
Genetic Purity of Cacao Criollo from Honduras Is Revealed by SSR Molecular Markers

 , , , , , and
, , , , , and 
Abstract
1. Introduction
2. Materials and Methods
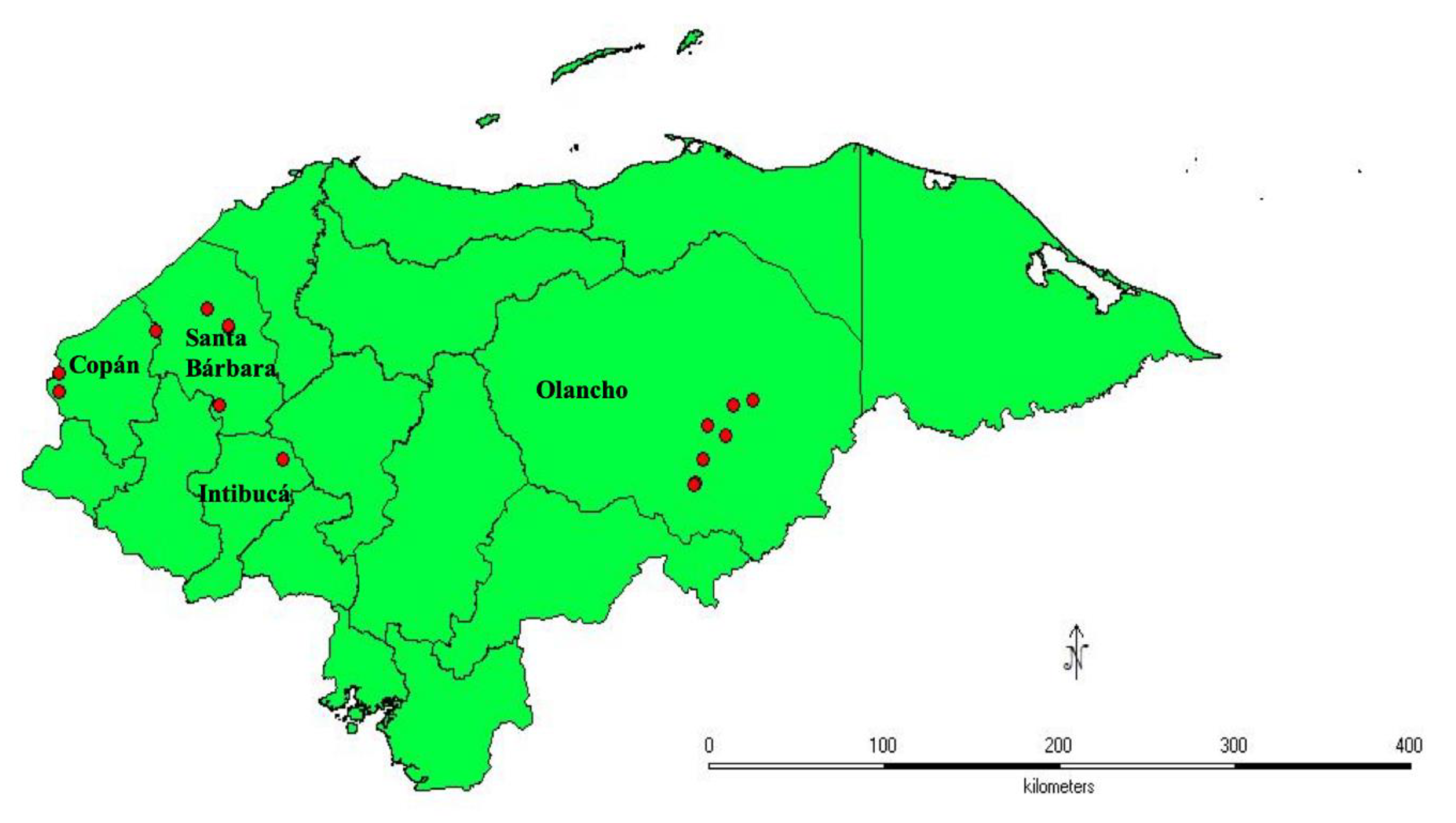
2.1. Plant Material
2.2. DNA Extraction and Quantification
2.3. SSR Markers
2.4. PCR Amplification and Electrophoresis
2.5. Data Analysis
2.6. Cluster Analysis
3. Results
3.1. Descriptive Genetic Parameters of the SSR Loci
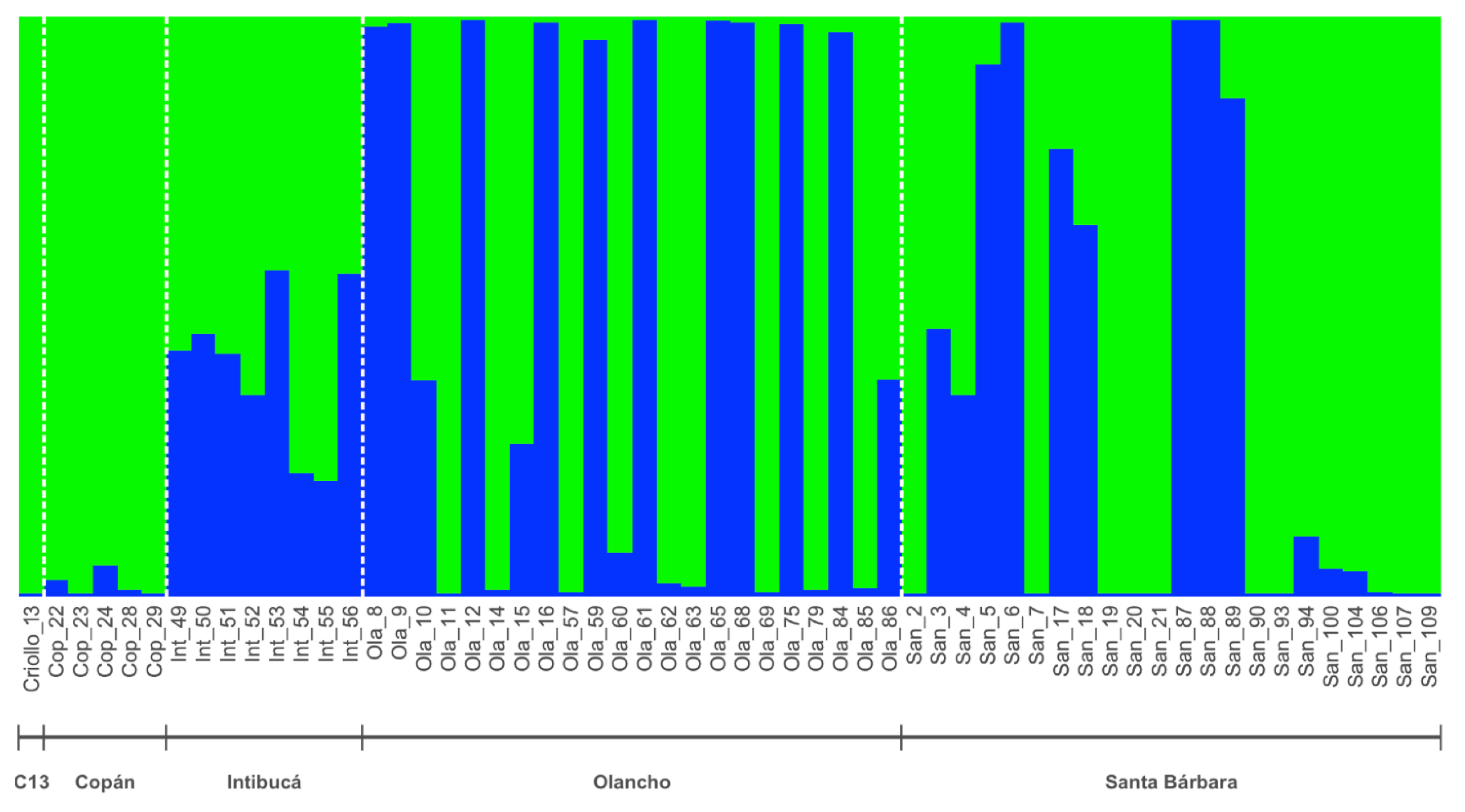
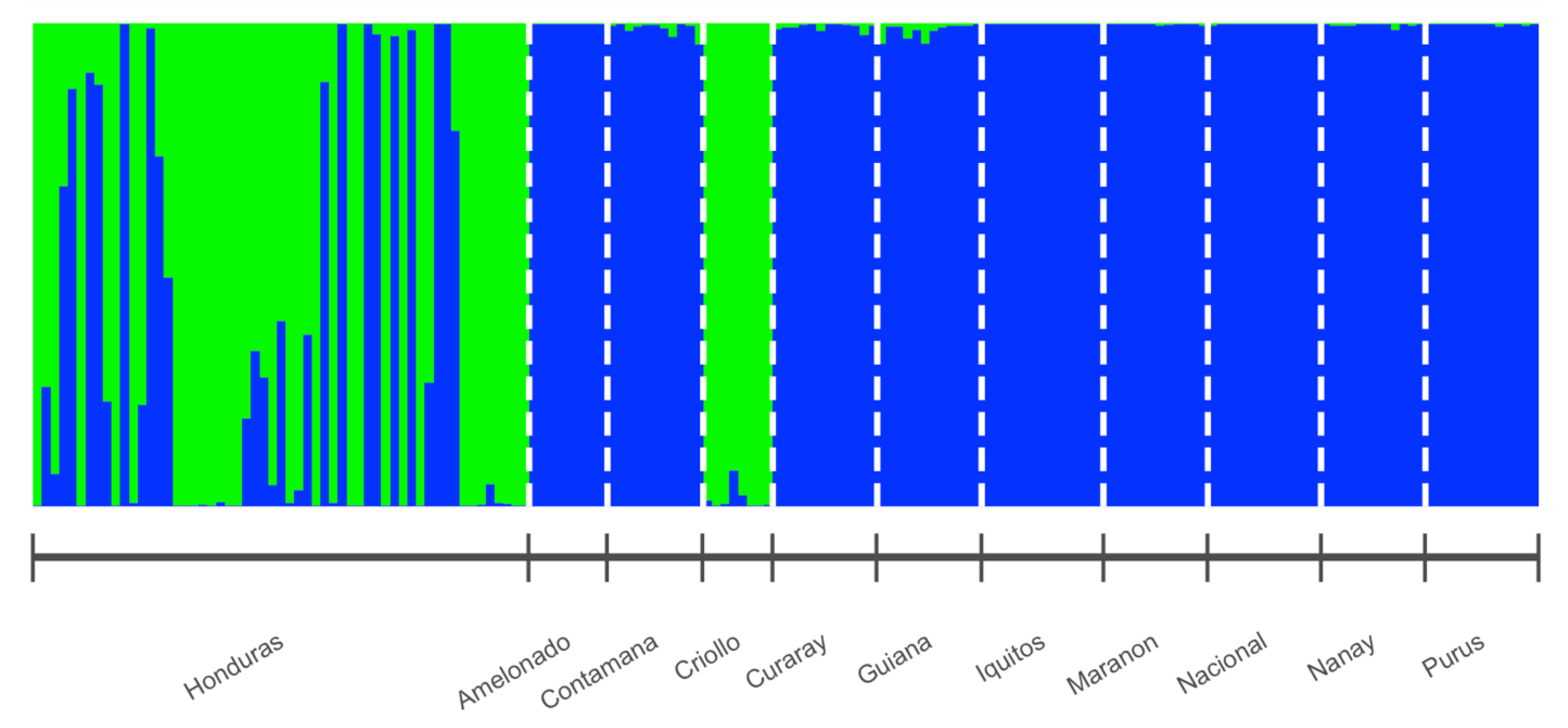
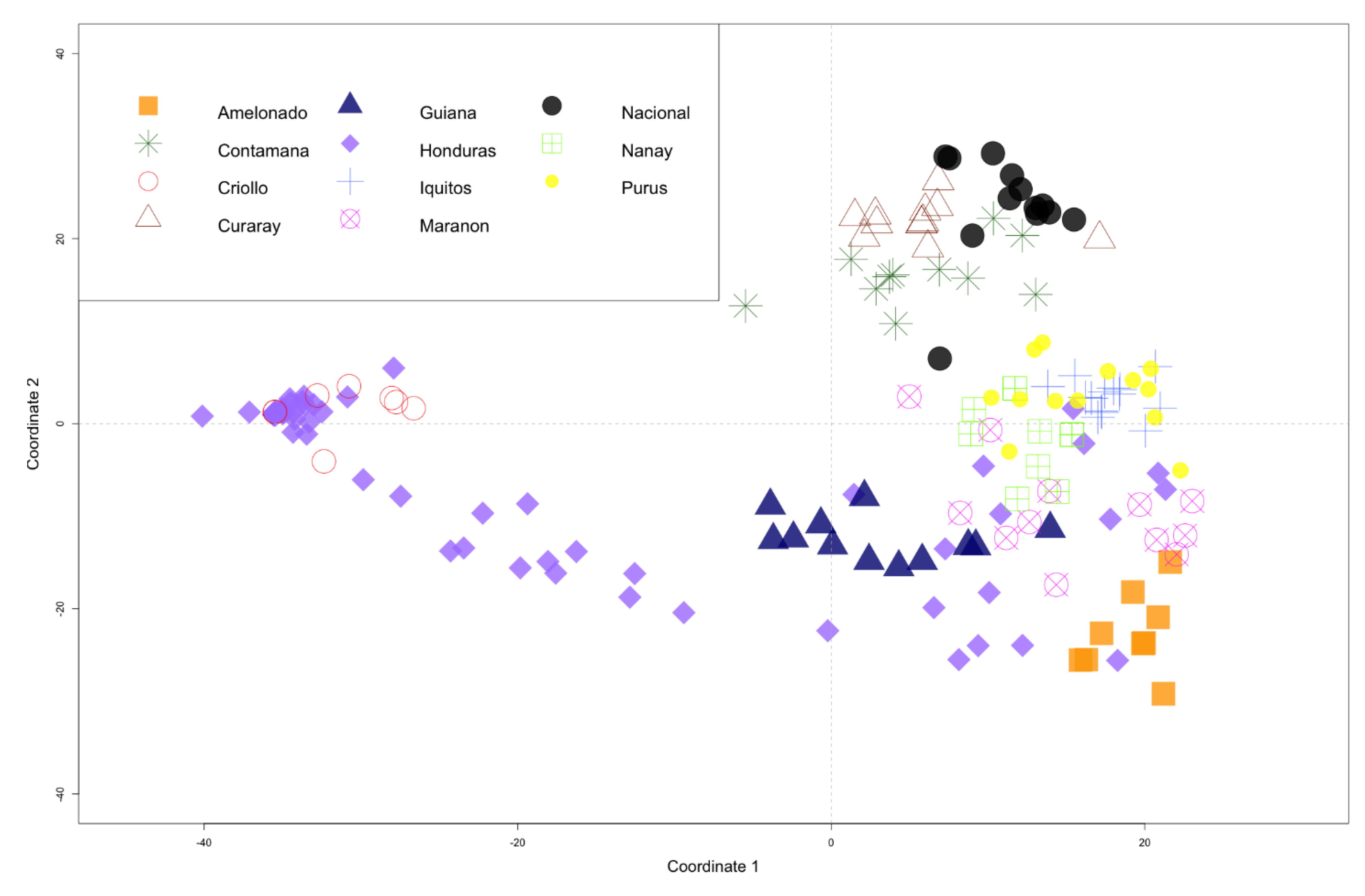
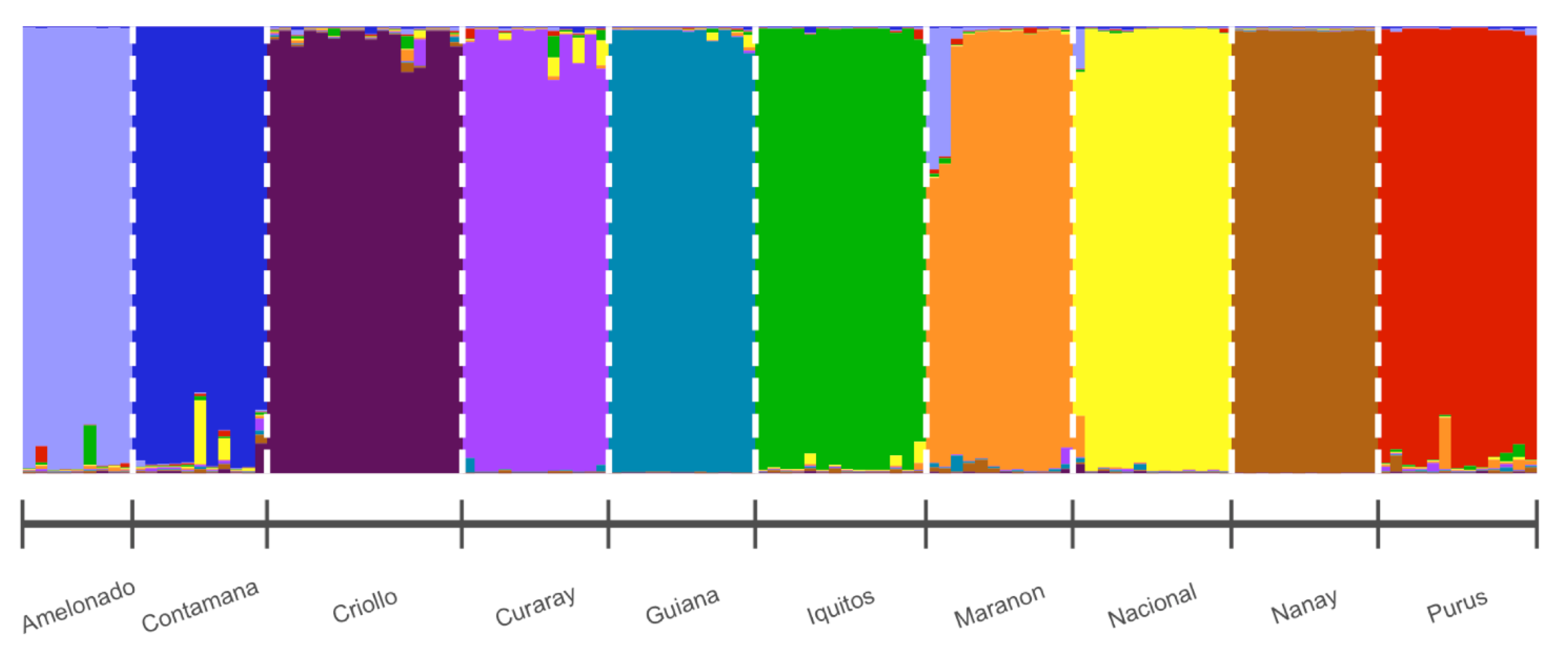
3.2. Cluster Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheesman, E. Notes on the Nomenclature, Classification and Possible Relationships of Cacao Populations; IPC Science and Technology Press: Guildford, UK, 1944. [Google Scholar]
- Bartley, B.G. The Genetic Diversity of Cacao and Its Utilization; CABI: Oxfordshire, UK, 2005; ISBN 1-84593-024-X. [Google Scholar]
- Flood, J.; Murphy, R. Cocoa Futures: A Source Book of Some Important Issues Facing the Cocoa Industry; Federación de Cafetaleros de Colombia: Bogotá, Colombia; CABI: London, UK, 2004; ISBN 958-97441-1-7. [Google Scholar]
- Cuatrecasas, J. Cacao and Its Allies: A Taxonomic Revision of the Genus Theobroma; Smithsonian Institution: Washington, DC, USA, 1964; Volume 35. [Google Scholar]
- Henderson, J.S.; Joyce, R.A.; Hall, G.R.; Hurst, W.J.; McGovern, P.E. Chemical and archaeological evidence for the earliest cacao beverages. Proc. Natl. Acad. Sci. USA 2007, 104, 18937–18940. [Google Scholar] [CrossRef] [PubMed]
- Powis, T.G.; Cyphers, A.; Gaikwad, N.W.; Grivetti, L.; Cheong, K. Cacao use and the San Lorenzo Olmec. Proc. Natl. Acad. Sci. USA 2011, 108, 8595–8600. [Google Scholar] [CrossRef] [PubMed]
- Ji, K.; Zhang, D.; Motilal, L.A.; Boccara, M.; Lachenaud, P.; Meinhardt, L.W. Genetic diversity and parentage in farmer varieties of cacao (Theobroma cacao L.) from Honduras and Nicaragua as revealed by single nucleotide polymorphism (SNP) markers. Genet. Resour. Crop Evol. 2013, 60, 441–453. [Google Scholar] [CrossRef]
- Crown, P.L.; Hurst, W.J. Evidence of cacao use in the Prehispanic American Southwest. Proc. Natl. Acad. Sci. USA 2009, 106, 2110–2113. [Google Scholar] [CrossRef]
- Wood, G.; Lass, R. Cocoa Tropical Agriculture; Wiley-Blackwell: Hoboken, NJ, USA, 1985. [Google Scholar]
- Zarrillo, S.; Gaikwad, N.; Lanaud, C.; Powis, T.; Viot, C.; Lesur, I.; Fouet, O.; Argout, X.; Guichoux, E.; Salin, F. The use and domestication of Theobroma cacao during the mid-Holocene in the upper Amazon. Nat. Ecol. Evol. 2018, 2, 1879–1888. [Google Scholar] [CrossRef]
- Coe, S.D.; Coe, M.D. The True History of Chocolate; Thames & Hudson: London, UK, 2013; ISBN 0-500-77093-X. [Google Scholar]
- Bergmann, J.F. The distribution of cacao cultivation in pre-Columbian America. Ann. Assoc. Am. Geogr. 1969, 59, 85–96. [Google Scholar] [CrossRef]
- Cornejo, O.E.; Yee, M.-C.; Dominguez, V.; Andrews, M.; Sockell, A.; Strandberg, E.; Livingstone, D.; Stack, C.; Romero, A.; Umaharan, P. Population genomic analyses of the chocolate tree, Theobroma cacao L., provide insights into its domestication process. Commun. Biol. 2018, 1, 1–12. [Google Scholar] [CrossRef]
- Bekele, F.; Bekele, I. A sampling of the phenetic diversity of cacao in the International Cocoa Gene Bank of Trinidad. Crop Sci. 1996, 36, 57–64. [Google Scholar] [CrossRef]
- Argout, X.; Salse, J.; Aury, J.-M.; Guiltinan, M.J.; Droc, G.; Gouzy, J.; Allegre, M.; Chaparro, C.; Legavre, T.; Maximova, S.N.; et al. The genome of Theobroma cacao. Nat. Genet. 2011, 43, 101–108. [Google Scholar] [CrossRef]
- Lachenaud, P.; Motamayor, J.C. The Criollo cacao tree (Theobroma cacao L.): A review. Genet. Resour. Crop Evol. 2017, 64, 1807–1820. [Google Scholar] [CrossRef]
- Motamayor, J.C.; Lachenaud, P.; Mota, J.W.d.S.e.; Loor, R.; Kuhn, D.N.; Brown, J.S.; Schnell, R.J. Geographic and genetic population differentiation of the Amazonian chocolate tree (Theobroma cacao L.). PLoS ONE 2008, 3, e3311. [Google Scholar] [CrossRef]
- Pound, F. A note on the cocoa population of South America. Rep. Proc. Cocoa Res. Conf. 1945, 1, 131–133. [Google Scholar]
- N’Goran, J.A.; Laurent, V.; Risterucci, A.-M.; Lanaud, C. Comparative genetic diversity studies of Theobroma cacao L. using RFLP and RAPD markers. Heredity 1994, 73, 589–597. [Google Scholar] [CrossRef]
- Lerceteau, E.; Robert, T.; Pétiard, V.; Crouzillat, D. Evaluation of the extent of genetic variability among Theobroma cacao accessions using RAPD and RFLP markers. Theor. Appl. Genet. 1997, 95, 10–19. [Google Scholar] [CrossRef]
- Whitkus, R.; De la Cruz, M.; Mota-Bravo, L.; Gómez-Pompa, A. Genetic diversity and relationships of cacao (Theobroma cacao L.) in southern Mexico. Theor. Appl. Genet. 1998, 96, 621–627. [Google Scholar] [CrossRef]
- Sounigo, O.; Umaharan, R.; Christopher, Y.; Sankar, A.; Ramdahin, S. Assessing the genetic diversity in the International Cocoa Genebank, Trinidad (ICG, T) using isozyme electrophoresis and RAPD. Genet. Resour. Crop Evol. 2005, 52, 1111–1120. [Google Scholar] [CrossRef]
- dos Santos, R.C.; Pires, J.L.; Lopes, U.V.; Gramacho, K.P.G.; Flores, A.B.; Bahia, R.d.C.S.; Ramos, H.C.C.; Corrêa, R.X.; Ahnert, D. Assessment of genetic diversity on a sample of cocoa accessions resistant to witches’ broom disease based on RAPD and pedigree data. Bragantia 2005, 64, 361–368. [Google Scholar] [CrossRef]
- Motamayor, J.C.; Risterucci, A.-M.; Lopez, P.A.; Ortiz, C.F.; Moreno, A.; Lanaud, C. Cacao domestication I: The origin of the cacao cultivated by the Mayas. Heredity 2002, 89, 380–386. [Google Scholar] [CrossRef]
- Zhang, D.; Mischke, S.; Johnson, E.S.; Phillips-Mora, W.; Meinhardt, L. Molecular characterization of an international cacao collection using microsatellite markers. Tree Genet. Genomes 2009, 5, 1–10. [Google Scholar] [CrossRef]
- Zhang, D.; Boccara, M.; Motilal, L.; Mischke, S.; Johnson, E.S.; Butler, D.R.; Bailey, B.; Meinhardt, L. Molecular characterization of an earliest cacao (Theobroma cacao L.) collection from Upper Amazon using microsatellite DNA markers. Tree Genet. Genomes 2009, 5, 595–607. [Google Scholar] [CrossRef]
- Zhang, D.; Martínez, W.J.; Johnson, E.S.; Somarriba, E.; Phillips-Mora, W.; Astorga, C.; Mischke, S.; Meinhardt, L.W. Genetic diversity and spatial structure in a new distinct Theobroma cacao L. population in Bolivia. Genet. Resour. Crop Evol. 2012, 59, 239–252. [Google Scholar] [CrossRef]
- Stack, J.C.; Royaert, S.; Gutiérrez, O.; Nagai, C.; Holanda, I.S.A.; Schnell, R.; Motamayor, J.-C. Assessing microsatellite linkage disequilibrium in wild, cultivated, and mapping populations of Theobroma cacao L. and its impact on association mapping. Tree Genet. Genomes 2015, 11, 19. [Google Scholar] [CrossRef]
- Motilal, L.A.; Zhang, D.; Mischke, S.; Meinhardt, L.W.; Umaharan, P. Microsatellite-aided detection of genetic redundancy improves management of the International Cocoa Genebank, Trinidad. Tree Genet. Genomes 2013, 9, 1395–1411. [Google Scholar] [CrossRef]
- Everaert, H.; Rottiers, H.; Pham, P.H.D.; Ha, L.T.V.; Nguyen, T.P.D.; Tran, P.D.; De Wever, J.; Maebe, K.; Smagghe, G.; Dewettinck, K. Molecular characterization of Vietnamese cocoa genotypes (Theobroma cacao L.) using microsatellite markers. Tree Genet. Genomes 2017, 13, 99. [Google Scholar] [CrossRef]
- Everaert, H.; De Wever, J.; Tang, T.K.H.; Vu, T.L.A.; Maebe, K.; Rottiers, H.; Lefever, S.; Smagghe, G.; Dewettinck, K.; Messens, K. Genetic classification of Vietnamese cacao cultivars assessed by SNP and SSR markers. Tree Genet. Genomes 2020, 16, 43. [Google Scholar] [CrossRef]
- Guiraud, B.S.H.B.; Tahi, M.G.; Fouet, O.; Trebissou, C.I.; Pokou, D.; Rivallan, R.; Argout, X.; Koffi, K.K.; Koné, B.; Zoro, B.I.A.; et al. Assessment of genetic diversity and structure in cocoa trees (Theobroma cacao L.) in Côte d’Ivoire with reference to their susceptibility to Cocoa swollen shoot virus disease (CSSVD). Tree Genet. Genomes 2018, 14, 52. [Google Scholar] [CrossRef]
- Lindo, A.A.; Robinson, D.E.; Tennant, P.F.; Meinhardt, L.W.; Zhang, D. Molecular characterization of cacao (Theobroma cacao) germplasm from Jamaica using single nucleotide polymorphism (SNP) markers. Trop. Plant Biol. 2018, 11, 93–106. [Google Scholar] [CrossRef]
- Adenet, S.; Regina, F.; Rogers, D.; Bharath, S.; Argout, X.; Rochefort, K.; Cilas, C. Study of the genetic diversity of cocoa populations (Theobroma cacao L.) of Martinique (FWI) and potential for processing and the cocoa industry. Genet. Resour. Crop Evol. 2020, 67, 1–11. [Google Scholar] [CrossRef]
- Arevalo-Gardini, E.; Meinhardt, L.W.; Zuñiga, L.C.; Arévalo-Gardni, J.; Motilal, L.; Zhang, D. Genetic identity and origin of “Piura Porcelana”—A fine-flavored traditional variety of cacao (Theoborma cacao) from the Peruvian Amazon. Tree Genet. Genomes 2019, 15, 11. [Google Scholar] [CrossRef]
- Cosme, S.; Cuevas, H.; Zhang, D.; Oleksyk, T.; Irish, B. Genetic diversity of naturalized cacao (Theobroma cacao L.) in Puerto Rico. Tree Genet. Genomes 2016, 12, 88. [Google Scholar] [CrossRef]
- Mata-Quirós, A.; Arciniegas-Leal, A.; Phillips-Mora, W.; Meinhardt, L.W.; Motilal, L.; Mischke, S.; Zhang, D. Assessing hidden parentage and genetic integrity of the “United Fruit Clones” of cacao (Theobroma cacao) from Costa Rica using SNP markers. Breed. Sci. 2018, 18057. [Google Scholar] [CrossRef] [PubMed]
- Olasupo, F.O.; Adewale, D.B.; Aikpokpodion, P.O.; Muyiwa, A.A.; Bhattacharjee, R.; Gutierrez, O.A.; Motamayor, J.C.; Schnell, R.J.; Ebai, S.; Zhang, D. Genetic identity and diversity of Nigerian cacao genebank collections verified by single nucleotide polymorphisms (SNPs): A guide to field genebank management and utilization. Tree Genet. Genomes 2018, 14, 32. [Google Scholar] [CrossRef]
- Motilal, L.A.; Zhang, D.; Umaharan, P.; Mischke, S.; Mooleedhar, V.; Meinhardt, L.W. The relic Criollo cacao in Belize–genetic diversity and relationship with Trinitario and other cacao clones held in the International Cocoa Genebank, Trinidad. Plant Genet. Resour. Charact. Util. 2010, 8, 106. [Google Scholar] [CrossRef]
- Durán, E.; Dubón, A. Tipos Genéticos de Cacao y Distribución Geográfica en Honduras; Centro de Comunicación Agrícola: Lima, Honduras, 2016. [Google Scholar]
- Murray, M.G.; Pitas, J.W. Plant DNA from alcohol-preserved samples. Plant Mol. Biol. Report. 1996, 14, 261–265. [Google Scholar] [CrossRef]
- Bressan, E.A.; Rossi, M.L.; Gerald, L.T.; Figueira, A. Extraction of high-quality DNA from ethanol-preserved tropical plant tissues. BMC Res. Notes 2014, 7, 268. [Google Scholar] [CrossRef] [PubMed]
- Saunders, J.A.; Mischke, S.; Leamy, E.A.; Hemeida, A.A. Selection of international molecular standards for DNA fingerprinting of Theobroma cacao. Theor. Appl. Genet. 2004, 110, 41–47. [Google Scholar] [CrossRef]
- Pugh, T.; Fouet, O.; Risterucci, A.-M.; Brottier, P.; Abouladze, M.; Deletrez, C.; Courtois, B.; Clément, D.; Larmande, P.; N’Goran, J.A. A new cacao linkage map based on codominant markers: Development and integration of 201 new microsatellite markers. Theor. Appl. Genet. 2004, 108, 1151–1161. [Google Scholar] [CrossRef]
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol. Ecol. 2007, 16, 1099–1106. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Liu, K.; Muse, S.V. PowerMarker: An Integrated Analysis Environment for Genetic Marker Analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A. Structure harvester: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Francis, R.M. Pophelper: An R package and web app to analyse and visualize population structure. Mol. Ecol. Resour. 2017, 17, 27–32. [Google Scholar] [CrossRef]
- Jombart, T. Adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef]
- Chessel, D.; Dufour, A.B.; Thioulouse, J. The ade4 package-I-One-table methods. R News 2004, 4, 5–10. [Google Scholar]
- Motamayor, J.C.; Risterucci, A.-M.; Heath, M.; Lanaud, C. Cacao domestication II: Progenitor germplasm of the Trinitario cacao cultivar. Heredity 2003, 91, 322–330. [Google Scholar] [CrossRef]
- Boza, E.J.; Motamayor, J.C.; Amores, F.M.; Cedeño-Amador, S.; Tondo, C.L.; Livingstone, D.S.; Schnell, R.J.; Gutiérrez, O.A. Genetic Characterization of the Cacao Cultivar CCN 51: Its Impact and Significance on Global Cacao Improvement and Production. J. Am. Soc. Hortic. Sci. 2014, 139, 219–229. [Google Scholar] [CrossRef]
- De la Cruz, M.; Whitkus, R.; Gómez-Pompa, A.; Mota-Bravo, L. Origins of cacao cultivation. Nature 1995, 375, 542–543. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SSR Name | EMBL Accession | Chromosome No. | Forward Primer | Reverse Primer | Size (bp) | Motif | Ref. |
|---|---|---|---|---|---|---|---|
| MTcCIR6 | Y16980 | 6 | 5′-TTCCCTCTAAACTACCCT AAAT-3′ | 5′-TAAAGCAAAGCAAT CTAACATA-3′ | 224–246 | (TG)7(GA)13 | [43] |
| MTcCIR7 | Y16981 | 7 | 5′-ATGCGAATGACAACTGG T-3′ | 5′-GCTTTCAGTCCTTTG CTT-3′ | 147–159 | (GA)11 | [43] |
| MTcCIR8 | Y16982 | 9 | 5′-CTAGTTTCCCATTTACCA-3′ | 5′-TCCTCAGCATTTTCTT TC-3′ | 286–302 | (TC)5TT(TC)17TTT(CT)4 | [43] |
| MTcCIR11 | Y16985 | 2 | 5′-TTTGGTGATTATTAGCA G-3′ | 5′-GATTCGATTTGATG TGAG-3′ | 290–316 | (TC)13 | [43] |
| MTcCIR12 | Y16986 | 4 | 5′-TCTGACCCCAAACCTGT A-3′ | 5′-ATTCCAGTTAAAGC ACAT-3′ | 187–219 | (CATA)4N18(TG)6 | [43] |
| MTcCIR18 | Y16991 | 4 | 5′-GATAGCTAAGGGGATTG AGGA-3′ | 5′-GGTAATTCAATCATT TGAGGATA-3′ | 331–345 | (GA)12 | [43] |
| MTcCIR22 | Y16995 | 1 | 5′-ATTCTCGCAAAAACTTA G-3′ | 5′-GATGGAAGGAGTGT AAATAG-3′ | 277–289 | (TC)12N146(CT)10 | [43] |
| MTcCIR24 | Y16996 | 9 | 5′-TTTGGGGTGATTTCTTC TGA-3′ | 5′-TCTGTCTCGTCTTTTG GTGA-3′ | 184–200 | (AG)13 | [43] |
| MTcCIR26 | Y16998 | 8 | 5′-GCATTCATCAATACAT TC-3′ | 5′-GCACTCAAAGTTCA TACTAC-3′ | 289–307 | (TC)9C(CT)4TT(CT)11 | [43] |
| MTcCIR33 | AJ271826 | 4 | 5′-TGGGTTGAAGATTTG GT-3′ | 5′-CAACAATGAAAATA GGCA-3′ | 270–344 | (TG)11 | [43] |
| MTcCIR37 | AJ271942 | 10 | 5′-CTGGGTGCTGATAGAT AA-3′ | 5′-AATACCCTCCACAC AAAT-3′ | 144–184 | (GT)15 | [43] |
| MTcCIR40 | AJ271943 | 3 | 5′-AATCCGACAGTCTTTAA TC-3′ | 5′-CCTAGGCCAGAGAA TTGA-3′ | 271–285 | (AC)15 | [43] |
| MTcCIR60 | AJ271958 | 2 | 5′-CGCTACTAACAAACATCA AA-3′ | 5′-AGAGCAACCATCAC TAATCA-3′ | 188–212 | (CT)7(CA)20 | [43] |
| MTcCIR115 | AJ566457 | 4 | 5′-GTGATTCAAATTCAAATA TG-3′ | 5′-AATAGCAAGAGAGT GATGAG-3′ | 189–195 | (TC)11 | [44] |
| MTcCIR158 | AJ566489 | 4 | 5′-TGTAGGTTATGCAGCGTG TTC-3′ | 5′-GATGAGGGGTGTAG CTGTTTG-3′ | 211–217 | (CT)8 | [44] |
| MTcCIR168 | AJ566497 | 4 | 5′-GGTAGTATTGAGGTGCGT AT-3′ | 5′-GTGAATGAATGGAT GTGAAA-3′ | 174–186 | (TC)9 | [44] |
| Locus | Na | Ne | Fa | Ho | He | F | PIC |
|---|---|---|---|---|---|---|---|
| MTcCIR6 | 6.0 | 2.10 | 0.63 | 0.26 | 0.52 | 0.50 | 0.46 |
| MTcCIR7 | 5.0 | 2.24 | 0.60 | 0.39 | 0.55 | 0.30 | 0.49 |
| MTcCIR8 | 5.0 | 2.01 | 0.66 | 0.33 | 0.50 | 0.34 | 0.45 |
| MTcCIR11 | 5.0 | 2.04 | 0.62 | 0.28 | 0.51 | 0.45 | 0.43 |
| MTcCIR12 | 7.0 | 2.18 | 0.61 | 0.35 | 0.54 | 0.35 | 0.48 |
| MTcCIR18 | 5.0 | 1.97 | 0.67 | 0.30 | 0.49 | 0.39 | 0.44 |
| MTcCIR22 | 3.0 | 1.80 | 0.69 | 0.25 | 0.44 | 0.44 | 0.37 |
| MTcCIR24 | 5.0 | 2.79 | 0.52 | 0.28 | 0.64 | 0.56 | 0.59 |
| MTcCIR26 | 6.0 | 2.71 | 0.54 | 0.26 | 0.63 | 0.58 | 0.58 |
| MTcCIR33 | 11.0 | 3.93 | 0.33 | 0.38 | 0.74 | 0.49 | 0.70 |
| MTcCIR37 | 8.0 | 2.67 | 0.54 | 0.30 | 0.62 | 0.52 | 0.58 |
| MTcCIR40 | 6.0 | 2.50 | 0.60 | 0.30 | 0.60 | 0.50 | 0.57 |
| MTcCIR60 | 5.0 | 1.67 | 0.75 | 0.37 | 0.40 | 0.08 | 0.35 |
| MTcCIR115 | 3.0 | 2.06 | 0.63 | 0.28 | 0.51 | 0.45 | 0.44 |
| MTcCIR158 | 3.0 | 1.84 | 0.66 | 0.14 | 0.46 | 0.69 | 0.36 |
| MTcCIR168 | 5.0 | 1.82 | 0.71 | 0.21 | 0.45 | 0.53 | 0.41 |
| Mean | 5.5 | 2.27 | 0.61 | 0.29 | 0.54 | 0.45 | 0.48 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
López, M.; Gori, M.; Bini, L.; Ordoñez, E.; Durán, E.; Gutierrez, O.; Masoni, A.; Giordani, E.; Biricolti, S.; Palchetti, E. Genetic Purity of Cacao Criollo from Honduras Is Revealed by SSR Molecular Markers. Agronomy 2021, 11, 225. https://doi.org/10.3390/agronomy11020225
López M, Gori M, Bini L, Ordoñez E, Durán E, Gutierrez O, Masoni A, Giordani E, Biricolti S, Palchetti E. Genetic Purity of Cacao Criollo from Honduras Is Revealed by SSR Molecular Markers. Agronomy. 2021; 11(2):225. https://doi.org/10.3390/agronomy11020225
Chicago/Turabian StyleLópez, Marlon, Massimo Gori, Lorenzo Bini, Erick Ordoñez, Erick Durán, Osman Gutierrez, Alberto Masoni, Edgardo Giordani, Stefano Biricolti, and Enrico Palchetti. 2021. "Genetic Purity of Cacao Criollo from Honduras Is Revealed by SSR Molecular Markers" Agronomy 11, no. 2: 225. https://doi.org/10.3390/agronomy11020225
APA StyleLópez, M., Gori, M., Bini, L., Ordoñez, E., Durán, E., Gutierrez, O., Masoni, A., Giordani, E., Biricolti, S., & Palchetti, E. (2021). Genetic Purity of Cacao Criollo from Honduras Is Revealed by SSR Molecular Markers. Agronomy, 11(2), 225. https://doi.org/10.3390/agronomy11020225