
Changes in Soil-Borne Communities of Arbuscular Mycorrhizal Fungi during Natural Regrowth of Abandoned Cattle Pastures Are Indicative of Ecosystem Restoration

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Sampling Design
2.2. Soil Sampling
2.3. Soil Physicochemical Analysis
2.4. Molecular Analysis
2.5. Bioinformatics
2.6. Statistical Analysis
3. Results
3.1. Soil Conditions
3.2. AM Fungal Sequencing Data
3.3. AM Fungal Abundance, Richness and Diversity
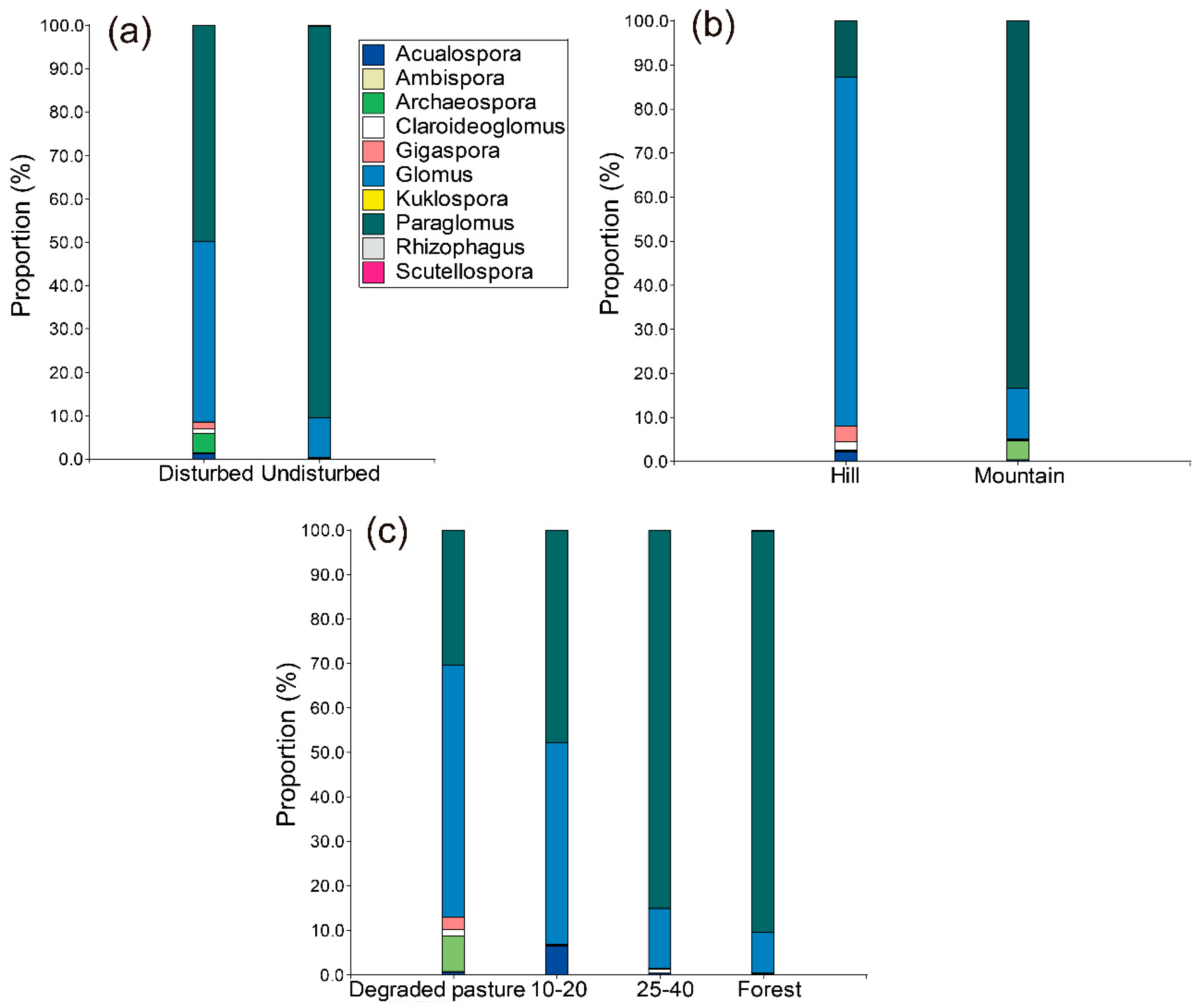
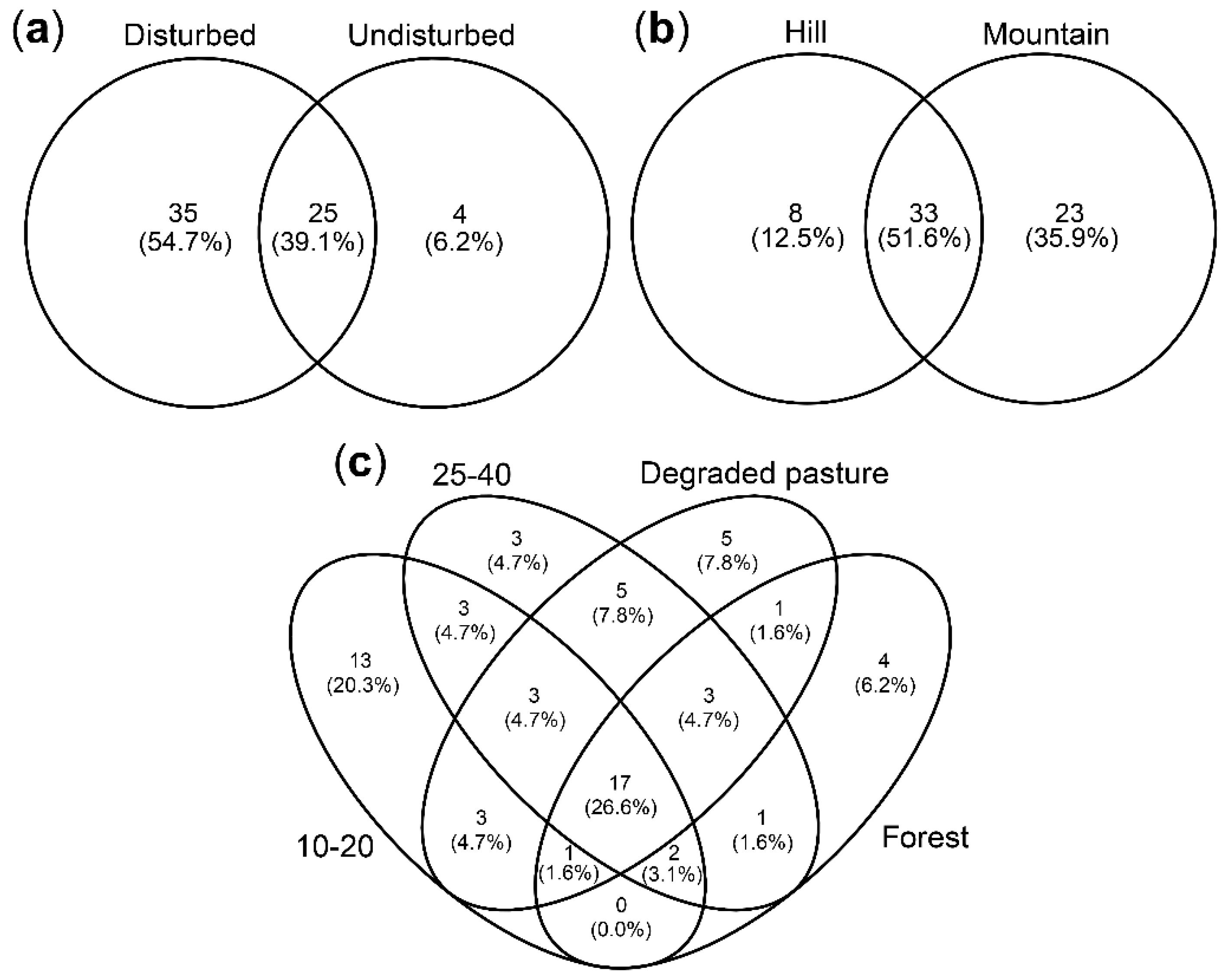
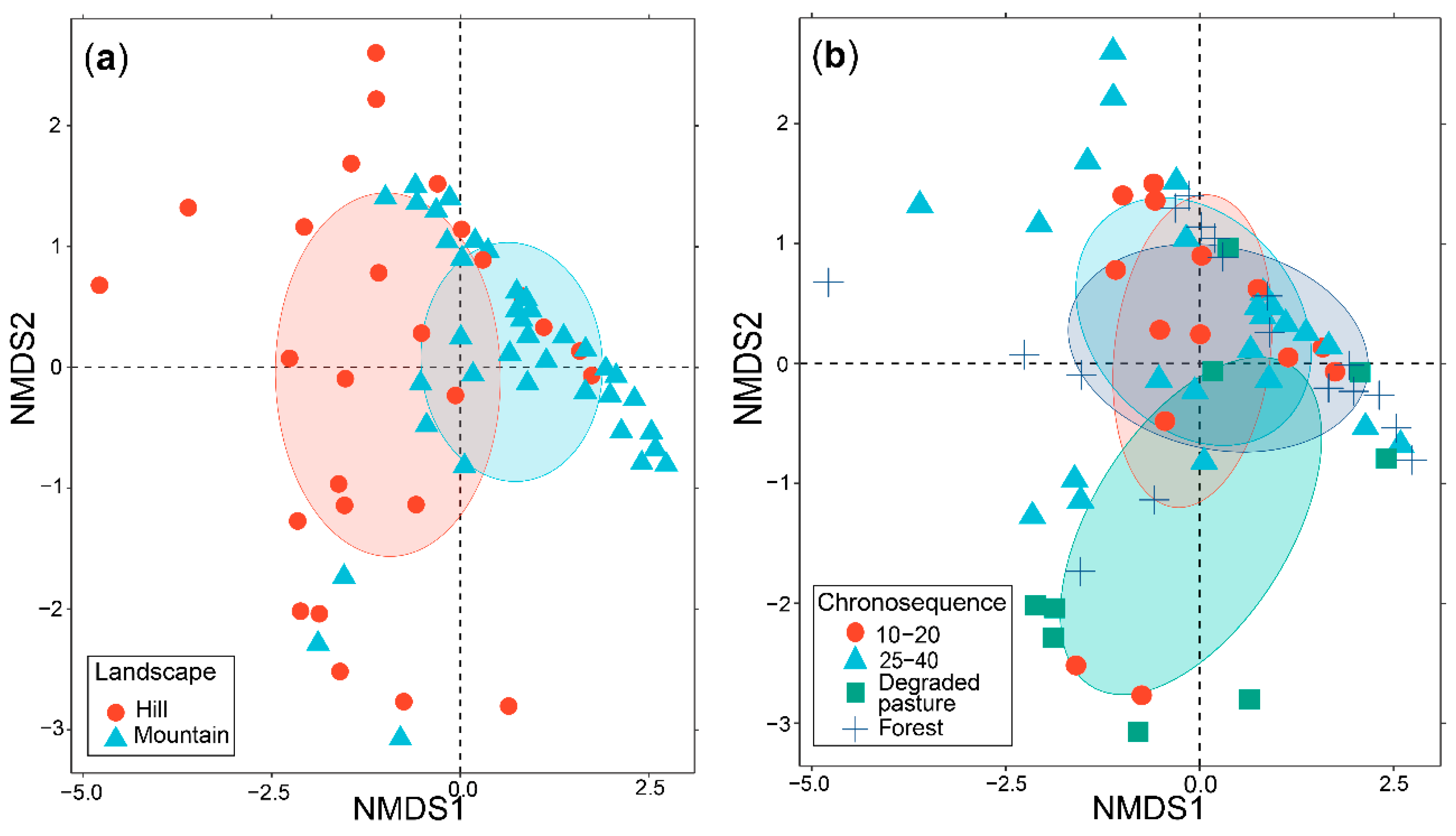
3.4. AM Fungal Community Composition
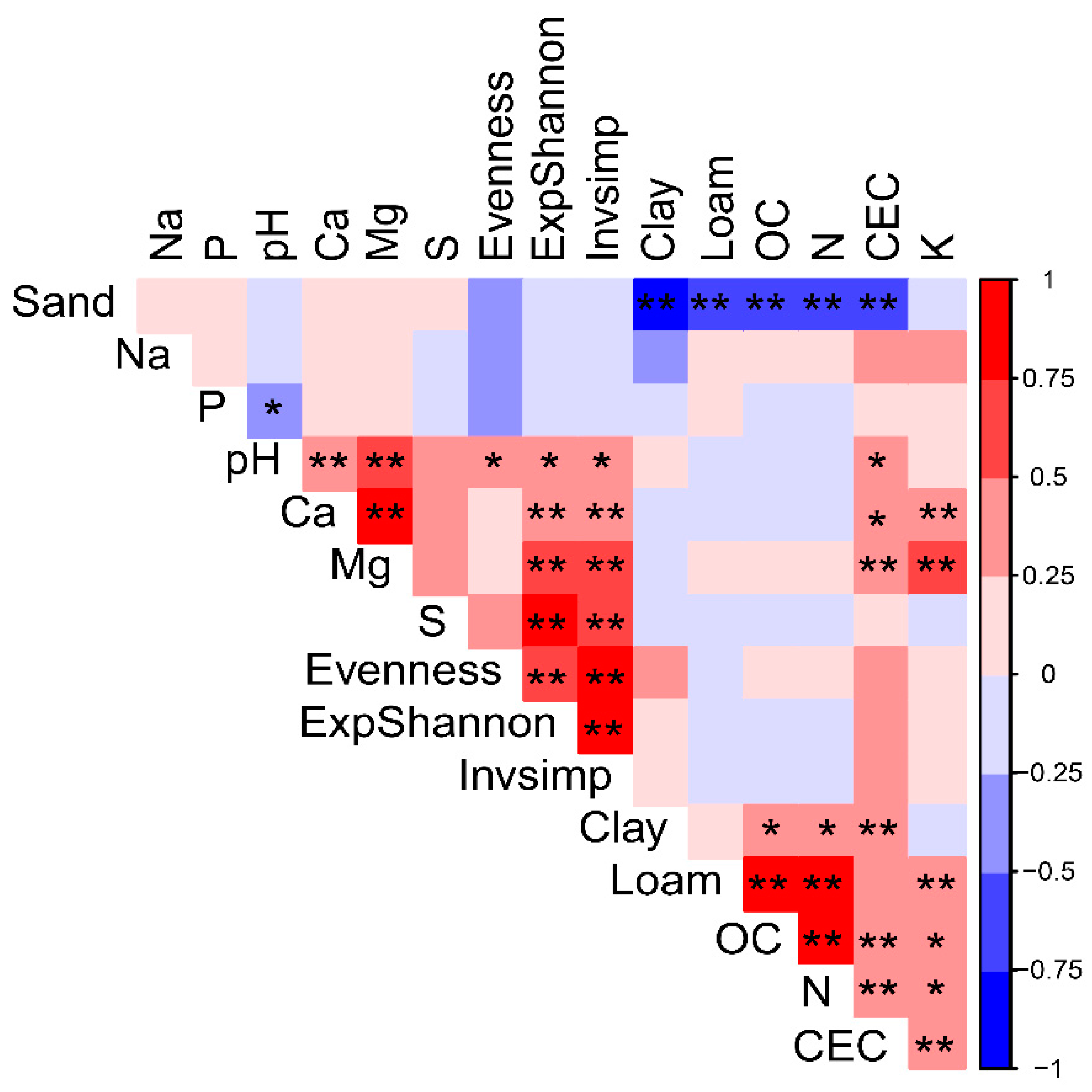
3.5. Relationship between Soil Properties and AM Fungal Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, S.E.; Read, D. Mycorrhizal Symbiosis; Elsevier: New York, NY, USA, 2008. [Google Scholar]
- Souza, T. Handbook of Arbuscular Mycorrhizal Fungi; Springer: Basel, Switzerland, 2015. [Google Scholar]
- Pérez-Lamarque, B.; Selosse, M.-A.; Öpik, M.; Morlon, H.; Martos, F. Constraints upon cheating emergence in mutualistic networks: Evidence in the mycorrhizal symbiosis. New Phytol. 2020, 226, 1822–1835. [Google Scholar] [PubMed]
- Sepp, S.-K.; Davison, J.; Jairus, T.; Vasar, M.; Moora, M.; Zobel, M.; Öpik, M. Non-random association patterns in plant-mycorrhizal fungal network reveal host-symbiont specificity. Mol. Ecol. 2019, 28, 365–378. [Google Scholar] [CrossRef]
- Zheng, C.; Ji, B.; Zhang, J.; Zhang, F.; Bever, J.D. Shading decreases plant carbon preferential allocation towards the most beneficial mycorrhizal mutualist. New Phytol. 2015, 205, 361–368. [Google Scholar] [CrossRef]
- Davison, J.; Moora, M.; Semchenk, M.; Adenan, S.B.; Ahmed, T.; Akhmetzhanova, A.A.; Alatalo, J.M.; Al-Qurishy, S.; Andriyanova, E.; Bahram, M.; et al. Temperature and pH define the realised niche space of arbuscular mycorrhizal fungi. New Phytol. 2021, 231, 763–776. [Google Scholar] [CrossRef]
- Entry, J.A.; Rygiewicz, P.T.; Watrud, L. S:; Donnelly, P.K. Influence of adverse soil conditions on the formation and function of arbuscular mycorrhizas. Adv. Environ. Res. 2002, 7, 123–138. [Google Scholar] [CrossRef]
- Moora, M.; Berger, S.; Davison, J.; Öpik, M.; Bommarco, R.; Bruelheide, H.; Kühn, I.; Kunin, W.E.; Metsis, M.; Rortais, A.; et al. Alien plants associate with widespread generalist arbuscular mycorrhizal fungal taxa: Evidence from a continental-scale study using massively parallel 454 sequencing. J. Biogeogr. 2011, 38, 1305–1317. [Google Scholar] [CrossRef]
- Cardil, A.; de-Miguel, S.; Silva, C.A.; Reich, P.B.; Calkin, D.; Brancalion, P.H.S.; Vibrans, A.C.; Gamarra, J.G.P.; Zhou, M.; Pijanowski, B.C.; et al. Recent deforestation drove the spikein Amazonian fires. Environ. Res. Lett. 2020, 15, 121003. [Google Scholar] [CrossRef]
- Sonter, L.J.; Herrera, D.; Barrett, D.J.; Galford, G.L.; Moran, C.J.; Soares-Filho, B.S. Mining drives extensive deforestationin the Brazilian Amazon. Nat. Commun. 2017, 8, 1013. [Google Scholar] [CrossRef] [Green Version]
- IDEAM. Boletín de Detección Temprana de Deforestación (DT-D); Instituto de Hidroglogìa, Meteorología y Estudos Ambientales IDEAM, Eds.; Instituto de Hidroglogìa, Metereología y Estudios Ambientales IDEAM: Bogotá, Colombia, 2020; p. 2. [Google Scholar]
- Siqueira, J.O.; Saggin-Junior, O.J. Dependency on arbuscular mycorrhizal fungi and responsiveness of some Brazilian native woody species. Mycorrhiza 2001, 11, 245–255. [Google Scholar] [CrossRef]
- Martínez, L.J.; Zinck, J.A. Temporal Variation of Soil Compactation and Deterioration of Soil Quality in Pasture Areas of Colombian Amazonia. Soil Tillage Res. 2004, 75, 3–18. [Google Scholar] [CrossRef]
- Marinho, F.; Silva, I.R.; Oehl, F.; Maia, L.C. Checklist of arbuscular mycorrhizal fungi in tropical forests. Sydowia 2018, 70, 107–127. [Google Scholar]
- Stürmer, S.L.; Siqueira, J.O. Species richness and spore abundance of arbuscular mycorrhizal fungi across distinct land uses in Western Brazilian Amazon. Mycorrhiza 2011, 21, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Heyde, M.; Bennett, J.A.; Pither, J.; Hart, M. Longterm effects of grazing on arbuscular mycorrhizal fungi. Agric. Ecosyst. Environ. 2017, 243, 27–33. [Google Scholar] [CrossRef]
- Leal, P.L.; Siqueira, J.O.; Stürmer, S.L. Switch of tropical Amazon forest to pasture affects taxonomic composition but not species abundance and diversity of arbuscular mycorrhizal fungal community. Appl. Soil Ecol. 2013, 71, 72–80. [Google Scholar] [CrossRef]
- Reyes, H.A.; Ferreira, P.F.A.; Silva, L.C.; da Costa, M.G.; Nobre, C.P.; Gehring, C. Arbuscular mycorrhizal fungi along secondary forest succession at the eastern periphery of Amazonia: Seasonal variability and impacts of soil fertility. Appl. Soil Ecol. 2019, 136, 1–10. [Google Scholar] [CrossRef]
- Sanders, I. Plant and arbuscular mycorrhizal fungal dliversity—Are we looking at the relevant levels of diversity and are we using the right techniques? New Phytol. 2004, 164, 415–418. [Google Scholar] [CrossRef] [Green Version]
- García de León, D.; Neuenkamp, L.; Moora, M.; Öpik, M.; Davison, J.; Peña-Venegas, C.P.; Vasar, M.; Jairus, T.; Zobel, M. Arbuscular mycorrhizal fungal communities in tropical rain forest are resilient to slash-and-burn agroculture. J. Trop. Ecol. 2018, 34, 186–199. [Google Scholar] [CrossRef]
- Murad, C.A.; Pearse, J. Landsat study of deforestation in the Amazon region of Colombia: Departments of Caquetá and Putumayo. Remote Sens. Appliction Soc. Environ. 2018, 11, 161–171. [Google Scholar] [CrossRef]
- IGAC. Estudio General de Suelos y Zonificación de Tierras Departamento del Caquetá Escala 1:100.000; Instituto Geográfico Agustín Codazzi IGAC, Ed.; Instituto Geofráfico Agustín Codazzi IGAC: Bogotá, Columbia, 2014. [Google Scholar]
- IGAC. Caquetá, Características Geográficas; Imprenta Nacional: Bogotá, Colombia, 2010. [Google Scholar]
- Rodríguez, L.; Suárez, J.C.; Pulleman, M.; Guaca, L.; Rico, A.; Romero, M.; Quintero, M.; Lavelle, P. Agroforestry Systems in the Colombian Amazon Improve the Provision of Soil Ecosystem Services. Appied Soil Ecol. 2021, 164, 103933. [Google Scholar] [CrossRef]
- Blanco, J.C.; Rojas, A.C.; Rodríguez, C.H.; Malagón, R. Relación entre índices de vegetación y estados de degradación de pasturas en zonas de lomerìo del departamento de Caquetá. Momentos Cienc. 2014, 11, 35–41. [Google Scholar]
- Dumbrell, A.J.; Ashton, P.D.; Aziz, N.; Feng, G.; Nelson, N.; Dytham, C.; Fitter, A.H.; Helgason, T. Distinct seasonal assemblages of arbuscular mycorrhizal fungi revealed by massively parallel pyrosequencing. New Phytol. 2011, 190, 794–804. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, S.; Young, J.P.W. Improved PCR primers for the detection and identi¢cation of arbuscular mycorrhizal fungi. FEMS Microb. Ecol. 2008, 65, 339–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peña-Venegas, C.P.; Sterling, A.; Andrade-Ramírez, T.K. Arbucular mycorrhization in Colombian and introduced rubber (Hevea brasiliensis) genotypes cultivated on degraded soils of the Amazon region. Agriculture 2021, 11, 361. [Google Scholar] [CrossRef]
- Vasar, M.; Anderson, R.; Davison, J.; Jairus, T.; Moora, M.; Remm, M.; Young, J.P.W.; Zobel, M.; Öpik, M. Increased sequencing depth does not increase captured diversity of arbuscular mycorrhizal fungi. Mycorrhiza 2017, 27, 761–773. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M..; Clamp, M.; Barton, G.J. Jalview version 2—A multiple sequence aligment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [Green Version]
- Milne, I.; Linder, D.; Bayer, M.; Husmeier, D.; McGuire, G.; Marshall, D.F.; Wright, F. TOPALi v2: A rich graphical interface for evolutionary analyses of multiple alignments on HPC clusters and multi-core desktops. Bioinformatics 2009, 25, 126–127. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadoupoulos, J.; Bealer, K.; Madden, T.L. BLAST +: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Kindt, R. Package ’Biodiversity R´ Community Ecology and Suitability Analysis. 2020. Available online: https://cran.r-project.org/web/packages/BiodiversityR/index.html (accessed on 25 May 2021).
- R Core Team. R: A Language and Environmental for Statistical Computing. 2020. Available online: http://r.meteo.uni.wroc.pl/web/packages/dplR/vignettes/intro-dplR.pdf (accessed on 25 May 2021).
- Oksanen, J.; Blanchet, G.; Friendly, M.; Kindt, R.; Lagendre, P.; McGlinn, D.; Minchin, P.; O´Hara, R.; Simpson, G.; Solymos, P.; et al. Package ´Vegan´: Community Ecology Package; The Comprensive R Archive Network: Vienna, Austra, 2018. [Google Scholar]
- Yan, L. Ggven: Draw Venn Diagram by Ggplot2 R Package. 2021. Available online: https://cran.r-project.org/web/packages/ggvenn/ggvenn.pdf (accessed on 27 September 2021).
- Di Renzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat; 2020. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. Nmle: Linear and Nonlinear Mixed Effects Models R Package. 2018. Available online: https://cran.r-project.org/web/packages/nlme/nlme.pdf (accessed on 27 September 2021).
- Roberts, D.W. Package: “labdsv”: Ordination and Multivariate Analysis for Ecology Package. 2019. Available online: https://rdrr.io/cran/labdsv/ (accessed on 27 September 2021).
- Wei, T.; Simko, V.; Levy, M.; Xie, Y.; Jin, Y.; Zemla, J. Package “corrplot”: Visualization of a Correlation Matrix Package. 2020. Available online: https://rdrr.io/cran/corrplot/ (accessed on 27 September 2021).
- Gu, Z. Package “circlize” Circular Visualization Package. 2021. Available online: https://rdrr.io/cran/circlize/man/circlize-package.html (accessed on 27 September 2021).
- Saks, U.l.; Davison, J.; Öpik, M.; Vasar, M.; Moora, M.; Zobel, M. Root-colonizing and soil-borne communities of arbuscular mycorrhizal fungi in a temperate forest understorey. Botany 2014, 92, 277–285. [Google Scholar] [CrossRef]
- Valera-Cervero, S.; Vasar, M.; Davison, J.; Barea, J.M.; Öpik, M.; Azcón-Aguilar, C. The composition of arbuscular mycorrhizal fungal communities differ among the roots, spores and extraradical mycelia associated with five mediterranean plant species. Environ. Microbiol. 2015, 17, 2882–2895. [Google Scholar] [CrossRef]
- García de León, D.; Moora, M.; Öpik, M.; Neuenkamp, L.; Gerz, M.; Jairus, T.; Vasar, M.; Bueno, C.G.; Davison, J.; Zobel, M. Symbiont dynamics during ecosystem succession: Co-occurring plant and arbuscular mycorrhizal fungal communities. FEMS Microb. Ecol. 2016, 92, fiw097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siqueira, J.O.; Cameiro, M.A.C.; Curi, N.; Rosado, S.C.S.; Davide, A.C. Mycorrhizal colonization and mycotrophic growth of native woody species as related to successional groups in Southeastern Brazil. For. Ecol. Manag. 1998, 107, 241–252. [Google Scholar] [CrossRef]
- Fitzsimons, M.S.; Miller, R.M.; Jastrow, J.D. Scale-dependent niche axes of arbuscular mycorrhizal fungi. Oecologia 2008, 158, 117–127. [Google Scholar] [CrossRef]
- Cofré, M.N.; Soteras, F.; Iglesias, M.R.; Velázquez, S.; Abarca, C.; Risio, L.; Ontivero, E.; Cabello, M.N.; Domínguez, L.S.; Lugo, M.A. Biodiversity of Arbuscular Mycorrhizal Fungi in South America: A Review. In Mycorrhizal Fungi in South America; Pagano, M.C., Lugo, M.A., Eds.; Springer Nature: Basel, Switzerland, 2019; pp. 49–72. [Google Scholar]
- Freitas, R.D.O.; Buscardo, E.; Nagy, L.; Maciel, A.B.S.; Carrenho, R.; Luizão, R.C.C. Arbuscular mycorrhizal fungal communities along a pedo-hydrological gradient in a Central Amazonian terra firme forest. Mycorrhiza 2014, 24, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Leal, P.C.L.; Stürmer, S.L.; Siqueira, J.O. Occurrence and diversity of arbuscular mycorrhizal fungi in trap cultures from soils under different land use systems in the Amazon, Brazil. Braz. J. Microbiol. 2009, 40, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Caproni, A.L.; Granha, J.R.D.O.; Fornaciari, A.J.; Nobre, C.P.; Mendonça, L.P.; Berbara, R.L.L. Diversity of arbuscular mycorrhizal fungi in an Amazon environment after mining. Floresta E Ambiente 2018, 25, e20150224. [Google Scholar] [CrossRef]
- Bhatia, N.; Sundari, K.; Adholeya, A. Diversity and selective dominance of vesicular-arbuscular mycorrhizal fungi. In Concepts in Mycorrhizal Research; Mukerji, K.G., Ed.; Springer: Amsterdam, The Netherlands, 1996; p. 374. [Google Scholar]
- Carrenho, R.; Trufem, S.F.B.; Bononi, V.L.R.; Silva, E.S. The effect of different soil properties on arbuscular mycorrhizal colonization of peanuts, sorghum and maize. Acta Bot. Bras. 2007, 21, 723–730. [Google Scholar] [CrossRef]
- Okiobé, S.T.; Angue, M.A.; Bougnom, B.P.; Onana, B.; Nwaga, D. Improvement of arbuscular mycorrhizal fungi inoculum production by nutrient solution concentration and soil texture variation. Int. J. Agron. Agric. Res. 2015, 6, 7–20. [Google Scholar]
- SchüBler, A.; Walker, C. The Glomeromycota: A Species List With New Families and New Genera; The Royal Botanic Garden Edinburgh: Gloucester, UK; The Royal Botanic Garden Kew, Botanische Staatssammlung: Munich, Germany; Oregon State University: Corvallis, OG, USA, 2010; p. 57. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Level | pH | CEC (meq/100 g) | OC (%) | Total N (%) | Ca (mg/kg) | Mg (mg/kg) |
|---|---|---|---|---|---|---|---|
| Landscape | Hill | 4.22 ± 0.11 a | 7.35 ± 0.55 a | 2.29 ± 0.20 a | 0.19 ± 0.02 a | 320.75 ± 51.62 a | 45.17 ± 20.11 a |
| Mountain | 4.04 ± 0.10 a | 5.55 ± 0.46 b | 2.04 ± 0.17 a | 0.17 ± 0.01 a | 236.42 ± 59.78 a | 76.54 ± 21.87 a | |
| Chronosequence | Degraded pasture | 4.26 ± 0.10 a | 4.97 ± 0.49 a | 1.67 ± 0.20 b | 0.14 ± 0.02 b | 236.92 ± 24.79 a | 39.67 ± 3.46 a |
| 10–20 | 4.25 ± 0.26 ab | 7.38 ± 1.06 a | 2.19 ± 0.36 ab | 0.18 ± 0.03 ab | 394.83 ± 154.03 a | 106.25 ± 66.24 a | |
| 25–40 | 4.15 ± 0.08 ab | 7.27 ± 0.49 a | 2.37 ± 0.13 a | 0.20 ± 0.01 a | 253.00 ± 15.68 a | 52.25 ± 5.05 a | |
| Forest | 3.84 ± 0.05 b | 6.19 ± 0.67 a | 2.42 ± 0.29 a | 0.20 ± 0.02 a | 229.58 ± 19.20 a | 45.25 ± 4.96 a | |
| Factor | Level | Na (mg/kg) | P (mg/kg) | K (mg/kg) | Texture | ||
| Loam (%) | Clay (%) | Sand (%) | |||||
| Landscape | Hill | 43.38 ± 2.64 a | 5.69 ± 0.53 a | 58.18 ± 6.55 a | 14.21 ± 2.62 a | 44.29 ± 3.47 a | 41.50 ± 4.18 b |
| Mountain | 45.04 ± 2.04 a | 5.32 ± 0.43 a | 71.21 ± 5.58 a | 13.67 ± 2.19 a | 21.50 ± 2.84 b | 64.83 ± 3.50 a | |
| Chronosequence | Degraded pasture | 45.25 ± 2.64 a | 4.86 ± 0.74 a | 47.76 ± 4.44 b | 7.75 ± 0.77 b | 28.75 ± 6.99 a | 63.50 ± 6.64 a |
| 10–20 | 44.58 ± 1.72 a | 6.01 ± 0.76 a | 75.13 ± 10.96 a | 16.17 ± 4.53 ab | 32.50 ± 2.96 a | 51.33 ± 5.86 a | |
| 25–40 | 44.17 ± 2.51 a | 5.06 ± 0.42 a | 72.49 ± 8.89 a | 16.33 ± 3.08 a | 35.00 ± 2.83 a | 48.67 ± 4.42 a | |
| Forest | 45.83 ± 5.33 a | 6.09 ± 0.76 a | 63.38 ± 8.82 ab | 15.50 ± 4.00 ab | 35.33 ± 3.84 a | 49.17 ± 4.59 a | |
| Factor | Level | Richness | Exponential Shannon (expH’) | Inverse Simpson 1/D | Equitability (Evenness-Piélou) |
|---|---|---|---|---|---|
| Landscape | Hill | 7.70 ± 1.04 a | 3.15 ± 0.573 a | 2.55 ± 0.41 a | 0.49 ± 0.07 a |
| Mountain | 6.50 ± 0.96 a | 2.45 ± 0.46 a | 1.94 ± 0.33 a | 0.32 ± 0.06 a | |
| Chronosequence | Degraded pasture | 12.12 ± 2.07 a | 3.43 ± 1.06 a | 2.50 ± 0.69 a | 0.53 ± 0.09 a |
| 10–20 | 6.60 ± 1.22 b | 3.57 ± 0.73 a | 2.89 ± 0.58 a | 0.46 ± 0.07 a | |
| 25–40 | 4.83 ± 0.52 b | 2.33 ± 0.37 a | 2.05 ± 0.29 a | 0.34 ± 0.12 a | |
| Forest | 4.83 ± 1.04 b | 1.88 ± 0.62 a | 1.54 ± 0.46 a | 0.29 ± 0.09 a |
| Factor | Level | Virtual Taxon | Identity | Indicator Value | Probability |
|---|---|---|---|---|---|
| Landscape | Mountain | VT444 | Paraglomus sp. | 0.9225 | 0.002 |
| Hill | VT70 | Glomus sp. | 0.2480 | 0.029 | |
| Chronosecuence | Degraded pasture | VT89 | Glomus sp. | 0.4239 | 0.011 |
| VT126 | Glomus sp. | 0.4095 | 0.030 | ||
| VT4 | Archaeospora sp. | 0.3326 | 0.010 | ||
| LH.Cl01 | Glomus sp. | 0.2914 | 0.018 | ||
| VT93 | Glomus sp. | 0.2556 | 0.014 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-León, C.H.; Peña-Venegas, C.P.; Sterling, A.; Muñoz-Ramirez, H.; Virguez-Díaz, Y.R. Changes in Soil-Borne Communities of Arbuscular Mycorrhizal Fungi during Natural Regrowth of Abandoned Cattle Pastures Are Indicative of Ecosystem Restoration. Agronomy 2021, 11, 2468. https://doi.org/10.3390/agronomy11122468
Rodríguez-León CH, Peña-Venegas CP, Sterling A, Muñoz-Ramirez H, Virguez-Díaz YR. Changes in Soil-Borne Communities of Arbuscular Mycorrhizal Fungi during Natural Regrowth of Abandoned Cattle Pastures Are Indicative of Ecosystem Restoration. Agronomy. 2021; 11(12):2468. https://doi.org/10.3390/agronomy11122468
Chicago/Turabian StyleRodríguez-León, Carlos H., Clara P. Peña-Venegas, Armando Sterling, Herminton Muñoz-Ramirez, and Yeny R. Virguez-Díaz. 2021. "Changes in Soil-Borne Communities of Arbuscular Mycorrhizal Fungi during Natural Regrowth of Abandoned Cattle Pastures Are Indicative of Ecosystem Restoration" Agronomy 11, no. 12: 2468. https://doi.org/10.3390/agronomy11122468
APA StyleRodríguez-León, C. H., Peña-Venegas, C. P., Sterling, A., Muñoz-Ramirez, H., & Virguez-Díaz, Y. R. (2021). Changes in Soil-Borne Communities of Arbuscular Mycorrhizal Fungi during Natural Regrowth of Abandoned Cattle Pastures Are Indicative of Ecosystem Restoration. Agronomy, 11(12), 2468. https://doi.org/10.3390/agronomy11122468