
Selection of Diploid and Tetraploid Banana Hybrids Resistant to Pseudocercospora fijiensis

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Disease Measurement and Selection Criteria
2.3. Data Analysis
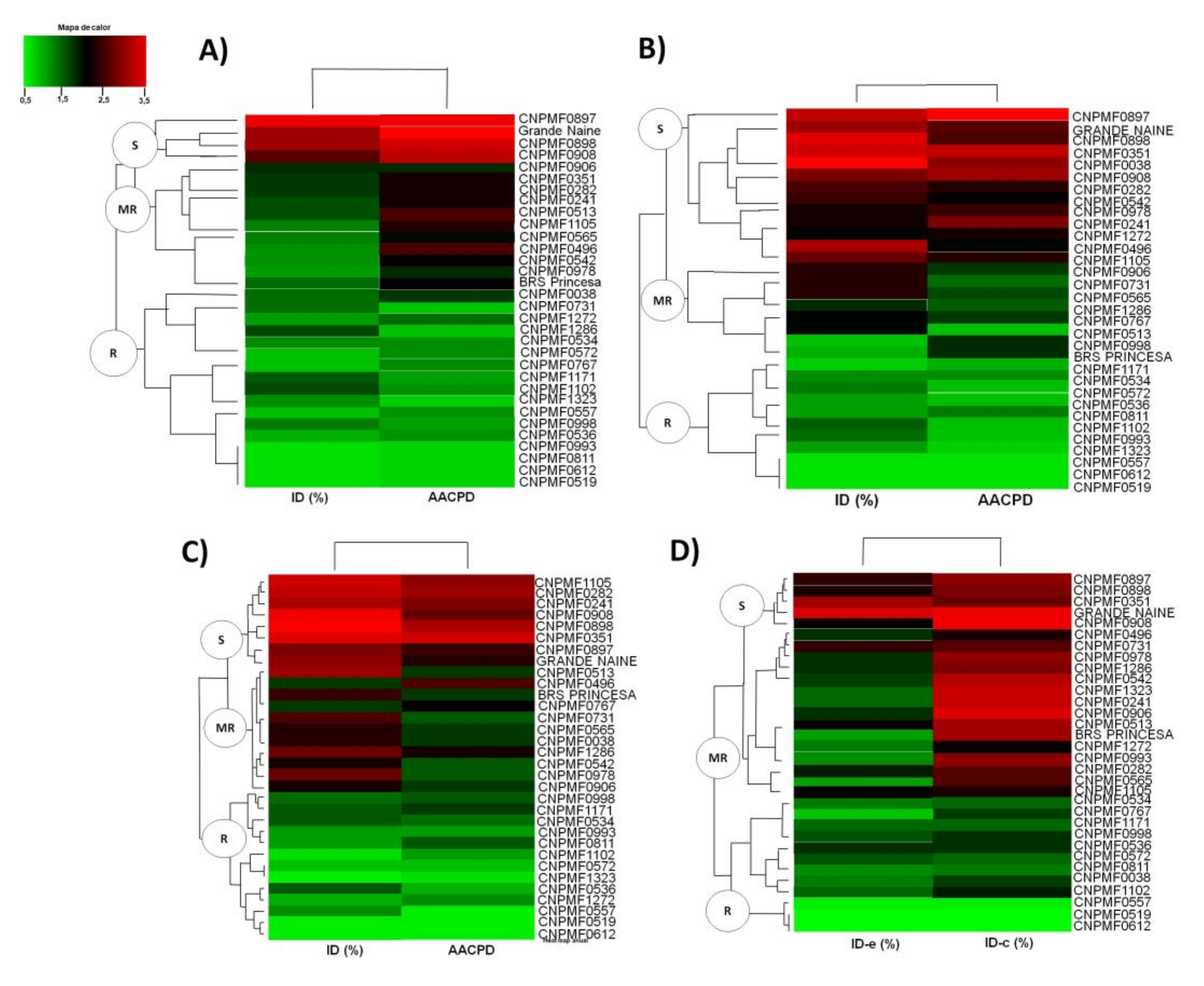
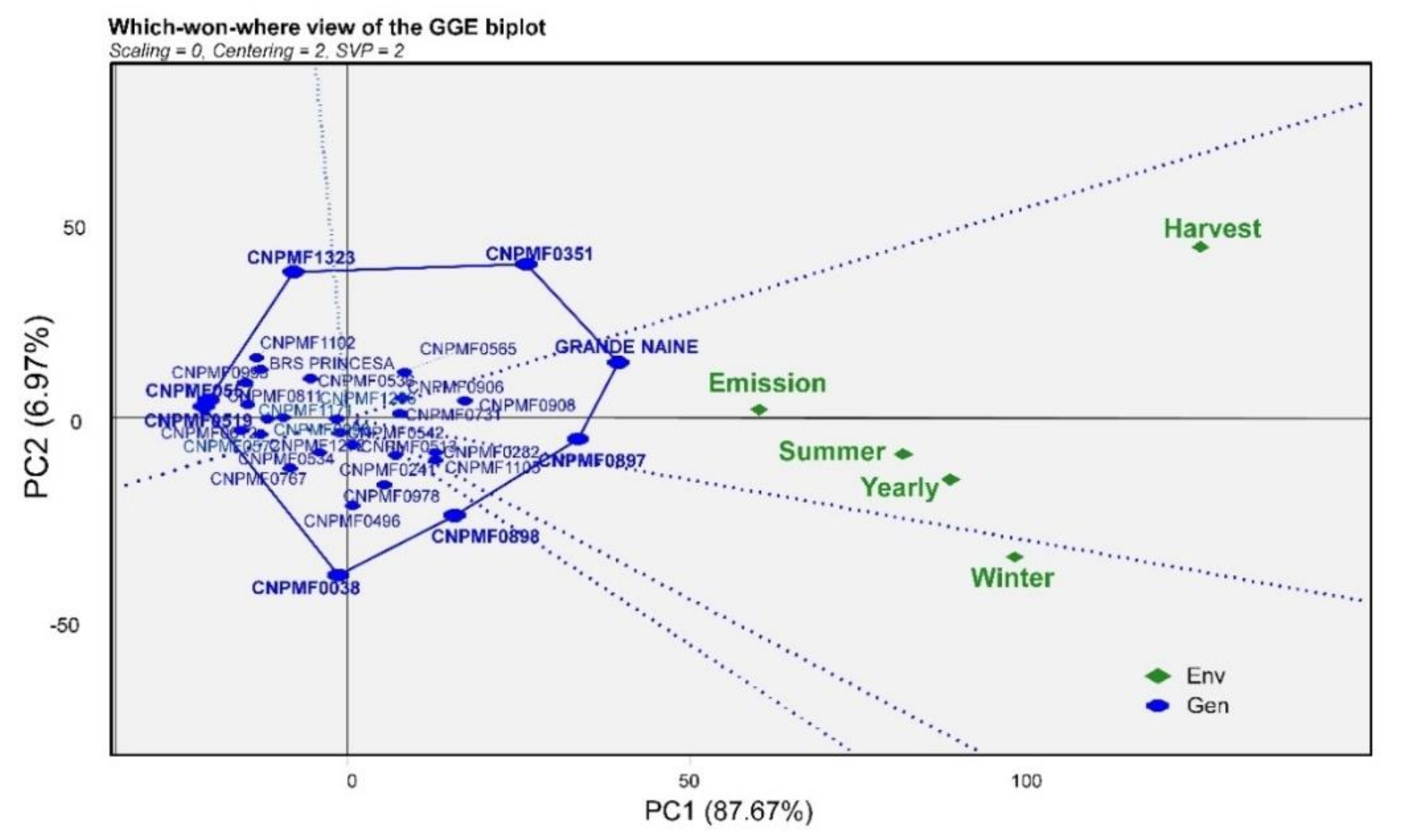
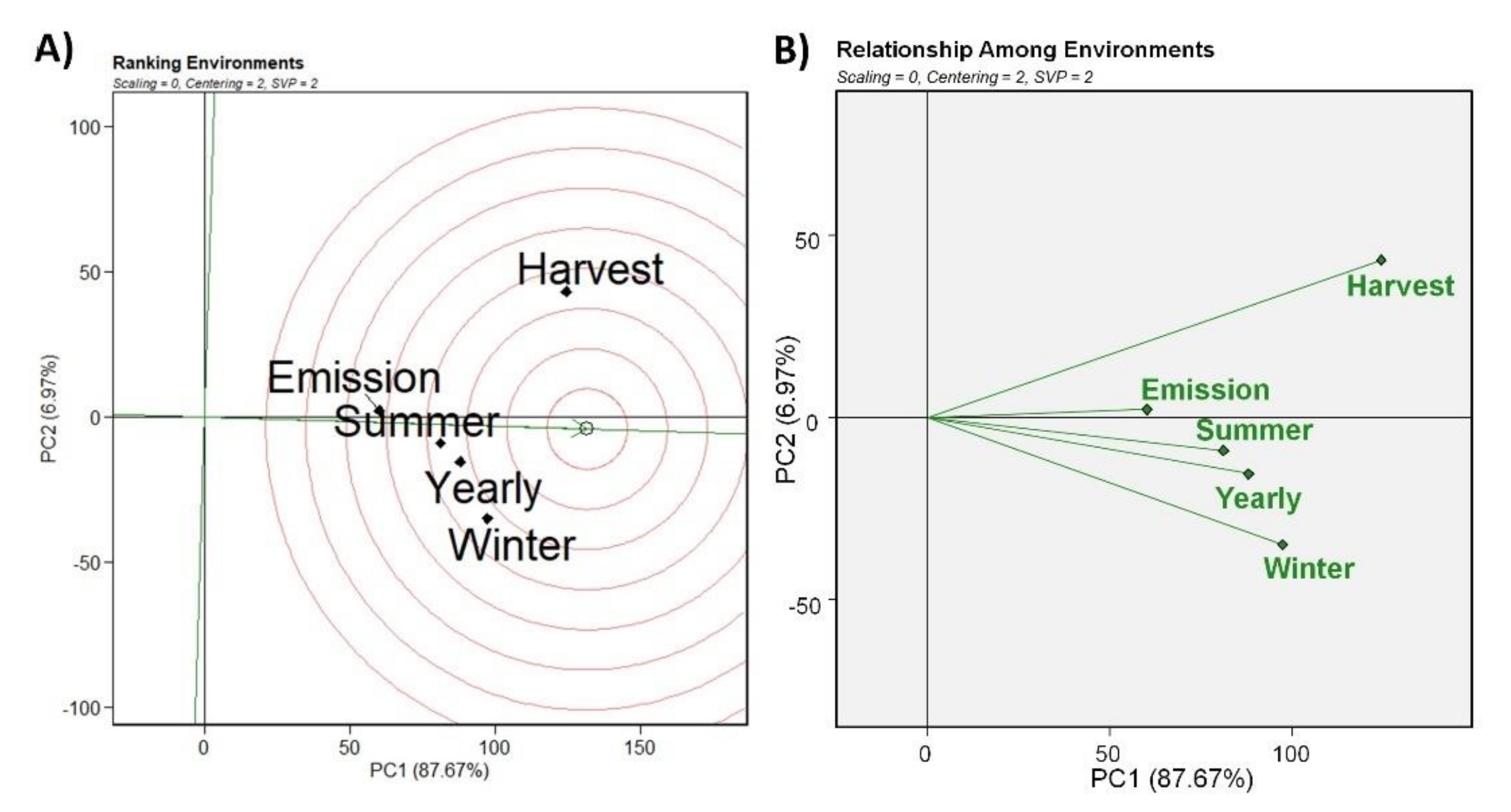
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weber, O.B.; Garruti, D.D.S.; Norões, N.P. Performance of banana genotypes with resistance to black leaf streak disease in Northeastern Brazil. Pesquisa Agropecuária Brasileira 2017, 52, 161–169. [Google Scholar] [CrossRef]
- FAOSTAT. Food and Agriculture Data. Available online: http://faostat.fao.org/ (accessed on 4 August 2021).
- Perrier, X.; De Langhe, E.; Donohue, M.; Lentfer, C.; Vrydaghs, L.; Bakry, F.; Carrel, F.; Hippolyte, I.; Horry, J.P.; Jenny, C.; et al. Multidisciplinary perspectives on banana (Musa spp.) domestication. Proc. Natl. Acad. Sci. USA 2011, 108, 11311–11318. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Qi, Y.X.; Zhang, H.; Zhang, H.Q.; Pu, J.J.; Xie, Y.X. Separation and identification of Musa acuminata Colla (banana) leaf proteins by two-dimensional gel electrophoresis and mass spectrometry. Genet. Mol. Res. 2013, 12, 6871–6881. [Google Scholar] [CrossRef]
- Isaza, R.E.A.; Dias-Trujillo, C.; Dhillon, B.; Aerts, A.; Carlier, J.; Crane, C.F.; De Jong, T.V.; De Vries, I.; Dietrich, R.; Farmer, A.D.; et al. Combating a global threat to a clonal crop: Banana black sigatoka pathogen Pseudocercospora fijiensis (Synonym Mycosphaerella fijiensis) genomes reveal clues for disease control. PLoS Genet. 2016, 12, 1–36. [Google Scholar] [CrossRef]
- Crous, P.W.; Tanaka, K.; Summerell, B.A.; Groenewald, J.Z. Additions to Mycosphaerella complex. IMA Fungus 2010, 2, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Gasparotto, L.; Pereira, J.C.R.; Trindade, D.R. Situação atual da Sigatoka negra da bananeira. Fitopatologia Brasileira 2001, 26, 449. [Google Scholar]
- Churchill, A.C.L. Mycosphaerella fijiensis, the black leaf streak pathogen of banana: Progress towards understanding pathogen biology and detection, disease development, and the challenges of control. Mol. Plant Pathol. 2011, 12, 307–328. [Google Scholar] [CrossRef] [PubMed]
- Castelana, F.P.; Saraiva, L.A.; Lange, F.; de Bellaire, L.D.L.; Cordenunsi, B.R.; Chillet, M. Effects of black leaf streak disease and Sigatoka disease on fruit quality and maturation process of bananas produced in the subtropical conditions of Southern Brazil. J. Crop. Prot. 2012, 35, 127–131. [Google Scholar] [CrossRef]
- Gagné, E.S.; Cluzet, S.; Merillon, J.M.; Geny, L. ABA initiates anthocyanin production in grape cell cultures. Plant Growth Regul. 2011, 30, 1–10. [Google Scholar] [CrossRef]
- Chillet, M.; Abadie, C.; Hubert, O.; Chilin-Charles, Y.; De Bellaire, L.D.L. Sigatoka disease reduces the greenlife of bananas. Crop Prot. 2009, 28, 41–45. [Google Scholar] [CrossRef]
- Jimenez, M.I.; Van Der Veken, L.; Neirynck, H.; Rodriguez, H.; Ruiz, O.; Swennen, R. Organic banana production in Ecuador: Its implications on black Sigatoka development and plant-soil nutritional status. Renew. Agric. Food Syst. 2007, 22, 297–306. [Google Scholar] [CrossRef]
- Alakonya, A.E.; Kimunye, J.; Mahuku, G.; Amah, D.; Uwimana, B.; Brown, A.; Swennen, R. Progress in understanding Pseudocercospora banana pathogens and the development of resistant Musa germplasm. Plant Pathol. 2018, 67, 59–770. [Google Scholar] [CrossRef]
- Soares, J.M.; Rocha, A.J.; Nascimento, F.S.; Santos, A.S.; Miller, R.N.; Ferreira, C.F.; Haddad, F.; Amorim, V.B.O.; Amorim, E.P. Genetic improvement for resistance to black Sigatoka in bananas: A systematic review. Front. Plant Sci. 2021, 12, 657916. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, R.; Vuylsteke, D. Inheritance of black Sigatoka resistance in plantain and banana (Musa spp.) hybrids. Theor. Appl. Genet. 1994, 89, 46–52. [Google Scholar] [CrossRef]
- Craenen, K.; Ortiz, R. Effect of the bs1 gene in plantain-banana hybrids on response to black Sigatoka. Theor. Appl. Genet. 1997, 95, 497–505. [Google Scholar] [CrossRef]
- Amorim, E.P.; Amorim, V.B.O.; Silva, M.S.; Haddad, F.; Ferreira, C.F.; Santos-Serejo, J.A. Developing hybrid banana varieties with improved properties. In Achieving Sustainable Cultivation of Bananas: Germplasm and Genetic Improvement; Kema, G.H.J., Drenth, A., Eds.; Burleigh Dodds Science Publishing: Cambridge, UK, 2021; Volume 2, pp. 1–17. ISBN 978 1 78676 344 0. [Google Scholar]
- Agritempo. Sistema de Monitoramento Agrometeorológico. Available online: http://www.agritempo.gov.br/agroclima/sumario (accessed on 4 August 2021).
- Ramos, J.B.; Bragança, C.A.D.; Oliveira, A.D.S.; Cordeiro, Z.J.M.; Haddad, F. First report of black Sigatoka of banana caused by Mycosphaerella fijiensis in Bahia, Brazil. Plant Dis. 2018, 102, 2035–2045. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, F.S.; Sousa, Y.M.; Rocha, A.J.; Ferreira, C.F.; Haddad, F.; Amorim, E.P. Sources of black Sigatoka resistance in wild banana diploids. Rev. Bras. Frutic. 2020, 42, 1–14. [Google Scholar] [CrossRef]
- Kimunye, J.; Were, E.; Swennen, R.; Viljoen, A.; Mahuku, G. Sources of resistence to Pseudocercospora fijiensis, the cause of black Sigatoka in banana. J. Plant Pathol. 2021, 71, 1651–1664. [Google Scholar] [CrossRef] [PubMed]
- Stover, R.H. Banana Plantain and Abaca Diseases; Commonwelth Mycological Institute: Richmond, UK, 1972; Volume 316. [Google Scholar]
- Gauhl, F.; Pasberg-Gauhl, C.; Vuylsteke, D.; Ortiz, R. Multilocational Evaluation of Black Sigatoka Resistance in Banana and Plantain; International Institute of Tropical Agriculture: Abuja, Nigeria, 1989; pp. 47–59. [Google Scholar]
- Fouré, E. Varietal reactions of bananas and plantains to black leaf streak disease. Banana Plantain Breed. Strateg. 1987, 21, 110–113. [Google Scholar]
- Mckinney, H.H. Influence of soil, temperature and moisture on infection of wheat seedlings by Helminthosporium sativum. J. Agric. Res. 1923, 26, 195–217. [Google Scholar]
- Madden, L.V.; Hugles, G.; Bosch, F.V.D. The study of plant disease epidemics. Am. Phytopathol. Soc. 2007, 32, 31–63. [Google Scholar]
- R Core Development Team. R: A Language and Environment for Statistical Computing, Version 2.12.1.; R Foundation for Statistical Computing: Vienna, Austria, 2011; ISBN 3-900051-07-0. [Google Scholar]
- Rebouças, T.A.; Haddad, F.; Ferreira, C.F.; Oliveira, S.A.S.; Ledo, C.A.S.; Amorim, E.P. Identification of banana genotypes resistant to Fusarium wilt race 1 under field and greenhouse conditions. Sci. Hortic. 2018, 239, 308–313. [Google Scholar] [CrossRef]
- Gonçalves, Z.S.; Haddad, F.; Amorim, V.B.O.; Ferreira, C.F.; Oliveira, S.A.S.; Amorim, E.P. Agronomic characterization and identification of banana genotypes resistant to Fusarium wilt race 1. Eur. J. Plant Pathol. 2019, 155, 1093–1103. [Google Scholar] [CrossRef]
- Yan, W.; Tinker, N.A. Biplot analysis of multi-environment trial data: Principles and applications. Can. J. Plant Sci. 2006, 86, 623–645. [Google Scholar] [CrossRef]
- Yan, W.; Kang, M.S.; Ma, B.; Woods, S.; Cornelius, P.L. GGE biplot vs. AMMI analysis of genotype-by-environment data. Crop Sci. 2007, 47, 641–653. [Google Scholar] [CrossRef]
- Olivoto, T.; Lúcio, A.D.C. Metan: An R package for multi-environment trial analysis. Methods Ecol. Evol. 2020, 11, 783–789. [Google Scholar] [CrossRef]
- Jones, D.R. Sigatoka. In Diseases of Banana, Abacá and Enset; CABI Publishing: New York, NY, USA, 2000; pp. 79–92. [Google Scholar]
- Cordeiro, Z.J.M.; Matos, A.P.; Haddad, F. Doenças fúngicas e bacterianas. In O Agronegórcio da Banana; Ferreira, C.F., Silva, S.O., Amorim, E.P., Santos-Serejo, J.A., Eds.; Embrapa: Brasília, Brasil, 2016; Volume 1, pp. 111–136. [Google Scholar]
- Barekye, A.; Tongoona, P.; Derera, J.; Laing, M.; Tushermereirwe, W.K. Appraisal of methods for assessing black Sigatoka resistance in diploid banana populations. Afr. J. Plant Sci. 2011, 5, 900–908. [Google Scholar] [CrossRef]
- Gauhl, F.; Pasberg-Gauhl, C.; Jones, D.R. Black leaf streak: Disease cycle and epidemiology. In Diseases of Banana, Abacá and Enset; CABI Publishing: New York, NY, USA, 2000; pp. 56–62. [Google Scholar]
- Kimunye, J.; Jomanga, K.; Tazuba, A.F.; Were, E.; Viljoen, A.; Swennen, R.; Mahuku, G. Genotype X environment response of ‘Matooke’ hybrids (Naritas) to Pseudocercospora fijiensis, the cause of black Sigatoka in banana. Agronomy 2021, 11, 1145. [Google Scholar] [CrossRef]
- Ortiz-Vasquez, E.; Kaemmer, D.; Zhang, H.B.; Muth, J.; Rodriguez-Mendiola, M.; Arias-Castro, C.; James, A. Construction and characterization of a plant transformation-competent BIBAC library of the black Sigatoka-resistant banana Musa acuminata cv. Tuu Gia (AA). Theor. Appl. Genet. 2005, 110, 706–713. [Google Scholar] [CrossRef]
- Timm, E.S.; Pardo, H.L.; Coello, R.P.; Navarrete, T.C.; Villegas, O.N.; Ordonez, E.S. Identification of differentially-expressed genes in response to Mycosphaerella fijiensis in the resistant Musa accession ‘Calcutta-4’ using suppression subtractive hybridization. PLoS ONE 2016, 11, 1–17. [Google Scholar] [CrossRef][Green Version]
- Amorim, E.P.; Santos-Serejo, J.A.; Amorim, V.B.O.; Ferreira, C.F.; Silva, S. Banana breeding at Embrapa cassava and fruits. Acta Hortic. 2013, 986, 171–176. [Google Scholar] [CrossRef]
- Chillet, M.; Castelana, F.P.; Abadie, C.; Hubert, O.; Chilin-Charles, Y.; De Bellaire, L.L. Effect of different levels of Sigatoka disease severity on banana pulp colour and early ripening. Can. J. Plant Pathol. 2014, 36, 48–53. [Google Scholar] [CrossRef]
- De Bellaire, L.L.; Four, E.; Abadie, C.; Carlier, J. Black leaf streak disease is challenging the banana industry. Fruits 2010, 65, 327–342. [Google Scholar] [CrossRef]
- Calberto, G.G.; Staver, C.; Siles, P. An assessment of global banana production and suitability under climate change scenarios. In Climate Change and Food Systems: Global Assessments and Implications for Food Security and Trade; Elbehri, A., Ed.; Food Agriculture Organization of the United Nations (FAO): Rome, Italy, 2015; pp. 266–291. [Google Scholar]
- Yonow, T.; Ramirez-Villegas, J.; Abadie, C.; Darnell, R.E.; Ota, N.; Kriticos, D.J. Black Sigatoka in bananas: Ecoclimatic suitability and disease pressure assessments. PLoS ONE 2019, 14, 1–25. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Codes | Genealogies |
|---|---|
| Improved Diploids | |
| CNPMF0038 | [(M53 × Madu)] × [(Malaccensis × Tjau Lagada)] |
| CNPMF0496 | [(M61 × Pisang Lilin)] × [(Terrinha × Calcutta 4)] |
| CNPMF0513 | [(M61 × Pisang Lilin)] × [(M53 × Kumburgh)] |
| CNPMF0519 | Self-fertilization (wild diploid Tambi) |
| CNPMF0534 | [(Calcutta 4 × Madang)] × [(Borneo × Guyod)] |
| CNPMF0536 | [(Calcutta 4 × Madang)] × [(Borneo × Guyod)] |
| CNPMF0542 | [(SH3263)] × [(Malaccensis × Sinwobogi)] |
| CNPMF0557 | [(M61 × Pisang Lilin)] × [(Malaccensis × Tjau Lagada)] |
| CNPMF0565 | [(Calcutta 4 × Pahang) × (Borneo × Madang)] × Khae |
| CNPMF0572 | [(Khai × (Calcutta 4 × Madang)] × [(Calcutta 4 × Madang)] |
| CNPMF0612 | [(M53 × Madu) × Madu)] × SH3263 |
| CNPMF0731 | [(Malaccensis × Madang)] × [(Tuugia × Calcutta 4)] |
| CNPMF0767 | [(Malaccensis × Madang)] × [(Khai × (Calcutta 4 × Madang)] |
| CNPMF0811 | [(Khai × (Calcutta 4 × Madang)] × [(Calcutta 4 × Pahang) × (Borneo × Madang)] |
| CNPMF0978 | [(Calcutta 4 × Madang)] × [(Terrinha × Calcutta 4)] |
| CNPMF0993 | [(Borneo × Guyod) × (Tuugia × Calcutta 4)] × [(Khai × (Calcutta 4 × Madang)] |
| CNPMF0998 | [(Borneo × Guyod)] × [(Borneo × Guyod) × SH3263] |
| CNPMF1102 | [(Jari Buaya × (Calcutta 4 × Madang)] × [(Borneo × Guyod) × (Tuugia × Calcutta 4)] |
| CNPMF1105 | [(Borneo × Guyod) × (Calcutta 4 × Heva)] × [(Calcutta 4 × Madang)] |
| CNPMF1171 | [(Malaccensis × Madang)] × [(M53 × (Tuugia × Calcutta 4)] |
| CNPMF1272 | [(Borneo × Guyod) × (Calcutta 4 × Heva)] × [(Tuugia × Calcutta 4)] |
| CNPMF1286 | [(Calcutta 4 × Madang)] × [(Terrinha × Calcutta 4)] |
| CNPMF1323 | [(Malaccensis × Sinwobogi)] × [(Calcutta 4 × Heva)] |
| Tetraploids hybrids | |
| CNPMF0241 | [(Pacovan × improved diploid *)] |
| CNPMF0282 | [(Pacovan × improved diploid *)] |
| CNPMF0351 | [(Prata Anã × improved diploid by FHIA 18)] |
| CNPMF0897 | [(Prata Anã)] × [(Malaccensis × Sinwobogi) × (Zebrina × Heva)] |
| CNPMF0898 | [(Prata Anã)] × [(Malaccensis × Sinwobogi) × (Calcutta 4 × Galeo)] |
| CNPMF0906 | [(Prata Santa Maria × improved diploid *)] |
| CNPMF0908 | [(Silk × improved diploid *)] |
| Commercial cultivars | |
| BRS Princesa | [(Yangambi × M53)] |
| Grand Naine | - |
| Genotypes | Summer | Winter | Whole Year | Emission | Harvest | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ID | AUDPC | LR | ID | AUDPC | LR | ID | AUDPC | LR | ID | ID | LR | |
| CNPMF0897 | 57.2a | 3014.0a | S | 53.6a | 3460.9a | S | 56.4a | 3408.2a | S | 40.7a | 75.9a | S |
| G. Naine | 68.0a | 3146.3a | S | 57.3a | 3974.8a | S | 55.8a | 3303.5a | S | 52.9a | 80.6a | S |
| CNPMF0351 | 33.3b | 1865.4b | MR | 50.0a | 3454.4a | S | 41.0a | 2832.6a | S | 30.7a | 97.2a | S |
| CNPMF0038 | 52.0a | 2748.8a | S | 32.8b | 2126.4a | S | 39.3a | 2471.6a | S | 36.1a | 93.5a | S |
| CNPMF0282 | 38.5b | 1922.3b | MR | 45.8a | 1857.0b | S | 38.5a | 2436.4a | S | 27.9a | 50.0a | S |
| CNPMF0908 | 45.2a | 2255.7a | S | 36.3b | 2896.4a | S | 38.1a | 2310.2a | S | 35.4a | 37.8a | S |
| CNPMF0241 | 33.8b | 1856.6b | MR | 46.4a | 2707.0a | S | 37.0a | 2338.5a | S | 15.8b | 36.1a | MR |
| CNPMF1105 | 3.7c | 347.3c | MR | 45.1a | 211.1d | S | 34.3a | 2227.1a | S | 14.9b | 25.0c | MR |
| CNPMF0898 | 33.2b | 2748.8a | S | 23.9b | 1340.2b | MR | 25.0a | 1535.1b | S | 16.6b | 25.0b | MR |
| CNPMF0998 | 30.6b | 2405.6a | MR | 31.0b | 2622.8a | MR | 23.3b | 2001.2b | MR | 14.3b | 16.7b | MR |
| CNPMF0906 | 30.6b | 1623.7b | MR | 28.9b | 1984.8b | MR | 30.0a | 1804.0b | MR | 22.2b | 44.4a | MR |
| CNPMF0542 | 37.1b | 2111.2b | MR | 36.7a | 2018.5a | S | 28.4a | 1680.0b | MR | 13.0c | 35.3a | MR |
| CNPMF0565 | 30.1b | 2313.3b | MR | 30.8b | 1327.8b | MR | 27.5a | 1337.4b | MR | 23.7b | 36.8a | MR |
| CNPMF0496 | 24.8b | 1816.7b | MR | 35.6a | 1816.7a | S | 23.3b | 1135.5b | MR | 11.1c | 20.3b | MR |
| CNPMF0513 | 30.4b | 1533.3b | MR | 10.7c | 403.8c | R | 22.0b | 1124.2b | MR | 19.0b | 24.8b | MR |
| CNPMF1102 | 10.8c | 1208.4b | R | 19.1b | 1123.2b | MR | 19.1b | 1190.3b | MR | 8.3c | 29.7d | R |
| CNPMF0731 | 8.8c | 659.8c | R | 20.4b | 905.6c | MR | 17.5b | 808.2c | MR | 17.1b | 18.5b | MR |
| CNPMF0978 | 6.9c | 125.0d | R | 48.7a | 1757.0a | S | 16.0b | 356.1c | MR | 16.0b | 21.5b | MR |
| BRSPrincesa | 19.5b | 438.8c | R | 13.3c | 860.2c | MR | 15.7b | 795.4c | MR | 14.9b | 25.9b | MR |
| CNPMF0767 | 6.3c | 573.0c | R | 12.6c | 778.7c | MR | 14.0b | 921.3c | MR | 3.8d | 7.4e | R |
| CNPMF1286 | 6.8c | 652.9c | R | 14.1c | 581.1c | MR | 13.4b | 590.2c | MR | 14.9b | 16.6b | MR |
| CNPMF0534 | 7.3c | 856.0b | R | 10.3c | 525.7c | R | 8.9b | 581.5c | R | 2.0d | 14.8b | R |
| CNPMF1272 | 10.8c | 757.0c | R | 43.4a | 1425.0b | S | 9.1b | 524.8c | R | 14.8b | 25.0d | MR |
| CNPMF0993 | 4.0c | 444.5c | R | 11.1c | 515.6c | R | 10.9b | 481.0c | R | 7.4c | 16.7b | MR |
| CNPMF1171 | 2.1c | 222.3c | R | 11.5c | 778.7c | R | 5.9c | 379.2c | R | 7.4c | 14.9d | R |
| CNPMF0572 | 3.7c | 474.1c | R | 9.5c | 357.9c | R | 4.5c | 291.5c | R | 1.9d | 1.9f | R |
| CNPMF0811 | 0.0d | 0.0d | R | 5.9d | 169.4d | R | 4.3c | 121.3d | R | 3.8d | 14.8d | R |
| CNPMF0536 | 0.4d | 20.9d | R | 4.1d | 238.1d | R | 4.3c | 233.6d | R | 3.8d | 5.7e | R |
| CNPMF1323 | 0.0d | 0.0d | R | 0.7d | 41.7d | R | 2.2c | 100.7d | R | 11.0b | 11.2e | R |
| CNPMF0519 | 0.0d | 0.0d | R | 0.0d | 0.0d | R | 0.0d | 0.0e | R | 0.0d | 0.0f | R |
| CNPMF0557 | 0.0d | 0.0d | R | 0.0d | 0.0d | R | 0.0d | 0.0e | R | 1.4d | 1.41f | R |
| CNPMF0612 | 0.0d | 0.0d | R | 0.0d | 0.0d | R | 0.0d | 0.0e | R | 2.8d | 2.9f | R |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonçalves, Z.S.; de Jesus Rocha, A.; Haddad, F.; de Oliveira Amorim, V.B.; Ferreira, C.F.; Amorim, E.P. Selection of Diploid and Tetraploid Banana Hybrids Resistant to Pseudocercospora fijiensis. Agronomy 2021, 11, 2483. https://doi.org/10.3390/agronomy11122483
Gonçalves ZS, de Jesus Rocha A, Haddad F, de Oliveira Amorim VB, Ferreira CF, Amorim EP. Selection of Diploid and Tetraploid Banana Hybrids Resistant to Pseudocercospora fijiensis. Agronomy. 2021; 11(12):2483. https://doi.org/10.3390/agronomy11122483
Chicago/Turabian StyleGonçalves, Zalmar Santana, Anelita de Jesus Rocha, Fernando Haddad, Vanusia Batista de Oliveira Amorim, Claudia Fortes Ferreira, and Edson Perito Amorim. 2021. "Selection of Diploid and Tetraploid Banana Hybrids Resistant to Pseudocercospora fijiensis" Agronomy 11, no. 12: 2483. https://doi.org/10.3390/agronomy11122483
APA StyleGonçalves, Z. S., de Jesus Rocha, A., Haddad, F., de Oliveira Amorim, V. B., Ferreira, C. F., & Amorim, E. P. (2021). Selection of Diploid and Tetraploid Banana Hybrids Resistant to Pseudocercospora fijiensis. Agronomy, 11(12), 2483. https://doi.org/10.3390/agronomy11122483