
Organic Cultivation and Deficit Irrigation Practices to Improve Chemical and Biological Activity of Mentha spicata Plants





Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Growing Conditions
2.2. Insects
2.3. Cultivation Plan
2.4. Physiological Measurements and Plant Growth Parameters
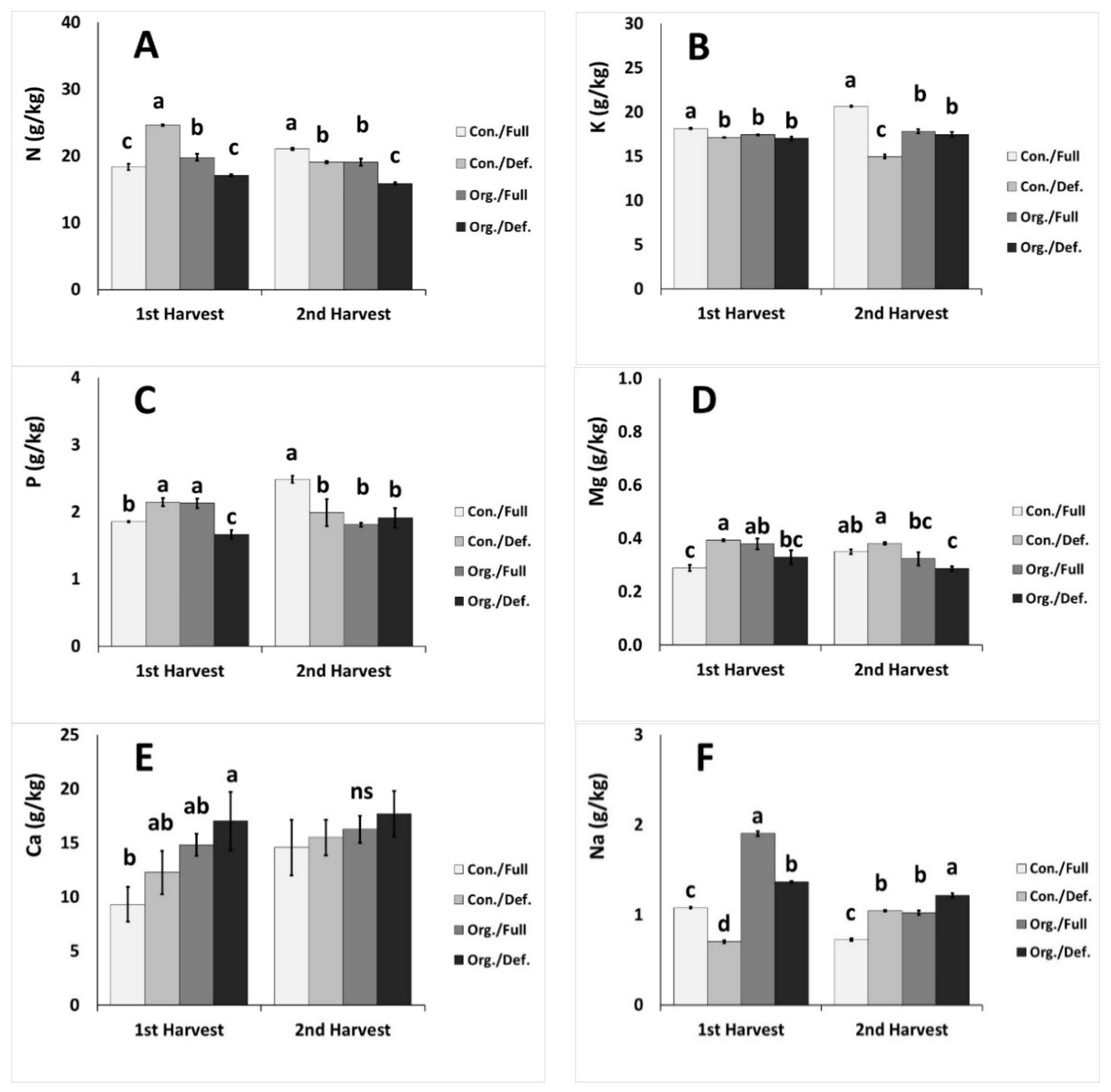
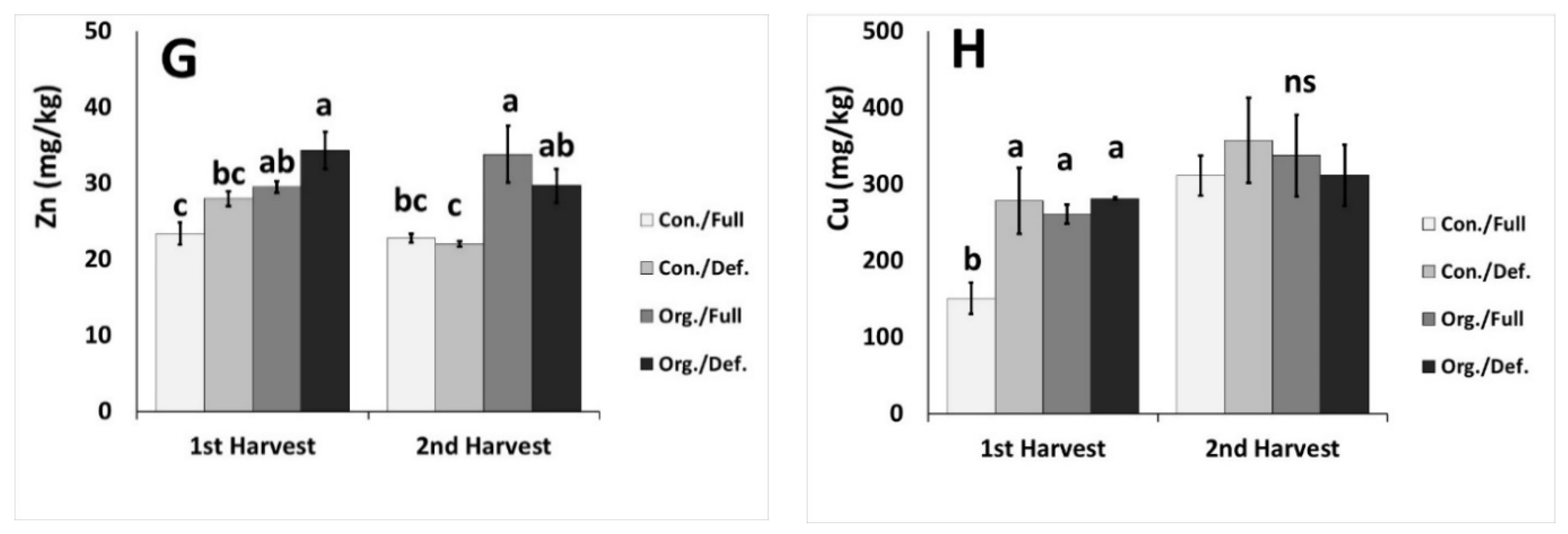
2.5. Nutrient Content in Plant Tissue
2.6. Essential Oil Extraction and Compound Identification
2.7. Total Phenols and Flavonoids Content and Antioxidant Activity of Plant Extracts
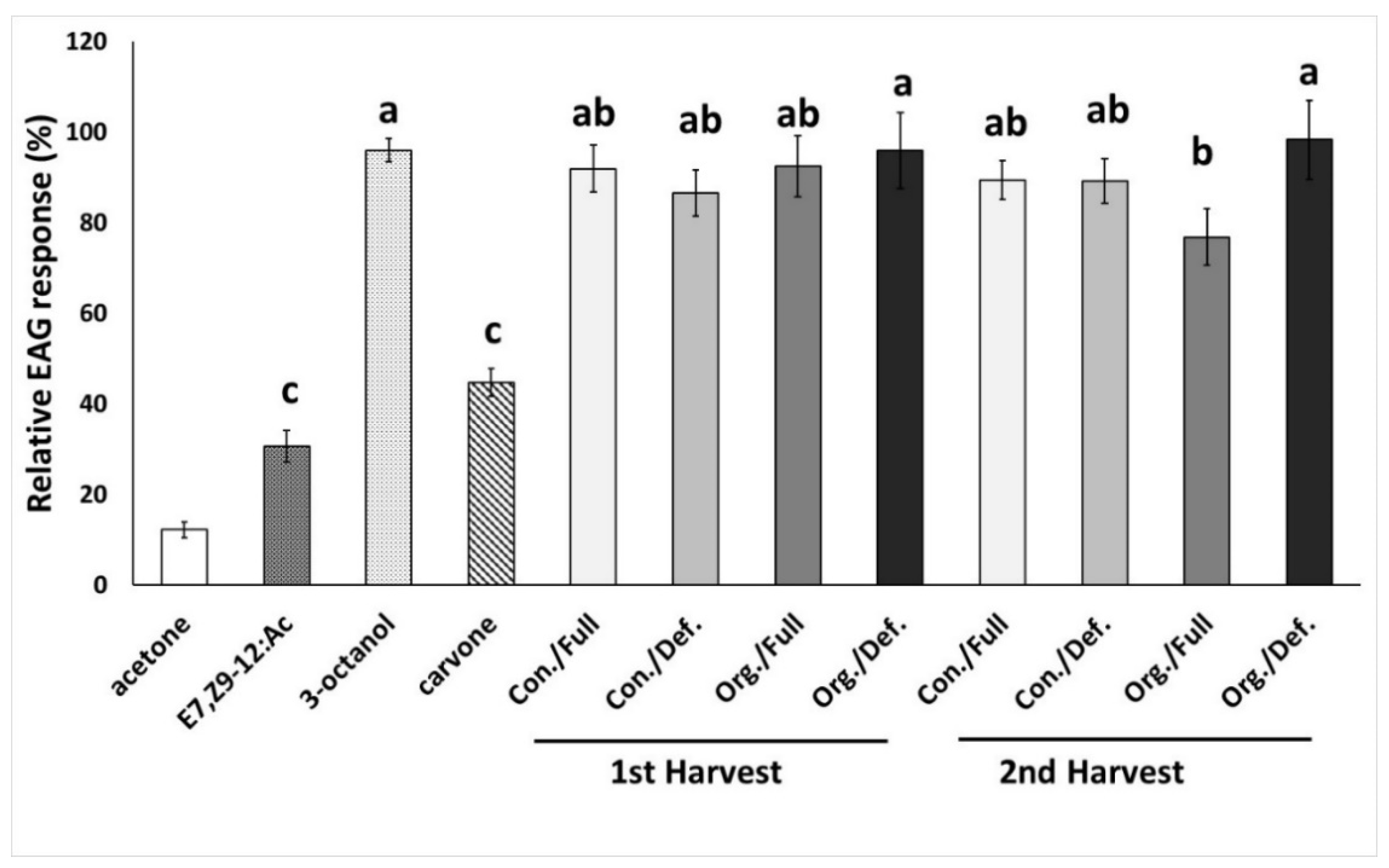
2.8. Electroantennographic Recordings
2.9. Larvicidal Bioassays
2.10. Statistical Methods
3. Results
3.1. Electroantennographic Response of Female L. botrana Adult Insects
3.2. Larvicidal Bioassays
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Setter, T.L.; Waters, I. Review of prospects for germplasm improvement for waterlogging tolerance in wheat, barley and oats. Plant Soil 2003, 253, 1–34. [Google Scholar] [CrossRef]
- Järvan, M.; Edesi, L. The effect of cultivation methods on the yield and biological quality of potato. Agron. Res. 2009, 7, 289–299. [Google Scholar]
- Chrysargyris, A.; Kloukina, C.; Vassiliou, R.; Tomou, E.-M.; Skaltsa, H.; Tzortzakis, N. Cultivation strategy to improve chemical profile and anti-oxidant activity of Sideritis perfoliata L. subsp. perfoliata. Ind. Crops Prod. 2019, 140, 111694. [Google Scholar] [CrossRef]
- Manik, S.M.N.; Pengilley, G.; Dean, G.; Field, B.; Shabala, S.; Zhou, M. Soil and crop management practices to minimize the impact of waterlogging on crop productivity. Front. Plant Sci. 2019, 10, 1–23. [Google Scholar] [CrossRef]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef]
- Kopittke, P.M.; Menzies, N.W.; Wang, P.; McKenna, B.A.; Lombi, E. Soil and the intensification of agriculture for global food security. Environ. Int. 2019, 132, 105078. [Google Scholar] [CrossRef] [PubMed]
- Parkash, V.; Singh, S. A review on potential plant-basedwater stress indicators for vegetable crops. Sustainability 2020, 12, 3945. [Google Scholar] [CrossRef]
- Bourn, D.; Prescott, J. A comparison of the nutritional value, sensory qualities, and food safety of organically and conventionally produced foods. Crit. Rev. Food Sci. Nutr. 2002, 42, 1–34. [Google Scholar] [CrossRef]
- Barbieri, P.; Pellerin, S.; Nesme, T. Comparing crop rotations between organic and conventional farming. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Litskas, V.; Chrysargyris, A.; Stavrinides, M.; Tzortzakis, N. Water-energy-food nexus: A case study on medicinal and aromatic plants. J. Clean. Prod. 2019, 233, 1334–1343. [Google Scholar] [CrossRef]
- Ahmad MALIK, A.; Suryapani, S.; Ahmad, J. Chemical Vs Organic Cultivation of Medicinal and Aromatic Plants: The choice is clear. Int. J. Med. Arom. Plants 2011, 1, 5–13. [Google Scholar]
- Lv, J.; Huang, H.; Yu, L.; Whent, M.; Niu, Y.; Shi, H.; Wang, T.T.Y.; Luthria, D.; Charles, D.; Yu, L.L. Phenolic composition and nutraceutical properties of organic and conventional cinnamon and peppermint. Food Chem. 2012, 132, 1442–1450. [Google Scholar] [CrossRef]
- Anwar, M.; Patra, D.D.; Chand, S.; Alpesh, K.; Naqvi, A.A.; Khanuja, S.P.S. Effect of organic manures and inorganic fertilizer on growth, herb and oil yield, nutrient accumulation, and oil quality of French basil. Commun. Soil Sci. Plant Anal. 2005, 36, 1737–1746. [Google Scholar] [CrossRef]
- Hallmann, E.; Sabała, P. Organic and conventional herbs quality reflected by their antioxidant compounds concentration. Appl. Sci. 2020, 10, 3468. [Google Scholar] [CrossRef]
- Bettaieb, I.; Zakhama, N.; Wannes, W.A.; Kchouk, M.E.; Marzouk, B. Water deficit effects on Salvia officinalis fatty acids and essential oils composition. Sci. Hortic. 2009, 120, 271–275. [Google Scholar] [CrossRef]
- Bettaieb Rebey, I.; Jabri-Karoui, I.; Hamrouni-Sellami, I.; Bourgou, S.; Limam, F.; Marzouk, B. Effect of drought on the biochemical composition and antioxidant activities of cumin (Cuminum cyminum L.) seeds. Ind. Crops Prod. 2012, 36, 238–245. [Google Scholar] [CrossRef]
- Okwany, R.O.; Peters, R.T.; Ringer, K.L.; Walsh, D.B. Sustained deficit irrigation effects on peppermint yield and oil quality in the semi-arid pacific northwest, USA. Appl. Eng. Agric. 2012, 28, 551–558. [Google Scholar] [CrossRef]
- García-Caparrós, P.; Romero, M.J.; Llanderal, A.; Cermeño, P.; Lao, M.T.; Segura, M.L. Effects of drought stress on biomass, essential oil content, nutritional parameters, and costs of production in six Lamiaceae species. Water 2019, 11, 573. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Laoutari, S.; Litskas, V.D.; Stavrinides, M.C.; Tzortzakis, N. Effects of water stress on lavender and sage biomass production, essential oil composition and biocidal properties against Tetranychus urticae (Koch). Sci. Hortic. 2016, 213, 96–103. [Google Scholar] [CrossRef]
- Németh-Zámbori, É.; Szabó, K.; Pluhár, Z.; Radácsi, P.; Inotai, K. Changes in biomass and essential oil profile of four Lamiaceae species due to different soil water levels. J. Essent. Oil Res. 2016, 28, 391–399. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Xylia, P.; Botsaris, G.; Tzortzakis, N. Antioxidant and antibacterial activities, mineral and essential oil composition of spearmint (Mentha spicata L.) affected by the potassium levels. Ind. Crops Prod. 2017, 103, 202–212. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Loupasaki, S.; Petropoulos, S.A.; Tzortzakis, N. Salinity and cation foliar application: Implications on essential oil yield and composition of hydroponically grown spearmint plants. Sci. Hortic. 2019, 256, 108581. [Google Scholar] [CrossRef]
- Telci, I.; Demirtas, I.; Bayram, E.; Arabaci, O.; Kacar, O. Environmental variation on aroma components of pulegone/piperitone rich spearmint (Mentha spicata L.). Ind. Crops Prod. 2010, 32, 588–592. [Google Scholar] [CrossRef]
- Hossain, M.B.; Barry-Ryan, C.; Martin-Diana, A.B.; Brunton, N.P. Effect of drying method on the antioxidant capacity of six Lamiaceae herbs. Food Chem. 2010, 123, 85–91. [Google Scholar] [CrossRef]
- Ruberto, G.; Baratta, M.T.; Deans, S.G.; Dorman, H.J.D. Antioxidant and antimicrobial activity of Foeniculum vulgare and Crithmum maritimum essential oils. Planta Med. 2000, 66, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Scherer, R.; Lemos, M.F.; Lemos, M.F.; Martinelli, G.C.; Martins, J.D.L.; da Silva, A.G. Antioxidant and antibacterial activities and composition of Brazilian spearmint (Mentha spicata L.). Ind. Crops Prod. 2013, 50, 408–413. [Google Scholar] [CrossRef]
- Yegen, O.; Berger, B.; Heitefuss, R. Investigations on the fungitoxicity of extracts of six selected plants from Turkey against phytopathogenic fungi. Z. Pflanzenkr. Pflanzenschutz 1992, 99, 349–359. [Google Scholar]
- Kumar, P.; Mishra, S.; Malik, A.; Satya, S. Insecticidal properties of Mentha species: A review. Ind. Crops Prod. 2011, 34, 802–817. [Google Scholar] [CrossRef]
- Ercan, F.S.; Baş, H.; Koç, M.; Pandir, D.; Öztemiz, S. Insecticidal activity of essential oil of Prangos ferulacea (Umbelliferae) against Ephestia kuehniella (Lepidoptera: Pyralidae) and Trichogramma embryophagum (Hymenoptera: Trichogrammatidae). Turk. J. Agric. For. 2013, 37, 719–725. [Google Scholar] [CrossRef]
- Sarma, R.; Adhikari, K.; Mahanta, S.; Khanikor, B. Combinations of Plant Essential Oil Based Terpene Compounds as Larvicidal and Adulticidal Agent against Aedes aegypti (Diptera: Culicidae). Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Singh, P.; Pandey, A.K. Prospective of essential oils of the genus mentha as biopesticides: A review. Front. Plant Sci. 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Attia, S.; Grissa, K.L.; Ghrabi, Z.G.; Mailleux, A.C.; Lognay, G.; Hance, T. Acaricidal activity of 31 essential oils extracted from plants collected in Tunisia. J. Essent. Oil Res. 2012, 24, 279–288. [Google Scholar] [CrossRef]
- Du, Y.; Zhou, A.; Chen, J. Olfactory and behavioral responses of red imported fire ants, solenopsis invicta, to ylang ylang oil and its components. J. Pest Sci. 2021. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Xylia, P.; Antoniou, O.; Tzortzakis, N. Climate change due to heat and drought stress can alter the physiology of Maratheftiko local cyprian grapevine variety. J. Water Clim. Chang. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Tzortzakis, N.; Chrysargyris, A.; Aziz, A. Adaptive response of a native mediterranean grapevine cultivar upon short-term exposure to drought and heat stress in the context of climate change. Agronomy 2020, 10, 249. [Google Scholar] [CrossRef]
- Heyman, L.; Chrysargyris, A.; Demeestere, K.; Tzortzakis, N.; Höfte, M. Responses to drought stress modulate the susceptibility to Plasmopara viticola in Vitis vinifera self-rooted cuttings. Plants 2021, 10, 273. [Google Scholar] [CrossRef] [PubMed]
- Chrysargyris, A.; Panayiotou, C.; Tzortzakis, N. Nitrogen and phosphorus levels affected plant growth, essential oil composition and antioxidant status of lavender plant (Lavandula angustifolia Mill.). Ind. Crops Prod. 2016, 83, 577–586. [Google Scholar] [CrossRef]
- Meyers, K.J.; Watkins, C.B.; Pritts, M.P.; Liu, R.H. Antioxidant and Antiproliferative Activities of Strawberries. J. Agric. Food Chem. 2003, 51, 6887–6892. [Google Scholar] [CrossRef] [PubMed]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.S.P. Response of plants to water stress. Front. Plant Sci. 2014, 5, 1–8. [Google Scholar] [CrossRef]
- Pirzad, A.; Shakiba, M.R.; Zehtab-Salmasi, S.; Mohammadi, S.A.; Darvishzadeh, R.; Samadi, A. Effect of water stress on leaf relative water content, chlorophyll, proline and soluble carbohydrates in Matricaria chamomilla L. J. Med. Plants Res. 2011, 5, 2483–2488. [Google Scholar]
- Kazimierczak, R.; Hallmann, E.; Kazimierczyk, M.; Sokołowska, O.; Rembiałkowska, E. Wpływ ekologicznego i konwencjonalnego systemu uprawy na zawartość substancji bioaktywnych w roślinach zielarskich. Veg. Crops Res. Bull. 2011, 75, 133–144. [Google Scholar] [CrossRef]
- Kazimierczak, R.; Hallmann, E.; Rembiałkowska, E. Effects of organic and conventional production systems on the content of bioactive substances in four species of medicinal plants. Biol. Agric. Hortic. 2015, 31, 118–127. [Google Scholar] [CrossRef]
- Ouzounidou, G.; Asfi, M.; Sotirakis, N.; Papadopoulou, P.; Gaitis, F. Olive mill wastewater triggered changes in physiology and nutritional quality of tomato (Lycopersicon esculentum Mill.) depending on growth substrate. J. Hazard. Mater. 2008, 158, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Coley, P.D.; Bryant, J.P.; Chapin, F.S. Resource availability and plant antiherbivore defense. Science 1985, 230, 895–899. [Google Scholar] [CrossRef]
- Pagare, S.; Bhatia, M.; Tripathi, N.; Pagare, S.; Bansal, Y.K. Secondary metabolites of plants and their role: Overview. Curr. Trends Biotechnol. Pharm. 2015, 9, 293–304. [Google Scholar]
- Okwany, R.O.; Peters, T.R.; Ringer, K.L.; Walsh, D.B.; Rubio, M. Impact of sustained deficit irrigation on spearmint (Mentha spicata L.) biomass production, oil yield, and oil quality. Irrig. Sci. 2012, 30, 213–219. [Google Scholar] [CrossRef]
- De Carvalho, C.C.C.R.; Da Fonseca, M.M.R. Carvone: Why and how should one bother to produce this terpene. Food Chem. 2006, 95, 413–422. [Google Scholar] [CrossRef]
- Snoussi, M.; Noumi, E.; Trabelsi, N.; Flamini, G.; Papetti, A.; De Feo, V. Mentha spicata essential oil: Chemical composition, antioxidant and antibacterial activities against planktonic and biofilm cultures of vibrio spp. strains. Molecules 2015, 20, 14402–14424. [Google Scholar] [CrossRef] [PubMed]
- Koundal, R.; Dolma, S.K.; Chand, G.; Agnihotri, V.K.; Reddy, S.G.E. Chemical composition and insecticidal properties of essential oils against diamondback moth (Plutella xylostella L.). Toxin Rev. 2020, 39, 371–381. [Google Scholar] [CrossRef]
- Marino, S.; Ahmad, U.; Ferreira, M.I.; Alvino, A. Evaluation of the effect of irrigation on biometric growth, physiological response, and essential oil of Mentha spicata (L.). Water 2019, 11, 2264. [Google Scholar] [CrossRef]
- Chauhan, R.S.; Kaul, M.K.; Shahi, A.K.; Kumar, A.; Ram, G.; Tawa, A. Chemical composition of essential oils in Mentha spicata L. accession [IIIM(J)26] from North-West Himalayan region, India. Ind. Crops Prod. 2009, 29, 654–656. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Nikolaidou, E.; Stamatakis, A.; Tzortzakis, N. Vegetative, physiological, nutritional and antioxidant behavior of spearmint (Mentha spicata L.) in response to different nitrogen supply in hydroponics. J. Appl. Res. Med. Aromat. Plants 2017, 6, 52–61. [Google Scholar] [CrossRef]
- Katerinopoulos, H.E.; Pagona, G.; Afratis, A.; Stratigakis, N.; Roditakis, N. Composition and insect attracting activity of the essential oil of Rosmarinus officinalis. J. Chem. Ecol. 2005, 31, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Tasin, M.; Anfora, G.; Ioriatti, C.; Carlin, S.; De Cristofaro, A.; Schmidt, S.; Bengtsson, M.; Versini, G.; Witzgall, P. Antennal and behavioral responses of grapevine moth Lobesia botrana females to volatiles from grapevine. J. Chem. Ecol. 2005, 31, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Tasin, M.; Bäckman, A.C.; Coracini, M.; Casado, D.; Ioriatti, C.; Witzgall, P. Synergism and redundancy in a plant volatile blend attracting grapevine moth females. Phytochemistry 2007, 68, 203–209. [Google Scholar] [CrossRef]
- Tasin, M.; Bäckman, A.C.; Bengtsson, M.; Ioriatti, C.; Witzgall, P. Essential host plant cues in the grapevine moth. Naturwissenschaften 2006, 93, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Abdelgaleil, S.A.M.; Mohamed, M.I.E.; Badawy, M.E.I.; El-Arami, S.A.A. Fumigant and contact toxicities of monoterpenes to Sitophilus oryzae (L.) and Tribolium castaneum (Herbst) and their inhibitory effects on acetylcholinesterase activity. J. Chem. Ecol. 2009, 35, 518–525. [Google Scholar] [CrossRef]
- Kedia, A.; Prakash, B.; Mishra, P.K.; Chanotiya, C.S.; Dubey, N.K. Antifungal, antiaflatoxigenic, and insecticidal efficacy of spearmint (Mentha spicata L.) essential oil. Int. Biodeterior. Biodegrad. 2014, 89, 29–36. [Google Scholar] [CrossRef]
- De Souza, V.N.; de Oliveira, C.R.F.; Matos, C.H.C.; de Almeida, D.K.F. Fumigation toxicity of essential oils against Rhyzopertha dominica (F.) in stored maize grain. Rev. Caatinga 2016, 29, 435–440. [Google Scholar] [CrossRef]
- Koliopoulos, G.; Pitarokili, D.; Kioulos, E.; Michaelakis, A.; Tzakou, O. Chemical composition and larvicidal evaluation of Mentha, Salvia, and Melissa essential oils against the West Nile virus mosquito Culex pipiens. Parasitol. Res. 2010, 107, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Konstantopoulou, I.; Vassilopoulou, L.; Mavragani-Tsipidou, P.; Scouras, Z.G. Insecticidal effects of essential oils. A study of the effects of essential oils extracted from eleven Greek aromatic plants on Drosophila auraria. Experientia 1992, 48, 616–619. [Google Scholar] [CrossRef] [PubMed]
- Eliopoulos, P.A.; Hassiotis, C.N.; Andreadis, S.S.; Porichi, A.E.E. Fumigant toxicity of essential oils from basil and spearmint against two major pyralid pests of stored products. J. Econ. Entomol. 2015, 108, 805–810. [Google Scholar] [CrossRef]
- Govindarajan, M.; Sivakumar, R.; Rajeswari, M.; Yogalakshmi, K. Chemical composition and larvicidal activity of essential oil from Mentha spicata (Linn.) against three mosquito species. Parasitol. Res. 2012, 110, 2023–2032. [Google Scholar] [CrossRef]
- Pavlidou, V.; Karpouhtsis, I.; Franzios, G.; Zambetaki, A.; Scouras, Z.; Mavragani-Tsipidou, P. Insecticidal and genotoxic effects of essential oils of Greek sage, Salvia fruticosa, and mint, Mentha pulegium, on Drosophila melanogaster and Bactrocera oleae (Diptera: Tephritidae). J. Agric. Urban Entomol. 2004, 21, 39–49. [Google Scholar]
- Lee, B.H.; Annis, P.C.; Tumaalii, F.; Lee, S.E. Fumigant toxicity of Eucalyptus blakelyi and Melaleuca fulgens essential oils and 1,8-cineole against different development stages of the rice weevil Sitophilus oryzae. Phytoparasitica 2004, 32, 498–506. [Google Scholar] [CrossRef]
- Lucia, A.; Licastro, S.; Zerba, E.; Masuh, H. Yield, chemical composition, and bioactivity of essential oils from 12 species of Eucalyptus on Aedes aegypti larvae. Entomol. Exp. Appl. 2008, 129, 107–114. [Google Scholar] [CrossRef]
- Pohlit, A.M.; Rezende, A.R.; Lopes Baldin, E.L.; Lopes, N.P.; De Andrade Neto, V.F. Plant extracts, isolated phytochemicals, and plant-derived agents which are lethal to arthropod vectors of human tropical diseases—A review. Planta Med. 2011, 77, 618–630. [Google Scholar] [CrossRef] [PubMed]
- Martinez, L.; Soti, P.; Kaur, J.; Racelis, A.; Kariyat, R.R. Impact of cover crops on insect community dynamics in organic farming. Agriculture 2020, 10, 209. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Three-Way Anova | Cultivation | Irrigation | Harvesting | Cult. × Irrig. | Cult. × Harv. | Irrig. × Harv. | Cult. × Irrig. × Harv. |
|---|---|---|---|---|---|---|---|
| Height (cm) | ns | ns | ns | ns | ** | ns | * |
| Fresh weight—Fw (g) | ns | ns | ns | ns | ns | ns | ns |
| Dry matter content (%) | ns | ** | ns | ns | ns | ns | ns |
| Stomatal conductance (cm/s) | ns | * | ** | ns | * | * | ns |
| Chlorophyll fluorescence Fv/Fm | ns | ns | ** | ns | ns | ns | ns |
| SPAD | * | ns | ns | ns | * | * | ns |
| Total phenols (μmol GAE/g Fw) | ns | ns | *** | ns | ns | ns | ns |
| total flavonoids (mg Rutin/g Fw) | * | ns | *** | ns | ns | ns | ns |
| ABTS (mg Trolox/g Fw) | ns | ns | *** | ns | ns | ns | ns |
| DPPH (mg Trolox/g Fw) | ** | ** | *** | * | * | ns | ns |
| FRAP (mg Trolox/g Fw) | ns | ns | *** | ns | ns | ns | ns |
| N (g/kg) | *** | ns | *** | *** | ns | *** | *** |
| K (g/kg) | * | *** | ** | *** | ns | *** | *** |
| P (g/kg) | * | ns | ns | ns | ns | ns | ns |
| Mg (g/kg) | ns | ns | ns | *** | ** | ** | ** |
| Ca (g/kg) | ns | ns | * | ns | ns | ns | ns |
| Na (g/kg) | *** | *** | *** | *** | *** | *** | ns |
| Zn (mg/kg) | *** | ns | ns | ns | ns | ns | ns |
| Cu (mg/kg) | ns | ns | * | ns | ns | ns | ns |
| Essential oil yield (%) | ns | ns | ns | * | ** | * | ns |
| D-Limonene (%) | *** | ** | *** | ns | ns | *** | ** |
| Eucalyptol (%) | ns | ns | *** | * | ns | ** | ns |
| Carvone (%) | *** | ** | *** | ns | ns | * | ** |
| Monoterpenes hydrocarbons (%) | *** | * | *** | ns | ns | ** | *** |
| Sesquiterpenes hydrocarbons (%) | *** | ns | *** | ns | ns | * | ** |
| Oxygenated monoterpenes (%) | ns | ns | *** | ns | ns | ns | ns |
| Oxygenated sesquiterpenes (%) | ns | ns | *** | ns | ns | ns | ns |
| Treatment | Plant Height (cm) | Fresh Weight (g) | Dry Matter Content (%) | EO Yield (%) | |
|---|---|---|---|---|---|
| Cultivation/Irrigation Plan | 1st harvest | ||||
| Conventional/full | CI | 44.16 ± 1.46 a | 65.31 ± 8.81 a | 25.97 ± 0.91 bc | 1.96 ± 0.11 a |
| Conventional/deficit | CD | 34.14 ± 1.36 b | 45.27 ± 3.70 b | 27.37 ± 0.33 ab | 2.24 ± 0.12 a |
| Organic/full | OI | 34.41 ± 2.20 b | 38.12 ± 3.07 b | 24.54 ± 0.47 c | 2.47 ± 0.10 a |
| Organic/deficit | OD | 33.50 ± 2.04 b | 35.32 ± 3.35 b | 28.19 ± 0.69 a | 2.54 ± 0.35 a |
| 2nd harvest | |||||
| Conventional/full | CI | 26.70 ± 1.63 c | 74.59 ± 4.62 a | 22.71 ± 0.73 c | 2.85 ± 0.06 a |
| Conventional/deficit | CD | 29.66 ± 1.63 c | 58.73 ± 6.21 ab | 29.11 ± 0.17 a | 2.41 ± 0.17 b |
| Organic/full | OI | 43.50 ± 2.12 a | 69.78 ± 9.56 ab | 26.28 ± 0.94 b | 2.12 ± 0.05 b |
| Organic/deficit | OD | 35.83 ± 2.35 b | 48.67 ± 5.87 b | 26.62 ± 0.93 b | 2.38 ± 0.06 b |
| Treatment | Stomatal Conductance (cm/s) | Chlorophyll Fluorescence Fv/Fm | SPAD | |
|---|---|---|---|---|
| Cultivation/Irrigation Plan | 1st harvest | |||
| Conventional/full | CI | 0.791 ± 0.07 ab | 0.776 ± 0.011 a | 46.18 ± 0.99 a |
| Conventional/deficit | CD | 0.940 ± 0.07 a | 0.775 ± 0.005 a | 47.45 ± 1.77 ab |
| Organic/full | OI | 0.877 ± 0.07 ab | 0.775 ± 0.006 a | 39.93 ± 1.26 bc |
| Organic/deficit | OD | 0.684 ± 0.08 b | 0.788 ± 0.011 a | 35.40 ± 2.13 c |
| 2nd harvest | ||||
| Conventional/full | CI | 2.451 ± 0.231 a | 0.809 ± 0.001 b | 42.73 ± 1.79 b |
| Conventional/deficit | CD | 1.393 ± 0.256 bc | 0.819 ± 0.007 ab | 47.45 ± 0.82 a |
| Organic/full | OI | 1.788 ± 0.190 b | 0.810 ± 0.002 ab | 42.70 ± 1.11 b |
| Organic/deficit | OD | 0.984 ± 0.113 c | 0.823 ± 0.003 a | 45.11 ± 0.57 ab |
| Compound | Total Phenols | Total Flavonoids | ABTS | DPPH | FRAP | |
|---|---|---|---|---|---|---|
| 1st harvest | ||||||
| Conventional/Full | CI | 384.17 ± 45.34 a | 24.99 ± 1.63 a | 45.81 ± 5.06 a | 23.65 ± 4.41 c | 176.70 ± 30.76 a |
| Conventional/Deficit | CD | 399.45 ± 11.83 a | 24.30 ± 1.13 a | 36.19 ± 1.35 a | 43.69 ± 4.00 b | 150.49 ± 10.41 a |
| Organic/Full | OI | 414.56 ± 48.56 a | 27.96 ± 1.95 a | 35.44 ± 1.30 a | 26.87 ± 3.96 c | 169.15 ± 7.83 a |
| Organic/Deficit | OD | 444.72 ± 28.67 a | 29.18 ± 1.29 a | 37.83 ± 3.62 a | 60.11 ± 4.81 a | 200.36 ± 8.47 a |
| 2nd harvest | ||||||
| Conventional/Full | CI | 75.62 ± 2.34 b | 7.39 ± 0.20 b | 8.94 ± 0.30 b | 53.40 ± 6.74 c | 27.20 ± 1.61 b |
| Conventional/Deficit | CD | 75.36 ± 2.60 b | 7.15 ± 0.36 b | 8.20 ± 0.47 b | 62.87 ± 1.16 c | 28.12 ± 1.25 b |
| Organic/Full | OI | 95.22 ± 1.89 a | 11.15 ± 0.41 a | 10.77 ± 0.39 a | 82.01 ± 2.36 b | 43.54 ± 3.16 a |
| Organic/Deficit | OD | 90.69 ± 5.60 a | 9.88 ± 0.81 a | 10.61 ± 0.38 a | 102.83 ± 7.58 a | 37.87 ± 1.23 a |
| 1st Harvest | 2nd Harvest | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Compound | RI | Conv. FI | Conv. DI | Org. FI | Org. DI | Conv. FI | Conv. DI | Org. FI | Org. DI |
| α Pinene | 933 | 0.847 b | 0.944 ab | 0.999 a | 0.980 a | 0.899 a | 0.772 b | 0.856 a | 0.870 a |
| Camphene | 948 | 0.066 a | 0.073 a | 0.068 a | 0.067 a | 0.046 a | 0.041 c | 0.044 ab | 0.042 bc |
| Sabinene | 973 | 0.628 a | 0.681 b | 0.687 b | 0.675 b | 0.651 a | 0.580 b | 0.629 a | 0.605 b |
| β Pinene | 977 | 1.301 b | 1.376 ab | 1.390 a | 1.386 a | 1.201 a | 1.088 b | 1.204 a | 1.159 b |
| β Myrcene | 989 | 0.712 b | 0.741 ab | 0.767 a | 0.709 b | 0.727 a | 0.626 b | 0.634 b | 0.619 b |
| 3 Octanol | 1003 | 0.168 a | 0.133 b | 0.139 b | 0.102 c | 0.105 a | 0.098 ab | 0.102 a | 0.085 b |
| α Terpinene | 1005 | 0.044 a | 0.040 a | 0.042 a | 0.044 a | - | - | - | - |
| D-Limonene | 1028 | 11.687 b | 12.253 b | 14.603 a | 13.354 a | 15.140 c | 16.149 b | 15.670 c | 18.594 a |
| Eucalyptol | 1031 | 6.042 b | 6.869 a | 6.516 ab | 6.545 ab | 4.764 b | 4.626 b | 5.004 a | 4.517 b |
| β Ocimene | 1036 | 0.142 a | 0.142 a | 0.129 ab | 0.127 b | 0.062 a | 0.054 ab | 0.051 b | 0.056 b |
| trans β Ocimene | 1046 | 0.024 b | 0.059 a | 0.043 ab | 0.041 ab | 0.063 b | 0.069 b | 0.066 b | 0.082 a |
| γ Terpinene | 1058 | 0.102 a | 0.085 a | 0.094 a | 0.100 a | 0.044 a | 0.035 a | 0.036 a | 0.022 a |
| cis Sabinene hydrate | 1067 | 0.384 b | 0.365 ab | 0.333 b | 0.407 a | 0.171 a | 0.161 a | 0.145 a | 0.162 a |
| iso Menthone | 1164 | 0.069 b | 0.088 a | 0.078 ab | 0.069 b | 0.133 b | 0.144 a | 0.133 b | 0.149 a |
| Borneol | 1166 | 0.372 a | 0.391 a | 0.333 b | 0.313 b | 0.198 a | 0.184 a | 0.170 ab | 0.155 b |
| Menthol | 1175 | - | - | - | - | 0.156 c | 0.252 b | 0.185 c | 0.332 a |
| Terpinen 4 ol | 1178 | 0.315 a | 0.313 a | 0.295 ab | 0.247 b | 0.150 a | 0.114 ab | 0.125 a | 0.058 b |
| α Terpineol | 1191 | 0.226 b | 0.259 a | 0.251 a | 0.218 b | 0.219 a | 0.200 a | 0.211 a | 0.163 b |
| Dihydro carveol | 1993 | 0.511 b | 0.290 c | 0.292 c | 0.621 a | - | - | - | - |
| neo Dihydro carveol | 1995 | 0.460 b | 0.414 c | 0.409 c | 0.505 a | 0.290 a | 0.269 b | 0.294 a | 0.300 a |
| trans Carveol | 1219 | 0.495 a | 0.293 b | 0.236 b | 0.172 b | 0.024 a | 0.000 a | 0.000 a | 0.000 a |
| cis Carveol | 1231 | 0.569 b | 0.475 bc | 0.384 c | 0.764 a | - | - | - | - |
| Pulegone | 1240 | 0.525 ab | 0.535 a | 0.509 ab | 0.475 b | 0.818 d | 1.066 b | 0.868 c | 1.188 a |
| Carvone | 1244 | 70.290 a | 69.285 a | 67.626 b | 68.260 b | 71.582 a | 71.049 a | 71.051 a | 68.173 b |
| iso Dihydro carveol acetate | 1326 | 0.064 b | 0.040 bc | 0.027 c | 0.159 a | - | - | - | - |
| cis Carvyl acetate | 1361 | 0.100 b | 0.088 b | 0.059 b | 0.234 a | - | - | - | - |
| β Bourbonene | 1386 | 0.679 a | 0.757 a | 0.684 a | 0.677 a | 0.463 c | 0.511 b | 0.536 ab | 0.564 a |
| β Elemene | 1393 | 0.217 a | 0.221 a | 0.215 a | 0.184 a | 0.106 a | 0.074 b | 0.061 b | 0.078 b |
| β Caryophyllene | 1425 | 1.098 a | 1.061 a | 1.093 a | 1.057 a | 0.832 a | 0.749 a | 0.830 a | 0.827 a |
| Germacrene D | 1497 | 0.537 a | 0.439 a | 0.475 a | 0.388 a | - | - | - | - |
| Bicyclogermacrene | 1512 | 0.230 a | 0.207 a | 0.237 a | 0.178 a | 0.138 a | 0.069 b | 0.074 b | 0.068 b |
| Germacrene A | 1519 | 0.075 a | 0.048 a | 0.072 a | 0.054 a | - | - | - | - |
| trans Calamene | 1534 | 0.204 a | 0.218 a | 0.204 a | 0.206 a | 0.227 a | 0.231 a | 0.253 a | 0.249 a |
| Cubenol 1,10 di epi | 1617 | 0.124 a | 0.122 a | 0.103 a | 0.132 a | 0.086 a | 0.094 a | 0.095 a | 0.105 a |
| α Cadinol | 1657 | 0.136 a | 0.113 a | 0.099 a | 0.102 a | 0.049 a | 0.059 a | 0.039 a | 0.049 a |
| Mint sulfide | 1737 | - | - | - | - | 0.093 a | 0.064 a | 0.078 a | 0.063 a |
| Total Identified | 99.509 ab | 99.425 b | 99.495 ab | 99.498 a | 99.452 a | 99.424 a | 99.451 a | 99.340 b | |
| Monoterpenes hydrocarbons | 15.603 c | 16.395 bc | 18.822 a | 17.485 ab | 18.834 b | 19.406 b | 19.192 b | 22.056 a | |
| Sesquiterpenes hydrocarbons | 3.041 a | 2.952 a | 2.981 a | 2.747 a | 1.775 a | 1.634 a | 1.755 a | 1.783 a | |
| Oxygenated monoterpenes | 80.271 a | 79.579 a | 77.263 b | 78.600 ab | 78.508 a | 78.068 a | 78.188 a | 75.199 b | |
| Oxygenated sesquiterpenes | 0.260 a | 0.235 a | 0.203 a | 0.235 a | 0.135 a | 0.153 a | 0.134 a | 0.155 a | |
| Others | 0.332 b | 0.262 bc | 0.225 c | 0.496 a | 0.198 a | 0.162 ab | 0.181 ab | 0.148 b | |
| Essential Oil | LD50 | CL 95% | Slope ± SE | Intercept ± SE | x2 | p | |
|---|---|---|---|---|---|---|---|
| 1st harvest | |||||||
| Conventional/Full | CI | 52.11 | 41.23–61.75 | 0.03 ± 0.001 | −1.45 ± 0.09 | 147.20 | 0.000 |
| Conventional/Deficit | CD | 53.29 | 44.76–61.90 | 0.03 ± 0.001 | −1.70 ± 0.83 | 159.16 | 0.000 |
| Organic/Full | OI | 50.84 | 42.45–59.02 | 0.03 ± 0.001 | −1.67 ± 0.08 | 151.53 | 0.000 |
| Organic/Deficit | OD | 57.72 | 52.96–62.40 | 0.03 ± 0.001 | −1.73 ± 0.08 | 53.78 | 0.000 |
| 2nd harvest | |||||||
| Conventional/Full | CI | 51.26 | 39.73–61.61 | 0.03 ± 0.001 | −1.44 ± 0.08 | 225.08 | 0.000 |
| Conventional/Deficit | CD | 47.03 | 33.49–53.59 | 0.03 ± 0.001 | −1.35 ± 0.08 | 244.57 | 0.000 |
| Organic/Full | OI | 51.96 | 42.00–61.86 | 0.04 ± 0.001 | −1.86 ± 0.09 | 235.51 | 0.000 |
| Organic/Deficit | OD | 51.43 | 44.99–57.36 | 0.03 ± 0.001 | −1.46 ± 0.08 | 79.50 | 0.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chrysargyris, A.; Koutsoumpeli, E.; Xylia, P.; Fytrou, A.; Konstantopoulou, M.; Tzortzakis, N. Organic Cultivation and Deficit Irrigation Practices to Improve Chemical and Biological Activity of Mentha spicata Plants. Agronomy 2021, 11, 599. https://doi.org/10.3390/agronomy11030599
Chrysargyris A, Koutsoumpeli E, Xylia P, Fytrou A, Konstantopoulou M, Tzortzakis N. Organic Cultivation and Deficit Irrigation Practices to Improve Chemical and Biological Activity of Mentha spicata Plants. Agronomy. 2021; 11(3):599. https://doi.org/10.3390/agronomy11030599
Chicago/Turabian StyleChrysargyris, Antonios, Eleni Koutsoumpeli, Panayiota Xylia, Anastasia Fytrou, Maria Konstantopoulou, and Nikolaos Tzortzakis. 2021. "Organic Cultivation and Deficit Irrigation Practices to Improve Chemical and Biological Activity of Mentha spicata Plants" Agronomy 11, no. 3: 599. https://doi.org/10.3390/agronomy11030599
APA StyleChrysargyris, A., Koutsoumpeli, E., Xylia, P., Fytrou, A., Konstantopoulou, M., & Tzortzakis, N. (2021). Organic Cultivation and Deficit Irrigation Practices to Improve Chemical and Biological Activity of Mentha spicata Plants. Agronomy, 11(3), 599. https://doi.org/10.3390/agronomy11030599