Total Soluble Solids Content (TSSC) and Titratable Acidity (TA)
2.4.2. Content in Chlorophyll and Carotenoids + Xanthophylls
- By taking SPAD index readings with a SPAD-502 Chlorophyll Meter (Konica-Minolta Co. Ltd., Tokyo, Japan) at two spots on the upper surface, 0.5–1 cm right and left from the central vein of each leaf. Measurements were taken from the two youngest but fully expanded leaves, individually in all 40 plants (ten plants per replication) that were sampled in each harvesting date [42].
- By quantifying chlorophyll a, b and total chlorophyll content of leaves in acetone extracts of homogenized samples, as described in Karapanos et al. [43]. Absorbance of the extracts at 663 and 647 nm was measured in a spectrophotometer (model Lambda 1A, PerkinElmer, Waltham, MA, USA) and the chlorophyll content was calculated according to the equations referred by Lichtenthaler and Buschmann [44].
2.4.3. Total Phenolics Content
2.5. Statistical Analysis
3. Results
3.1. The Effect of Nutrient Solution pH on Taraxacum officinale
3.2. The Effect of Nutrient Solution pH on Reichardia picroides
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elia, A.; Santamaria, P. Biodiversity in vegetable crops: A heritage to save. The case of the Puglia region. Ital. J. Agron. 2013, 8, 21–34. [Google Scholar] [CrossRef]
- Renna, M.; Montesano, F.; Signore, A.; Gonnella, M.; Santamaria, P. BiodiverSO: A Case Study of Integrated Project to Preserve the Biodiversity of Vegetable Crops in Puglia (Southern Italy). Agriculture 2018, 8, 128. [Google Scholar] [CrossRef]
- Pieroni, A. Medicinal plants and food medicines in the folk traditions of the upper Lucca Province, Italy. J. Ethnopharmacol. 2000, 70, 235–273. [Google Scholar] [CrossRef]
- Garcia-Oliveira, P.; Barral, M.; Carpena, M.; Gullón, P.; Fraga-Corral, M.; Otero, P.; Prieto, M.A.; Simal-Gandara, J. Traditional plants from Asteraceae family as potential candidates for functional food industry. Food Funct. 2021, 12, 2850–2873. [Google Scholar] [CrossRef]
- Vanzani, P.; Rossetto, M.; De Marco, V.; Sacchetti, L.E.; Paoletti, M.G.; Rigo, A. Wild Mediterranean plants as traditional food: A valuable source of antioxidants. J. Food Sci. 2011, 76, 46–51. [Google Scholar] [CrossRef]
- Recio, M.C.; Giner, R.M.; Hermenegildo, M.; Peris, J.B.; Iez, I.S.M.A.I.; Rios, J. Phenolics of Reichardia and their taxonomic implications. Biochem. Syst. Ecol. 1992, 20, 449–452. [Google Scholar] [CrossRef]
- Gatto, M.A.; Ippolito, A.; Linsalata, V.; Cascarano, N.A.; Nigro, F.; Vanadia, S.; Di Venere, D. Activity of extracts from wild edible herbs against postharvest fungal diseases of fruit and vegetables. Postharvest Biol. Technol. 2011, 61, 72–82. [Google Scholar] [CrossRef]
- Jeon, H.J.; Kang, H.J.; Jung, H.J.; Kang, Y.S.; Lim, C.J.; Kim, Y.M.; Park, E.H. Anti-inflammatory activity of Taraxacum officinale. J. Ethnopharmacol. 2008, 115, 82–88. [Google Scholar] [CrossRef]
- Petropoulos, S.; Karkanis, A.; Martins, N.; Ferreira, I.C.F.R. Phytochemical composition and bioactive compounds of common purslane (Portulaca oleracea L.) as affected by crop management practices. Trends Food Sci. Technol. 2016, 55, 1–10. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Fernandes, Â.; Tzortzakis, N.; Sokovic, M.; Ciric, A.; Barros, L.; Ferreira, I.C.F.R. Bioactive compounds content and antimicrobial activities of wild edible Asteraceae species of the Mediterranean flora under commercial cultivation conditions. Food Res. Int. 2019, 119, 859–868. [Google Scholar] [CrossRef]
- Petropoulos, S.; Ntatsi, G.; Levizou, E.; Barros, L.; Ferreira, I. Nutritional profile and chemical composition of Cichorium spinosum ecotypes. LWT-Food Sci. Technol. 2016, 73, 95–101. [Google Scholar] [CrossRef]
- Chatzopoulou, E.; Carocho, M.; Di Gioia, F.; Petropoulos, S.A. The beneficial health effects of vegetables and wild edible greens: The case of the Mediterranean diet and its sustainability. Appl. Sci. 2020, 10, 9144. [Google Scholar] [CrossRef]
- Corrêa, R.C.G.; Gioia, F.; Ferreira, I.C.F.R.; Petropoulos, S.A. Wild greens used in the Mediterranean diet. In The Mediterranean Diet: An Evidence-Based Approach; Preedy, V., Watson, R., Eds.; Academic Press: London, UK, 2020; pp. 209–228. ISBN 9788578110796. [Google Scholar]
- Sánchez-Mata, M.C.; Loera, R.D.C.; Morales, P.; Fernández-Ruiz, V.; Cámara, M.; Marqués, C.D.; Pardo-de-Santayana, M.; Tardío, J.; Cabrera Loera, R.D.; Morales, P.; et al. Wild vegetables of the Mediterranean area as valuable sources of bioactive compounds. Genet. Resour. Crop Evol. 2012, 59, 431–443. [Google Scholar] [CrossRef]
- Ceccanti, C.; Landi, M.; Benvenuti, S.; Pardossi, A.; Guidi, L. Mediterranean wild edible plants: Weeds or “new functional crops”? Molecules 2018, 23, 2299. [Google Scholar] [CrossRef]
- Correa, R.C.G.; Di Gioia, F.; Ferreira, I.; Petropoulos, S.A. Halophytes for future horticulture: The case of small-scale farming in the Mediterranean basin. In Halophytes for Future Horticulture: From Molecules to Ecosystems towards Biosaline Agriculture; Grigore, M.-N., Ed.; Springer Nature Switzerland AG: Cham, Switzerland, 2020; pp. 1–28. ISBN 9783030178543. [Google Scholar]
- Guiomar, N.; Godinho, S.; Pinto-Correia, T.; Almeida, M.; Bartolini, F.; Bezák, P.; Biró, M.; Bjørkhaug, H.; Bojnec, Š.; Brunori, G.; et al. Typology and distribution of small farms in Europe: Towards a better picture. Land Use Policy 2018, 75, 784–798. [Google Scholar] [CrossRef]
- Singh, R.; Singh, G.S. Traditional agriculture: A climate-smart approach for sustainable food production. Energy Ecol. Environ. 2017, 2, 296–316. [Google Scholar] [CrossRef]
- Frison, E.A.; Cherfas, J.; Hodgkin, T. Agricultural biodiversity is essential for a sustainable improvement in food and nutrition security. Sustainability 2011, 3, 238–253. [Google Scholar] [CrossRef]
- Sardare, M.D.; Admane, S.A. Review on plant without soil-hydroponics. Int. J. Res. Eng. Technol. 2013, 02, 299–304. [Google Scholar]
- Giménez, A.; Fernández, J.A.; Pascual, J.A.; Ros, M.; López-Serrano, M.; Egea-Gilabert, C. An agroindustrial compost as alternative to peat for production of baby leaf red lettuce in a floating system. Sci. Hortic. 2019, 246, 907–915. [Google Scholar] [CrossRef]
- Maggini, R.; Kiferle, C.; Pardossi, A. Hydroponic production of medicinal plants. In Medicinal Plants: Antioxidant Properties, Traditional Uses and Conservation Strategies; Pereira, D., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2014; pp. 91–116. [Google Scholar]
- Sgherri, C.; Cecconami, S.; Pinzino, C.; Navari-Izzo, F.; Izzo, R. Levels of antioxidants and nutraceuticals in basil grown in hydroponics and soil. Food Chem. 2010, 123, 416–422. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234. [Google Scholar] [CrossRef]
- Rouphael, Y.; Petropoulos, S.A.; Cardarelli, M.; Colla, G. Salinity as eustressor for enhancing quality of vegetables. Sci. Hortic. 2018, 234, 361–369. [Google Scholar] [CrossRef]
- Putra, P.A.; Yuliando, H. Soilless culture system to support water use efficiency and product quality: A review. Agric. Agric. Sci. Procedia 2015, 3, 283–288. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2011. [Google Scholar]
- Singh, H.; Dunn, B.; Payton, M.; Brandenberger, L. Fertilizer and cultivar selection of lettuce, Basil, and Swiss chard for hydroponic production. HortTechnology 2019, 29, 50–56. [Google Scholar] [CrossRef]
- Gillespie, D.P.; Kubota, C.; Miller, S.A. Effects of Low pH of hydroponic nutrient solution on plant growth, nutrient uptake, and root rot disease incidence of Basil (Ocimum basilicum L.). HortScience 2020, 55, 1251–1258. [Google Scholar] [CrossRef]
- Alam, S.; Naqvi, S.; Ansari, R. Impact of soil pH on nutrient uptake by crop plants. In Handbook of Plant and Crop Stress; Pessarakli, M., Ed.; Marcel Dekker: New York, NY, USA, 1999; pp. 51–60. [Google Scholar]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett. 2006, 4, 147–157. [Google Scholar] [CrossRef]
- Chalker-Scott, L. Environmental significance of anthocyanins in plant stress responses. Photochem. Photobiol. 1999, 70, 1–9. [Google Scholar] [CrossRef]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef] [PubMed]
- Michalak, A. Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress. Polish J. Environ. Stud. 2006, 15, 523–530. [Google Scholar]
- Young, A.J. The photoprotective role of carotenoids in higher plants. Physiol. Plant. 1991, 83, 702–708. [Google Scholar] [CrossRef]
- Petropoulos, S.; Levizou, E.; Ntatsi, G.; Fernandes, Â.; Petrotos, K.; Akoumianakis, K.; Barros, L.; Ferreira, I. Salinity effect on nutritional value, chemical composition and bioactive compounds content of Cichorium spinosum L. Food Chem. 2017, 214, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Boltz, D.F.; Lueck, C.H. Colorimetric Determination of Non-Metals; Boltz, D.F., Ed.; Interscience Publishers: New York, NY, USA, 1958; pp. 29–46. [Google Scholar]
- Kalra, Y. Handbook of Reference Methods for Plant Analysis; CRC Press: New York, NY, USA, 1998. [Google Scholar]
- Allen, S. Chemical Analysis of Ecological Materials, 2nd ed.; Blackwell Scientific Publications: Oxford/London, UK, 1989. [Google Scholar]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Olympios, C.; Ropokis, A.; Vlachou, G.; Ntatsi, G. Fruit volatiles, quality, and yield of watermelon as affected by grafting. J. Agric. Sci. Technol. 2014, 16, 873–885. [Google Scholar]
- Karkanis, A.C.; Petropoulos, S.A. Physiological and growth responses of several genotypes of common purslane (Portulaca oleracea L.) under Mediterranean semi-arid conditions. Not. Bot. Horti Agrobot. Cluj-Napoca 2017, 45, 569–575. [Google Scholar] [CrossRef]
- Karapanos, I.; Papandreou, A.; Skouloudi, M.; Makrogianni, D.; Fernández, J.A.; Rosa, E.; Ntatsi, G.; Bebeli, P.J.; Savvas, D. Cowpea fresh pods—A new legume for the market: Assessment of their quality and dietary characteristics of 37 cowpea accessions grown in southern Europe. J. Sci. Food Agric. 2017, 97, 4343–4352. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Singleton, V.; Rossi, J.J. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Anugoolprasert, O.; Kinoshita, S.; Naito, H.; Shimizu, M.; Ehara, H. Effect of low pH on the growth, physiological characteristics and nutrient absorption of sago palm in a hydroponic system. Plant Prod. Sci. 2012, 15, 125–131. [Google Scholar] [CrossRef]
- Zhao, D.; Hao, Z.; Wang, J.; Tao, J. Effects of pH in irrigation water on plant growth and flower quality in herbaceous peony (Paeonia lactiflora Pall.). Sci. Hortic. 2013, 154, 45–53. [Google Scholar] [CrossRef]
- Roosta, H.R.; Rezaei, I. Effect of nutrient solution pH on the vegetative and reproductive growth and physiological characteristics of rose cv. ‘Grand gala’ in hydroponic system. J. Plant Nutr. 2014, 37, 2179–2194. [Google Scholar] [CrossRef]
- Sonneveld, C. Composition of nutrient solutions. In Hydroponic Production of Vegetables and Ornamentals; Savvas, D., Passam, H., Eds.; Embryo Publications: Athens, Greece, 2002; pp. 179–210. [Google Scholar]
- Maggini, R.; Benvenuti, S.; Leoni, F.; Pardossi, A. Terracrepolo (Reichardia picroides (L.) Roth.): Wild food or new horticultural crop? Sci. Hortic. 2018, 240, 224–231. [Google Scholar] [CrossRef]
- Hajnos, M. Buffer Capacity of soils. In Encyclopedia of Agrophysics; Gliński, J., Horabik, J., Lipiec, J., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 94–95. ISBN 978-90-481-3585-1. [Google Scholar]
- Acevedo-Whitehouse, K.; Duffus, A.L.J. Effects of environmental change on wildlife health. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 3429–3438. [Google Scholar] [CrossRef] [PubMed]
- Rosas, A.; Rengel, Z.; Mora, M.D.L.L. Manganese supply and pH influence growth, carboxylate exudation and peroxidase activity of ryegrass and white clover. J. Plant Nutr. 2007, 30, 253–270. [Google Scholar] [CrossRef]
- Schubert, S.; Schubert, E.; Mengel, K. Effect of low pH of the root medium on proton release, growth, and nutrient uptake of field beans (Vicia faba). Plant Soil 1990, 124, 239–244. [Google Scholar] [CrossRef]
- Yan, F.; Schubert, S.; Mengel, K. Effect of low root medium pH on net proton release, root respiration, and root growth of corn (Zea mays L.) and broad bean (Vicia faba L.). Plant Physiol. 1992, 99, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.I.; Park, J.M.; Kim, S.H.; Kang, N.J.; Park, K.S.; Lee, S.Y.; Jeong, B.R. Effects of root zone pH and nutrient concentration on the growth and nutrient uptake of tomato seedlings. J. Plant Nutr. 2011, 34, 640–652. [Google Scholar] [CrossRef]
- Findenegg, G.R. A comparative study of ammonium toxicity at different constant pH of the nutrient solution. Plant Soil 1987, 103, 239–243. [Google Scholar] [CrossRef]
- Assimakopoulou, A. Effect of iron supply and nitrogen form on growth, nutritional status and ferric reducing activity of spinach in nutrient solution culture. Sci. Hortic. 2006, 110, 21–29. [Google Scholar] [CrossRef]
- Dyśko, J.; Kaniszewski, S.; Kowalczyk, W. The effect of nutrient solution pH on phosphorus availability in soilless culture of tomato. J. Elem. 2008, 13, 189–198. [Google Scholar]
- Savvas, D.; Karagianni, V.; Kotsiras, A.; Demopoulos, V.; Karkamisi, I.; Pakou, P. Interactions between ammonium and pH of the nutrient solution supplied to gerbera (Gerbera jamesonii) grown in pumice. Plant Soil 2003, 254, 393–402. [Google Scholar] [CrossRef]
- Lippmann, R.; Babben, S.; Menger, A.; Delker, C.; Quint, M. Development of wild and cultivated plants under global warming conditions. Curr. Biol. 2019, 29, R1326–R1338. [Google Scholar] [CrossRef]
- Rietra, R.P.J.J.; Heinen, M.; Dimkpa, C.O.; Bindraban, P.S. Effects of nutrient antagonism and synergism on yield and fertilizer use efficiency. Commun. Soil Sci. Plant Anal. 2017, 48, 1895–1920. [Google Scholar] [CrossRef]
- Assimakopoulou, A.; Holevas, C.D.; Fasseas, K. Relative susceptibility of some Prunus rootstocks in hydroponics to iron deficiency. J. Plant Nutr. 2011, 34, 1014–1033. [Google Scholar] [CrossRef]
- Clark, M.B.; Mills, H.A.; Robacker, C.D.; Latimer, J.G. Influence of nitrate: Ammonium ratios on growth and elemental concentration in two Azalea cultivars. J. Plant Nutr. 2003, 26, 2503–2520. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 3rd ed.; Sinauer Associates Publishers: Sunderland, MA, USA, 2002. [Google Scholar]
- Arnon, D.I.; Johnson, C.M. Influence of hydrogen ion concentration on the growth of higher plants under controlled conditions. Plant Physiol. 1941, 525–539. [Google Scholar] [CrossRef]
- Colonna, E.; Rouphael, Y.; Barbieri, G.; De Pascale, S. Nutritional quality of ten leafy vegetables harvested at two light intensities. Food Chem. 2016, 199, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, P. Nitrate in vegetables: Toxicity, content, intake and EC regulation. J. Sci. Food Agric. 2006, 86, 10–17. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Fernandes, Â.; Dias, M.I.; Pereira, C.; Calhelha, R.C.; Chrysargyris, A.; Tzortzakis, N.; Ivanov, M.; Sokovic, M.D.; Barros, L.; et al. Chemical composition and plant growth of Centaurea raphanina subsp. mixta plants cultivated under saline conditions. Molecules 2020, 25, 2204. [Google Scholar] [CrossRef]
- Petropoulos, S.; Fernandes, A.; Barros, L.; Ferreira, I. A comparison of the phenolic profile and antioxidant activity of different Cichorium spinosum L. ecotypes. J. Sci. Food Agric. 2017, 98, 183–189. [Google Scholar] [CrossRef]
- Petropoulos, S.; Fernandes, Â.; Karkanis, A.; Ntatsi, G.; Barros, L.; Ferreira, I. Successive harvesting affects yield, chemical composition and antioxidant activity of Cichorium spinosum L. Food Chem. 2017, 237, 83–90. [Google Scholar] [CrossRef]



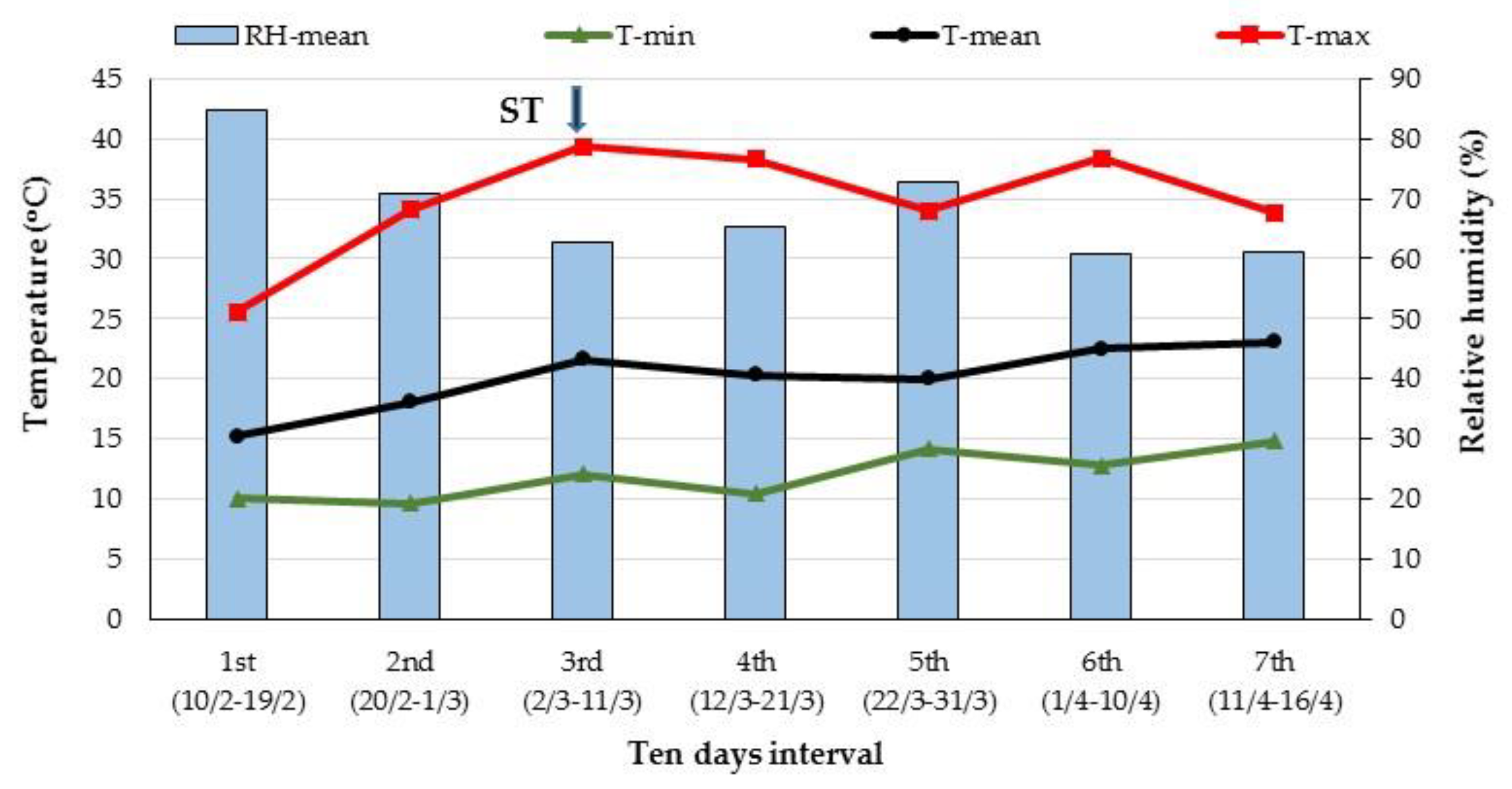{kind=link}
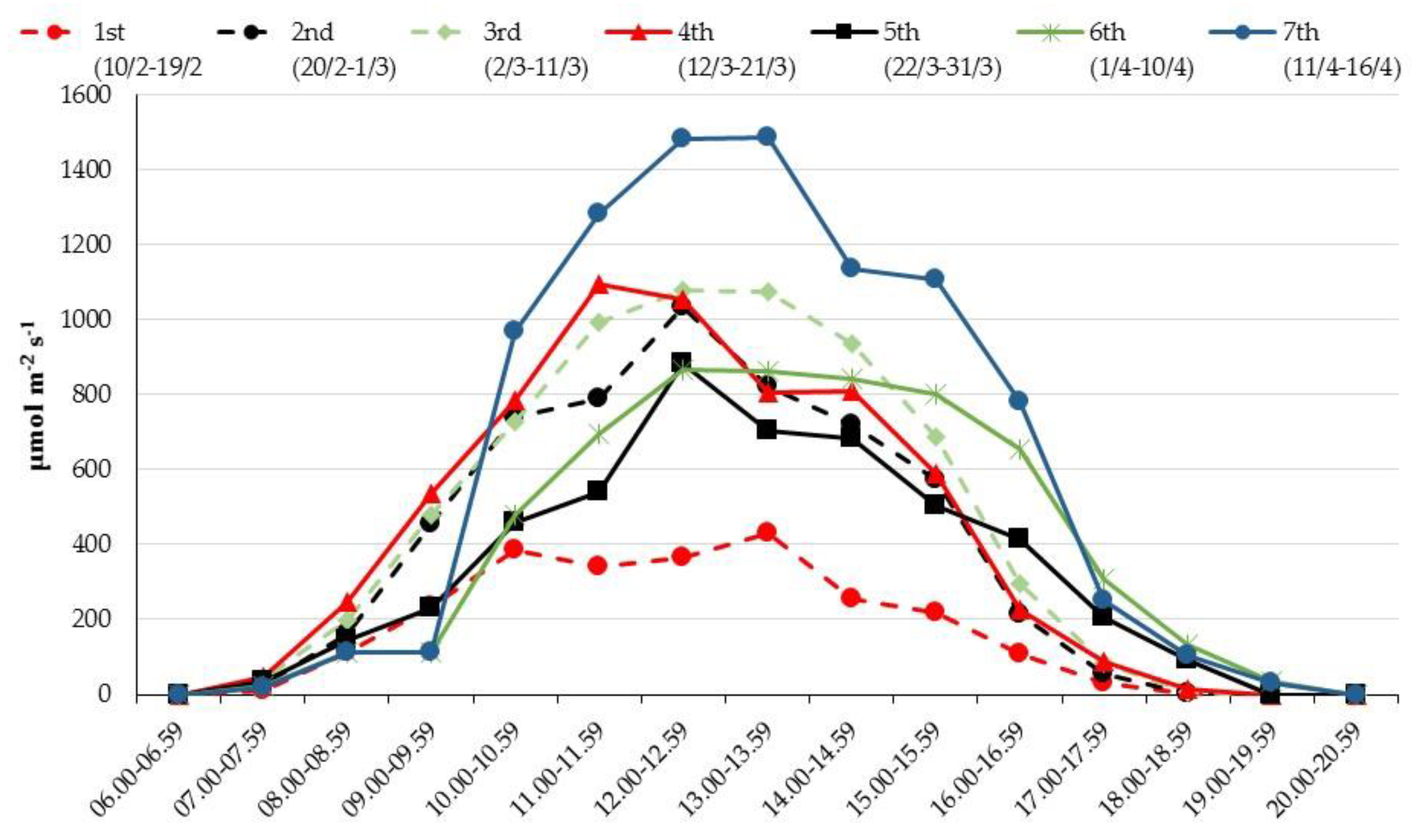{kind=link}
| pH | H1 | H2 | H1 | H2 | H1 | H2 | H1 | H2 |
|---|---|---|---|---|---|---|---|---|
| Leaf Number Plant−1 | Rosette Diameter (cm) | Non-Marketable Leaves (%) Plant−1 | SPAD Index | |||||
| 4.0 | 18.9 a(b) | 27.9 a(a) | 24.2 b(b) | 36.7 b(a) | 12.9 a(a) | 12.0 a(a) | 33.1 a(b) | 43.2 a(a) |
| 5.5 | 18.6 a(b) | 29.9 a(a) | 27.4 a(b) | 45.8 a(a) | 12.5 a(a) | 14.7 a(a) | 35.5 a(b) | 42.1 a(a) |
| 7.0 | 24.4 a(b) | 31.1 a(a) | 30.1 a(b) | 47.5 a(a) | 9.7 a(a) | 11.8 a(a) | 35.0 a(b) | 40.7 a(a) |
| pH | Total plant FW (g) | Root FW (g) plant−1 | Marketable leaves FW (g) plant−1 | Leaf DMC (%) | ||||
| 4.0 | 8.8 b(b) | 36.3 b(a) | 1.3 a (b) | 5.8 b(a) | 7.4 b(b) | 28.4 a(a) | 9.9 a(b) | 12.7 a(a) |
| 5.5 | 11.3 ab(b) | 47.3 a(a) | 1.4 a (b) | 7.9 a(a) | 9.9 a(b) | 32.9 a(a) | 11.3 a(b) | 13.3 a(a) |
| 7.0 | 12.5 a(b) | 41.1 ab(a) | 1.5 a (b) | 7.3 ab(a) | 10.9 a(b) | 31.7 a(a) | 11.0 a(b) | 13.0 a(a) |
| pH | H1 | H2 | H1 | H2 | H1 | H2 | H1 | H2 | H1 | H2 |
|---|---|---|---|---|---|---|---|---|---|---|
| N (g kg−1 DW) | P (g kg−1 DW) | K (g kg−1 DW) | Ca (g kg−1 DW) | Mg (g kg−1 DW) | ||||||
| 4.0 | 50.9 a(a) | 44.2 a(b) | 11.5 a(a) | 10.8 a(a) | 62.3 b(a) | 56.2 b(b) | 9.0 a(a) | 8.5 a(a) | 3.3 a(a) | 3.1 a(a) |
| 5.5 | 51.7 a(a) | 46.2 a(b) | 11.7 a(a) | 10.3 a(a) | 62.3 b(b) | 67.7 a(a) | 8.5 a(a) | 9.3 a(a) | 3.4 a(a) | 3.5 a(a) |
| 7.0 | 53.8 a(a) | 44.9 a(b) | 10.5 a(a) | 8.9 b(b) | 67.3 a(a) | 66.1 a(a) | 8.2 a(a) | 7.4 a(a) | 3.7 a(a) | 3.4 a(a) |
| pH | Fe (mg kg−1 DW) | Mn (mg kg−1 DW) | Zn (mg kg−1 DW) | Cu (mg kg−1 DW) | B (mg kg−1 DW) | |||||
| 4.0 | 101.3 a(a) | 92.0 a(a) | 25.8 a(a) | 25.7 a(a) | 28.5 a(b) | 37.8 a(a) | 5.0 c(b) | 8.7 a(a) | 52.2 a(a) | 60.1 a(a) |
| 5.5 | 98.2 a(a) | 88.4 a(a) | 23.1 a(a) | 21.2 b(a) | 22.4 b(b) | 40.3 a(a) | 6.5 b(b) | 8.1 a(a) | 47.7 a(a) | 49.5 a(a) |
| 7.0 | 98.7 a(a) | 97.2 a(a) | 24.7 a(a) | 13.9 c(b) | 25.0 b(a) | 16.8 b(b) | 8.0 a(a) | 8.9 a(a) | 40.4 a(a) | 46.2 a(a) |
| pH | H1 | H2 | H1 | H2 | H1 | H2 | H1 | H2 | H1 | H2 |
|---|---|---|---|---|---|---|---|---|---|---|
| N (g kg−1 DW) | P (g kg−1 DW) | K (g kg−1 DW) | Ca (g kg−1 DW) | Mg (g kg−1 DW) | ||||||
| 4.0 | 43.2 a(a) | 45.6 a(a) | 10.4 a(a) | 11.1 a(a) | 38.5 c(a) | 33.5 b(a) | 3.4 a(a) | 2.9 a(a) | 1.6 a(a) | 1.5 a(a) |
| 5.5 | 45.4 a(a) | 42.5 a(a) | 11.4 a(a) | 12.2 a(a) | 46.2 b(a) | 43.9 a(a) | 3.6 a(a) | 3.8 a(a) | 1.7 a(a) | 1.7 a(a) |
| 7.0 | 46.9 a(a) | 44.5 a(a) | 12.3 a(a) | 9.2 b(b) | 56.6 a(a) | 44.3 a(a) | 3.2 a(a) | 3.5 a(a) | 1.6 a(a) | 2.0 a(a) |
| pH | Fe (mg kg−1 DW) | Mn (mg kg−1 DW) | Zn (mg kg−1 DW) | Cu (mg kg−1 DW) | B (mg kg−1 DW) | |||||
| 4.0 | 146.6 a(a) | 140.0 a(a) | 11.2 b(a) | 11.0 b(a) | 29.5 a(a) | 32.6 b(a) | 9.9 b(b) | 16.7 a(a) | 28.8 a(a) | 28.7 a(a) |
| 5.5 | 129.0 b(a) | 121.7 ab(a) | 11.3 b(b) | 13.3 b(a) | 29.4 a(b) | 37.9 a(a) | 12.1 b(b) | 14.1 a(a) | 29.6 a(a) | 35.2 a(a) |
| 7.0 | 120.3 b(a) | 108.7 b(b) | 23.9 a(b) | 29.4 a(a) | 30.6 a(a) | 20.3 c(b) | 16.7 a(a) | 16.1 a(a) | 31.5 a(a) | 28.8 a(a) |
| pH | H1 | H2 | H1 | H2 | H1 | H2 | H1 | H2 |
|---|---|---|---|---|---|---|---|---|
| Chlorophyll a (mg kg−1 FW) | Chlorophyll b (mg kg−1 FW) | Total Chlorophyll (mg kg−1 FW) | Nitrates (mg kg−1 FW) | |||||
| 4.0 | 866.9 a(a) | 815.6 a(a) | 412.5 a(a) | 400.1 a(a) | 1279.3 a(a) | 1215.7 a(a) | 2621.5 b(a) | 2694.9 b(a) |
| 5.5 | 819.4 ab(a) | 727.1 ab(b) | 396.1 ab(a) | 361.8 a(b) | 1215.6 ab(a) | 1089.0 ab(b) | 3396.8 ab(b) | 4405.0 a(a) |
| 7.0 | 750.3 b(a) | 700.4 b(a) | 359.8 b(a) | 368.9 a(a) | 1110.1 b(a) | 1069.3 b(a) | 4170.1 a(a) | 4032.6 a(a) |
| pH | TSSC (oBrix) | TA (g malic acid kg−1 FW) | Carotenoids + xanthophylls (mg kg−1 FW) | Total phenolics (mg GAE kg−1 FW) | ||||
| 4.0 | 5.58 a(a) | 4.58 a(b) | 2.81 a(a) | 1.89 a(b) | 117.3 a(a) | 105.9 a(a) | 735.1 a(a) | 758.4 a(a) |
| 5.5 | 4.23 b(a) | 4.13 a(a) | 2.19 b(a) | 1.75 a(b) | 115.2 a(a) | 97.3 a(b) | 593.5 b(a) | 636.1 b(a) |
| 7.0 | 4.30 b(a) | 4.28 a(a) | 1.99 b(a) | 1.51 b(b) | 105.1 a(a) | 97.0 a(a) | 537.6 b(b) | 702.2 ab(a) |
| pH | H1 | H2 | H1 | H2 | H1 | H2 | H1 | H2 |
|---|---|---|---|---|---|---|---|---|
| Leaf Number Plant−1 | Rosette Diameter (cm) | Non-Marketable Leaves (%) Plant−1 | SPAD Index | |||||
| 4.0 | 21.6 b(b) | 33.8 c(a) | 22.7 b(b) | 29.1 b(a) | 13.3 a(a) | 8.6 a(a) | 47.5 a(a) | 50.7 a(a) |
| 5.5 | 27.1 a(b) | 68.9 a(a) | 32.0 a(b) | 44.0 a(a) | 12.4 a(a) | 9.8 a(a) | 50.0 a(a) | 53.7 a(a) |
| 7.0 | 29.7 a(b) | 56.9 b(a) | 30.6 a(b) | 42.2 a(a) | 12.9 a(a) | 13.1 a(a) | 49.6 a(a) | 50.2 a(a) |
| pH | Total plant FW (g) | Root FW (g) plant−1 | Marketable leaves FW (g) plant−1 | Leaf DMC (%) | ||||
| 4.0 | 11.6 b(b) | 30.5 c(a) | 1.4 b(b) | 5.6 b(a) | 8.3 b(b) | 21.1 c(a) | 7.6 a(b) | 9.7 a(a) |
| 5.5 | 22.0 a(b) | 72.1 a(a) | 3.4 a(b) | 8.6 a(a) | 16.8 a(b) | 54.8 a(a) | 7.9 a(b) | 10.1 a(a) |
| 7.0 | 25.4 a(b) | 55.3 b(a) | 3.2 a(b) | 7.9 ab(a) | 20.5 a(b) | 42.6 b(a) | 7.9 a(b) | 10.3 a(a) |
| pH | H1 | H2 | H1 | H2 | H1 | H2 | H1 | H2 | H1 | H2 |
|---|---|---|---|---|---|---|---|---|---|---|
| N (g kg−1 DW) | P (g kg−1 DW) | K (g kg−1 DW) | Ca (g kg−1 DW) | Mg (g kg−1 DW) | ||||||
| 4.0 | 38.1 a(a) | 35.8 a(a) | 6.4 b(a) | 7.8 a(a) | 51.9 b(a) | 51.5 a(a) | 7.0 b(a) | 7.8 a(a) | 1.8 a(a) | 1.8 a(a) |
| 5.5 | 43.2 a(a) | 29.1 a(b) | 9.8 a(a) | 6.0 ab(b) | 66.2 a(a) | 56.2 a(a) | 9.6 a(a) | 8.7 a(a) | 2.4 a(a) | 2.0 a(a) |
| 7.0 | 45.6 a(a) | 31.6 a(b) | 6.9 b(a) | 4.1 b(b) | 69.6 a(a) | 63.5 a(a) | 8.4 ab(a) | 6.0 b(b) | 2.4 a(a) | 1.7 a(b) |
| pH | Fe (mg kg−1 DW) | Mn (mg kg−1 DW) | Zn (mg kg−1 DW) | Cu (mg kg−1 DW) | B (mg kg−1 DW) | |||||
| 4.0 | 156.1 a(a) | 112.0 a(b) | 53.5 a(a) | 48.3 a(a) | 40.3 a(b) | 52.8 a(a) | 7.4 a(b) | 10.7 a(a) | 29.6 a(b) | 64.9 a(a) |
| 5.5 | 108.1 b(a) | 82.0 b(a) | 52.7 a(a) | 31.9 b(b) | 29.0 b(a) | 34.6 b(a) | 8.6 a(a) | 3.3 c(b) | 45.2 a(a) | 64.3 a(a) |
| 7.0 | 124.4 ab(a) | 89.3 b(b) | 38.5 b(a) | 14.5 c(b) | 20.4 c(a) | 15.3 c(b) | 8.2 a(a) | 8.8 b(a) | 35.9 a(b) | 60.0 a(a) |
| pH | H1 | H2 | H1 | H2 | H1 | H2 | H1 | H2 | H1 | H2 |
|---|---|---|---|---|---|---|---|---|---|---|
| N (g kg−1 DW) | P (g kg−1 DW) | K (g kg−1 DW) | Ca (g kg−1 DW) | Mg (g kg−1 DW) | ||||||
| 4.0 | 46.3 a(a) | 44.4 a(a) | 13.7 b(a) | 13.1 a(a) | 46.2 c(a) | 43.5 a(a) | 3.7 a(a) | 4.0 a(a) | 1.7 a(a) | 1.8 a(a) |
| 5.5 | 53.0 a(a) | 44.4 a(b) | 18.1 a(a) | 10.5 b(b) | 67.3 a(a) | 47.7 a(b) | 4.1 a(a) | 4.5 a(a) | 1.7 a(a) | 1.9 a(a) |
| 7.0 | 47.9 a(a) | 45.6 a(a) | 14.5 b(a) | 6.3 c(b) | 58.1 b(a) | 50.0 a(b) | 4.3 a(a) | 4.6 a(a) | 1.7 a(b) | 2.0 a(a) |
| Fe (mg kg−1 DW) | Mn (mg kg−1 DW) | Zn (mg kg−1 DW) | Cu (mg kg−1 DW) | B (mg kg−1 DW) | ||||||
| 4.0 | 599.9 a(a) | 517.1 a(a) | 29.9 a(a) | 19.1 b(b) | 119.1 a(a) | 74.9 a(b) | 19.9 a(a) | 20.2 a(a) | 32.4 a(b) | 54.7 a(a) |
| 5.5 | 221.7 b(a) | 219.0 b(a) | 23.6 a(b) | 39.0 a(a) | 46.9 b(b) | 59.7 b(a) | 18.6 ab(a) | 13.0 b(b) | 49.1 a(a) | 54.2 a(a) |
| 7.0 | 184.7 b(a) | 175.4 b(a) | 22.6 a(a) | 20.0 b(a) | 33.0 b(a) | 25.4 c(b) | 15.9 b(a) | 16.3 ab(a) | 36.5 a(a) | 55.5 a(a) |
| pH | H1 | H2 | H1 | H2 | H1 | H2 | H1 | H2 |
|---|---|---|---|---|---|---|---|---|
| Chlorophyll a (mg kg−1 FW) | Chlorophyll b (mg kg−1 FW) | Total Chlorophyll (mg kg−1 FW) | Nitrates (mg kg−1 FW) | |||||
| 4.0 | 520.1 a(a) | 456.8 a(b) | 258.3 a(a) | 207.9 a(b) | 778.5 a(a) | 664.8 a(b) | 2142.4 b(b) | 2893.5 a(a) |
| 5.5 | 413.5 b(a) | 458.3 a(a) | 208.7 b(a) | 203.7 a (a) | 622.2 b(a) | 662.1 a(a) | 3487.8 a(a) | 3496.6 a(a) |
| 7.0 | 405.1 b(a) | 453.3 a(a) | 212.4 b(a) | 210.4 a (a) | 617.4 b(a) | 663.7 a(a) | 3389.1 a(a) | 3289.8 a(a) |
| pH | TSSC (oBrix) | TA (g malic acid kg−1 FW) | Carotenoids + xanthophylls (mg kg−1 FW) | Total phenolics (mg GAE kg−1 FW) | ||||
| 4.0 | 4.35 a(a) | 5.23 a(a) | 1.37 a(a) | 1.49 a(a) | 79.7 a(a) | 74.9 a(a) | 1043.6 a(a) | 1214.8 a(a) |
| 5.5 | 3.87 a(a) | 3.30 b(a) | 1.36 a(a) | 1.44 a(a) | 65.3 b(a) | 70.2 a(a) | 775.2 b(a) | 755.3 b(a) |
| 7.0 | 3.70 a(a) | 3.43 b(a) | 1.13 a(a) | 1.13 b(a) | 63.6 b(a) | 72.6 a(a) | 800.0 b(a) | 672.6 b(a) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alexopoulos, A.A.; Marandos, E.; Assimakopoulou, A.; Vidalis, N.; Petropoulos, S.A.; Karapanos, I.C. Effect of Nutrient Solution pH on the Growth, Yield and Quality of Taraxacum officinale and Reichardia picroides in a Floating Hydroponic System. Agronomy 2021, 11, 1118. https://doi.org/10.3390/agronomy11061118
Alexopoulos AA, Marandos E, Assimakopoulou A, Vidalis N, Petropoulos SA, Karapanos IC. Effect of Nutrient Solution pH on the Growth, Yield and Quality of Taraxacum officinale and Reichardia picroides in a Floating Hydroponic System. Agronomy. 2021; 11(6):1118. https://doi.org/10.3390/agronomy11061118
Chicago/Turabian StyleAlexopoulos, Alexios A., Efstathios Marandos, Anna Assimakopoulou, Nikolina Vidalis, Spyridon A. Petropoulos, and Ioannis C. Karapanos. 2021. "Effect of Nutrient Solution pH on the Growth, Yield and Quality of Taraxacum officinale and Reichardia picroides in a Floating Hydroponic System" Agronomy 11, no. 6: 1118. https://doi.org/10.3390/agronomy11061118
APA StyleAlexopoulos, A. A., Marandos, E., Assimakopoulou, A., Vidalis, N., Petropoulos, S. A., & Karapanos, I. C. (2021). Effect of Nutrient Solution pH on the Growth, Yield and Quality of Taraxacum officinale and Reichardia picroides in a Floating Hydroponic System. Agronomy, 11(6), 1118. https://doi.org/10.3390/agronomy11061118