
Rotation and Fertilization Effects on Soil Quality and Yields in a Long Term Field Experiment


 ,
,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
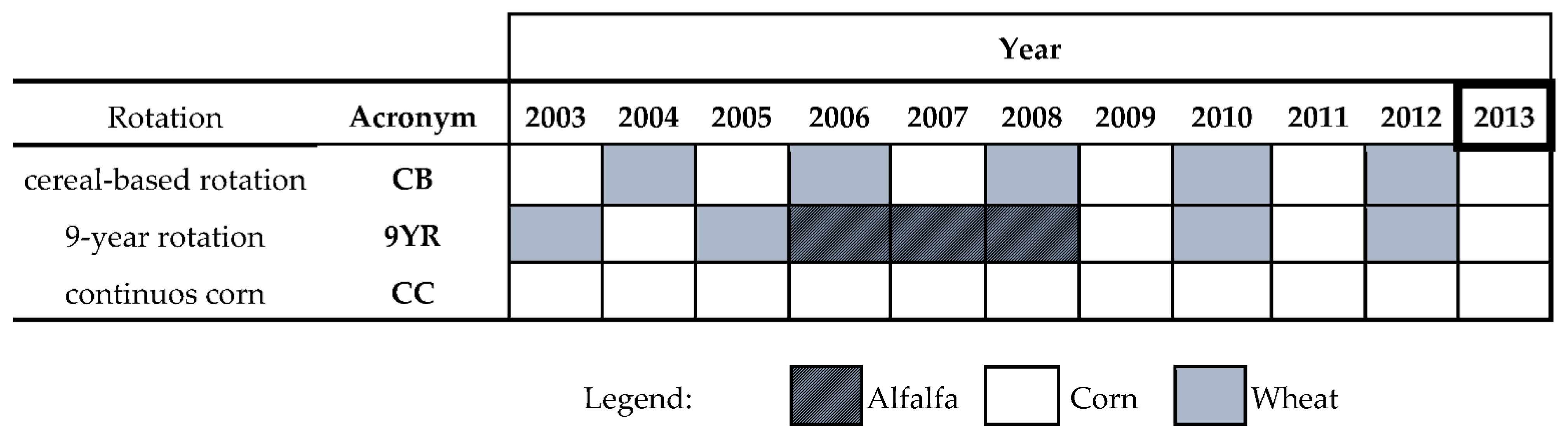
2.1. Study Site and Experimental Design
- CC: continuous corn (Zea mays L.);
- CB: 2 year cereal-based rotation of corn (Zea mays L.) and wheat (Triticum aestivum L.);
- 9YR: 9 year rotations consisting of corn-wheat-corn-wheat-corn-wheat-alfalfa-alfalfa-alfalfa (Medicago sativa L.).
- CK: unfertilized control;
- NP: mineral N and phosphorous (P) fertilization;
- M: cattle manure fertilization.
2.2. Soil Sampling and Analysis
2.3. Data Handling and Statistics
3. Results
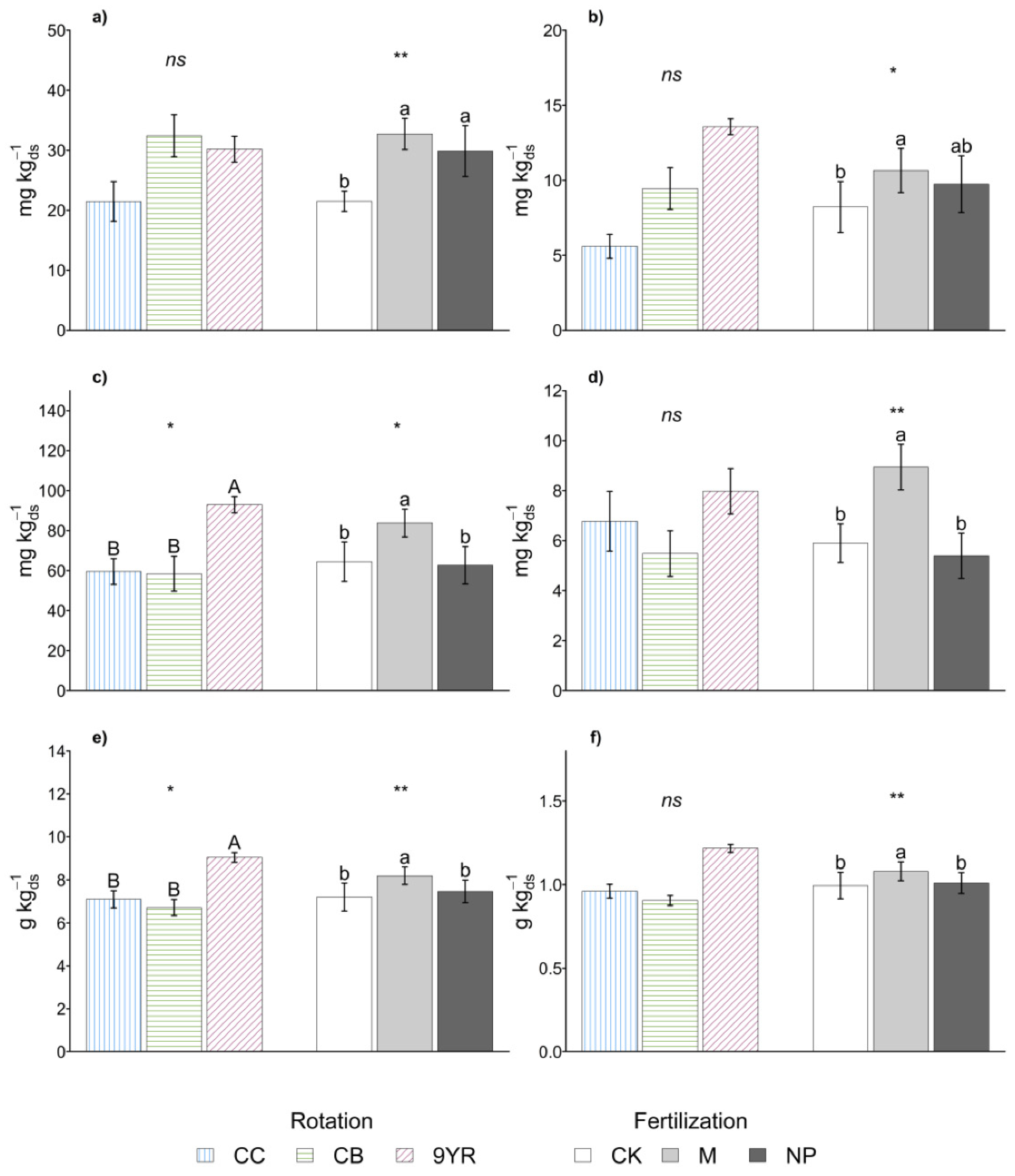
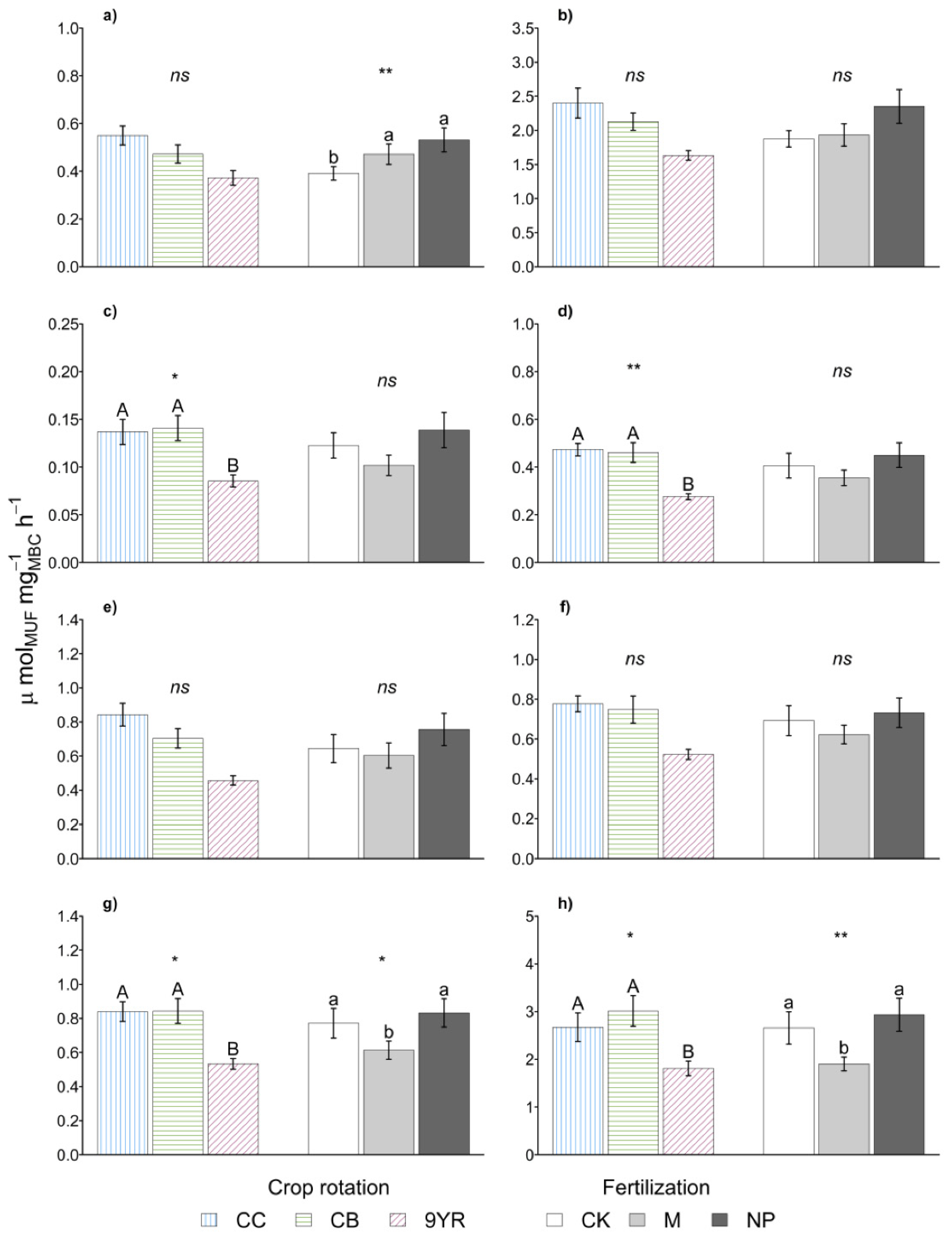
3.1. Crop Rotations Effects on Soil Biochemical Indicators
3.2. Fertilization Treatments Effects on Soil Biochemical Indicators
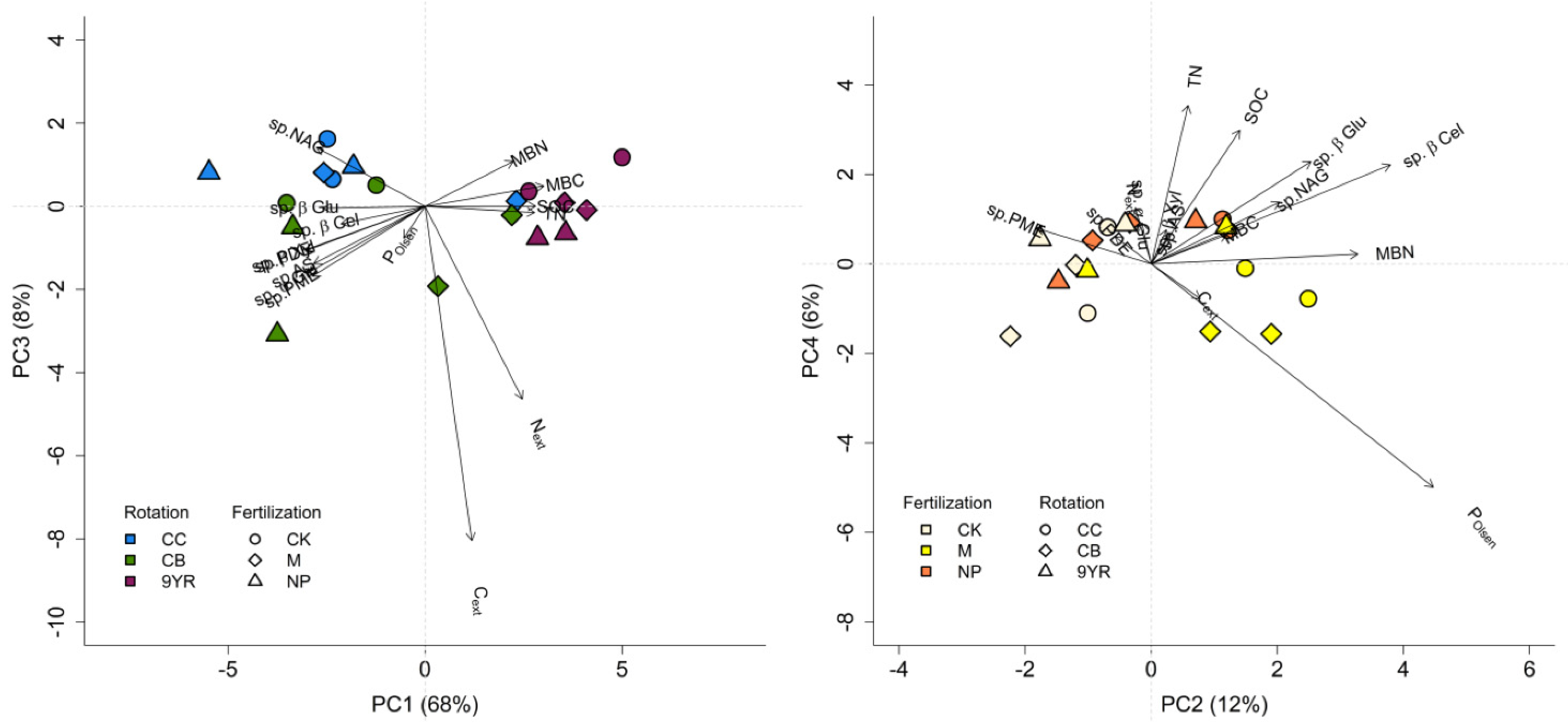
3.3. Principal Component Analysis
4. Discussion
4.1. The Crop Rotations Factor
4.2. The Fertilization Factor
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Riedell, W.E.; Pikul, J.L.; Jaradat, A.A.; Schumacher, T.E. Crop rotation and nitrogen input effects on soil fertility, maize mineral nutrition, yield and seed composition. Agron. J. 2009, 101, 870–879. [Google Scholar] [CrossRef]
- Liu, X.; Herbert, S.J.; Hashemi, A.M.; Zhang, X.; Ding, G. Effects of agricultural management on soil organic matter and carbon transformation—A review. Plant Soil Environ. 2006, 52, 531–543. [Google Scholar] [CrossRef]
- Norris, C.E.; Congreves, K.A. Alternative management practices improve soil health indices in intensive vegetable cropping systems: A review. Front. Environ. Sci. 2018, 6, 50. [Google Scholar] [CrossRef]
- Acosta-Martinez, V.; Zobeck, T.M.; Allen, V. Soil microbial, chemical and physical properties in continuous cotton and integrated crop-livestock systems. Soil Sci. Soc. Am. J. 2004, 68, 1875–1884. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Van Der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Ball, B.C.; Bingham, I.; Rees, R.M.; Watson, C.A.; Litterick, A. The role of crop rotations in determining soil structure and crop growth conditions. Can. J. Soil Sci. 2005, 85, 557–577. [Google Scholar] [CrossRef]
- Wortman, S.E.; Galusha, T.D.; Mason, S.T.; Francis, C.A. Soil fertility and crop yields in long-term organic and conventional cropping systems in Eastern Nebraska. Renew. Agric. Food Syst. 2011, 27, 200–216. [Google Scholar] [CrossRef]
- Karlen, D.L.; Varvel, G.E.; Bullock, D.G.; Cruse, R.M. Crop rotations for the 21st century. Adv. Agron. 1994, 53, 1–44. [Google Scholar]
- Kaiser, V.M. Metabolism and growth. In Mineral Nutrition of Plants: Principles and Perspectives; Epstein, E., Bloomm, A.J., Eds.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2005. [Google Scholar]
- Riley, H.; Pommeresche, R.; Eltun, R.; Hansen, S.; Korsaeth, A. Soil structure, organic matter and earthworm activity in a comparison of cropping systems with contrasting tillage, rotations, fertilizer levels and manure use. Agric. Ecosyst. Environ. 2008, 124, 275–284. [Google Scholar] [CrossRef]
- Lehmann, J.; Lan, Z.; Hyland, C.; Sato, S.; Solomon, D.; Ketterins, Q.M. Long-Term Dynamics of Phosphorus Forms and Retention in Manure-Amended Soils. Environ. Sci. Technol. 2005, 39, 6672–6680. [Google Scholar] [CrossRef]
- Ogiyama, S.; Sakamoto, K.; Suzuki, H.; Ushio, S.; Anzai, T.; Inubushi, K. Accumulation of Zinc and Copper in an Arable Field after Animal Manure Application. Soil Sci. Plant Nutr. 2005, 51, 801–808. [Google Scholar] [CrossRef]
- Ghirardini, A.; Grillini, V.; Verlicchi, P. A review of the occurrence of selected micropollutants and microorganisms in different raw and treated manure—Environmental risk due to antibiotics after application to soil. Sci. Total Environ. 2020, 707, 136118. [Google Scholar] [CrossRef]
- Courtier-Murias, D.; Simpson, A.J.; Marzadori, C.; Baldoni, G.; Ciavatta, C.; Fernández, J.M.; López-de-Sá, E.G.; Plaza, C. Unraveling the long-term stabilization mechanisms of organic materials in soils by physical fractionation and NMR spectroscopy. Agric. Ecosyst. Environ. 2013, 171, 9–18. [Google Scholar] [CrossRef]
- Giacometti, C.; Demyan, M.S.; Cavani, L.; Marzadori, C.; Ciavatta, C.; Kandeler, E. Chemical and microbiological soil quality indicators and their potential to differentiate fertilization regimes in temperate agroecosystems. Appl. Soil Ecol. 2013, 64, 32–48. [Google Scholar] [CrossRef]
- Mazzon, M.; Cavani, L.; Margon, A.; Sorrenti, G.; Ciavatta, C.; Marzadori, C. Changes in soil phenol oxidase activities due to long-term application of compost and mineral N in a walnut orchard. Geoderma 2018, 316, 70–77. [Google Scholar] [CrossRef]
- Triberti, L.; Nastri, A.; Baldoni, G. Long-term effects of crop rotation, manure and mineral fertilisation on carbon sequestration and soil fertility. Eur. J. Agron. 2016, 74, 47–55. [Google Scholar] [CrossRef]
- West, T.O.; Post, W.M. Soil organic carbon sequestration rates by tillage and crop rotation: A global data analysis. Soil Sci. Soc. Am. J. 2002, 66, 1930–1946. [Google Scholar] [CrossRef]
- Campbell, C.A.; Zentener, R.P.; Liang, B.C.; Roloff, G.; Gregorich, E.G.; Blomert, B. Organic C accumulation in soil over 30 years in semiarid southwestern Saskatchewan—Effect of crop rotation and fertilizers. Can. J. Soil Sci. 2000, 80, 179–192. [Google Scholar] [CrossRef]
- Karlen, D.L.; Cambardella, C.A.; Kovar, J.L.; Colvin, T.S. Soil quality response to long-term tillage and crop rotation practices. Soil Tillage Res. 2013, 133, 54–64. [Google Scholar] [CrossRef]
- Körschens, M. The importance of long-term field experiments for soil science and environmental research—A review. Plant Soil Environ. 2006, 52, 1–8. [Google Scholar]
- D’Hose, T.; Cougnon, M.; De Vliegher, A.; Vandecasteele, B.; Viaene, N.; Cornelis, W.; Van Bockstaele, E.; Reheul, D. The positive relationship between soil quality and crop production: A case study on the effect of farm compost application. Appl. Soil Ecol. 2014, 75, 189–198. [Google Scholar] [CrossRef]
- Luo, G.; Li, L.; Friman, V.P.; Guo, J.; Guo, S.; Shen, Q.; Ling, N. Organic amendments increase crop yields by improving microbe-mediated soil functioning of agroecosystems: A meta-analysis. Soil Biol. Biochem. 2018, 124, 105–115. [Google Scholar] [CrossRef]
- Song, Y.N.; Zhang, F.S.; Marschner, P.; Fan, F.L.; Gao, H.M.; Bao, X.G.; Sun, J.H.; Li, L. Effect of intercropping on crop yield and chemical and microbiological properties in rhizosphere of wheat (Triticum aestivum L.), maize (Zea mays L.) and faba bean (Vicia faba L.). Biol. Fertil. Soils 2007, 43, 565–574. [Google Scholar] [CrossRef]
- Plaza, C.; Gollany, H.T.; Baldoni, G.; Polo, A.; Ciavatta, C. Predicting long-term organic carbon dynamics in organically amended soils using the CQESTR model. J. Soils Sediments 2012, 12, 486–493. [Google Scholar] [CrossRef]
- Triberti, L.; Nastri, A.; Giordani, G.; Comellini, F.; Baldoni, G.; Toderi, G. Can mineral and organic fertilization help sequestrate carbon dioxide in cropland? Eur. J. Agron. 2008, 29, 13–20. [Google Scholar] [CrossRef]
- ISO 14256-2. Soil Quality—Determination of Nitrate, Nitrite and Ammonium in Field-Moist Soils by Extraction with Potassium Chloride Solution—Part 2: Automated Method with Segmented Flow Analysis; International Organization for Standardization: Geneva, Switzerland, 2005. [Google Scholar]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate (No. 939); US Department of Agriculture Circular: Washington, DC, USA, 1954. [Google Scholar]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil N: A rapid direct extraction method to measure microbial biomass in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Swisher, R.; Carroll, G.C. Fluorescein diacetate hydrolysis as an estimator of microbial biomass on coniferous needle surfaces. Microb. Ecol. 1980, 6, 217–226. [Google Scholar] [CrossRef]
- Giacometti, C.; Cavani, L.; Baldoni, G.; Ciavatta, C.; Marzadori, C.; Kandeler, E. Microplate-scale fluorometric soil enzyme assays as tools to assess soil quality in a long-term agricultural field experiment. Appl. Soil Ecol. 2014, 75, 80–85. [Google Scholar] [CrossRef]
- German, D.P.; Weintraub, M.N.; Grandy, A.S.; Lauber, C.L.; Rinkes, Z.L.; Allison, S.D. Optimization of hydrolytic and oxidative enzyme methods for ecosystem studies. Soil Biol. Biochem. 2011, 43, 1387–1397. [Google Scholar] [CrossRef]
- Gil-Sotres, F.; Trasar-Cepeda, C.; Leirós, M.C.; Seoane, S. Different approaches to evaluating soil quality using biochemical properties. Soil Biol. Biochem. 2005, 37, 877–887. [Google Scholar] [CrossRef]
- Kandeler, E.; Eder, G. Effect of cattle slurry in grassland on microbial biomass and on activities of various enzymes. Biol. Fertil. Soils 1993, 16, 249–254. [Google Scholar] [CrossRef]
- Trasar-Cepeda, C.; Leirós, M.C.; Gil-Sotres, F. Hydrolytic enzyme activities in agricultural and forest soils. Some implications for their use as indicators of soil quality. Soil Biol. Biochem. 2008, 40, 2146–2155. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- McDaniel, M.D.; Grandy, S. Soil microbial biomass and function are altered by 12 years of crop rotation. Soil J. 2016, 2, 583–599. [Google Scholar] [CrossRef]
- Doran, J.W.; Coleman, D.C.; Bezdicek, D.F.; Stewart, B.A. Defining Soil Quality for a Sustainable Environment; Special Pubblication 35; Soil Science Society of America: Madison, WI, USA, 1994. [Google Scholar]
- Wiesmeier, M.; Urbanskia, L.; Hobleya, E.; Langc, B.; von Lützowa, M.; Marin-Spiottad, E.; van Wesemaele, B.; Rabotf, E.; Ließf, M.; Garcia-Francoa, N.; et al. Soil organic carbon storage as a key function of soils—A review of drivers and indicators at various scales. Geoderma 2019, 333, 149–162. [Google Scholar] [CrossRef]
- Glasener, K.M.; Wagger, M.G.; MacKown, C.T.; Volk, R.J. Contributions of shoot and root nitrogen-15 labelled legume nitrogen sources to a sequence of three cereal crops. Soil Biol. Biochem. 2002, 66, 523–530. [Google Scholar]
- Drinkwater, L.E.; Wagoner, P.; Sarrantonio, M. Legume-based cropping systems have reduced carbon and nitrogen losses. Nature 1998, 396, 262–265. [Google Scholar] [CrossRef]
- Welsh, C.; Tenuta, M.; Flaten, D.N.; Thiessen-Martens, J.R.; Entz, M.H. High yielding organic crop management decreases plant-available but not recalcitrant soil phosphorous. Agron. J. 2009, 101, 1027–1035. [Google Scholar] [CrossRef]
- Moore, J.M.; Klose, S.; Tabatabai, M.A. Soil microbial biomass carbon and nitrogen as affected by cropping systems. Boil Fertil Soils 2000, 31, 200–210. [Google Scholar] [CrossRef]
- Acosta-Martínez, V.; Tabatabai, M.A. Phosphorous Cycle Enzymes. In Methods of Soil Enzymology; Dick, R.P., Ed.; Soil Science Society of America: Madison, WI, USA, 2011; pp. 161–183. [Google Scholar]
- Dick, R.P. A review: Long-term effects of agricultural systems on soil biochemical and microbial parameters. Agric. Ecosyst. Environ. 1992, 40, 25–36. [Google Scholar] [CrossRef]
- Green, C.J.; Blackmer, A.M.; Horton, R. Nitrogen effects on conservation of carbon during corn residue decomposition in soil. Soil Sci. Soc. Am. J. 1995, 59, 453–459. [Google Scholar] [CrossRef]
- Bullock, L.R.; Brosius, M.; Evanylo, G.K.; Ristaino, J.B. Organic and synthetic fertility amendments influence soil microbial, physical and chemical properties on organic and conventional farms. Appl. Soil Ecol. 2002, 19, 147–160. [Google Scholar] [CrossRef]
- Kandeler, E.; Stemmer, M.; Klimanek, E. Response of soil microbial biomass, urease and xylanase within particle size fractions to long-term soil management. Soil Biol. Biochem. 1999, 31, 261–273. [Google Scholar] [CrossRef]
- Böhme, L.; Langer, U.; Böhme, F. Microbial biomass, enzyme activities and microbial community structure in two European long-term field experiments. Agric. Ecosyst. Environ. 2005, 109, 141–152. [Google Scholar] [CrossRef]
- Marinari, S.; Mancinelli, R.; Campiglia, E.; Grego, S. Chemical and biological indicators of soil quality in organic and conventional farming systems in central Italy. Ecol. Indic. 2006, 6, 701–711. [Google Scholar] [CrossRef]
- Liang, B.C.; MacKenzie, A.F.; Schnitzer, M.; Monreal, C.M.; Voroney, P.R.; Beyaert, R.P. Management-induced changes in labile soil organic matter under continuous corn in eastern Canadian soils. Biol. Fertil. Soils 1998, 26, 88–94. [Google Scholar] [CrossRef]
- Wallenstein, M.D.; McNulty, S.; Fernandez, I.J.; Boggs, J.; Schlesinger, W.H. Nitrogen fertilization decreases forest soil fungal and bacterial biomass in three long-term experiments. For. Ecol. Manag. 2006, 222, 459–468. [Google Scholar] [CrossRef]
- Chen, J.; Luo, Y.; Van Groenigen, K.J.; Hungate, B.A.; Cao, J.; Zhou, X.; Wang, R.W. A keystone microbial enzyme for nitrogen control of soil carbon storage. Sci. Adv. 2018, 4, 2–8. [Google Scholar] [CrossRef]
- Zhang, Q.; Feng, J.; Wu, J.; Zhang, D.; Chen, Q.; Li, Q.; Long, C.; Feyissa, A.; Cheng, X. Variations in carbon-decomposition enzyme activities respond differently to land use change in central China. Land Degrad. Dev. 2019, 30, 459–469. [Google Scholar] [CrossRef]
- Hardarson, G.; Atkins, C. Optimising biological N2 fixation by legumes in farming systems. Plant Soil 2003, 252, 41–54. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | pH | C:N | NH4+-N | NO3−-N | POlsen | MBC:SOC | FDA | Yield |
|---|---|---|---|---|---|---|---|---|
| mg kg−1 | mg kg−1 | mg kg−1 | % | mg kg−1 h−1 | Mgdm ha−1 | |||
| Rotation (R) | ||||||||
| CC | 6.40 | 7.37 | 5.51 | 6.46 | 23.6 | 0.828 | 44.1 | 4.89 |
| CB | 6.03 | 7.40 | 6.12 | 10.6 | 22.5 | 0.854 | 43.4 | 3.60 |
| 9YR | 6.00 | 7.43 | 2.37 | 17.1 | 14.3 | 1.03 | 54.2 | 4.74 |
| p-value | 0.724 | 0.955 | 0.263 | 0.073 | 0.166 | 0.058 | 0.194 | 0.087 |
| Fertilization (F) | ||||||||
| CK | 6.07 b | 7.22 | 5.23 | 9.69 b | 14.1 c | 0.874 | 43.7 b | 2.87 b |
| NP | 5.94 b | 7.38 | 4.77 | 11.5 ab | 18.1 b | 0.820 | 48.1 ab | 5.72 a |
| M | 6.42 a | 7.65 | 3.44 | 12.6 a | 26.4 a | 1.03 | 50.0 a | 4.64 a |
| p-value | 0.025 | 0.058 | 0.086 | 0.047 | 0.002 | 0.064 | 0.041 | 0.004 |
| R × F | ||||||||
| p-value | 0.216 | 0.180 | 0.436 | 0.177 | 0.063 | 0.475 | 0.225 | 0.093 |
| Factors | Hydrolytic Enzyme Activities (nmolMUF g−1ds h−1) | |||||||
|---|---|---|---|---|---|---|---|---|
| β-Cell | β-glu | α-glu | β-xyl | NAG | AS | PDE | PME | |
| Rotation (R) | ||||||||
| CC | 32.2 | 139 | 7.79 | 27.5 | 48.7 | 45.1 | 48.4 | 151 |
| CB | 27.3 | 121 | 7.76 | 25.4 | 40.2 | 41.3 | 46.2 | 162 |
| 9YR | 34.8 | 152 | 7.83 | 25.5 | 42.3 | 48.5 | 49.1 | 165 |
| p-value | 0.281 | 0.147 | 0.990 | 0.650 | 0.100 | 0.237 | 0.761 | 0.793 |
| Fertilization (F) | ||||||||
| CK | 24.6 c | 116 b | 7.31 | 23.9 b | 41.4 | 41.4 b | 45.7 | 155 |
| NP | 31.4 b | 139 a | 7.86 | 25.9 b | 42.8 | 42.8 b | 48.4 | 168 |
| M | 39.4 a | 160 a | 8.29 | 28.6 a | 50.6 | 50.6 a | 49.5 | 153 |
| p-value | <0.001 | 0.011 | 0.468 | 0.008 | 0.065 | 0.032 | 0.275 | 0.284 |
| R × F | ||||||||
| p-value | 0.042 | 0.139 | 0.625 | 0.316 | 0.266 | 0.442 | 0.306 | 0.225 |
| Factors | Ecosystem Ratios | ||
|---|---|---|---|
| enz C:N | enz C:P | enz N:P | |
| Rotation (R) | |||
| CC | 1.37 | 1.01 | 0.733 |
| CB | 1.42 | 0.969 | 0.684 |
| 9YR | 1.44 | 1.00 | 0.696 |
| p-value | 0.297 | 0.401 | 0.240 |
| Fertilization (F) | |||
| CK | 1.41 | 0.969 b | 0.686 b |
| NP | 1.41 | 0.986 b | 0.700 b |
| M | 1.40 | 1.03 a | 0.738 a |
| p-value | 0.967 | 0.001 | 0.013 |
| R × F | |||
| p-value | 0.487 | 0.025 | 0.075 |
| ANOVA Parameter | PC1 | PC2 | PC3 | PC4 | ||||
|---|---|---|---|---|---|---|---|---|
| Proportion of variance (%) | 68 | 12 | 8 | 6 | ||||
| ANOVA factor | F | p-value | F | p-value | F | p-value | F | p-value |
| Rotation (R) | 24.5 | <0.001 | 4.19 | 0.057 | 6.44 | 0.022 | 4.97 | 0.040 |
| Fertilization (F) | 13.8 | 0.029 | 13.2 | 0.003 | 3.95 | 0.064 | 6.86 | 0.018 |
| R × F | 1.12 | 0.410 | 1.17 | 0.392 | 0.72 | 0.602 | 2.70 | 0.109 |
| Significant comparison | 9YR > CB = CC M > CK > NP | CC = CB = 9YR M > NP > CK | CC > 9YR > CB NP = M = CK | 9YR > CC > CB NP > CK > M | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giacometti, C.; Mazzon, M.; Cavani, L.; Triberti, L.; Baldoni, G.; Ciavatta, C.; Marzadori, C. Rotation and Fertilization Effects on Soil Quality and Yields in a Long Term Field Experiment. Agronomy 2021, 11, 636. https://doi.org/10.3390/agronomy11040636
Giacometti C, Mazzon M, Cavani L, Triberti L, Baldoni G, Ciavatta C, Marzadori C. Rotation and Fertilization Effects on Soil Quality and Yields in a Long Term Field Experiment. Agronomy. 2021; 11(4):636. https://doi.org/10.3390/agronomy11040636
Chicago/Turabian StyleGiacometti, Caterina, Martina Mazzon, Luciano Cavani, Loretta Triberti, Guido Baldoni, Claudio Ciavatta, and Claudio Marzadori. 2021. "Rotation and Fertilization Effects on Soil Quality and Yields in a Long Term Field Experiment" Agronomy 11, no. 4: 636. https://doi.org/10.3390/agronomy11040636
APA StyleGiacometti, C., Mazzon, M., Cavani, L., Triberti, L., Baldoni, G., Ciavatta, C., & Marzadori, C. (2021). Rotation and Fertilization Effects on Soil Quality and Yields in a Long Term Field Experiment. Agronomy, 11(4), 636. https://doi.org/10.3390/agronomy11040636