
Performance of Hydroponically Cultivated Geranium and Common Verbena under Salinity and High Electrical Conductivity Levels


 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
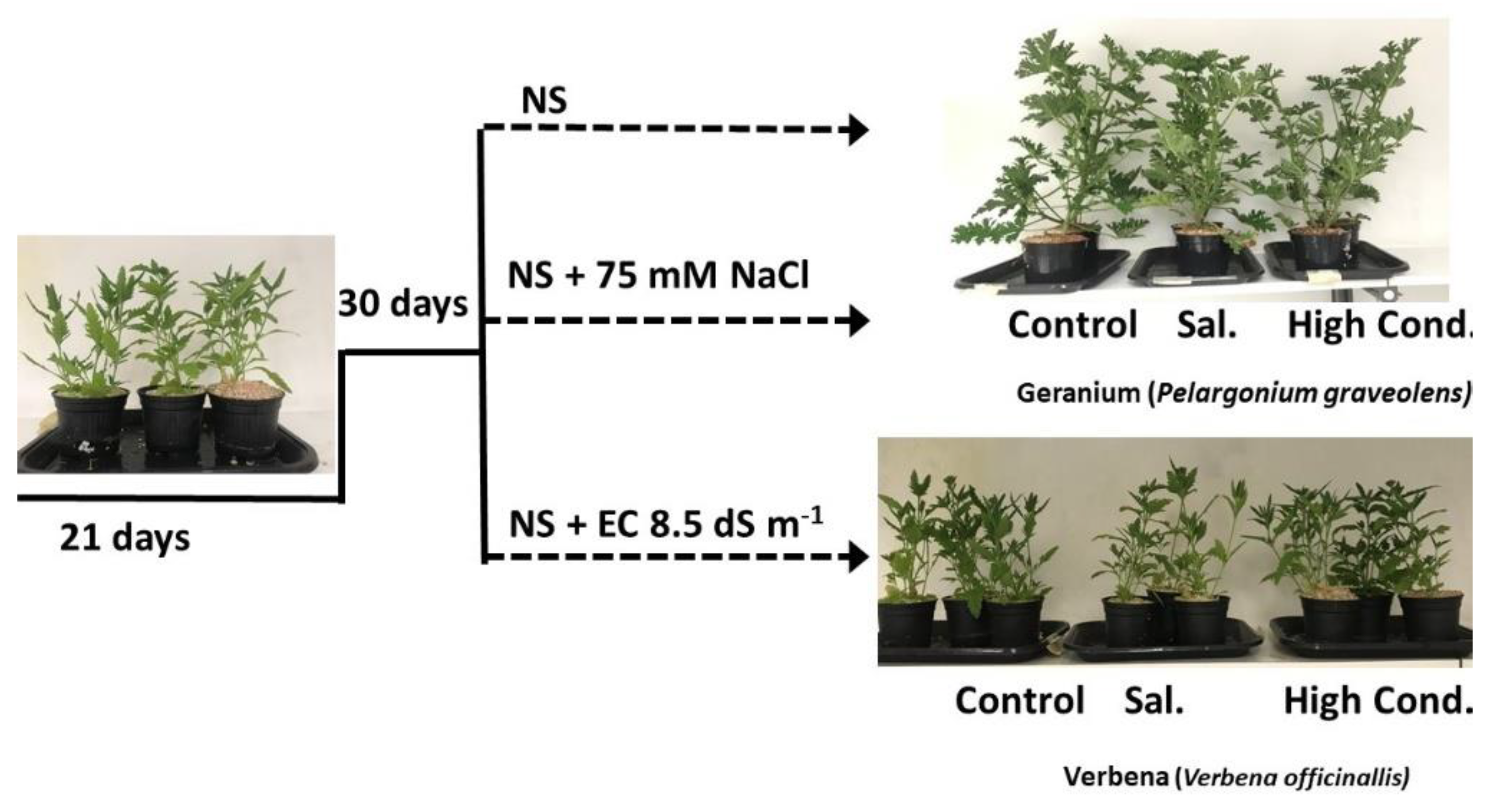
2.1. Experimental Setup and Plant Material
2.2. Plant Growth and Physiological Measurements
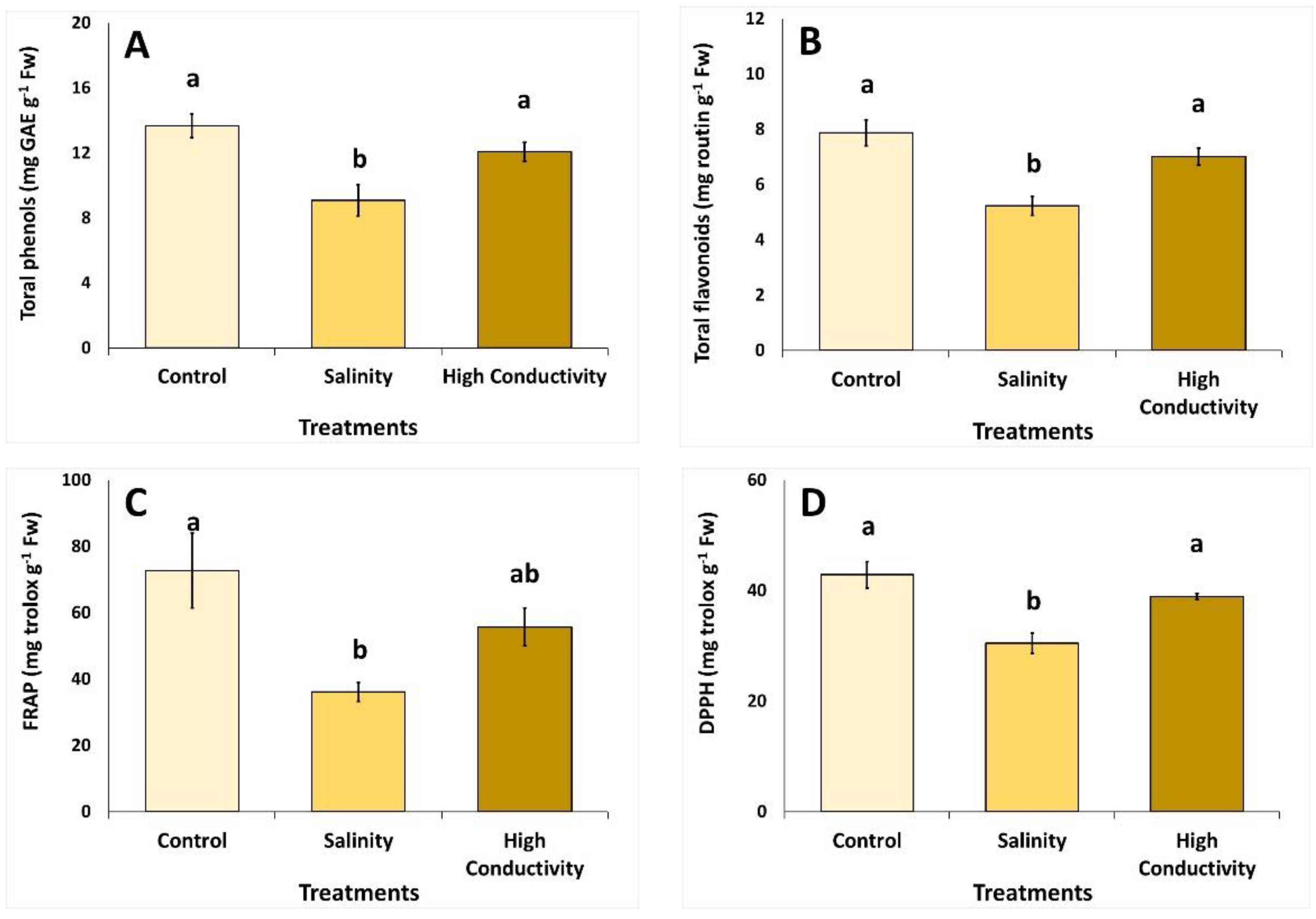
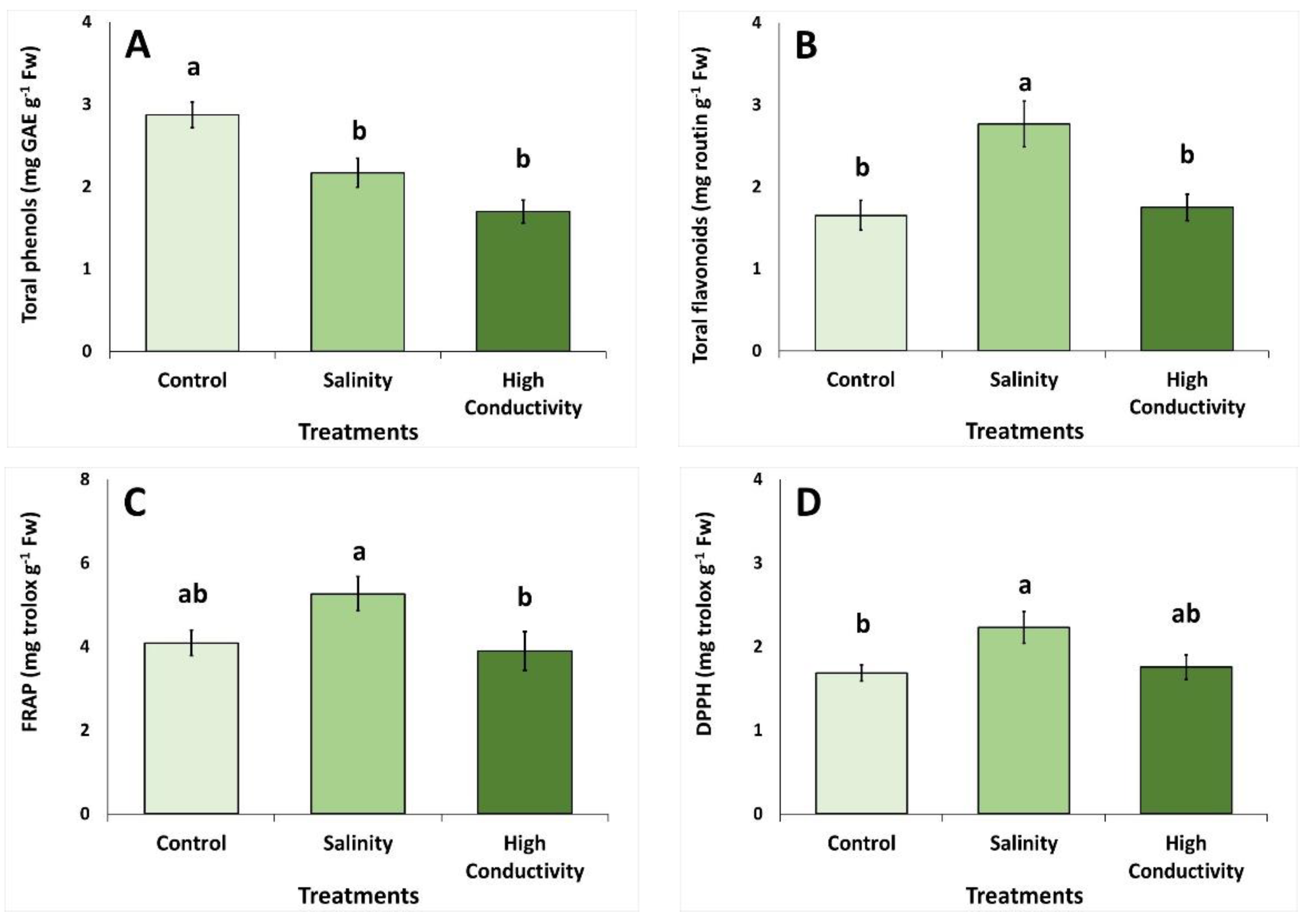
2.3. Antioxidant Activity, Total Phenolic and Total Flavonoid Content
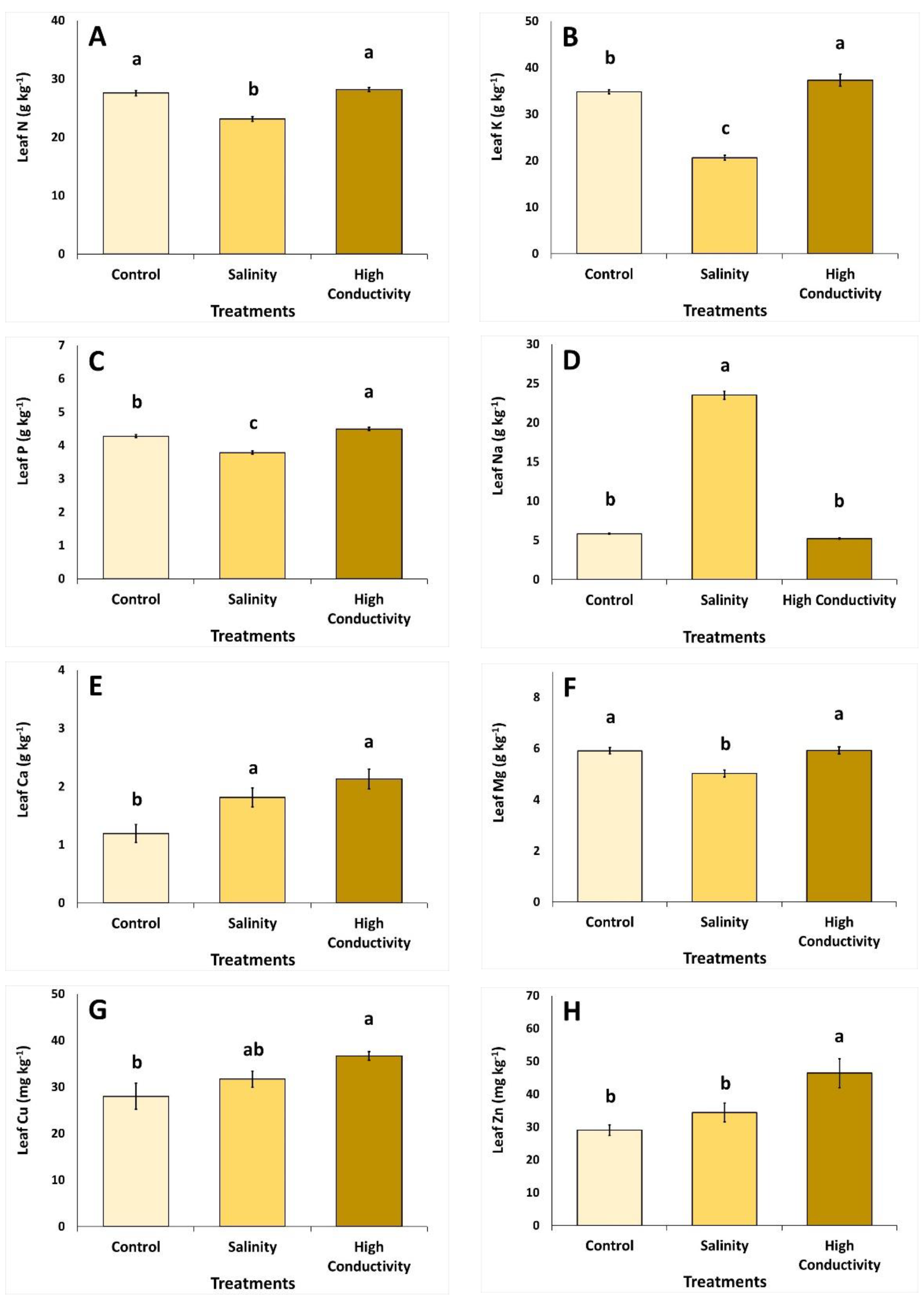
2.4. Mineral Content
2.5. Hydrogen Peroxide, Lipid Peroxidation, Antioxidative Enzyme Activities and Proline Content
2.6. Statistical Analysis
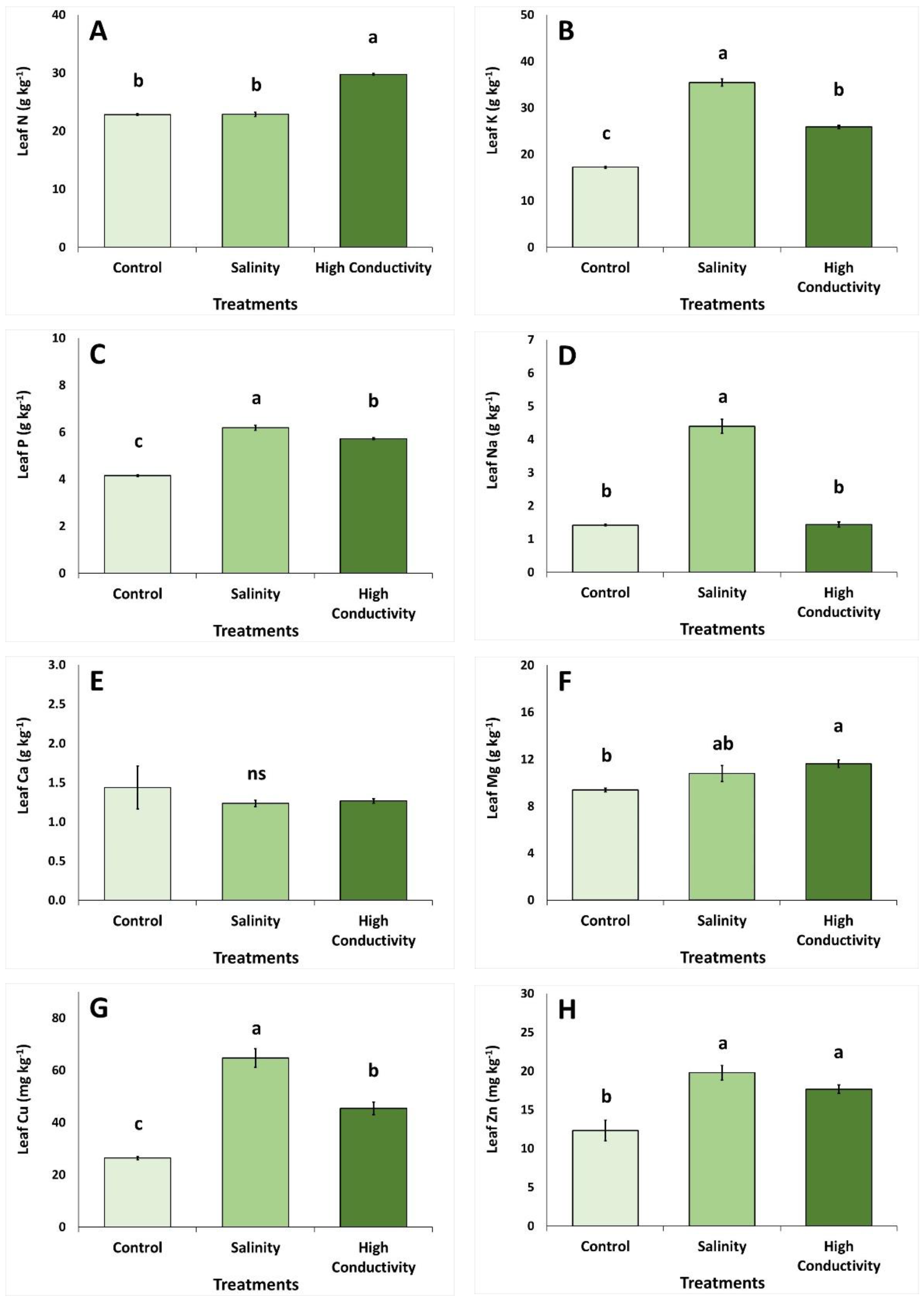
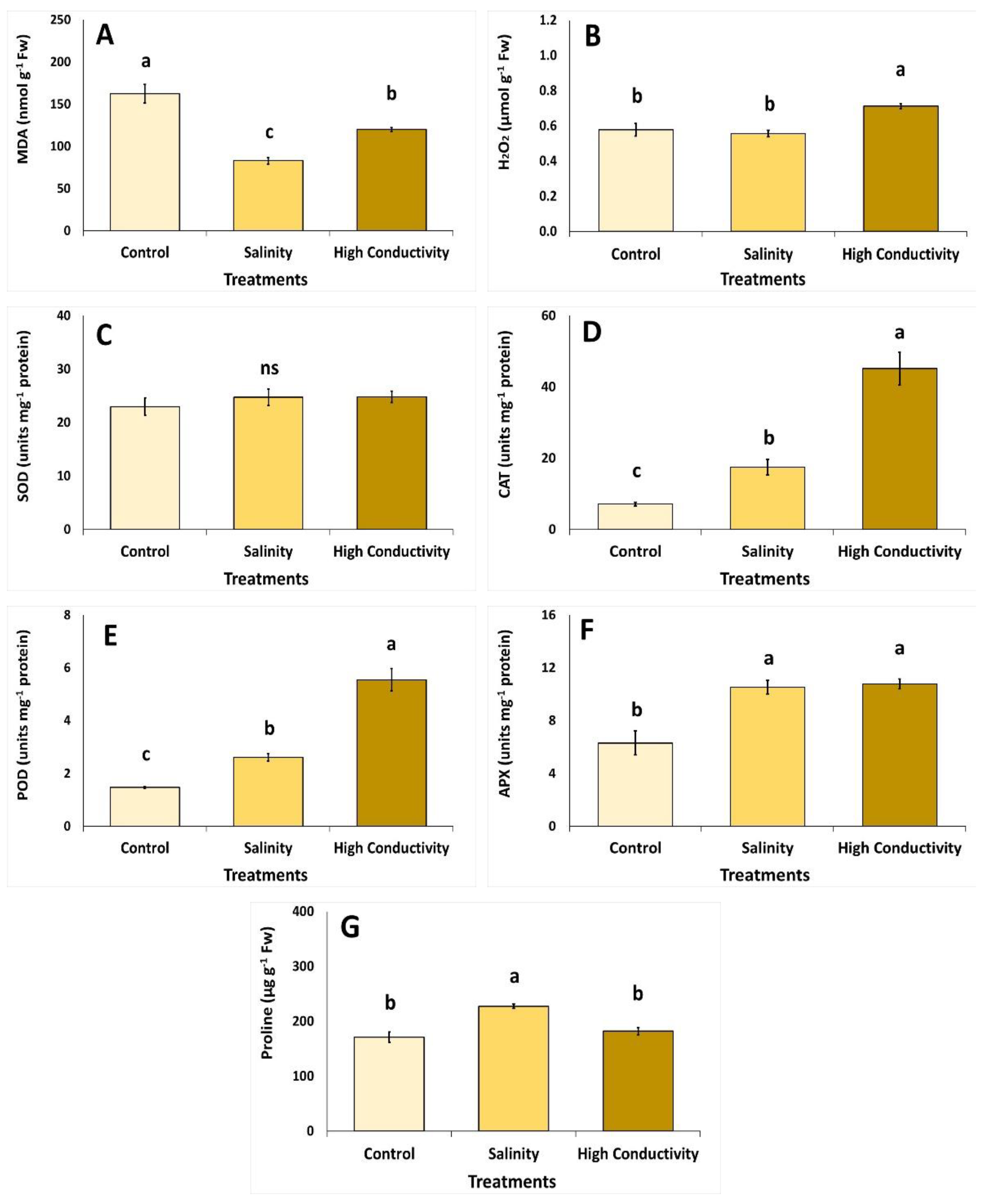
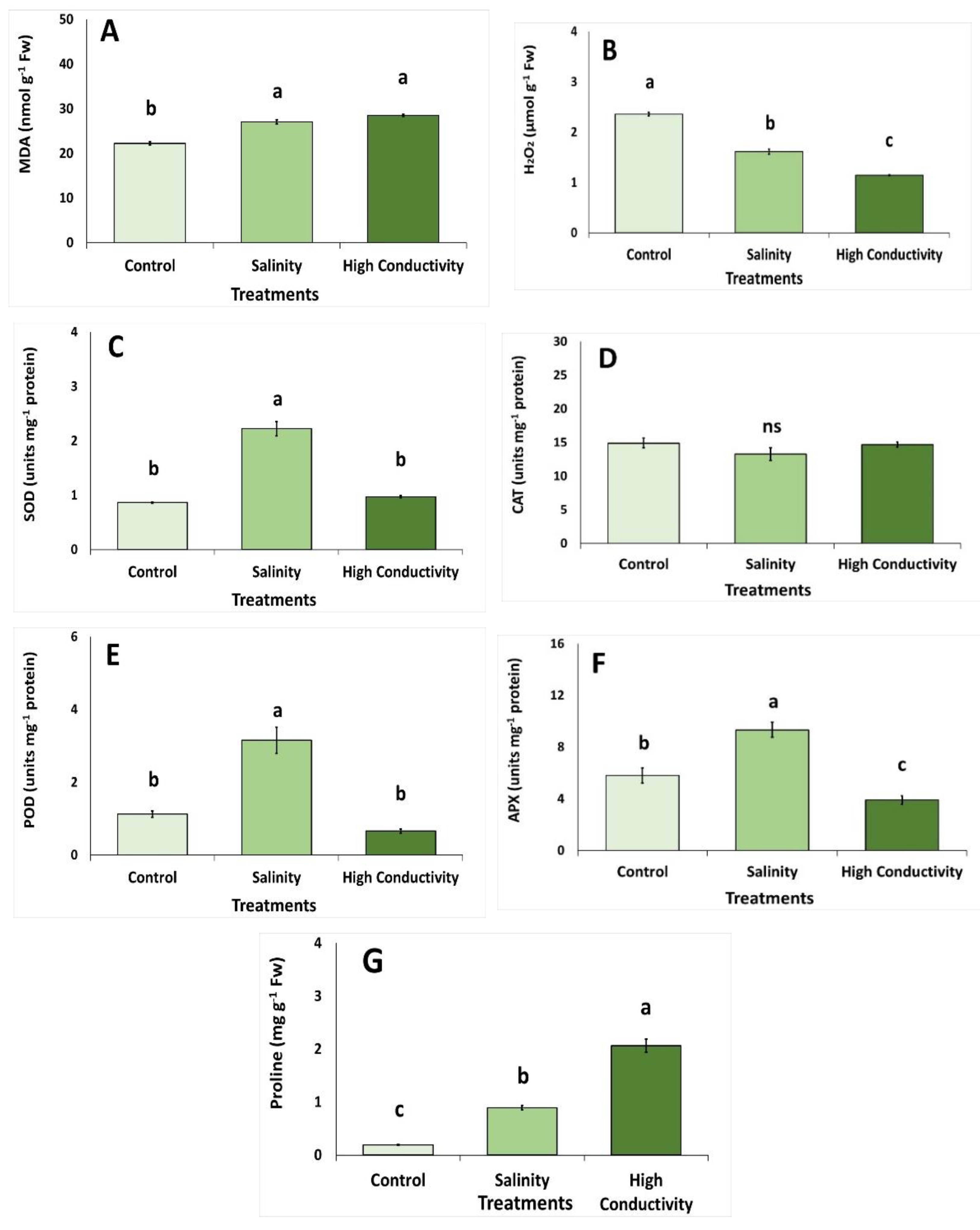
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Niu, G.; Rodriguez, D.S. Relative salt tolerance of five herbaceous perennials. HortScience 2006, 41, 1493–1497. [Google Scholar] [CrossRef]
- Guo, H.; Hu, Z.; Zhang, H.; Min, W.; Hou, Z. Comparative Effects of Salt and Alkali Stress on Antioxidant System in Cotton (Gossypium Hirsutum L.) Leaves. Open Chem. 2019, 17, 1352–1360. [Google Scholar] [CrossRef]
- Tzortzakis, N.G. Influence of NaCl and calcium nitrate on lettuce and endive growth using nutrient film technique. Int. J. Veg. Sci. 2009, 15, 44–56. [Google Scholar] [CrossRef]
- Koubouris, G.C.; Tzortzakis, N.; Kourgialas, N.N.; Darioti, M.; Metzidakis, I. Growth, photosynthesis and pollen performance in saline water treated olive plants under high temperature. Int. J. Plant Biol. 2015, 6, 6. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Solomou, M.; Petropoulos, S.A.; Tzortzakis, N. Physiological and biochemical attributes of Mentha spicata when subjected to saline conditions and cation foliar application. J. Plant Physiol. 2019, 232, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.Q.; Ma, B.L.; Ren, C.Z. Growth, gas exchange, chlorophyll fluorescence, and ion content of naked oat in response to salinity. Crop. Sci. 2007, 47, 123–131. [Google Scholar] [CrossRef]
- Giordano, M.; Petropoulos, S.A.; Cirillo, C.; Rouphael, Y. Biochemical, Physiological, and Molecular Aspects of Ornamental Plants Adaptation to Deficit Irrigation. Horticulturae 2021, 7, 107. [Google Scholar] [CrossRef]
- Giordano, M.; Petropoulos, S.A.; Rouphael, Y. Response and Defence Mechanisms of Vegetable Crops against Drought, Heat and Salinity Stress. Agriculture 2021, 11, 463. [Google Scholar] [CrossRef]
- Kalantari, E.; Hassanli, A.; Ghanbarian, G.A.; Ghaemi, A.A.; Mousavi, S.R. Local desalination treatment plant wastewater reuse and evaluation potential absorption of salts by the halophyte plants. Eurasian J. Soil Sci. 2018, 7, 43–50. [Google Scholar] [CrossRef][Green Version]
- Kelepesi, S.; Tzortzakis, N.G. Olive mill wastes: A growing medium component for seedling and crop production of lettuce and chicory. Int. J. Veg. Sci. 2009, 15, 325–339. [Google Scholar] [CrossRef]
- Razmjoo, K.; Heydarizadeh, P.; Sabzalian, M.R. Effect of salinity and drought stresses on growth parameters and essential oil content of Matricaria chamomila. Int. J. Agric. Biol. 2008, 10, 451–454. [Google Scholar]
- Rozema, J. Growth, water and ion relationships of halophytic monocotyledonae and dicotyledonae; a unified concept. Aquat. Bot. 1991, 39, 17–33. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Tzortzakis, N.; Nicola, S.; Savvas, D.; Voogt, W. Editorial: Soilless Cultivation Through an Intensive Crop Production Scheme. Management Strategies, Challenges and Future Directions. Front. Plant Sci. 2020, 11, 10–12. [Google Scholar] [CrossRef]
- Grattan, S.; Grieve, C.M. Salinity-mineral nutrient relations in horticultural crops. Sci. Hortic. 1999, 78, 127–157. [Google Scholar] [CrossRef]
- Moya, C.; Oyanedel, E.; Verdugo, G.; Flores, M.F.; Urrestarazu, M.; Álvaro, J.E. Increased electrical conductivity in nutrient solution management enhances dietary and organoleptic qualities in soilless culture tomato. HortScience 2017, 52, 868–872. [Google Scholar] [CrossRef]
- Dorais, M.; Papadopoulos, A.P.; Gosselin, A. Influence of electric conductivity management on greenhouse tomato yield and fruit quality. Agronomie 2001, 21, 367–383. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Loupasaki, S.; Petropoulos, S.A.; Tzortzakis, N. Salinity and cation foliar application: Implications on essential oil yield and composition of hydroponically grown spearmint plants. Sci. Hortic. 2019, 256, 108581. [Google Scholar] [CrossRef]
- Taârit, M.B.; Msaada, K.; Hosni, K.; Marzouk, B. Fatty acids, phenolic changes and antioxidant activity of clary sage (Salvia sclarea L.) rosette leaves grown under saline conditions. Ind. Crops Prod. 2012, 38, 58–63. [Google Scholar] [CrossRef]
- Parejo, I.; Viladomat, F.; Bastida, J.; Rosas-Romero, A.; Flerlage, N.; Burillo, J.; Codina, C. Comparison between the radical scavenging activity and antioxidant activity of six distilled and nondistilled mediterranean herbs and aromatic plants. J. Agric. Food Chem. 2002, 50, 6882–6890. [Google Scholar] [CrossRef]
- Xylia, P.; Clark, A.; Chrysargyris, A.; Romanazzi, G.; Tzortzakis, N. Quality and safety attributes on shredded carrots by using Origanum majorana and ascorbic acid. Postharvest Biol. Technol. 2019, 155, 120–129. [Google Scholar] [CrossRef]
- Hendawy, S.F.; Khalid, K.A. Response of Sage (Salvia officinalis L.) Plants to Zinc Application Under Different Salinity Levels. J. Appl. Sci. Res. 2014, 1, 147–155. [Google Scholar]
- Salachna, P.; Piechocki, R. Effects of sodium chloride on growth and mineral nutrition of Purpletop vervain. J. Ecol. Eng. 2016, 17, 148–152. [Google Scholar] [CrossRef]
- Fiz, O.; Vargas, P.; Alarcón, M.; Aedo, C.; García, J.L.; Aldasoro, J.J. Phylogeny and historical biogeography of Geraniaceae in relation to climate changes and pollination ecology. Syst. Bot. 2008, 33, 326–342. [Google Scholar] [CrossRef]
- Colling, J.; Groenewald, J.H.; Makunga, N.P. Genetic alterations for increased coumarin production lead to metabolic changes in the medicinally important Pelargonium sidoides DC (Geraniaceae). Metab. Eng. 2010, 12, 561–572. [Google Scholar] [CrossRef]
- Pandey, J.; Chand, S.; Chaurasiya, S.; Kumari, R.; Patra, D.D.; Verma, R.K.; Singh, S. Effect of tannery sludge amendments on the activity of soil enzymes and phytoremediation potential of two economically important cultivars of geranium (Pelargonium graveolens). Soil Sediment Contam. 2019, 28, 395–410. [Google Scholar] [CrossRef]
- Khan, A.W.; Khan, A.U.; Ahmed, T. Anticonvulsant, anxiolytic, and sedative activities of Verbena officinalis. Front. Pharmacol. 2016, 7, 499. [Google Scholar] [CrossRef]
- Batista, M.C.; Nascimento, R.; Júnior, S.D.O.M. Physiology and production of cherry tomato cultivars in a hydroponic system using brackish water. Braz. J. Agric. Environ. Eng. 2021, 25, 219–227. [Google Scholar]
- Irakoze, W.; Prodjinoto, H.; Nijimbere, S.; Bizimana, J.B.; Bigirimana, J.; Rufyikiri, G.; Lutts, S. NaCl- and Na2SO4-Induced Salinity Differentially Affect Clay Soil Chemical Properties and Yield Components of Two Rice Cultivars (Oryza sativa L.) in Burundi. Agronomy 2021, 11, 571. [Google Scholar] [CrossRef]
- Tzortzakis, N.G.; Economakis, C.D. Shredded maize stems as an alternative substrate medium: Effect on growth, flowering and yield of tomato in soilless culture. J. Veg. Sci. 2005, 11, 57–70. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Papakyriakou, E.; Petropoulos, S.A.; Tzortzakis, N. The combined and single effect of salinity and copper stress on growth and quality of Mentha spicata plants. J. Hazard. Mater. 2019, 368, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Chrysargyris, A.; Nikou, A.; Tzortzakis, N. Effectiveness of Aloe vera gel coating for maintaining tomato fruit quality. N. Zeal. J. Crop. Hortic. Sci. 2016, 44, 203–217. [Google Scholar] [CrossRef]
- Wojdyło, A.; Oszmiański, J.; Czemerys, R. Antioxidant activity and phenolic compounds in 32 selected herbs. Food Chem. 2007, 105, 940–949. [Google Scholar] [CrossRef]
- Tzortzakis, N.G.; Tzanakaki, K.; Economakis, C.D. Effect of origanum oil and vinegar on the maintenance of postharvest quality of tomato. Food Nutr. Sci. 2011, 02, 974–982. [Google Scholar] [CrossRef][Green Version]
- Meyers, K.J.; Watkins, C.B.; Pritts, M.P.; Liu, R.H. Antioxidant and Antiproliferative Activities of Strawberries. J. Agric. Food Chem. 2003, 51, 6887–6892. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Prasad, M.; Kavanagh, A.; Tzortzakis, N. Biochar type, ratio, and nutrient levels in growing media affects seedling production and plant performance. Agronomy 2020, 10, 1421. [Google Scholar] [CrossRef]
- Loreto, F.; Velikova, V. Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products, and reduces lipid peroxidation of cellular membranes. Plant Physiol. 2001, 127, 1781–1787. [Google Scholar] [CrossRef]
- de Azevedo Neto, A.D.; Prisco, J.T.; Enéas-Filho, J.; de Abreu, C.E.B.; Gomes-Filho, E. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot. 2006, 56, 87–94. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Xylia, P.; Botsaris, G.; Tzortzakis, N. Antioxidant and antibacterial activities, mineral and essential oil composition of spearmint (Mentha spicata L.) affected by the potassium levels. Ind. Crops Prod. 2017, 103, 202–212. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Charalambous, S.; Xylia, P.; Litskas, V.; Stavrinides, M.; Tzortzakis, N. Assessing the biostimulant effects of a novel plant-based formulation on tomato crop. Sustainability 2020, 12, 8432. [Google Scholar] [CrossRef]
- Khedr, A.H.A.; Abbas, M.A.; Abdel Wahid, A.A.; Quick, W.P.; Abogadallah, G.M. Proline induces the expression of salt-stress-responsive proteins and may improve the adaptation of Pancratium maritimum L. to salt-stress. J. Exp. Bot. 2003, 54, 2553–2562. [Google Scholar] [CrossRef]
- Economakis, C. Effect of solution conductivity of growth and yield of Origanum dictamnus L. in nutrient film culture. Acta Hortic. 1992, 306, 204–209. [Google Scholar] [CrossRef]
- Ciriello, M.; Pannico, A.; El-Nakhel, C.; Formisano, L.; Cristofano, F.; Duri, L.G.; Pizzolongo, F.; Romano, R.; De Pascale, S.; Colla, G.; et al. Sweet basil functional quality as shaped by genotype and macronutrient concentration reciprocal action. Plants 2020, 9, 1786. [Google Scholar] [CrossRef]
- Wu, M.; Kubota, C. Effects of electrical conductivity of hydroponic nutrient solution on leaf gas exchange of five greenhouse tomato cultivars. Horttechnology 2008, 18, 271–277. [Google Scholar] [CrossRef]
- Romero-Aranda, R.; Soria, T.; Cuartero, J. Tomato plant-water uptake and plant-water relationships under saline growth conditions. Plant Sci. 2001, 160, 265–272. [Google Scholar] [CrossRef]
- Mohammadkhani, N.; Heidari, R. Water stress induced stomatal closure in two maize cultivars. Res. J. Biol. Sci. 2008, 3, 750–754. [Google Scholar]
- Sirault, X.R.R.; James, R.A.; Furbank, R.T. A new screening method for osmotic component of salinity tolerance in cereals using infrared thermography. Funct. Plant Biol. 2009, 36, 970–977. [Google Scholar] [CrossRef]
- Sucre, B.; Suárez, N. Effect of salinity and PEG-induced water stress on water status, gas exchange, solute accumulation, and leaf growth in Ipomoea pes-caprae. Environ. Exp. Bot. 2011, 70, 192–203. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Tzionis, A.; Xylia, P.; Tzortzakis, N. Effects of salinity on tagetes growth, physiology, and shelf life of edible flowers stored in passive modified atmosphere packaging or treated with ethanol. Front. Plant Sci. 2018, 871, 1–13. [Google Scholar] [CrossRef]
- Dorta-Santos, M.A.; Barriola, I.; Wassner, D.F.; Ploschuk, E.L. Photosynthesis, fluorescence and mesophyll conductance responses to increasing salinity levels in Jatropha curcas at early vegetative stages. J. Agron. Crop. Sci. 2020, 206, 52–63. [Google Scholar] [CrossRef]
- Xie, Z.X.; Duan, L.S.; Li, Z.H.; Wang, X.D.; Liu, X. Dose-Dependent Effects of Coronatine on Cotton Seedling Growth Under Salt Stress. J. Plant Growth Regul. 2015, 34, 651–664. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Diaz-Vivancos, P.; Álvarez, S.; Fernández-García, N.; Sánchez-Blanco, M.J.; Hernández, J.A. NaCl-induced physiological and biochemical adaptative mechanisms in the ornamental Myrtus communis L. plants. J. Plant Physiol. 2015, 183, 41–51. [Google Scholar] [CrossRef]
- Albornoz, F.; Heinrich Lieth, J. Over fertilization limits lettuce productivity because of osmotic stress. Chil. J. Agric. Res. 2015, 75, 284–290. [Google Scholar] [CrossRef]
- Naczk, M.; Shahidi, F. Extraction and analysis of phenolics in food. J. Chromatogr. A 2004, 1054, 95–111. [Google Scholar] [CrossRef]
- Krauss, S.; Schnitzler, W.H.; Grassmann, J.; Woitke, M. The influence of different electrical conductivity values in a simplified recirculating soilless system on inner and outer fruit quality characteristics of tomato. J. Agric. Food Chem. 2006, 54, 441–448. [Google Scholar] [CrossRef]
- Ali, H.M.Z.; Ismail, G.S.M.; Mohamed, H.E.; Ismail, G.S.M. Tomato fruit quality as influenced by salinity and nitric oxide. Turk. J. Bot. 2014, 38, 122–129. [Google Scholar] [CrossRef]
- Dewir, Y.H.; Chakrabarty, D.; Ali, M.B.; Hahn, E.J.; Paek, K.Y. Effects of hydroponic solution EC, substrates, PPF and nutrient scheduling on growth and photosynthetic competence during acclimatization of micropropagated Spathiphyllum plantlets. Plant Growth Regul. 2005, 46, 241–251. [Google Scholar] [CrossRef]
- Toderich, K.; Shuyskaya, E.; Rakhmankulova, Z.; Bukarev, R.; Khujanazarov, T.; Zhapaev, R.; Ismail, S.; Gupta, S.K.; Yamanaka, N.; Boboev, F. Threshold tolerance of new genotypes of Pennisetum glaucum (L.) R. Br. to salinity and drought. Agronomy 2018, 8, 230. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Azarmi, F.; Mozafari, V.; Abbaszadeh Dahaji, P.; Hamidpour, M. Biochemical, physiological and antioxidant enzymatic activity responses of pistachio seedlings treated with plant growth promoting rhizobacteria and Zn to salinity stress. Acta Physiol. Plant 2016, 38, 1–16. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Diaz-Vivancos, P.; Álvarez, S.; Fernández-García, N.; Sanchez-Blanco, M.J.; Hernández, J.A. Physiological and biochemical mechanisms of the ornamental Eugenia myrtifolia L. plants for coping with NaCl stress and recovery. Planta 2015, 242, 829–846. [Google Scholar] [CrossRef]
- Martínez, J.P.; Kinet, J.M.; Bajji, M.; Lutts, S. NaCl alleviates polyethylene glycol-induced water stress in the halophyte species Atriplex halimus L. J. Exp. Bot. 2005, 56, 2421–2431. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Geranium | Verbena | |||||
|---|---|---|---|---|---|---|
| Control (2.1 dS m−1) | Salinity (75 mM NaCl) (8.5 dS m−1) | High Conductivity (8.5 dS m−1) | Control (2.1 dS m−1) | Salinity (75 mM NaCl) (8.5 dS m−1) | High Conductivity (8.5 dS m−1) | |
| Height (cm) | 48.11 ± 1.52 a | 39.38 ± 1.36 b | 48.10 ± 3.56 a | 16.41 ± 0.18 b | 17.40 ± 0.18 a | 15.63 ± 0.29 b |
| Leaf No | 38.00 ± 2.42 a | 36.00 ± 0.44 a | 40.66 ± 6.16 a | 56.00 ± 1.65 a | 42.66 ± 4.75 b | 65.50 ± 0.22 a |
| Biomass fresh weight (g) | 62.02 ± 2.47 a | 48.70 ± 3.37 b | 56.92 ± 5.24 ab | 4.63 ± 0.33 a | 2.86 ± 0.24 b | 5.14 ± 0.29 a |
| Biomass dry weight (g) | 8.10 ± 0.61 a | 7.42 ± 0.49 a | 9.26 ± 1.52 a | 0.85 ± 0.05 a | 0.50 ± 0.05 b | 0.94 ± 0.08 a |
| Geranium | Verbena | |||||
|---|---|---|---|---|---|---|
| Control (2.1 dS m−1) | Salinity (75 mM NaCl) (8.5 dS m−1) | High Conductivity (8.5 dS m−1) | Control (2.1 dS m−1) | Salinity (75 mM NaCl) (8.5 dS m−1) | High Conductivity (8.5 dS m−1) | |
| Stomatal resistance (cm s−1) | 1.16 ± 0.36 c | 10.80 ± 0.87 a | 4.87 ± 0.74 b | 2.61 ± 0.31 c | 11.90 ± 1.55 a | 8.59 ± 0.75 b |
| Chlorophyll fluorescence (Fv/Fm) | 0.83 ± 0.00 a | 0.78 ± 0.01 b | 0.79 ± 0.00 b | 0.81 ± 0.01 a | 0.76 ± 0.00 b | 0.75 ± 0.01 b |
| SPAD | 110.80 ± 1.72 a | 59.86 ± 3.84 c | 84.20 ± 8.17 b | 43.17 ± 3.06 a | 25.47 ± 1.71 b | 28.92 ± 2.62 b |
| Chlorophyll a (mg g−1 Fw) | 0.85 ± 0.06 a | 0.80 ± 0.04 a | 0.86 ± 0.05 a | 1.15 ± 0.16 ab | 0.84 ± 0.12 b | 1.44 ± 0.13 a |
| Chlorophyll b (mg g−1 Fw) | 0.28 ± 0.02 a | 0.25 ± 0.03 a | 0.28 ± 0.02 a | 0.34 ± 0.04 a | 0.30 ± 0.03 a | 0.37 ± 0.02 a |
| Total Chlorophylls (mg g−1 Fw) | 1.13 ± 0.08 a | 1.05 ± 0.07 a | 1.14 ± 0.07 a | 1.49 ± 0.21 ab | 1.14 ± 0.11 b | 1.81 ± 0.15 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chrysargyris, A.; Petropoulos, S.A.; Prvulovic, D.; Tzortzakis, N. Performance of Hydroponically Cultivated Geranium and Common Verbena under Salinity and High Electrical Conductivity Levels. Agronomy 2021, 11, 1237. https://doi.org/10.3390/agronomy11061237
Chrysargyris A, Petropoulos SA, Prvulovic D, Tzortzakis N. Performance of Hydroponically Cultivated Geranium and Common Verbena under Salinity and High Electrical Conductivity Levels. Agronomy. 2021; 11(6):1237. https://doi.org/10.3390/agronomy11061237
Chicago/Turabian StyleChrysargyris, Antonios, Spyridon A. Petropoulos, Dejan Prvulovic, and Nikolaos Tzortzakis. 2021. "Performance of Hydroponically Cultivated Geranium and Common Verbena under Salinity and High Electrical Conductivity Levels" Agronomy 11, no. 6: 1237. https://doi.org/10.3390/agronomy11061237
APA StyleChrysargyris, A., Petropoulos, S. A., Prvulovic, D., & Tzortzakis, N. (2021). Performance of Hydroponically Cultivated Geranium and Common Verbena under Salinity and High Electrical Conductivity Levels. Agronomy, 11(6), 1237. https://doi.org/10.3390/agronomy11061237