
Pea Breeding Lines Adapted to Autumn Sowings in Broomrape Prone Mediterranean Environments

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Assessments
2.3. Statistical Analysis
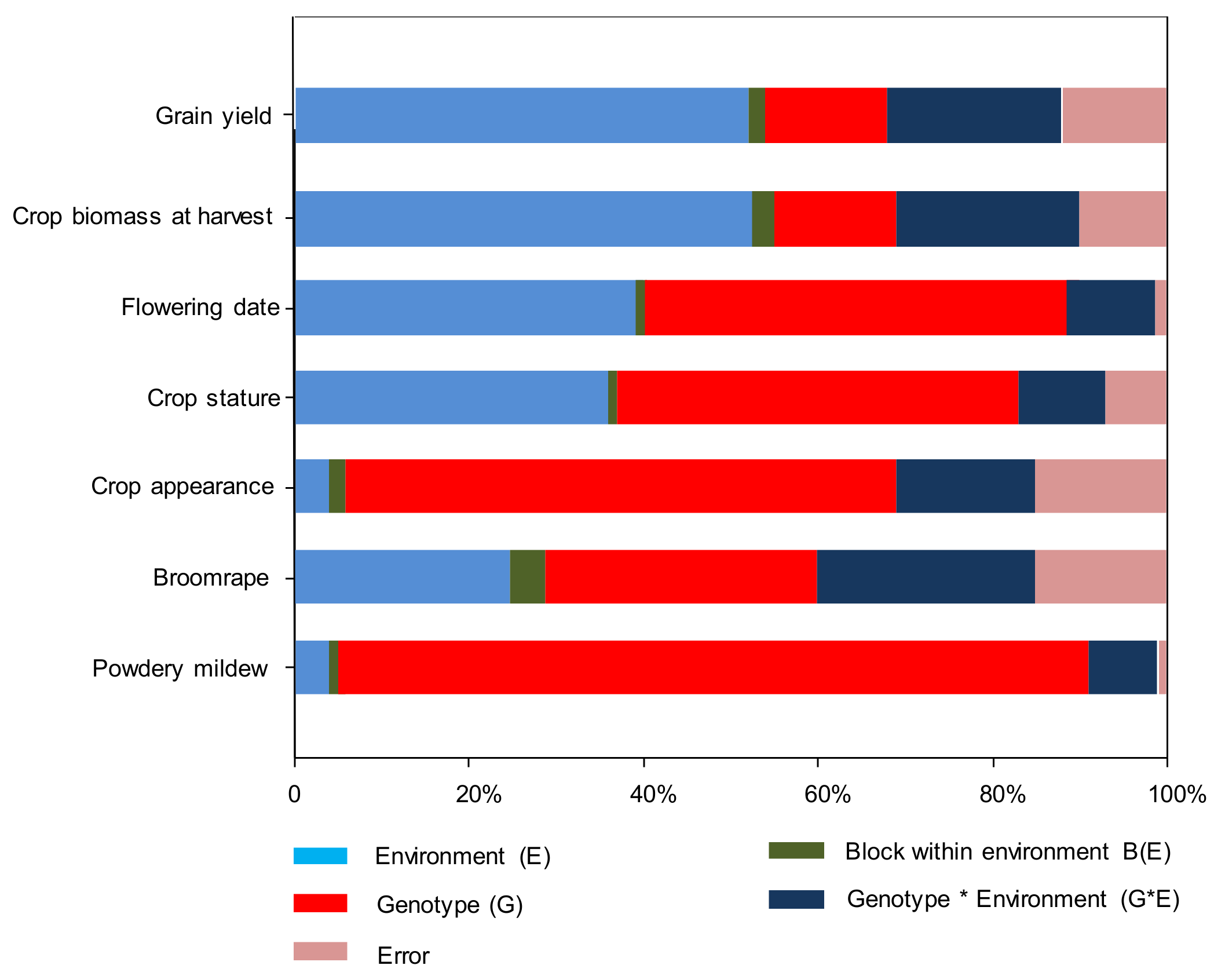
2.3.1. Variances Analyses
2.3.2. Heritability-Adjusted GGE Biplot (HA-GGE)
2.3.3. Multi-Trait Stability Index (MTSI) Based on Factor Analysis
2.3.4. Non-Metric Multi-Dimensional Scaling Ordination (NMDS)
3. Results
3.1. Multi-Trait Stability Index (MTSI)
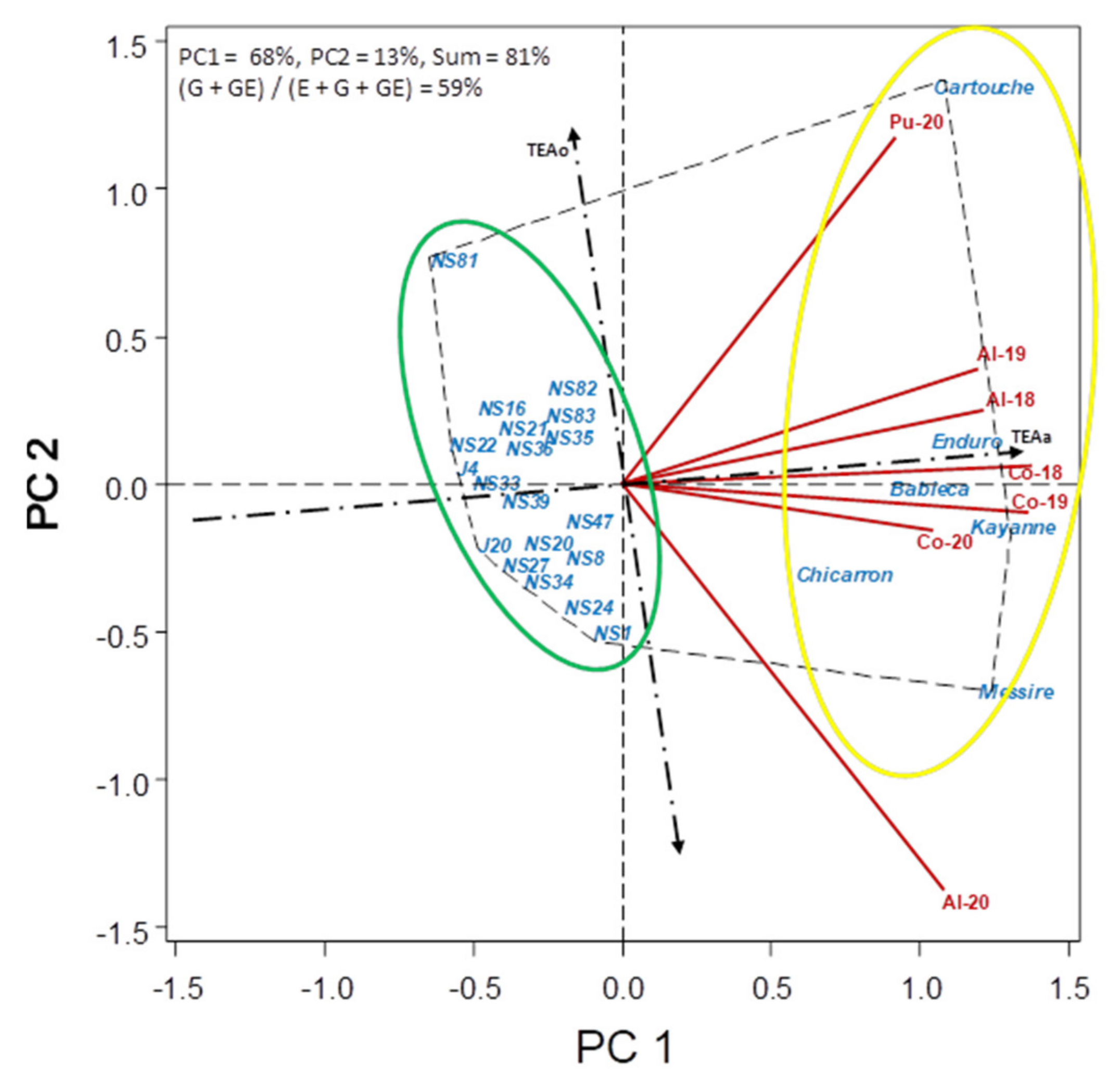
3.2. Correlations between Traits and Non-Metric Multi-Dimensional Scaling Ordination (NMDS)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Stagnari, F.; Maggio, A.; Galieni, A.; Pisante, M. Multiple benefits of legumes for agriculture sustainability: An overview. Chem. Biol. Technol. Agric. 2017, 4, 2. [Google Scholar] [CrossRef]
- Guinet, M.; Nicolardot, B.; Voisin, A.S. Nitrogen benefits of ten legume pre-crops for wheat assessed by field measurements and modelling. Eur. J. Agron. 2020, 120, 126–151. [Google Scholar] [CrossRef]
- FAOSTAT 2020. Available online: http://www.fao.org/faostat (accessed on 4 February 2021).
- Tulbek, M.C.; Lam, R.S.H.; Wang, Y.C.; Asavajaru, P.; Lam, A. Chapter 9—Pea: A Sustainable Vegetable Protein Crop. In Sustainable protein sources; Nadathur, S.R., Wanasundara, J.P.D., Scanlin, L., Eds.; Academis Press: Cambridge, MA, USA, 2017; pp. 145–164. [Google Scholar] [CrossRef]
- Parihar, A.K.; Dixit, G.P.; Bohra, A.; Gupta, D.S.; Singh, A.K.; Kumar, N.; Singh, D.; Singh, N.P. Genetic Advancement in Dry Pea (Pisum sativum L.): Retrospect and Prospect. In Accelerated Plant Breeding; Gosal, S.S., Wani, S.H., Eds.; Springer Nature: Cham, Switzerland, 2020; Volume 3. [Google Scholar] [CrossRef]
- Rubiales, D.; Fondevilla, S.; Chen, W.; Gentzbittel, L.; Higgins, T.J.V.; Castillejo, M.A.; Singh, K.B.; Rispail, N. Achievements and challenges in legume breeding for pest and disease resistance. Crit. Rev. Plant Sci. 2015, 34, 195–236. [Google Scholar] [CrossRef]
- Neugschwandtner, R.W.; Bernhuber, A.; Kammlander, S.; Wagentristl, H.; Klimek-Kopyra, A.; Kaul, H.P. Yield structure components of autumn- and spring-sown pea (Pisum sativum L.). Acta Agric. Scand. Sect. B—Soil Plant Sci 2020, 70, 109–116. [Google Scholar] [CrossRef]
- Rubiales, D.; Fernández-Aparicio, M.; Moral, A.; Barilli, E.; Sillero, J.C.; Fondevilla, S. Disease resistance in pea (Pisum sativum L.) types for autumn sowings in Mediterranean environments. Czech. J. Genet. Plant Breed. 2009, 45, 135–142. [Google Scholar] [CrossRef]
- Rubiales, D.; Fernández-Aparicio, M.; Pérez-De-Luque, A.; Castillejo, M.A.; Prats, E.; Sillero, J.C.; Rispail, N.; Fondevilla, S. Breeding approaches for crenate broomrape (Orobanche crenata Forsk.) management in pea (Pisum sativum L.). Pest. Manag. Sci. 2009, 65, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Rubiales, D.; Fernández-Aparicio, M. Innovations in parasitic weeds management in legume crops. A review. Agron. Sustain. Dev. 2011, 32, 433–449. [Google Scholar] [CrossRef]
- Rubiales, D.; Moreno, M.T.; Sillero, J.C. Search for Resistance to Crenate Broomrape (Orobanche crenata Forsk.) in Pea Germplasm. Genet. Resour. Crop Evol. 2005, 52, 853–861. [Google Scholar] [CrossRef]
- Fondevilla, S.; Flores, F.; Emeran, A.A.; Kharrat, M.; Rubiales, D. High productivity of dry pea genotypes resistant to crenate broomrape in Mediterranean environments. Agron. Sustain. Dev. 2017, 37, 61. [Google Scholar] [CrossRef]
- Rubiales, D.; Fondevilla, S.; Fernández-Aparicio, M. Development of Pea Breeding Lines with Resistance to Orobanche crenata Derived from Pea Landraces and Wild Pisum spp. Agronomy 2021, 11, 36. [Google Scholar] [CrossRef]
- Fondevilla, S.; Fernández-Aparicio, M.; Satovic, Z.; Emeran, A.A.; Torres, A.M.; Moreno, M.T.; Rubiales, D. Identification of quantitative trait loci for specific mechanisms of resistance to Orobanche crenata in pea. Mol. Breed. 2010, 25, 259–272. [Google Scholar] [CrossRef]
- Rubiales, D.; Fondevilla, S. Future prospects for ascochyta blight resistance breeding in cool season food legumes. Front. Plant Sci. 2012, 3, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.; Timmerman-Vaughan, G.; Rubiales, D.; Warkentin, T.; Siddique, K.; Erskine, W.; Barbetti, M. Didymella pinodes and its management in field pea: Challenges and opportunities. Field Crop Res. 2013, 148, 61–77. [Google Scholar] [CrossRef]
- Iglesias-García, R.; Prats, E.; Fondevilla, S.; Satovic, Z.; Rubiales, D. Quantitative Trait Loci Associated to Drought Adaptation in Pea (Pisum sativum L.). Plant Mol. Biol. Rep. 2015, 33, 1768–1778. [Google Scholar] [CrossRef]
- Tiwari, K.R.; Penner, G.A.; Warkentin, T.D. Inheritance of powdery mildew resistance in pea. Can. J. Plant Sci. 1997, 77, 307–310. [Google Scholar] [CrossRef]
- Fondevilla, S.; Rubiales, D. Powdery mildew control in pea. A review. Agron. Sustain. Dev. 2011, 32, 401–409. [Google Scholar] [CrossRef]
- Yan, W.; Holland, J.B. A heritability-adjusted GGE biplot for test environment evaluation. Euphytica 2009, 171, 355–369. [Google Scholar] [CrossRef]
- Flores, F.; Nadal, S.; Solis, I.; Winkler, J.; Sass, O.; Stoddard, F.L.; Link, W.; Raffiot, B.; Muel, F.; Rubiales, D.; et al. Faba bean adaptation to autumn sowing under European climates. Agron. Sustain. Dev. 2012, 32, 727–734. [Google Scholar] [CrossRef]
- Iglesias-García, R.; Prats, E.; Flores, F.; Amri, M.; Mikić, A.; Rubiales, D. Assessment of field pea (Pisum sativum L.) grain yield, aerial biomass and flowering date stability in Mediterranean environments. Crop Pasture Sci. 2017, 68, 915–923. [Google Scholar] [CrossRef]
- Rubiales, D.; Emeran, A.A.; Flores, F. Adaptation of Grass Pea (Lathyrus sativus) to Mediterranean Environments. Agronomy 2020, 10, 1295. [Google Scholar] [CrossRef]
- RAEA. Resultados de Ensayos de Variedades de Guisantes Proteaginosos en Andalucía. Campaña 2014/2015. 2015. Available online: File:///E:/Descargas/RAEA%20Resultados%20guisantes%2014-15%20 (accessed on 21 December 2020).
- Red de Información Agroclimática de Andalucía (RIA). Available online: https://www.juntadeandalucia.es/agriculturaypesca/ifapa/riaweb/web/ (accessed on 21 December 2020).
- McIntosh, M.S. Analysis of combined experiments. Agron. J. 1983, 75, 153–155. [Google Scholar] [CrossRef]
- Aznar-Fernández, T.; Carrillo-Perdomo, E.; Flores, F.; Rubiales, D. Identification and multi-environment validation of resistance to pea weevil (Bruchus pisorum) in Pisum germplasm. J. Pest Sci. 2018, 91, 505–514. [Google Scholar] [CrossRef]
- Burgueño, J.; Crossa, J.; Vargas, M. SAS Programs for Graphing GE and GGE Biplots; CIMMYT: El Batan, Mexico, 2003. [Google Scholar]
- Olivoto, T.; Lúcio, A.D.C.; Da Silva, J.A.G.; Sari, B.G.; Diel, M.I. Mean Performance and Stability in Multi-Environment Trials II: Selection Based on Multiple Traits. Agron. J. 2019, 111, 2961–2969. [Google Scholar] [CrossRef]
- Olivoto, T.; Lúcio, A.D. metan: An R package for multi-environment trial analysis. Methods Ecol. Evol. 2020, 11, 783–789. [Google Scholar] [CrossRef]
- Kruskal, J.B. Multidimensional scaling by optimizing goodness of fit to a nonmetric hypothesis. Psychometrika 1964, 29, 1–27. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Yang, R.-C.; Crossa, J.; Cornelius, P.L.; Burgueño, J. Biplot Analysis of Genotype × Environment Interaction: Proceed with Caution. Crop Sci. 2009, 49, 1564–1576. [Google Scholar] [CrossRef]
- Smykal, P.; Aubert, G.; Burstin, J.; Coyne, C.J.; Ellis, N.T.H.; Flavell, A.J.; Ford, R.; Hýbl, M.; Macas, J.; Neumann, P.; et al. Pea (Pisum sativum L.) in the Genomic Era. Agronomy 2012, 2, 74–115. [Google Scholar] [CrossRef]
- Rubiales, D.; González-Bernal, M.J.; Warkentin, T.; Bueckert, R.; VazPatto, M.C.; McPhee, K.; McGee, R.; Smýkal, P. CH20—Advances in breeding of peas. In Achieving Sustainable Cultivation of Vegetables; Hochmuth, G., Ed.; Burleig Dodds Science Publishing Limited: Cambridge, UK, 2019. [Google Scholar]
- Fernández-Aparicio, M.; Flores, F.; Rubiales, D. The Effect of Orobanche crenata Infection Severity in Faba Bean, Field Pea, and Grass Pea Productivity. Front. Plant Sci. 2016, 7, 1409. [Google Scholar] [CrossRef]
- Parker, C. Parasitic Weeds: A World Challenge. Weed Sci. 2012, 60, 269–276. [Google Scholar] [CrossRef]
- Parker, C. Orobanche crenata in UK—An update. Haustorium 2014, 65, 5–6. [Google Scholar]
- Rubiales, D. Can we breed for durable resistance to broomrapes? Phytopathol. Mediterr. 2018, 57, 170–185. [Google Scholar]
- Grenz, J.; Sauerborn, J. Mechanisms limiting the geographical range of the parasitic weed Orobanche crenata. Agric. Ecosyst. Environ. 2007, 122, 275–281. [Google Scholar] [CrossRef]
- Pérez-De-Luque, A.; Jorrín, J.; Cubero, J.I.; Rubiales, D. Orobanche crenata resistance and avoidance in pea (Pisum spp.) operate at different developmental stages of the parasite. Weed Res. 2005, 45, 379–387. [Google Scholar] [CrossRef]
- Pérez-De-Luque, A.; Sillero, J.C.; Moral, A.; Cubero, J.I.; Rubiales, D. Effect of sowing date and host resistance on the establishment and development of Orobanche crenata in faba bean and common vetch. Weed Res. 2004, 44, 282–288. [Google Scholar] [CrossRef]
- Rubiales, D.; Alcántara, C.; Pérez-De-Luque, A.; Gil, J.; Sillero, J.C. Infection of chickpea (Cicer arietinum) by crenate broomrape (Orobanche crenata) as influenced by sowing date and weather conditions. Agrononie 2003, 23, 359–362. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Flores, F.; Rubiales, D. Escape and true resistance to crenate broomrape (Orobanche crenata Forsk.) in grass pea (Lathyrus sativus L.) germplasm. Field Crops Res. 2012, 125, 92–97. [Google Scholar] [CrossRef]
- Moral, J.; Lozano-Baena, M.D.; Rubiales, D. Temperature and water stress during conditioning and incubation phase affecting Orobanche crenata seed germination and radicle growth. Front. Plant Sci. 2015, 6, 408. [Google Scholar] [CrossRef]
- Pérez-De-Luque, A.; Flores, F.; Rubiales, D. Differences in Crenate Broomrape Parasitism Dynamics on Three Legume Crops Using a Thermal Time Model. Front. Plant Sci. 2016, 7, 1910. [Google Scholar] [CrossRef] [PubMed]
- Rubiales, D.; Barilli, E.; Flores, F. Broomrape (Orobanche crenata) as a major constraint for grass pea (Lathyrus sativus) production in Mediterranean rain-fed environments. Agronomy 2020, 10, 1931. [Google Scholar] [CrossRef]
- Yu, J.; Holland, J.B.; McMullen, M.D.; Buckler, E.S. Genetic Design and Statistical Power of Nested Association Mapping in Maize. Genetics 2008, 178, 539–551. [Google Scholar] [CrossRef]
- Gutiérrez, N.; Palomino, C.; Šatović, Z.; Ruiz-Rodríguez, M.D.; Vitale, S.; Gutiérrez, M.V.; Rubiales, D.; Kharrat, M.; Amri, M.; Emeran, A.A.; et al. QTLs for Orobanche spp. resistance in faba bean: Identification and validation across different environments. Mol. Breed. 2013, 32, 909–922. [Google Scholar] [CrossRef]
- Maalouf, F.; Khalil, S.; Ahmed, S.; Akintunde, A.N.; Kharrat, M.; El Shama’A, K.; Hajjar, S.; Malhotra, R.S. Yield stability of faba bean lines under diverse broomrape prone production environments. Field Crops Res. 2011, 124, 288–294. [Google Scholar] [CrossRef]
- Rubiales, D.; Flores, F.; Emeran, A.A.; Kharrat, M.; Amri, M.; Rojas-Molina, M.M.; Sillero, J.C. Identification and multi-environment validation of resistance against broomrapes (Orobanche crenata and O. foetida) in faba bean (Vicia faba). Field Crops Res. 2014, 166, 58–65. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Moral, A.; Kharrat, M.; Rubiales, D. Resistance against broomrapes (Orobanche and Phelipanche spp.) in faba bean (Vicia faba) based in low induction of broomrape seed germination. Euphytica 2012, 186, 897–905. [Google Scholar] [CrossRef]
- Ejeta, G. Breeding for Striga Resistance in Sorghum: Exploitation of an Intricate Host-Parasite Biology. Crop Sci. 2007, 47, S216–S227. [Google Scholar] [CrossRef]
- Vogler, R.K.; Ejeta, G.; Butler, L.G. Inheritance of Low Production of Striga Germination Stimulant in Sorghum. Crop Sci. 1996, 36, 1185–1191. [Google Scholar] [CrossRef]
- Pavan, S.; Schiavulli, A.; Marcotrigiano, A.R.; Bardaro, N.; Bracuto, V.; Ricciardi, F.; Charnikhova, T.; Lotti, C.; Bouwmeester, H.; Ricciardi, L. Characterization of Low-Strigolactone Germplasm in Pea (Pisum sativum L.) Resistant to Crenate Broomrape (Orobanche crenata Forsk.). Mol. Plant-Microbe Interact. 2016, 29, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Bardaro, N.; Marcotrigiano, A.R.; Bracuto, V.; Mazzeo, R.; Ricciardi, F.; Lotti, C.; Pavan, S.; Ricciardi, L. Genetic analysis of resistance to Orobanche crenata (Forsk.) in a pea (Pisum sativum L.) low-strigolactone line. J. Plant Pathol. 2016, 98, 671–675. [Google Scholar]
- Fondevilla, S.; Torres, A.M.; Moreno, M.T.; Rubiales, D. Identification of a New Gene for Resistance to Powdery Mildew in Pisum fulvum, a Wild Relative of Pea. Breed. Sci. 2007, 57, 181–184. [Google Scholar] [CrossRef]
- Iglesias-García, R.; Rubiales, D.; Fondevilla, S. Penetration resistance to Erysiphe pisi in pea mediated by er1 gene is associated with protein cross-linking but not with callose apposition or hypersensitive response. Euphytica 2015, 201, 381–387. [Google Scholar] [CrossRef]
- Carrillo, E.; Satovic, Z.; Aubert, G.; Boucherot, K.; Rubiales, D.; Fondevilla, S. Identification of quantitative trait loci and candidate genes for specific cellular resistance responses against Didymella pinodes in pea. Plant Cell Rep. 2014, 33, 1133–1145. [Google Scholar] [CrossRef]
- Banniza, S.; Hashemi, P.; Warkentin, T.D.; Vandenberg, A.; Davis, A.R. The relationships among lodging, stem anatomy, degree of lignification, and resistance to mycosphaerella blight in field pea (Pisum sativum). Can. J. Bot. 2005, 83, 954–967. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Russi, L.; Romani, M.; Pecetti, L.; Nazzicari, N. Farmer-participatory vs. conventional market-oriented breeding of inbred crops using phenotypic and genome-enabled approaches: A pea case study. Field Crops Res. 2019, 232, 30–39. [Google Scholar] [CrossRef]
- Tayeh, N.; Klein, A.; Le Paslier, M.C.; Jacquin, F.; Houtin, H.; Rond, C.; Chabert-Martinello, M.; Magnin-Robert, J.B.; Marget, P.; Aubert, G.; et al. Genomic prediction in pea: Effect of marker density and training population size and composition on prediction accuracy. Front. Plant Sci. 2015, 6, 941. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Rubiales, D.; Wang, Y.; Fang, P.; Sun, T.; Liu, N.; Xu, P. Omics resources and omics-enabled approaches for achieving high productivity and improved quality in pea (Pisum sativum L.). Theor. Appl. Genet. 2021, 134, 755–776. [Google Scholar] [CrossRef]
- Cvejić, S.; Radanović, A.; Dedić, B.; Jocković, M.; Jocić, S.; Miladinović, D. Genetic and Genomic Tools in Sunflower Breeding for Broomrape Resistance. Genes 2020, 11, 152. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Derived from Cross * or Obtentor | Progeny | |
|---|---|---|---|
| Breeding lines | J4 | Ps624 × Messire | Ps624/Messire |
| J20 | [(Messire × Pf660) × Messire] × Ballet | Messire/Pf660//Ballet | |
| NS1 | J20 × J23 | Messire/Pf660/Ballet//Ps624/Ps423/Radley | |
| NS8 | [(3070 × J4) × (3054 × J20)] × Viriato | Franklin/Ps624/Messire//B99-100/Messire/Pf660/Ballet//Viriato | |
| NS16 | J2 × Cartouche | Ps624/Messire//Cartouche | |
| NS20 | 3062 × J20 | 3062//Messire/Pf660/Ballet | |
| NS21 | 3066 × J4 | 3066//Ps624/Messire | |
| NS22 | 3066 × J4 | 3066//Ps624/Messire | |
| NS24 | Baccara × J6 | Baccara//Ps565/Ps624 | |
| NS27 | 3066 × J4 | 3066//Ps624/Messire | |
| NS33 | J4 × J23 | Ps624/Messire//Ps423/Radley | |
| NS34 | 3064 × J20 | 3064//Messire/Pf660/Ballet | |
| NS35 | 3066 × J4 | 3066//Ps624/Messire | |
| NS36 | 3066 × J4 | 3066//Ps624/Messire | |
| NS39 | (J4 × Baccara) × Chicarrón | Ps624/Messire/Baccara//Chicarron | |
| NS47 | [3070 × (6NIL × Baccara)] × (J22 × RMS1) | Franklin//6NIL/Baccara/Messire/Pf660/P665/RMS1 | |
| NS81 | 3071 × (6NIL × Baccara) | Lifter//Messire/Pf660/Baccara | |
| NS82 | 3066 × J4 | 3066//Ps624/Messire | |
| NS83 | 3066 × J4 | 3066//Ps624/Messire/ | |
| Check cultivars | Messire | SERASEM, France | |
| Kayanne | MOMONT, France | ||
| Chicarrón | ITACyL, Spain | ||
| Cartouche | SERASEM, France | ||
| Enduro | FLORIM. D., France | ||
| Babieca | INIA, Spain |
| Environments | Season | Site, Level of Broomrape (Oc) Infestation | Soil Type | Soil pH | Organic Matter (g/100 g) | Available Phosphorus (mg/kg) | Average Tmax (°C) | Average Tmin (°C) | Rain (mm) |
| Al-18 | 2017–2018 | Córdoba Almezos: high Oc | Cambisol | - | - | - | 19.8 | 7.6 | 442 |
| Al-19 | 2018–2019 | Córdoba Almezos: high Oc | Cambisol | 7.5 | 1.2 | 15.0 | 21.6 | 6.4 | 206 |
| Al-20 | 2019–2020 | Córdoba Almezos: high Oc | Cambisol | - | - | - | 19.9 | 6.7 | 362 |
| Co-18 | 2017–2018 | Córdoba Cortijo: high Oc | Cambisol | - | - | - | 19.5 | 7.5 | 441 |
| Co-19 | 2018–2019 | Córdoba Cortijo: high Oc | Cambisol | 7.5 | 1.2 | 15.1 | 22.2 | 6.3 | 206 |
| Co-20 | 2019–2020 | Córdoba Cortijo: high Oc | Cambisol | - | - | - | 20.1 | 6.6 | 363 |
| Pu-18 | 2017–2018 | Córdoba Puente: low Oc | Vertisol | - | - | - | 19.6 | 7.6 | 441 |
| Pu-19 | 2018–2019 | Córdoba Puente: low Oc | Vertisol | 7.8 | 0.7 | 9.9 | 22.0 | 5.9 | 129 |
| Pu-20 | 2019–2020 | Córdoba Puente: low Oc | Vertisol | - | - | - | 20.0 | 6.7 | 322 |
| Accession | Al-18 | Al-19 | Al-20 | Co-18 | Co-19 | Co-20 | Pu-18 | Pu-19 | Pu-20 | Mean | SE | S2xi a | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Breeding lines | NS22 | 4811 | 3693 | 2292 | 4144 | 3077 | 1494 | 7333 | 7933 | 6560 | 4593 | 463 | 5,098,509 |
| NS34 | 4552 | 4685 | 3136 | 3656 | 3360 | 2842 | 6430 | 5593 | 5753 | 4445 | 263 | 1,640,110 | |
| NS8 | 4387 | 4463 | 1564 | 2193 | 3679 | 2112 | 7610 | 6007 | 4790 | 4089 | 404 | 3,855,415 | |
| NS39 | 4273 | 4627 | 1516 | 2823 | 2542 | 2192 | 6600 | 6239 | 5870 | 4076 | 384 | 3,579,527 | |
| NS35 | 4123 | 2917 | 2372 | 3124 | 3083 | 1904 | 6267 | 5865 | 5581 | 3915 | 315 | 2,611,513 | |
| NS21 | 4282 | 3611 | 2618 | 3554 | 3572 | 2027 | 3904 | 4669 | 5786 | 3780 | 265 | 1,203,486 | |
| NS83 | 3498 | 3798 | 2489 | 3041 | 2824 | 1963 | 4094 | 5859 | 5678 | 3694 | 261 | 1,808,051 | |
| NS33 | 3290 | 2763 | 2616 | 3330 | 2531 | 1262 | 5667 | 4392 | 4923 | 3419 | 293 | 1,850,394 | |
| NS27 | 4643 | 3193 | 1876 | 2522 | 3004 | 1304 | 4596 | 3530 | 5693 | 3374 | 316 | 1,991,276 | |
| NS81 | 3145 | 3874 | 1776 | 2457 | 2925 | 1320 | 2296 | 5757 | 5933 | 3276 | 348 | 2,677,509 | |
| NS36 | 3498 | 3935 | 1721 | 2441 | 2665 | 1579 | 4715 | 2788 | 5038 | 3153 | 258 | 1,516,254 | |
| NS82 | 2900 | 3256 | 1705 | 2584 | 3097 | 1571 | 2584 | 5633 | 5025 | 3151 | 265 | 1,872,430 | |
| NS47 | 3578 | 3750 | 1644 | 2137 | 2337 | 1814 | 4912 | 4363 | 3060 | 3066 | 266 | 1,350,504 | |
| NS1 | 3437 | 3911 | 1796 | 1996 | 2174 | 1812 | 4952 | 3644 | 3493 | 3024 | 217 | 1,254,387 | |
| NS20 | 3434 | 3712 | 1788 | 1993 | 2362 | 2134 | 4944 | 1887 | 4007 | 2918 | 236 | 1,288,321 | |
| NS24 | 3218 | 3057 | 472 | 1552 | 3149 | 854 | 4378 | 3468 | 5297 | 2827 | 314 | 2,536,403 | |
| J20 | 3484 | 3169 | 2140 | 2009 | 2530 | 2120 | 3904 | 2482 | 3473 | 2812 | 169 | 496,512 | |
| J4 | 3271 | 3499 | 2636 | 2074 | 2380 | 1498 | 2459 | 3015 | 3827 | 2740 | 186 | 540,092 | |
| NS16 | 2444 | 2777 | 604 | 1003 | 1222 | 762 | 2540 | 1972 | 2957 | 1809 | 183 | 845,190 | |
| Check cultivars | Messire | 4905 | 2987 | 180 | 1267 | 2117 | 832 | 7933 | 6126 | 5800 | 3572 | 521 | 7,383,456 |
| Kayanne | 3690 | 3383 | 128 | 2249 | 2759 | 794 | 5185 | 7208 | 5230 | 3403 | 427 | 5,044,151 | |
| Chicarrón | 3549 | 3643 | 68 | 2494 | 2837 | 904 | 7180 | 5191 | 3920 | 3310 | 440 | 4,530,520 | |
| Cartouche | 3537 | 2508 | 56 | 1276 | 722 | 124 | 5514 | 4630 | 2870 | 2360 | 382 | 3,866,770 | |
| Enduro | 2634 | 2789 | 4 | 975 | 973 | 368 | 4930 | 4873 | 3270 | 2313 | 355 | 3,418,513 | |
| Babieca | 2607 | 1752 | 8 | 1166 | 1690 | 348 | 2554 | 5026 | 3580 | 2081 | 300 | 2,477,425 | |
| Mean | 3647 | 3430 | 1488 | 2322 | 2544 | 1437 | 4939 | 4726 | 4696 | 3248 | |||
| SE | 95 | 117 | 147 | 122 | 94 | 84 | 215 | 217 | 142 | 69 |
| Accession | Al-18 | Al-19 | Al-20 | Co-18 | Co-19 | Co-20 | Pu-18 | Pu-19 | Pu-20 | Mean | SE | S2xi a | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Breeding lines | NS22 | 0.00 | 0.01 | 0.06 | 0.14 | 0.08 | 0.18 | 0.00 | 0.00 | 0.16 | 0.06 | 0.02 | 0.0056 |
| J4 | 0.00 | 0.00 | 0.57 | 0.36 | 0.03 | 0.37 | 0.00 | 0.00 | 0.00 | 0.15 | 0.05 | 0.0495 | |
| NS16 | 0.05 | 0.03 | 0.78 | 0.47 | 0.20 | 0.21 | 0.00 | 0.00 | 0.08 | 0.20 | 0.05 | 0.0694 | |
| J20 | 0.10 | 0.01 | 1.15 | 0.12 | 0.03 | 0.41 | 0.00 | 0.00 | 0.01 | 0.20 | 0.07 | 0.1430 | |
| NS33 | 0.05 | 0.35 | 0.90 | 0.18 | 0.18 | 0.20 | 0.00 | 0.00 | 0.02 | 0.21 | 0.06 | 0.0809 | |
| NS27 | 0.00 | 0.05 | 1.38 | 0.21 | 0.20 | 0.24 | 0.00 | 0.00 | 0.04 | 0.23 | 0.08 | 0.1939 | |
| NS81 | 0.03 | 0.06 | 1.29 | 0.50 | 0.04 | 0.25 | 0.00 | 0.00 | 0.06 | 0.25 | 0.08 | 0.1792 | |
| NS39 | 0.14 | 0.05 | 1.04 | 0.14 | 0.07 | 1.17 | 0.00 | 0.00 | 0.06 | 0.30 | 0.09 | 0.2136 | |
| NS20 | 0.16 | 0.04 | 1.14 | 0.58 | 0.21 | 0.71 | 0.00 | 0.00 | 0.02 | 0.32 | 0.07 | 0.1629 | |
| NS36 | 0.04 | 0.01 | 0.86 | 0.16 | 0.15 | 1.56 | 0.00 | 0.00 | 0.14 | 0.32 | 0.11 | 0.2869 | |
| NS21 | 0.06 | 0.10 | 0.87 | 0.05 | 0.12 | 1.57 | 0.00 | 0.00 | 0.16 | 0.33 | 0.11 | 0.2905 | |
| NS8 | 0.15 | 0.07 | 1.37 | 0.44 | 0.28 | 0.85 | 0.00 | 0.00 | 0.10 | 0.36 | 0.09 | 0.2160 | |
| NS24 | 0.28 | 0.04 | 1.78 | 0.43 | 0.19 | 0.52 | 0.00 | 0.00 | 0.08 | 0.37 | 0.11 | 0.3152 | |
| NS47 | 0.24 | 0.27 | 1.20 | 0.66 | 0.19 | 0.82 | 0.00 | 0.00 | 0.06 | 0.38 | 0.08 | 0.1754 | |
| NS34 | 0.03 | 0.16 | 1.25 | 0.07 | 0.65 | 1.36 | 0.00 | 0.00 | 0.02 | 0.39 | 0.13 | 0.3093 | |
| NS35 | 0.18 | 0.26 | 0.92 | 0.13 | 0.13 | 1.71 | 0.00 | 0.00 | 0.15 | 0.39 | 0.12 | 0.3227 | |
| NS82 | 0.03 | 0.16 | 0.87 | 0.30 | 0.03 | 1.94 | 0.00 | 0.00 | 0.25 | 0.40 | 0.14 | 0.4096 | |
| NS1 | 0.28 | 0.02 | 1.77 | 0.70 | 0.12 | 0.74 | 0.00 | 0.00 | 0.04 | 0.41 | 0.11 | 0.3461 | |
| NS83 | 0.02 | 0.13 | 0.86 | 0.29 | 0.02 | 2.24 | 0.00 | 0.00 | 0.21 | 0.42 | 0.15 | 0.5405 | |
| Check cultivars | Cartouche | 0.84 | 1.34 | 0.90 | 1.61 | 1.94 | 0.90 | 0.00 | 0.00 | 0.65 | 0.91 | 0.15 | 0.4331 |
| Chicarrón | 0.21 | 0.88 | 1.81 | 0.60 | 2.56 | 2.42 | 0.00 | 0.00 | 0.19 | 0.96 | 0.21 | 1.0686 | |
| Babieca | 0.80 | 0.84 | 1.61 | 1.22 | 2.38 | 2.11 | 0.00 | 0.00 | 0.25 | 1.02 | 0.17 | 0.7725 | |
| Enduro | 0.47 | 1.56 | 1.88 | 2.06 | 2.80 | 1.47 | 0.00 | 0.00 | 0.28 | 1.17 | 0.22 | 1.0269 | |
| Kayanne | 1.07 | 1.83 | 1.95 | 1.49 | 2.08 | 2.12 | 0.00 | 0.00 | 0.18 | 1.19 | 0.19 | 0.8251 | |
| Messire | 0.84 | 0.57 | 2.72 | 1.57 | 2.33 | 2.41 | 0.00 | 0.00 | 0.26 | 1.19 | 0.21 | 1.1847 | |
| Mean | 0.24 | 0.35 | 1.24 | 0.57 | 0.68 | 1.14 | 0.00 | 0.00 | 0.14 | 0.48 | |||
| SE | 0.04 | 0.07 | 0.07 | 0.06 | 0.11 | 0.11 | 0.00 | 0.00 | 0.02 | 0.03 |
| Trait | Factor | XO | XS | SD | h2 | SG(%) |
|---|---|---|---|---|---|---|
| Grain yield | FA1 | 3054 | 3709 | 656 | 0.76 | 16.43 |
| Dry biomass | FA1 | 6624 | 8372 | 1748 | 0.78 | 20.74 |
| Crop appearance | FA1 | 3.33 | 4.27 | 0.93 | 0.95 | 26.93 |
| Broomrape infection | FA2 | 0.59 | 0.41 | −0.18 | 0.77 | 23.13 |
| Grain Yield | Dry Biomass | Dtf | Crop Stature | Crop Appearance # | Broomrape | Powdery Mildew | Ascochyta Blight ## | |
|---|---|---|---|---|---|---|---|---|
| Grain yield | 0.9649 *** | −0.0972 | 0.2056 | 0.8955 *** | −0.9650 *** | −0.1269 | −0.1772 | |
| Dry biomass | 0.0426 | 0.1602 | 0.8052 *** | −0.9115 *** | −0.1304 | −0.6560 ** | ||
| Dtf | 0.5760 * | −0.1314 | −0.0714 | 0.6284 ** | −0.8432 *** | |||
| Crop stature | 0.3971 | −0.2999 | 0.3615 | −0.7356 *** | ||||
| Crop appearance # | −0.8858 *** | −0.0482 | −0.7937 *** | |||||
| Broomrape | −0.0604 | 0.1012 | ||||||
| Powdery mildew | −0.5859 * | |||||||
| Ascochyta blight ## |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubiales, D.; Osuna-Caballero, S.; González-Bernal, M.J.; Cobos, M.J.; Flores, F. Pea Breeding Lines Adapted to Autumn Sowings in Broomrape Prone Mediterranean Environments. Agronomy 2021, 11, 769. https://doi.org/10.3390/agronomy11040769
Rubiales D, Osuna-Caballero S, González-Bernal MJ, Cobos MJ, Flores F. Pea Breeding Lines Adapted to Autumn Sowings in Broomrape Prone Mediterranean Environments. Agronomy. 2021; 11(4):769. https://doi.org/10.3390/agronomy11040769
Chicago/Turabian StyleRubiales, Diego, Salvador Osuna-Caballero, María J. González-Bernal, María J. Cobos, and Fernando Flores. 2021. "Pea Breeding Lines Adapted to Autumn Sowings in Broomrape Prone Mediterranean Environments" Agronomy 11, no. 4: 769. https://doi.org/10.3390/agronomy11040769
APA StyleRubiales, D., Osuna-Caballero, S., González-Bernal, M. J., Cobos, M. J., & Flores, F. (2021). Pea Breeding Lines Adapted to Autumn Sowings in Broomrape Prone Mediterranean Environments. Agronomy, 11(4), 769. https://doi.org/10.3390/agronomy11040769