
The Challenge of Combining High Yields with Environmentally Friendly Bioproducts: A Review on the Compatibility of Pesticides with Microbial Inoculants


 , and
, and 
Abstract
1. Introduction
2. History and Use of Pesticides
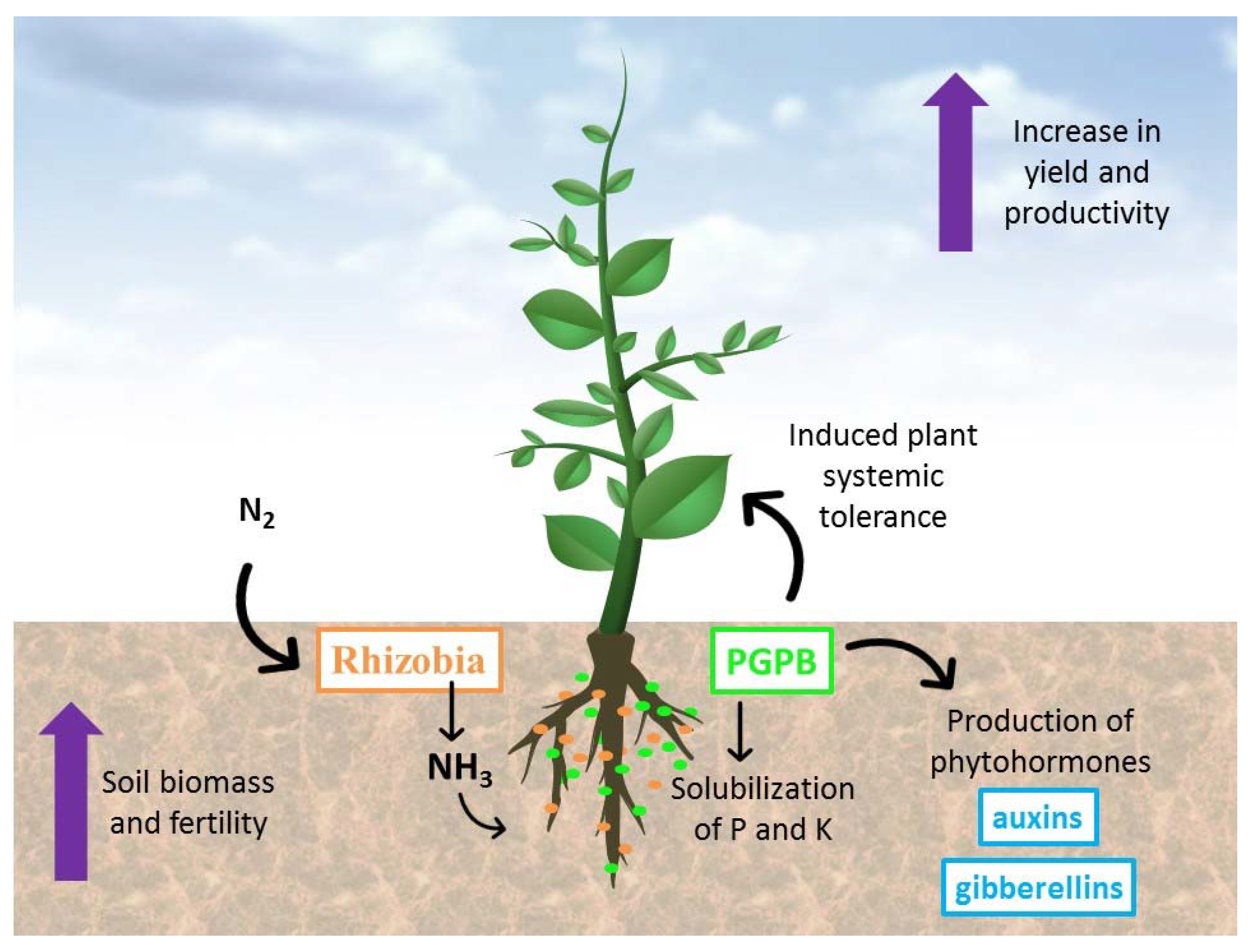
3. Inoculants or Biofertilizers
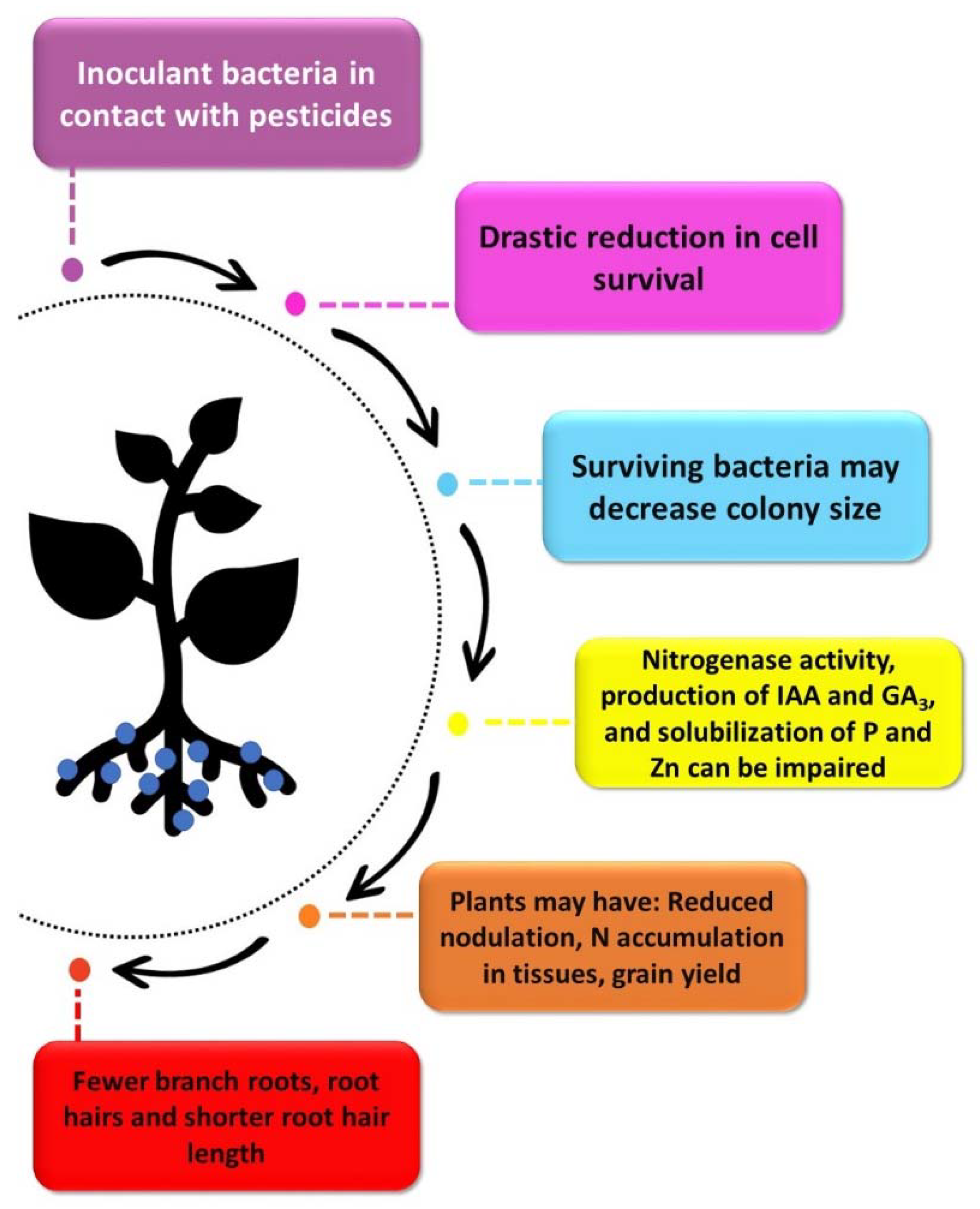
4. Are Pesticides and Microbial Inoculants Compatible?
4.1. Compatibility with Fungicides
4.2. Compatibility with Insecticides
4.3. Compatibility with Herbicides
4.4. Compatibility with Mixtures (Fungicides, Insecticides, and Herbicides)
5. Are There Alternatives to the Challenges of Using Pesticides and Microbial Inoculants?
6. Final Remarks
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations, Department of Economic and Social Affairs, Population Division. World Population Prospects 2019 Highlights. Available online: https://population.un.org/wpp/Publications/Files/WPP2019_Highlights.pdf (accessed on 4 January 2021).
- Ameen, A.; Raza, S. Green Revolution: A Review. Int. J. Adv. Sci. Res. 2018, 3, 129–137. [Google Scholar] [CrossRef]
- Armanda, D.T.; Guinée, J.B.; Tukker, A. The second green revolution: Innovative urban agriculture’s contribution to food security and sustainability—A review. Glob. Food Secur. 2019, 22, 13–24. [Google Scholar] [CrossRef]
- Arora, N.K. Agricultural sustainability and food security. Environ. Sustain. 2018, 1, 217–219. [Google Scholar] [CrossRef]
- Arora, N.K.; Fatima, T.; Mishra, I.; Verma, S. Microbe-based Inoculants: Role in Next Green Revolution. In Environmental Concerns and Sustainable Development; Metzler, J.B., Ed.; Springer Singapore: Singapore, 2019; Volume 1, pp. 191–246. [Google Scholar]
- Harwood, J. Could the adverse consequences of the green revolution have been foreseen? How experts responded to unwelcome evidence. Agroecol. Sustain. Food Syst. 2019, 44, 509–535. [Google Scholar] [CrossRef]
- Singh, Z.; Kaur, J.; Kaur, R.; Hundal, S.S. Toxic Effects of Organochlorine Pesticides: A Review. Am. J. Biosci. 2016, 4, 11. [Google Scholar] [CrossRef]
- Arora, N.K.; Tewari, S.; Singh, S.; Lal, N.; Maheshwari, D.K. PGPR for Protection of Plant Health Under Saline Conditions. In Bacteria in Agrobiology: Stress Management; Metzler, J.B., Ed.; Springer Singapore: Singapore, 2011; pp. 239–258. [Google Scholar]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef]
- Hungria, M.; Nogueira, M.A. Microrganismos e a sustentabilidade de sistemas agrícolas de alta produtividade. In FertBio 2016; SBCS: Goiânia, Brazil, 2016; ISBN 978-85-86504-15-0. [Google Scholar]
- Hungria, M.; Nogueira, M.A.; Araujo, R.S. Inoculation of Brachiaria spp. with the plant growth-promoting bacterium Azospirillum brasilense: An environment-friendly component in the reclamation of degraded pastures in the tropics. Agric. Ecosyst. Environ. 2016, 221, 125–131. [Google Scholar] [CrossRef]
- Llewellyn, D. Does Global Agriculture Need Another Green Revolution? Engineering 2018, 4, 449–451. [Google Scholar] [CrossRef]
- Malusá, E.; Vassilev, N. A contribution to set a legal framework for biofertilisers. Appl. Microbiol. Biotechnol. 2014, 98, 6599–6607. [Google Scholar] [CrossRef]
- Hungria, M.; Mendes, I.C. Nitrogen Fixation with Soybean: The Perfect Symbiosis? In Biological Nitrogen Fixation; Wiley: Hoboken, NJ, USA, 2015; pp. 1009–1024. [Google Scholar]
- Martin-Guay, M.-O.; Paquette, A.; Dupras, J.; Rivest, D. The new Green Revolution: Sustainable intensification of agriculture by intercropping. Sci. Total Environ. 2018, 615, 767–772. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.S.; Nogueira, M.A.; Hungria, M. Microbial inoculants: Reviewing the past, discussing the present and previewing an outstanding future for the use of beneficial bacteria in agriculture. AMB Express 2019, 9, 1–22. [Google Scholar] [CrossRef]
- FAOSTAT. 2019. Available online: http://www.fao.org/faostat/en/#home (accessed on 15 March 2021).
- Fishel, F.M. Pest Management and Pesticides: A Historical Perspective. Agronomy Department, UF/IFAS Extension, 2016. Available online: http://edis.ifas.ufl.edu (accessed on 16 November 2020).
- Hungria, M.; Nogueira, M.A.; Campos, L.J.M.; Menna, P.; Brandi, F.; Ramos, Y.G. Seed pre-inoculation with Bradyrhizobium as time-optimizing option for large-scale soybean cropping systems. Agron. J. 2020, 112, 5222–5236. [Google Scholar] [CrossRef]
- Anupama, B.; Nidhi, S.; Kiran, S.; Bikrol, A.; Saxena, N.; Singh, K. Response of Glycine max in relation to nitrogen fixation as influenced by fungicide seed treatment. Afr. J. Biotechnol. 2005, 4, 667–671. [Google Scholar] [CrossRef]
- Fox, J.E.; Gulledge, J.; Engelhaupt, E.; Burow, M.E.; McLachlan, J.A. Pesticides reduce symbiotic efficiency of nitrogen-fixing rhizobia and host plants. Proc. Natl. Acad. Sci. USA 2007, 104, 10282–10287. [Google Scholar] [CrossRef] [PubMed]
- Ahemad, M. Growth suppression of legumes in pyriproxyfen stressed soils: A comparative study. Emir. J. Food Agric. 2014, 26, 66–72. [Google Scholar] [CrossRef]
- Campo, R.J.; Araujo, R.S.; Hungria, M. Nitrogen fixation with the soybean crop in Brazil: Compatibility between seed treatment with fungicides and bradyrhizobial inoculants. Symbiosis 2009, 48, 154–163. [Google Scholar] [CrossRef]
- Rodrigues, T.F.; Bender, F.R.; Sanzovo, A.W.S.; Ferreira, E.; Nogueira, M.A.; Hungria, M. Impact of pesticides in properties of Bradyrhizobium spp. and in the symbiotic performance with soybean. World J. Microbiol. Biotechnol. 2020, 36, 172. [Google Scholar] [CrossRef]
- Santos, M.S.; Rondina, A.B.L.; Nogueira, M.A.; Hungria, M. Compatibility of Azospirillum brasilense with Pesticides Used for Treatment of Maize Seeds. Int. J. Microbiol. 2020, 2020, 1–8. [Google Scholar] [CrossRef]
- Santos, M.S.; Rodrigues, T.F.; Ferreira, E.; Megias, M.; Nogueira, M.A.; Hungria, M. Method for Recovering and Counting Viable Cells from Maize Seeds Inoculated with Azospirillum brasilense. J. Pure Appl. Microbiol. 2020, 14, 195–204. [Google Scholar] [CrossRef]
- Pereira, L.C.; De Carvalho, C.; Suzukawa, A.K.; Correia, L.V.; Pereira, R.C.; Dos Santos, R.F.; Braccini, A.L.; Osipi, E.A.F. Toxicity of seed-applied pesticides to Azospirillum spp.: An approach based on bacterial count in the maize rhizosphere. Seed Sci. Technol. 2020, 48, 241–246. [Google Scholar] [CrossRef]
- Mahmood, I.; Imadi, S.R.; Shazadi, K.; Gul, A.; Hakeem, K.R. Effects of Pesticides on Environment; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2016; pp. 253–269. [Google Scholar]
- Abubakar, Y.; Tijjani, H.; Egbuna, C.; Adetunji, C.O.; Kala, S.; Kryeziu, T.L.; Ifemeje, J.C.; Patrick-Iwuanyanwu, K.C. Pesticides, History, and Classification. In Natural Remedies for Pest, Disease and Weed Control; Elsevier BV: Amsterdam, The Netherlands, 2020; pp. 29–42. [Google Scholar]
- Costa, L.G.; Galli, C.L.; Murphy, S.D. Toxicology of Pesticides: Experimental, Clinical and Regulatory Aspects, 1st ed.; Springer: Berlin/Heidelberg, Germany, 1987. [Google Scholar]
- Carson, R. Silent Spring, 1st ed.; Houghton Mifflin: Boston, MA, USA, 1962; p. 368. [Google Scholar]
- Stern, V.M.; Smith, R.F.; Van Der Bosch, R.; Hagen, R.S. The integrated control concept. Hilgardia 1959, 29, 81–101. [Google Scholar] [CrossRef]
- Moura, A.P. Manejo Integrado de Pragas: Estratégias e Táticas de Manejo Para o Controle de Insetos e Ácaros-Praga em Hortaliças; Embrapa Hortaliças: Brasília, Brazil, 2015; p. 28. [Google Scholar]
- Donley, N. The USA lags behind other agricultural nations in banning harmful pesticides. Environ. Health 2019, 18, 1–12. [Google Scholar] [CrossRef]
- Kim, K.-H.; Kabir, E.; Jahan, S.A. Exposure to pesticides and the associated human health effects. Sci. Total Environ. 2017, 575, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kumar, V.; Shahzad, B.; Tanveer, M.; Sidhu, G.P.S.; Handa, N.; Kohli, S.K.; Yadav, P.; Bali, A.S.; Parihar, R.D.; et al. Worldwide pesticide usage and its impacts on ecosystem. SN Appl. Sci. 2019, 1, 1446. [Google Scholar] [CrossRef]
- Rani, L.; Thapa, K.; Kanojia, N.; Sharma, N.; Singh, S.; Grewal, A.S.; Srivastav, A.L.; Kaushal, J. An extensive review on the consequences of chemical pesticides on human health and environment. J. Clean. Prod. 2021, 283, 124657. [Google Scholar] [CrossRef]
- Williams, P.M. Current Use of Legume Inoculant Technology. In Biological Nitrogen Fixation; Metzler, J.B., Ed.; Springer: Berlin/Heidelberg, Germany, 1984; pp. 173–200. [Google Scholar]
- Smith, R.S. Legume inoculant formulation and application. Can. J. Microbiol. 1992, 38, 485–492. [Google Scholar] [CrossRef]
- Hungria, M.; Campo, R.J. Economical and Environmental Benefits of Inoculation and Biological Nitrogen Fixation with Soybean: Situation in South America. Available online: http://ainfo.cnptia.embrapa.br/digital/bitstream/item/173978/1/ID-23469-pt-1.pdf (accessed on 1 April 2021).
- Hungria, M.; Campo, R.J.; Mendes, I.C. A Importância do Processo de Fixação Biológica do Nitrogênio para a Cultura da Soja: Componente Essencial Para a Competitividade do Produto Brasileiro; Documentos, 283; Embrapa Soja: Londrina, Brazil, 2007; p. 80. [Google Scholar]
- Ormeño-Orrillo, E.; Hungria, M.; Martínez-Romero, E. Dinitrogen-fixing prokaryotes. In The Prokaryotes—Prokaryotic Physiology and Biochemistry; Rosemberg, E., De Long, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 427–451. [Google Scholar] [CrossRef]
- Hungria, M.; Campo, R.J.; Souza, E.M.; Pedrosa, F.O. Inoculation with selected strains of Azospirillum brasilense and A. lipoferum improves yields of maize and wheat in Brazil. Plant Soil 2010, 331, 413–425. [Google Scholar] [CrossRef]
- Gomes, D.F.; Ormeño-Orrillo, E.; Hungria, M. Biodiversity, symbiotic efficiency and genomics of Rhizobium tropici and related species. In Biological Nnitrogen Fixation; De Bruijn, F.J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 747–756. [Google Scholar]
- Zhang, Y.J.; Zheng, W.T.; Everall, I.; Young, J.P.W.; Zhang, X.X.; Tian, C.F.; Sui, X.H.; Wang, E.T.; Chen, W.X. Rhizobium anhuiense sp. nov., isolated from effective nodules of Vicia faba and Pisum sativum. Int. J. Syst. Evol. Microbiol. 2015, 65, 2960–2967. [Google Scholar] [CrossRef] [PubMed]
- Mercante, F.M.; Otsubo, A.A.; Brito, O.R. New Native Rhizobia Strains for Inoculation of Common Bean in the Brazilian Savanna. Rev. Bras. Ciência Solo 2017, 41, 1–11. [Google Scholar] [CrossRef]
- Pandey, R.P.; Srivastava, A.K.; Gupta, V.K.; O’Donovan, A.; Ramteke, P.W. Enhanced yield of diverse varieties of chickpea (Cicer arietinum L.) by different isolates of Mesorhizobium ciceri. Environ. Sustain. 2018, 1, 425–435. [Google Scholar] [CrossRef]
- Daur, I.; Saad, M.M.; Eida, A.A.; Ahmad, S.; Shah, Z.H.; Ihsan, M.Z.; Muhammad, Y.; Sohrab, S.S.; Hirt, H. Boosting Alfalfa (Medicago sativa L.) Production with Rhizobacteria from Various Plants in Saudi Arabia. Front. Microbiol. 2018, 9, 477. [Google Scholar] [CrossRef] [PubMed]
- Mariana, S.S.; Mariangela, H.; Marco, A.N.; Santos, M.S.; Hungria, M.; Nogueira, M.A. Production of polyhydroxybutyrate (PHB) and biofilm by Azospirillum brasilense aiming at the development of liquid inoculants with high performance. Afr. J. Biotechnol. 2017, 16, 1855–1862. [Google Scholar] [CrossRef]
- Gundi, J.S.; Santos, M.S.; Oliveira, A.L.M.; Nogueira, M.A.; Hungria, M. Development of liquid inoculants for strains of Rhizobium tropici group using response surface methodology. Afr. J. Biotechnol. 2018, 17, 411–421. [Google Scholar] [CrossRef]
- Hungria, M.; Andrade, D.D.S.; De O’Chueire, L.M.; Probanza, A.; Guttierrez-Mañero, F.J.; Megías, M. Isolation and characterization of new efficient and competitive bean (Phaseolus vulgaris L.) rhizobia from Brazil. Soil Biol. Biochem. 2000, 32, 1515–1528. [Google Scholar] [CrossRef]
- Hungria, M.; Loureiro, M.F.; Mendes, I.C.; Campo, R.J.; Graham, P.H. Inoculant Preparation, Production and Application. In Nitrogen Fixation in Agriculture, Forestry, Ecology, and the Environment; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2005; pp. 223–253. [Google Scholar]
- Bashan, Y.; De-Bashan, L.E.; Prabhu, S.R.; Hernandez, J.-P. Advances in plant growth-promoting bacterial inoculant technology: Formulations and practical perspectives (1998–2013). Plant Soil 2014, 378, 1–33. [Google Scholar] [CrossRef]
- Cassán, F.; Okon, Y.; Creus, C.M. Handbook for Azospirillum: Technical Issues and Protocols; Springer International Publishing: London, UK, 2015; ISBN 978-3-319-06541-0. [Google Scholar]
- Campo, R.J.; Araujo, R.S.; Mostasso, F.L.; Hungria, M. In-furrow inoculation of soybean as alternative to fungicide and micronutrient seed treatment. Rev. Bras. Ciência Solo 2010, 34, 1103–1112. [Google Scholar] [CrossRef]
- Fukami, J.; Nogueira, M.A.; Araujo, R.S.; Hungria, M. Accessing inoculation methods of maize and wheat with Azospirillum brasilense. AMB Express 2016, 6, 1–13. [Google Scholar] [CrossRef]
- Moretti, L.G.; Lazarini, E.; Bossolani, J.W.; Parente, T.L.; Caioni, S.; Araujo, R.S.; Hungria, M. Can additional inoculations increase soybean nodulation and grain yield? Agron. J. 2018, 110, 715–721. [Google Scholar] [CrossRef]
- Drew, E.; Herridge, D.; Ballard, R.; O’hara, G.; Deaker, R.; Denton, M.; Yates, R.; Gemell, G.; Hartley, E.; Phillips, L.; et al. Inoculating Legumes: A Practical Guide, 2nd ed.; Grains Research and Development Corporation: Barton, Australian, 2009; p. 72. ISBN 978-1-921779-45-9. [Google Scholar]
- MAPA—Ministério da Agricultura, Pecuária e Abastecimento. Instrução Normativa No 5, de 6 de Agosto de 2004. Diário Oficial da União da República Federativa do Brasil, 6 August 2004.
- MAPA—Ministério da Agricultura, Pecuária e Abastecimento. Instrução Normativa No 13, de 24 de Março de 2011. Diário Oficial da União da República Federativa do Brasil, 24 March 2011.
- Hungria, M.; Campo, R.J. Inoculantes microbianos: Situação no Brasil. In Biofertilizantes en Iberoamérica: Visión Técnica, Científica y Empresarial; Izaguirre-Mayoral, M.L., Labandera, C., Sanjuan, J., Eds.; Cyted/Biofag: Montevideo, Uruguay, 2007; pp. 22–31. [Google Scholar]
- Herrmann, L.; Lesueur, D. Challenges of formulation and quality of biofertilizers for successful inoculation. Appl. Microbiol. Biotechnol. 2013, 97, 8859–8873. [Google Scholar] [CrossRef]
- Deaker, R.; Hartley, E.; Gemell, G.; Herridge, D.F.; Karanja, N. Inoculant production and quality control. In Working with Rhizobia; Howieson, J.G., Dilworth, J.G., Eds.; ACIAR: Canberra, Australia, 2016; pp. 167–186. [Google Scholar]
- ANPII, Associação Nacional dos Produtores e Importadores de Inoculantes. Estatísticas. 2020. Available online: http://www.anpii.org.br/estatisticas/ (accessed on 22 November 2020).
- Santos, M.S.; Nogueira, M.A.; Hungria, M. Outstanding impact of Azospirillum brasilense strains Ab-V5 and Ab-V6 on the Brazilian agriculture: Lessons that farmers are receptive to adopt new microbial inoculants. Rev. Bras. Ciência Solo 2021, 45, e0200128. [Google Scholar] [CrossRef]
- Hungria, M.; Nogueira, M.A.; Araujo, R.S. Co-inoculation of soybeans and common beans with rhizobia and azospirilla: Strategies to improve sustainability. Biol. Fertil. Soils 2013, 49, 791–801. [Google Scholar] [CrossRef]
- Hungria, M.; Nogueira, M.A.; Araujo, R.S. Soybean Seed Co-Inoculation with Bradyrhizobium spp. and Azospirillum brasilense: A New Biotechnological Tool to Improve Yield and Sustainability. Am. J. Plant Sci. 2015, 6, 811–817. [Google Scholar] [CrossRef]
- Bashan, Y.; De-Bashan, L.E. How the Plant Growth-Promoting Bacterium Azospirillum Promotes Plant Growth—A Critical Assessment. Adv. Agron. 2010, 108, 77–136. [Google Scholar] [CrossRef]
- Cerezini, P.; Kuwano, B.H.; dos Santos, M.B.; Terassi, F.; Hungria, M.; Nogueira, M.A. Strategies to promote early nodulation in soybean under drought. Field Crop. Res. 2016, 196, 160–167. [Google Scholar] [CrossRef]
- Mahanty, T.; Bhattacharjee, S.; Goswami, M.; Bhattacharyya, P.; Das, B.; Ghosh, A.; Tribedi, P. Biofertilizers: A potential approach for sustainable agriculture development. Environ. Sci. Pollut. Res. 2016, 24, 3315–3335. [Google Scholar] [CrossRef] [PubMed]
- Fukami, J.; Cerezini, P.; Hungria, M. Azospirillum: Benefits that go far beyond biological nitrogen fixation. AMB Express 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Silva, E.R.; Zoz, J.; Oliveira, C.E.S.; Zuffo, A.M.; Steiner, F.; Zoz, T.; Vendruscolo, E.P. Can co-inoculation of Bradyrhizobium and Azospirillum alleviate adverse effects of drought stress on soybean (Glycine max L. Merrill.)? Arch. Microbiol. 2019, 201, 325–335. [Google Scholar] [CrossRef]
- Aeron, A.; Khare, E.; Jha, C.K.; Meena, V.S.; Aziz, S.M.A.; Islam, M.T.; Kim, K.; Meena, S.K.; Pattanayak, A.; Rajashekara, H.; et al. Revisiting the plant growth-promoting rhizobacteria: Lessons from the past and objectives for the future. Arch. Microbiol. 2019, 202, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Vishwakarma, K.; Kumar, N.; Shandilya, C.; Mohapatra, S.; Bhayana, S.; Varma, A. Revisiting Plant–Microbe Interactions and Microbial Consortia Application for Enhancing Sustainable Agriculture: A Review. Front. Microbiol. 2020, 11, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Cassán, F.; López, G.; Nievas, S.; Coniglio, A.; Torres, D.; Donadio, F.; Molina, R.; Mora, V. What Do We Know About the Publications Related with Azospirillum? A Metadata Analysis. Microb. Ecol. 2021, 81, 278–281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hussain, S.; Zhao, F.; Zhu, L.; Cao, X.; Yu, S.; Jin, Q. Effects of Azospirillum brasilense and Pseudomonas fluorescens on nitrogen transformation and enzyme activity in the rice rhizosphere. J. Soils Sediments 2018, 18, 1453–1465. [Google Scholar] [CrossRef]
- Thirumal, G.; Reddy, R.S.; Triveni, S.; Nagaraju, Y.; Prasannakumar, B. Screening of Native Rhizobia and Pseudomonas Strains for Plant Growth Promoting Activities. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 616–625. [Google Scholar] [CrossRef]
- Sandini, I.E.; Pacentchuk, F.; Hungria, M.; Nogueira, M.A.; Cruz, S.P.; Nakatani, A.S.; Araujo, R.S. Seed inoculation with Pseudomonas fluorescens promotes growth, yield and reduces nitrogen application in maize. Int. J. Agric. Biol. 2019, 22, 1369–1375. [Google Scholar] [CrossRef]
- Araujo, F.F.; Guaberto, L.M.; Silva, I.F. Bioprospecção de bactérias promotoras de crescimento em Brachiaria brizantha. Rev. Bras. Zootec. 2012, 41, 521–527. [Google Scholar] [CrossRef]
- Ribeiro, V.P.; Marriel, I.E.; De Sousa, S.M.; Lana, U.G.D.P.; Mattos, B.B.; De Oliveira, C.A.; Gomes, E.A. Endophytic Bacillus strains enhance pearl millet growth and nutrient uptake under low-P. Braz. J. Microbiol. 2018, 49, 40–46. [Google Scholar] [CrossRef]
- O’Callaghan, M. Microbial inoculation of seed for improved crop performance: Issues and opportunities. Appl. Microbiol. Biotechnol. 2016, 100, 5729–5746. [Google Scholar] [CrossRef]
- Nogueira, M.A.; Prando, A.M.; Oliveira, A.B.; Lima, D.; Conte, O.; Harger, N.; Oliveira, F.T.; Hungria, M. Ações de Transferência de Tecnologia em Inoculação/Coinoculação com Bradyrhizobium e Azospirillum na Cultura da Soja na Safra 2017/18 no Estado do Paraná; Embrapa Soja: Londrina, Brazil, 2018; p. 15. [Google Scholar]
- de Souza, J.E.B.; Ferreira, E.P.D.B. Improving sustainability of common bean production systems by co-inoculating rhizobia and azospirilla. Agric. Ecosyst. Environ. 2017, 237, 250–257. [Google Scholar] [CrossRef]
- Reis, V.M. Uso de Bactérias Fixadores de Nitrogênio Como Inoculante Para Aplicação em Gramíneas; Embrapa Agrobiologia: Seropédica, Brazil, 2007; p. 22. [Google Scholar]
- Galindo, F.S.; Filho, M.C.M.T.; Buzetti, S.; Rodrigues, W.L.; Santini, J.M.K.; Alves, C.J. Nitrogen fertilisation efficiency and wheat grain yield affected by nitrogen doses and sources associated with Azospirillum brasilense. Acta Agric. Scand. Sect. B Plant Soil Sci. 2019, 69, 606–617. [Google Scholar] [CrossRef]
- Fukami, J.; Ollero, F.J.; Megías, M.; Hungria, M. Phytohormones and induction of plant-stress tolerance and defense genes by seed and foliar inoculation with Azospirillum brasilense cells and metabolites promote maize growth. AMB Express 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Pereg, L.; De-Bashan, L.E.; Bashan, Y. Assessment of affinity and specificity of Azospirillum for plants. Plant Soil 2016, 399, 389–414. [Google Scholar] [CrossRef]
- Racca, R.; González, N. Nutrición nitrogenada de la alfalfa e impacto de la fijación simbiótica del nitrógeno. In El Cultivo de la Alfalfa en la Argentina; Gasigalup, D.H., Ed.; Ediciones INTA: Buenos Aires, Argentina, 2007; pp. 69–79. [Google Scholar]
- Buntić, A.V.; Stajković-Srbinović, O.S.; Knežević, M.M.; Kuzmanović, Đ.Ž.; Rasulić, N.V.; Delić, D.I. Development of liquid rhizobial inoculants and pre-inoculation of alfalfa seeds. Arch. Biol. Sci. 2019, 71, 379–387. [Google Scholar] [CrossRef]
- Leite, R.C.; Santos, A.C.; Dos Santos, J.G.D.; Leite, R.C.; Oliveira, L.B.T.; Hungria, M. Mitigation of mombasa grass (Me-gathyrsus maximus) dependence on nitrogen fertilization as a function of inoculation with Azospirillum brasilense. Rev. Bras. Ciên. Solo. 2019, 43, 180–234. [Google Scholar] [CrossRef]
- Heinrichs, R.; Universidade Estadual Paulista; Meirelles, G.C.; Santos, L.F.D.M.; Lira, M.V.D.S.; Lapaz, A.D.M.; Nogueira, M.A.; Bonini, C.D.S.B.; Filho, C.V.S.; Moreira, A.; et al. Azospirillum inoculation of ’Marandu’ palisade grass seeds: Effects on forage production and nutritional status. Semin. Ciências Agrárias 2020, 41, 465–478. [Google Scholar] [CrossRef]
- Steinfeld, H.; Gerber, P.; Wassenaar, T.; Castel, V.; Rosales, M.; De Haan, C. Livestock’s Long Shadow: Environmental Issues and Options, 1st ed.; United Nations Food and Agriculture Organization: Rome, Italy, 2006. [Google Scholar]
- Fonte, S.J.; Nesper, M.; Hegglin, D.; Velásquez, J.E.; Ramirez, B.; Rao, I.M.; Bernasconi, S.M.; Bünemann, E.K.; Frossard, E.; Oberson, A. Pasture degradation impacts soil phosphorus storage via changes to aggregate-associated soil organic matter in highly weathered tropical soils. Soil Biol. Biochem. 2014, 68, 150–157. [Google Scholar] [CrossRef]
- Braccini, A.; Mariucci, G.; Suzukawa, A.; Lima, L.; Piccinin, G. Co-inoculação e Modos de Aplicação de Bradyrhizobium japonicum e Azospirillum brasilense e Adubação Nitrogenada na Nodulação das Plantas e Rendimento da Cultura da Soja. Sci. Agrar. Parana. 2016, 15, 27–35. [Google Scholar] [CrossRef]
- Garcia, A. Fungicidas I: Utilização no Controle Químico de Doenças e sua Ação contra os Fitopatógenos; Embrapa Rondônia: Porto Velho, Brazil, 1999; p. 34. [Google Scholar]
- Curley, R.L.; Burton, J.C. Compatibility of Rhizobium japonicum with Chemical Seed Protectants 1. Agron. J. 1975, 67, 807–808. [Google Scholar] [CrossRef]
- Leterme, P.; Revellin, C.; Catroux, G. Effect of some fungicide seed treatments on the survival of Bradyrhizobium japonicum and on the nodulation and yield of soybean [Glycine max. (L) Merr.]. Biol. Fertil. Soils 1993, 16, 211–214. [Google Scholar] [CrossRef]
- Cattelan, A.J.; Hungria, M. Nitrogen nutrition and inoculation. In Tropical Soybean, Improvement and Production; FAO, Plant Production and Protection Series, No. 27; Embrapa-CNPSo: Londrina, Brazil, 1994; pp. 201–215. [Google Scholar]
- Campo, R.J.; Hungria, M. Compatibilidade de Uso de Inoculantes e Fungicidas no Tratamento de Sementes de Soja; Embrapa Soja: Londrina, Brazil, 2000; p. 31. [Google Scholar]
- Zilli, J.É.; Ribeiro, K.G.; Campo, R.J.; Hungria, M. Influence of fungicide seed treatment on soybean nodulation and grain yield. Rev. Bras. Ciência Solo 2009, 33, 917–923. [Google Scholar] [CrossRef]
- Gomes, Y.C.B.; Valadão, F.C.D.A.; Dalchiavon, F.C. Joint use of fungicides, insecticides and inoculants in the treatment of soybean seeds. Rev. Ceres 2017, 64, 258–265. [Google Scholar] [CrossRef][Green Version]
- Schulz, T.J.; Thelen, K.D. Soybean Seed Inoculant and Fungicidal Seed Treatment Effects on Soybean. Crop. Sci. 2008, 48, 1975–1983. [Google Scholar] [CrossRef]
- Ahmed, T.H.M.; Elsheikh, E.A.E.; Mahdi, A.A. The in vitro compatibility of some Rhizobium and Bradyrhizobium strains with fungicides. Afr. Crop. Sci. Conf. Proc. 2007, 8, 1171–1178. [Google Scholar] [CrossRef]
- Rathjen, J.; Ryder, M.; Riley, I.; Lai, T.; Denton, M. Impact of seed-applied pesticides on rhizobial survival and legume nodulation. J. Appl. Microbiol. 2020, 129, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Madhaiyan, M.; Poonguzhali, S.; Hári, K.; Saravanan, V.; Sa, T. Influence of pesticides on the growth rate and plant-growth promoting traits of Gluconacetobacter diazotrophicus. Pestic. Biochem. Physiol. 2006, 84, 143–154. [Google Scholar] [CrossRef]
- Fernandes, M.F.; Procópio, S.D.O.; Teles, D.A.; Filho, J.G.D.S.; Filho, A.C.; Machado, T.N. Toxicidade de inseticidas utilizados na cultura da cana-de-açúcar à bactéria diazotrófica Herbaspirillum seropedicae. Rev. Ciências Agrar. Amaz. J. Agric. Environ. Sci. 2012, 55, 318–326. [Google Scholar] [CrossRef]
- Insecticidal efficacy of lichens and their metabolites—A mini review. J. Appl. Pharm. Sci. 2018, 8, 159–164. [CrossRef]
- Nauen, R.; Slater, R.; Sparks, T.C.; Elbert, A.; McCaffery, A. IRAC: Insecticide Resistance and Mode-of-action Classification of Insecticides. In Modern Crop Protection Compounds; Wiley: Weinheim, Germany, 2019; pp. 995–1012. [Google Scholar]
- De Souza, C.P.; Guedes, T.D.A.; Fontanetti, C.S. Evaluation of herbicides action on plant bioindicators by genetic biomarkers: A review. Environ. Monit. Assess. 2016, 188, 694. [Google Scholar] [CrossRef]
- Martinez, D.A.; Loening, U.E.; Graham, M.C. Impacts of glyphosate-based herbicides on disease resistance and health of crops: A review. Environ. Sci. Eur. 2018, 30, 1–14. [Google Scholar] [CrossRef]
- Barros, V.M.D.S.; Pedrosa, J.L.F.; Gonçalves, D.R.; De Medeiros, F.C.L.; Carvalho, G.R.; Gonçalves, A.H.; Teixeira, P.V.V.Q. Herbicides of biological origin: A review. J. Hortic. Sci. Biotechnol. 2020, 1–9. [Google Scholar] [CrossRef]
- Wilms, W.; Woźniak-Karczewska, M.; Syguda, A.; Niemczak, M.; Ławniczak, Ł.; Pernak, J.; Rogers, R.D.; Chrzanowski, L. Herbicidal ionic liquids—A promising future for old herbicides? Review on synthesis, toxicity, biodegradation and efficacy studies. J. Agric. Food Chem. 2020, 68, 10456–10488. [Google Scholar] [CrossRef]
- Adegas, F.S.; Vergas, L.; Gazziero, D.L.P.; Karam, D. Impacto Econômico da Resistência de Plantas Daninhas a Herbicidas no Brasil; Embrapa Soja: Londrina, Brazil, 2017; p. 12. [Google Scholar]
- Angelini, J.; Silvina, G.; Taurian, T.; Ibáñez, F.; Tonelli, M.L.; Valetti, L.; Anzuay, M.S.; Ludueña, L.; Muñoz, V.; Fabra, A. The effects of pesticides on bacterial nitrogen fixers in peanut-growing area. Arch. Microbiol. 2013, 195, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Dash, N.P.; Kumar, A.; Kaushik, M.S.; Abraham, G.; Singh, P.K. Agrochemicals influencing nitrogenase, biomass of N2-fixing cyanobacteria and yield of rice in wetland cultivation. Biocatal. Agric. Biotechnol. 2017, 9, 28–34. [Google Scholar] [CrossRef]
- Procópio, S.O.; Santos, J.B.; Jacques, R.J.S.; Kasuya, M.C.M.; Silva, A.A.; Werlang, R.C. crescimento de estirpes de Bradyrhizobium sob influência dos herbicidas glyphosate potássico, fomesafen, imazethapyr e carfentrazone-ethyl. Rev. Ceres. 2004, 51, 179–188. [Google Scholar]
- Drouin, P.; Sellami, M.; Prévost, D.; Fortin, J.; Antoun, H. Tolerance to agricultural pesticides of strains belonging to four genera ofRhizobiaceae. J. Environ. Sci. Health Part B 2010, 45, 757–765. [Google Scholar] [CrossRef] [PubMed]
- King, C.; Purcell, L.C.; Vories, E.D. Plant Growth and Nitrogenase Activity of Glyphosate-Tolerant Soybean in Response to Foliar Glyphosate Applications. Agron. J. 2001, 93, 179–186. [Google Scholar] [CrossRef]
- Hungria, M.; Mendes, I.C.; Nakatani, A.S.; Dos Reis-Junior, F.B.; Morais, J.Z.; De Oliveira, M.C.N.; Fernandes, M.F. Effects of the glyphosate-resistance gene and herbicides on soybean: Field trials monitoring biological nitrogen fixation and yield. Field Crop. Res. 2014, 158, 43–54. [Google Scholar] [CrossRef]
- Hungria, M.; Nakatani, A.S.; Souza, R.A.; Sei, F.B.; Chueire, L.M.D.O.; Arias, C.A. Impact of the ahas transgene for herbicides resistance on biological nitrogen fixation and yield of soybean. Transgenic Res. 2014, 24, 155–165. [Google Scholar] [CrossRef]
- Zablotowicz, R.M.; Reddy, K.N. Nitrogenase activity, nitrogen content, and yield responses to glyphosate in glyphosate-resistant soybean. Crop. Prot. 2007, 26, 370–376. [Google Scholar] [CrossRef]
- Zobiole, L.H.; Kremer, R.J.; Oliveira, R.S.; Constantin, J. Glyphosate affects chlorophyll, nodulation and nutrient accumulation of “second generation” glyphosate-resistant soybean (Glycine max L.). Pestic. Biochem. Physiol. 2011, 99, 53–60. [Google Scholar] [CrossRef]
- Liu, C.-M.; McLean, P.A.; Sookdeo, C.C.; Cannon, F.C. Degradation of the Herbicide Glyphosate by Members of the Family Rhizobiaceae. Appl. Environ. Microbiol. 1991, 57, 1799–1804. [Google Scholar] [CrossRef]
- Sene, L.; Converti, A.; Secchi, G.A.R.; Simão, R.D.C.G. New aspects on atrazine biodegradation. Braz. Arch. Biol. Technol. 2010, 53, 487–496. [Google Scholar] [CrossRef]
- Hungria, M.; Nogueira, M.A. Tecnologias de inoculação da cultura da soja: Mitos, verdades e desafios. In Boletim de Pesquisa 2019/2020; Fundação MT: Rondonópolis, Brazil, 2019; pp. 50–62. [Google Scholar]
- Mourgues, F.; Brisset, M.-N.; Chevreau, E. Strategies to improve plant resistance to bacterial diseases through genetic engineering. Trends Biotechnol. 1998, 16, 203–210. [Google Scholar] [CrossRef]
- Molinari, S. Natural genetic and induced plant resistance, as a control strategy to plant-parasitic nematodes alternative to pesticides. Plant Cell Rep. 2010, 30, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lubberstedt, T.; Xu, M. The Genetic and Molecular Basis of Plant Resistance to Pathogens. J. Genet. Genom. 2013, 40, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Ahemad, M.; Khan, M.S. Comparative toxicity of selected insecticides to pea plants and growth promotion in response to insecticide-tolerant and plant growth promoting Rhizobium leguminosarum. Crop. Prot. 2010, 29, 325–329. [Google Scholar] [CrossRef]
- Kadouri, D.; Jurkevitch, E.; Okon, Y. Involvement of the Reserve Material Poly-β-Hydroxybutyrate in Azospirillum brasilense Stress Endurance and Root Colonization. Appl. Environ. Microbiol. 2003, 69, 3244–3250. [Google Scholar] [CrossRef]
- Bhat, S.G.; Subin, R.S. Bacterial polyhydroxyalkanoates production and its applications. In Microbial Bioproducts; Bhat, S.G., Nambisan, P., Eds.; Directorate of Public Relation and Publication: Kerala, India, 2015; pp. 70–96. [Google Scholar]
- Fukami, J.; Abrantes, J.L.F.; Del Cerro, P.; Nogueira, M.A.; Ollero, F.J.; Megias, M.; Hungria, M. Revealing strategies of quorum sensing in Azospirillum brasilense strains Ab-V5 and Ab-V6. Arch. Microbiol. 2017, 200, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Mariangela, H.; Marco, A.N.; Ricardo, S.A. Alternative methods of soybean inoculation to overcome adverse conditions at sowing. Afr. J. Agric. Res. 2015, 10, 2329–2338. [Google Scholar] [CrossRef][Green Version]
- Correia, L.V.; Felber, P.H.; Pereira, L.C.; Braccini, A.L.; Carvalho, D.U.; Da Cruz, M.A.; Matera, T.C.; Santos, R.F.; Marteli, D.C.V.; Osipi, E.A.F. Inoculation of Wheat with Azospirillum spp.: A Comparison Between Foliar and In-furrow Applications. J. Agric. Sci. 2019, 12, 194. [Google Scholar] [CrossRef]
- DunhamTrimmer® Global Biocontrol Market Overview. Trends, Drivers & Insights, 2019. DunhamTrimmer®, Florida-USA. Available online: http://dunhamtrimmer.com/wp-content/uploads/2019/05/TOCDT_Global_Biocontrol_Overview_Links.pdf (accessed on 2 February 2021).



{kind=link}
{kind=link}
| Pesticide | Concentration | Microorganism | Effect | Reference |
|---|---|---|---|---|
| Monocrotophos i, Malathion i, Chlorpyripho i, Dichlorvos i, Lindano i e Endosulphan i | Recommended dose of each product | Gluconacetobacter diazotrophicus | With the exception of Malathion, all insecticides reduced cell viability. Nitrogenase activity was totally inhibited by Monocrotophos, Dichlorvos and Lindano. Production of IAA and GA3, and solubilization of P and Zn were impaired | [106] |
| Butachlor h, Alachlor h Atrazine h and 2,4-D h | Recommended dose of each product | Cell growth was hindered by 2,4-D. All herbicides reduced the activity of nitrogenase, the production of IAA and GA3, and the solubilization of P and Zn | ||
| Captan f, Thiram f, Luxan f and Fernasan-D f | 1000 µg L−1 | Bradyrhizobium sp. and Rhizobium sp. | Decreased colony diameter and inhibited growth in areas close to the fungicide application site | [104] |
| Benomyl f, Captan f, Carbendazin f, Carboxin f, Difenoconazole f, Thiabendazole f, Thiram f, Tolylfluanid f | Recommended dose for soybean | B. elkanii and B. japonicum | All fungicides caused mortality of microorganisms | [24] |
| Imidacloprid i, Fipronil i, Thiamethoxam i, Endosulphan i e Carbofuran i | 250 g ha−1, 400 g ha−1, 480 g ha−1, 2.800 g ha−1, 1.650 g ha−1, respectively | Herbaspirillum seropedicae | Endosulphan increased the lag phase. Carbofuran increased generation time and reduced lag phase | [107] |
| Pyraclostrobin f, thiophanato-methyl f e fipronil i | 2 mL kg−1 maize seed | A. brasilense (strains Ab-V5 and Ab-V6) | Drastic reduction in cell concentration 24 h after inoculation in treated seeds | [26] |
| Carbendazin f + Thiram f | 40–60 mL 20 kg−1 maize seed | Drastic reduction in cell concentration 24 h after inoculation in treated seeds | [27] | |
| metalaxil-m + fludioxonil + tiametoxame + abamectin | Recommended dose for maize | Drastic reduction in cell concentration 12 h after inoculation in treated seeds | [28] | |
| Thiram f, Thiram + Thiabendazole f and PPT f | >200 µg L−1 | Rhizobium leguminosarum bv. viciae | Formation of growth inhibition halos greater than 10 mm around the fungicide | [105] |
| Imidacloprid i | 0, 100, 200 e 300 µg L−1 | Formation of growth inhibition halos greater than 10 mm around the insecticide for all concentrations evaluated | ||
| Pyraclostrobin f, thiophanato-methyl f e fipronil i | Recommended dose for soybean | B. elkanii and B. japonicum | Drastic decrease in cell concentration after 7 days of exposure. Colony formation with smaller diameter | Rodrigues et al. [25] |
| Culture | Fungicide | Microorganism | local | Effect | Reference |
|---|---|---|---|---|---|
| Soybean (Glycine max) | Thiram f | B. japonicum | Greenhouse | Lower nodule number, nodule dry weight and nitrogenase activity | [21] |
| Benomyl f, Captan f, Carbendazin f, Carboxin f, Difenoconazole f, Thiabendazole f, Thiram, Tolylfluanid f | B. elkanii (strain SEMIA 5019) + B. japonicum (strain SEMIA 5079) | Greenhouse and field | Reduction in the number of nodules and in the total N in grains | [24] | |
| Carbendazin f + Thiram f; Carboxin f + Thiram f | B. elkanii (strains 5019 + 587), B. japonicum (strain 5079) + B. diazoefficiens (strain SEMIA 5080) | Field | Reduction of nodulation efficiency. Reduction of N content and grain yield to SEMIA 587 with Carbendazin + Thiram | [101] | |
| Carbendazin f + Thiram f | B. japonicum (strain 5079) + B. diazoefficiens (strain SEMIA 5080) | Field | Reduction in the number of pods per plant and grains per plant | [102] | |
| Mefenoxam f + Fludioxonil f | B. japonicum | Field | Reduced grain yield and protein and oleic acid content | [103] | |
| Pyraclostrobin f, thiophanato-methyl f and fipronil i | B. japonicum (strain 5079) and B. diazoefficiens (strain 5080) | Field | Less N accumulation in grains | [25] | |
| Alfafa (Medicago sativa) | Methyl parathion i, DDT i e pentachlorophenol i | Sinorhizobium meliloti, | Greenhouse | Reduction of nitrogenase activity, number of nodules and plant dry weight | [22] |
| Peanut (Arachis hypogaea). | Imazetapir h, Imazapic h, S-metachloro h, Dichlosulam h and Glyphosate h | Diazotrophic bacteria present in the soil | Greenhouse | Reduction of cell concentration of free and symbiotic diazotrophic bacteria | [115] |
| Field | Reduced cell concentration of free and symbiotic diazotrophic bacteria and reduced nitrogenase activity except for glyphosate | ||||
| Chickpeas (Cicer arietinum L.), pea (Pisum sativum L.), Mung beans (Vigna radiata L. Wiclzek) and lentil (Lens esculentus, = Lens culinaris Medik). | Pyriproxyfen i | Bacteria commonly present in the soil used | Pots in the field | Reduction in the number of nodules, in the dry weight of nodules, in the concentration of N in roots and in protein concentration in the grains | [23] |
| Rice (Oryza sativa) | Benthicarb h | Cyanobacteria naturally found in the rice paddy soil | Field | Decreased cell growth, nitrogenase activity and Naccumulation | [116] |
| Maize (Zea mays) | Pyraclostrobin f, thyophanato-methyl f and fipronil i | A. brasilense (strains Ab-V5 and Ab-V6) | Greenhouse | Fewer branch roots, root hair and shorter root hair length | [26] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, M.S.; Rodrigues, T.F.; Nogueira, M.A.; Hungria, M. The Challenge of Combining High Yields with Environmentally Friendly Bioproducts: A Review on the Compatibility of Pesticides with Microbial Inoculants. Agronomy 2021, 11, 870. https://doi.org/10.3390/agronomy11050870
Santos MS, Rodrigues TF, Nogueira MA, Hungria M. The Challenge of Combining High Yields with Environmentally Friendly Bioproducts: A Review on the Compatibility of Pesticides with Microbial Inoculants. Agronomy. 2021; 11(5):870. https://doi.org/10.3390/agronomy11050870
Chicago/Turabian StyleSantos, Mariana Sanches, Thiago Fernandes Rodrigues, Marco Antonio Nogueira, and Mariangela Hungria. 2021. "The Challenge of Combining High Yields with Environmentally Friendly Bioproducts: A Review on the Compatibility of Pesticides with Microbial Inoculants" Agronomy 11, no. 5: 870. https://doi.org/10.3390/agronomy11050870
APA StyleSantos, M. S., Rodrigues, T. F., Nogueira, M. A., & Hungria, M. (2021). The Challenge of Combining High Yields with Environmentally Friendly Bioproducts: A Review on the Compatibility of Pesticides with Microbial Inoculants. Agronomy, 11(5), 870. https://doi.org/10.3390/agronomy11050870