
In Silico Characterization and Expression Profiles of Heat Shock Transcription Factors (HSFs) in Maize (Zea mays L.)


 ,
,  , , ,
, , ,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sequence Retrieval
2.2. Sequence Analysis
2.3. Sequence Alignment and Phylogenetic Analysis
2.4. Gene Duplication and Evolutionary Analysis
2.5. Chromosomal Distribution
2.6. Expression Profiling of HSF Genes
2.7. Protein 3D Structure, Network Interaction, and Gene Ontology Analysis
3. Results
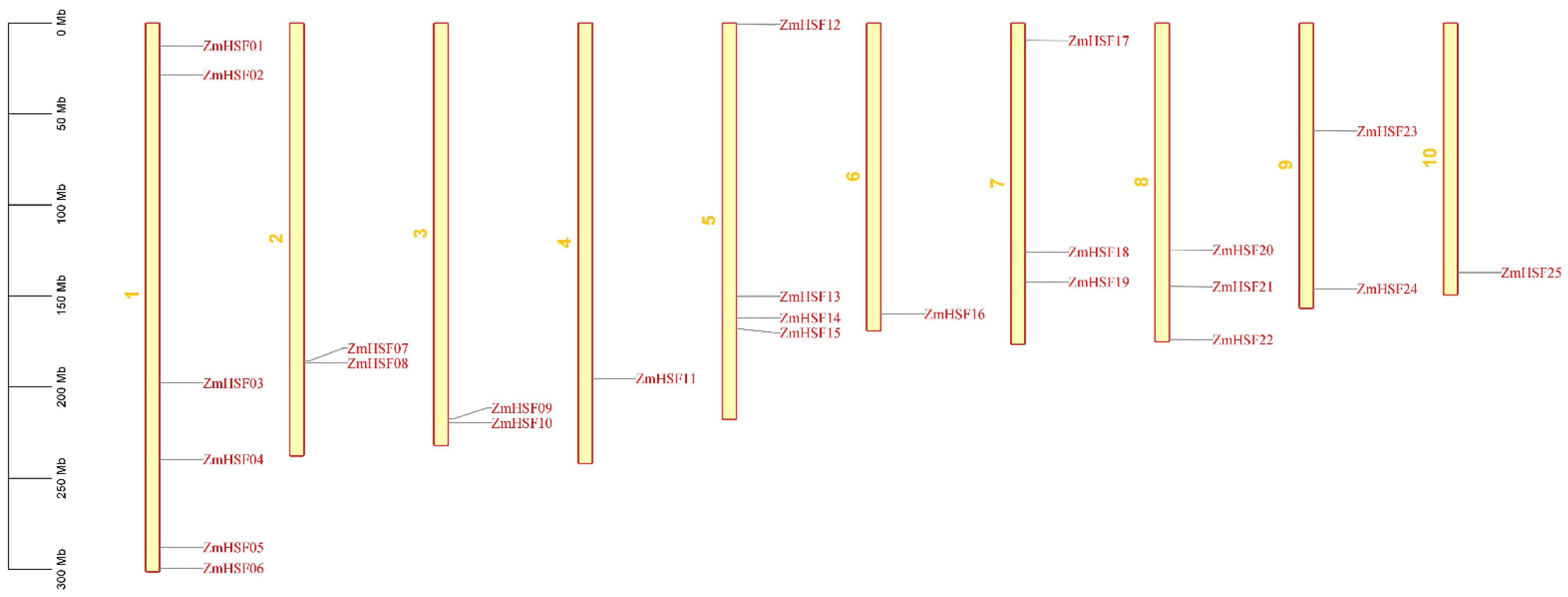
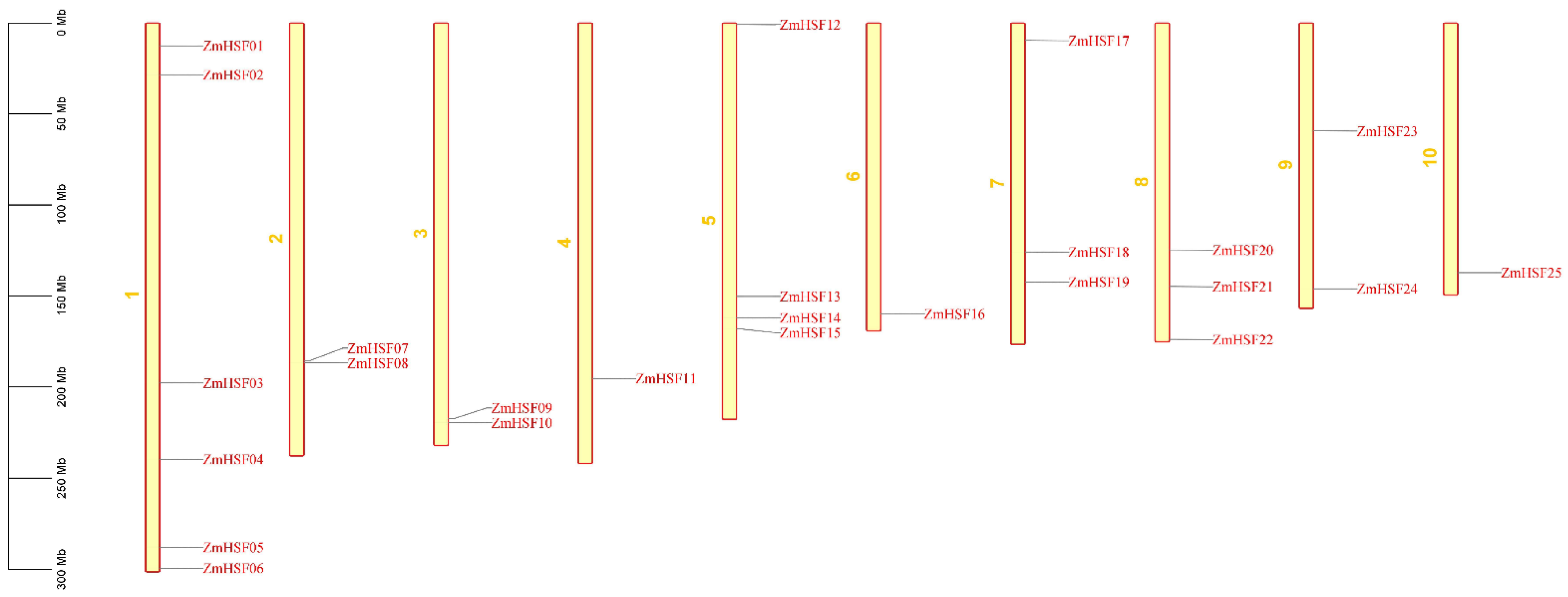
3.1. Identification and Chromosomal Distribution of Maize HSFs
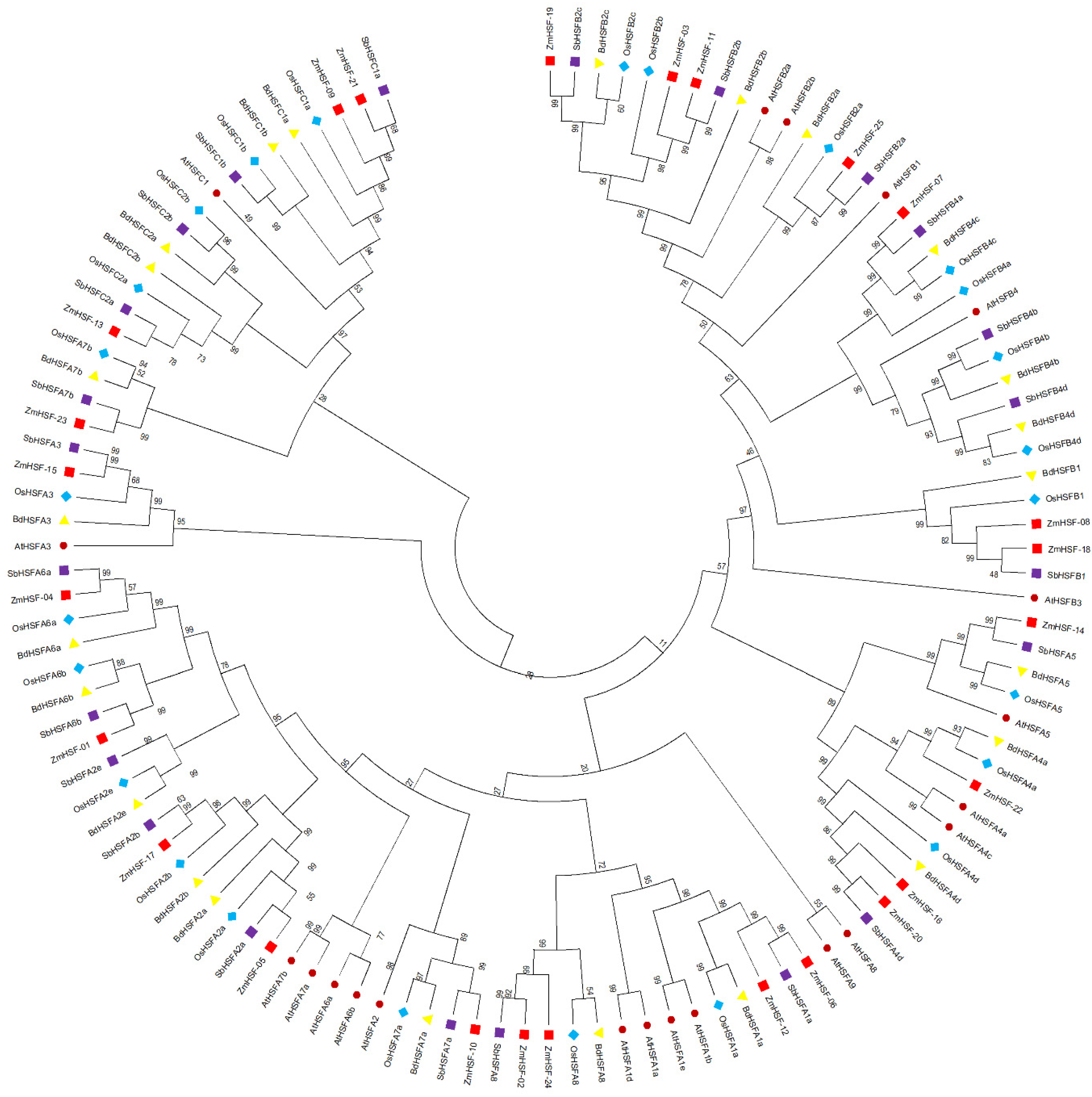
3.2. Phylogenetic Analysis and Classification of Maize HSFs
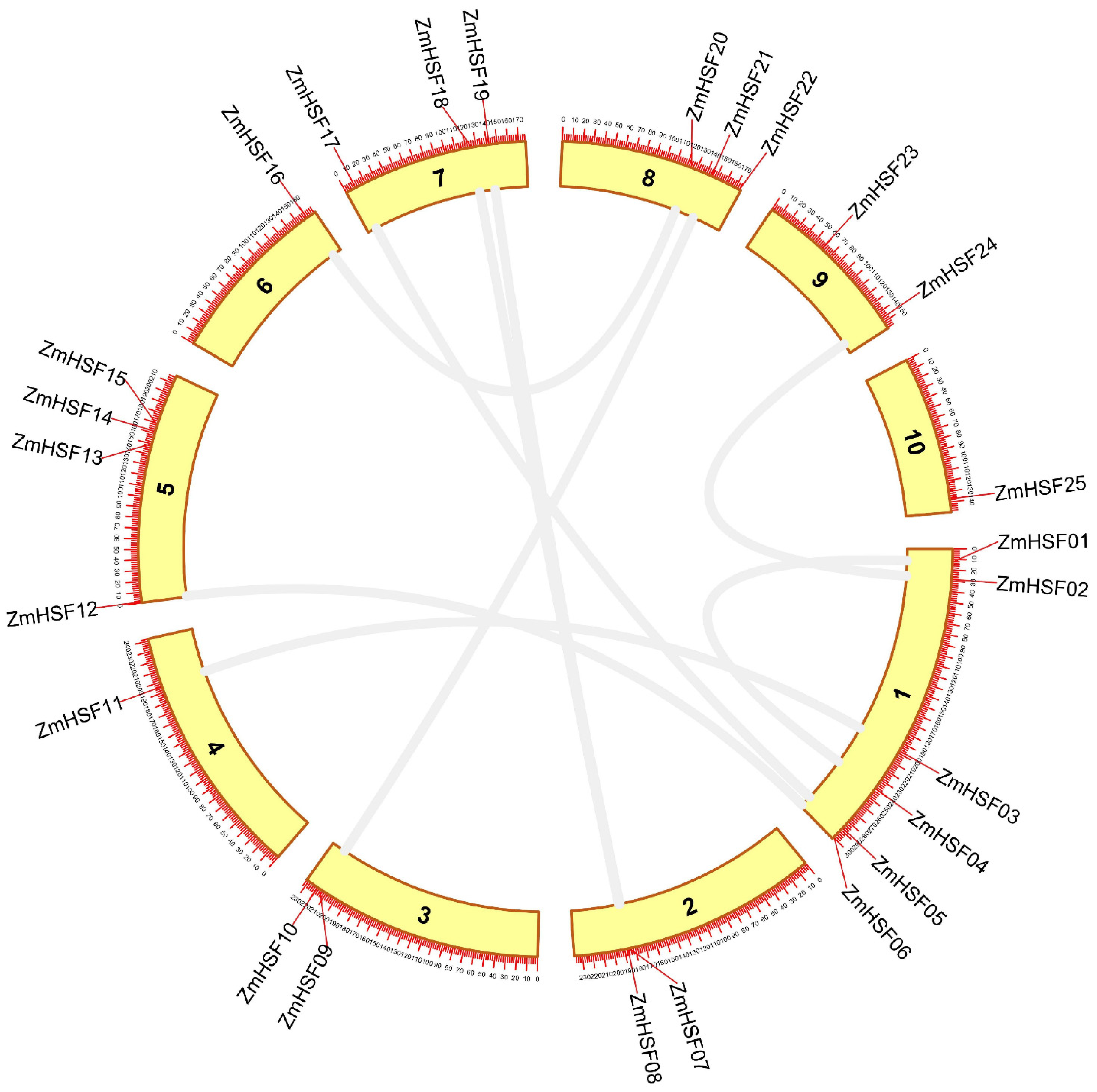
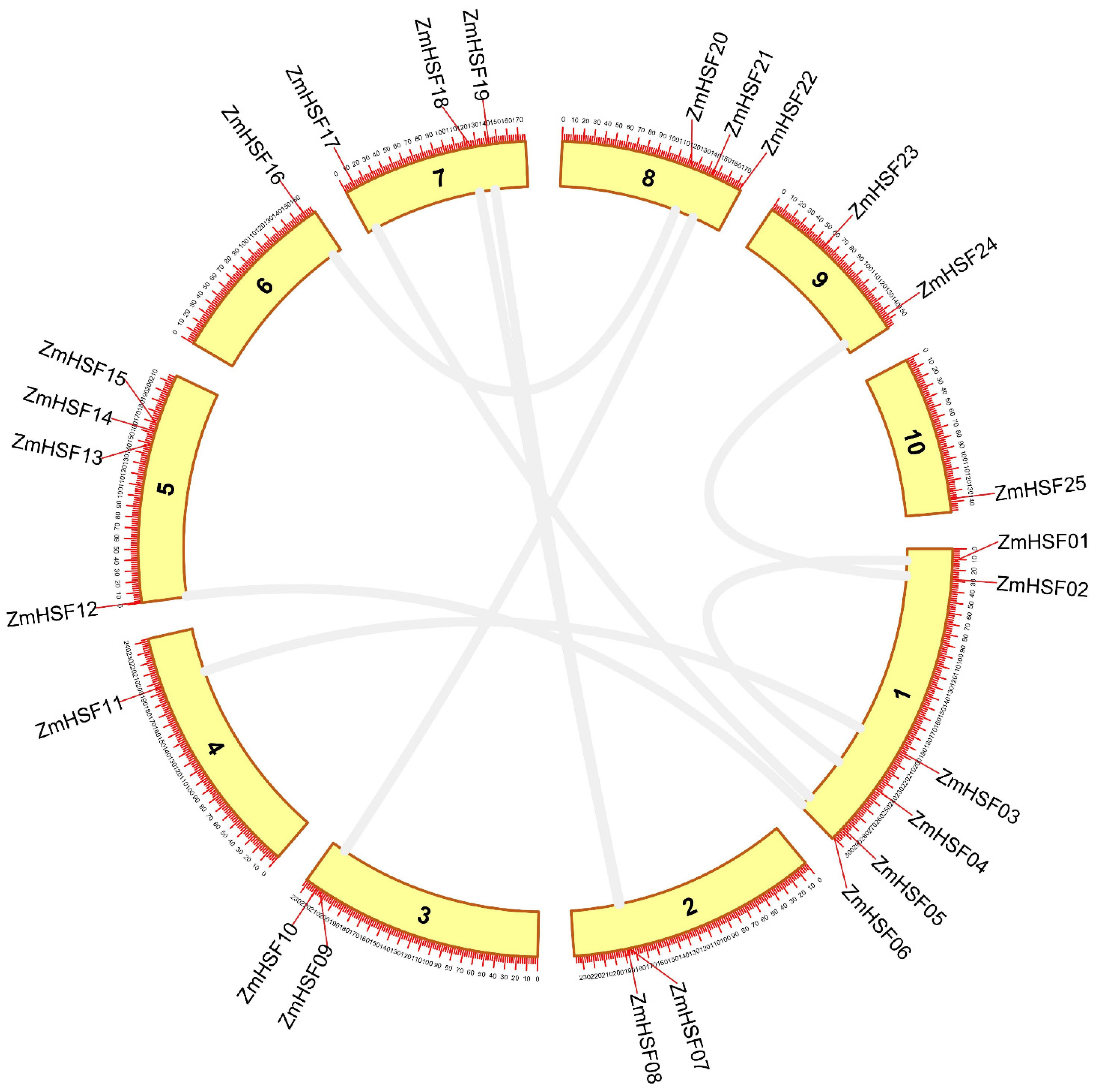
3.3. Gene Duplication Analysis and Evolutionary Rate Calculation
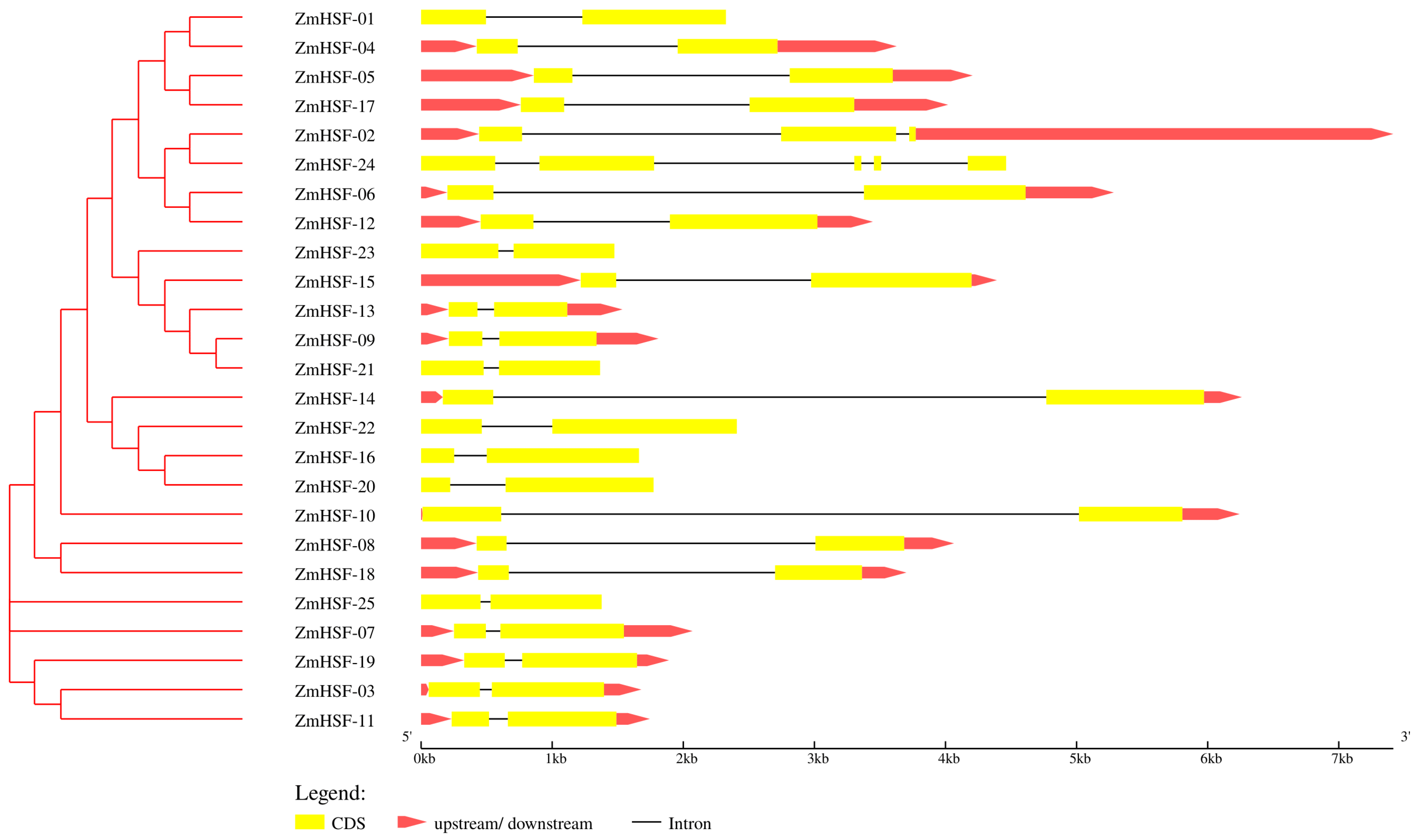
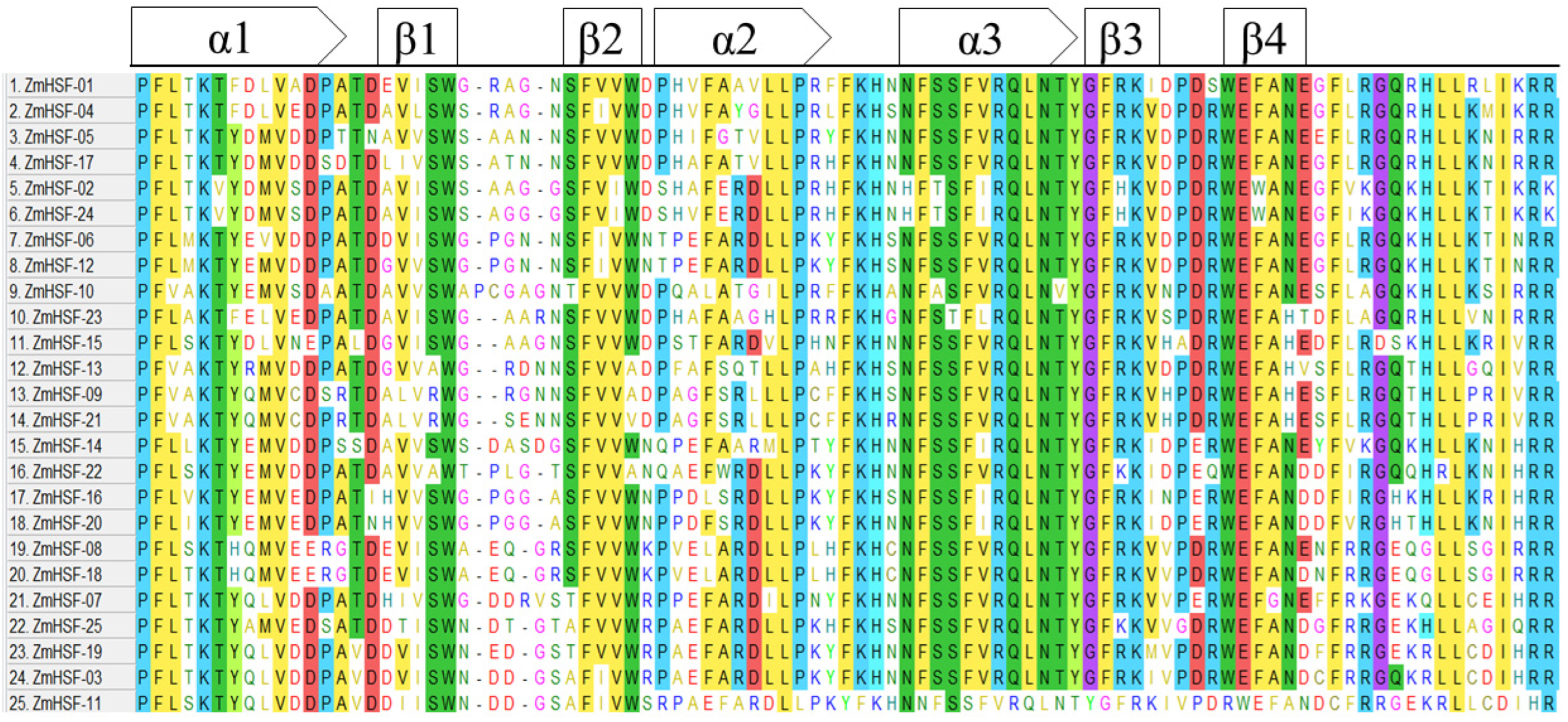
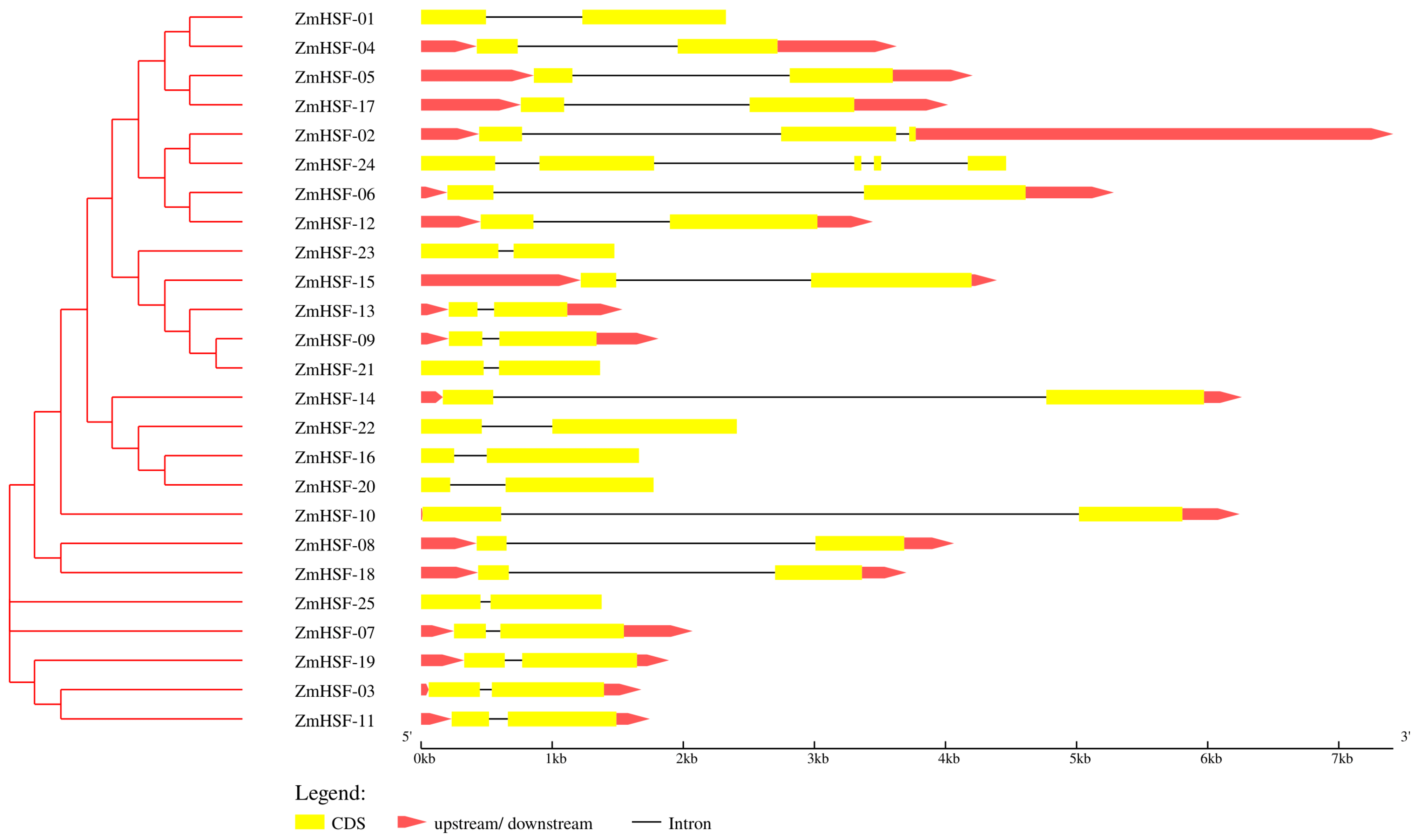
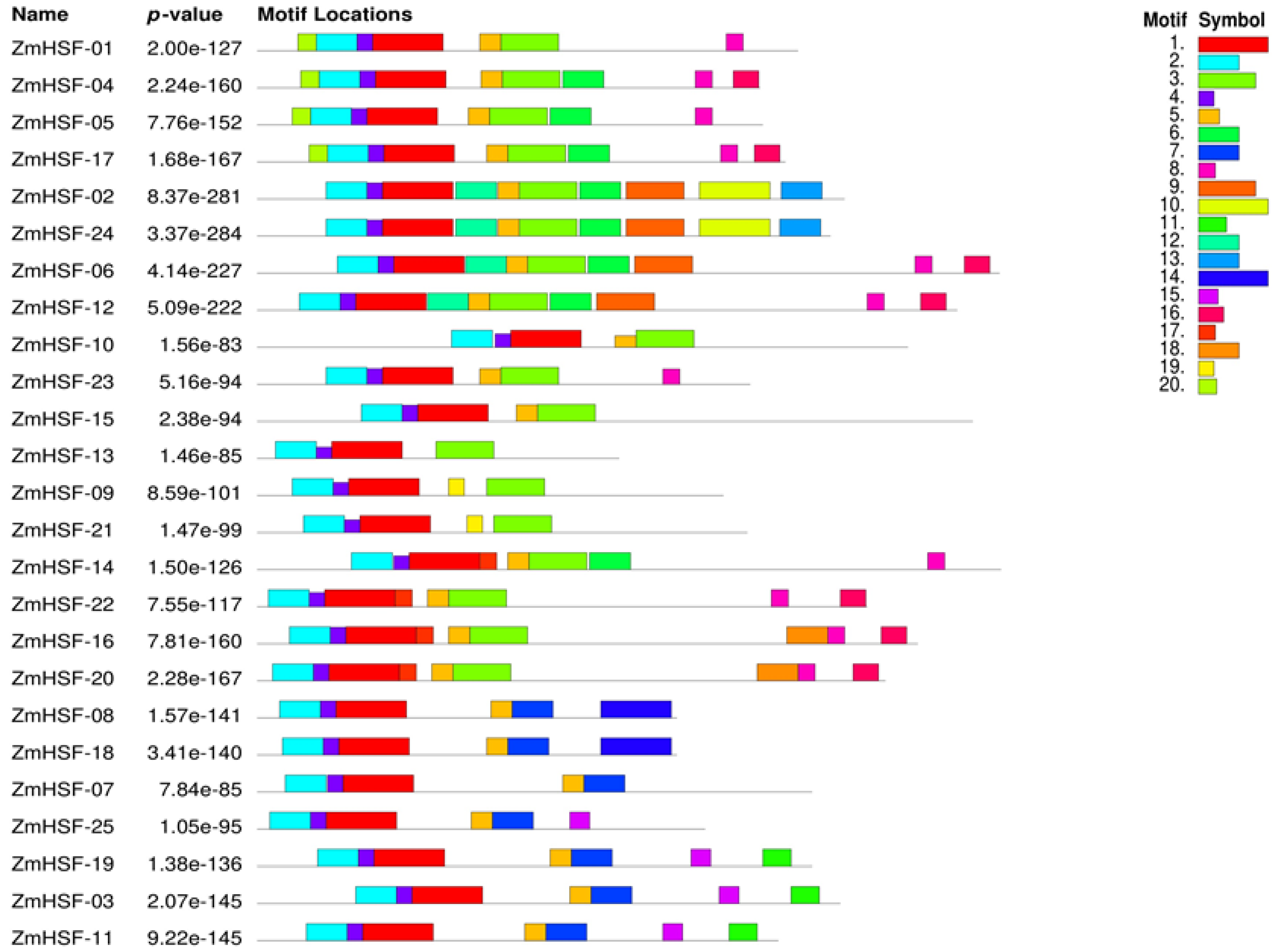
3.4. Gene Structure and Protein Motif Analysis
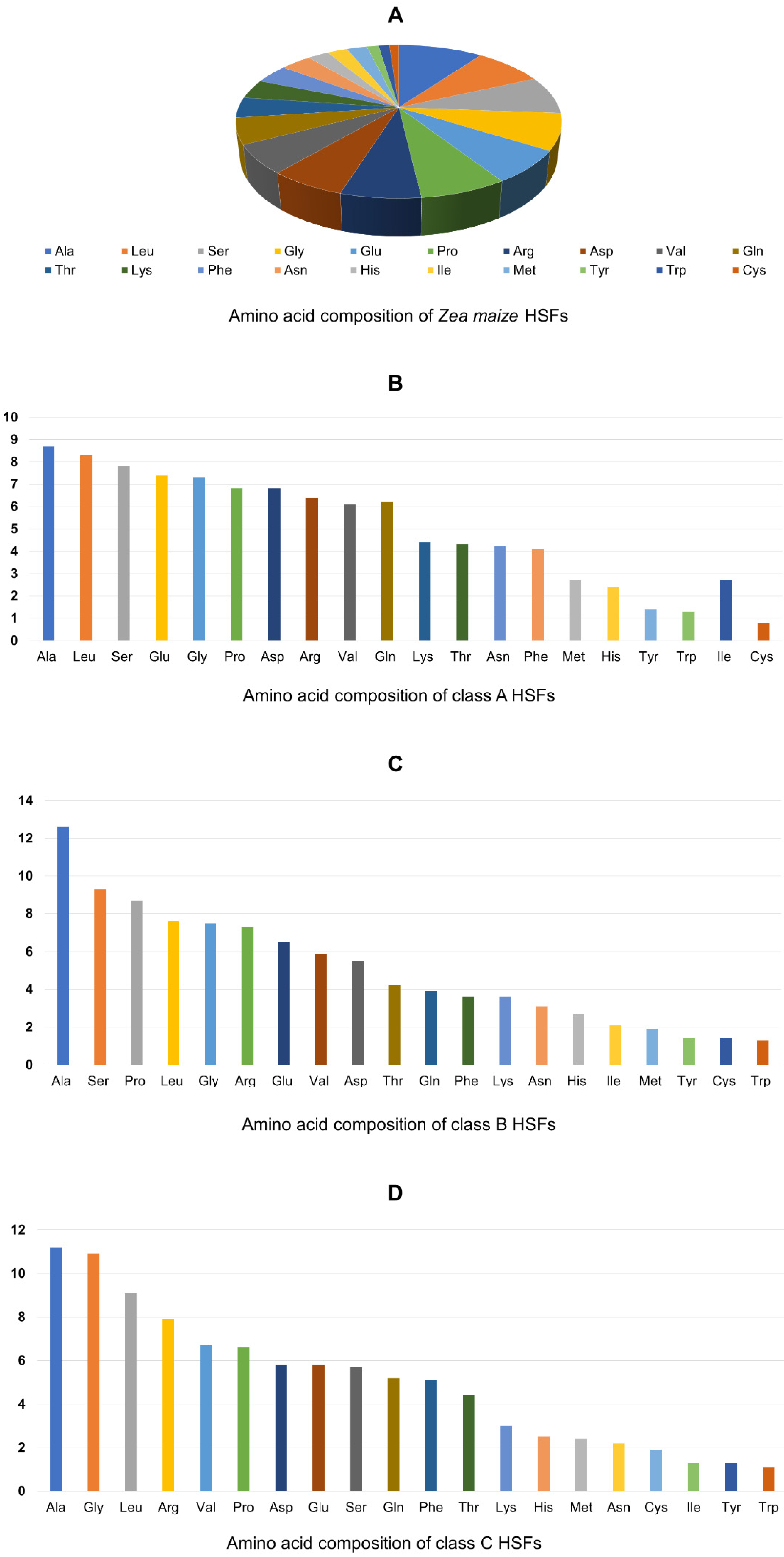
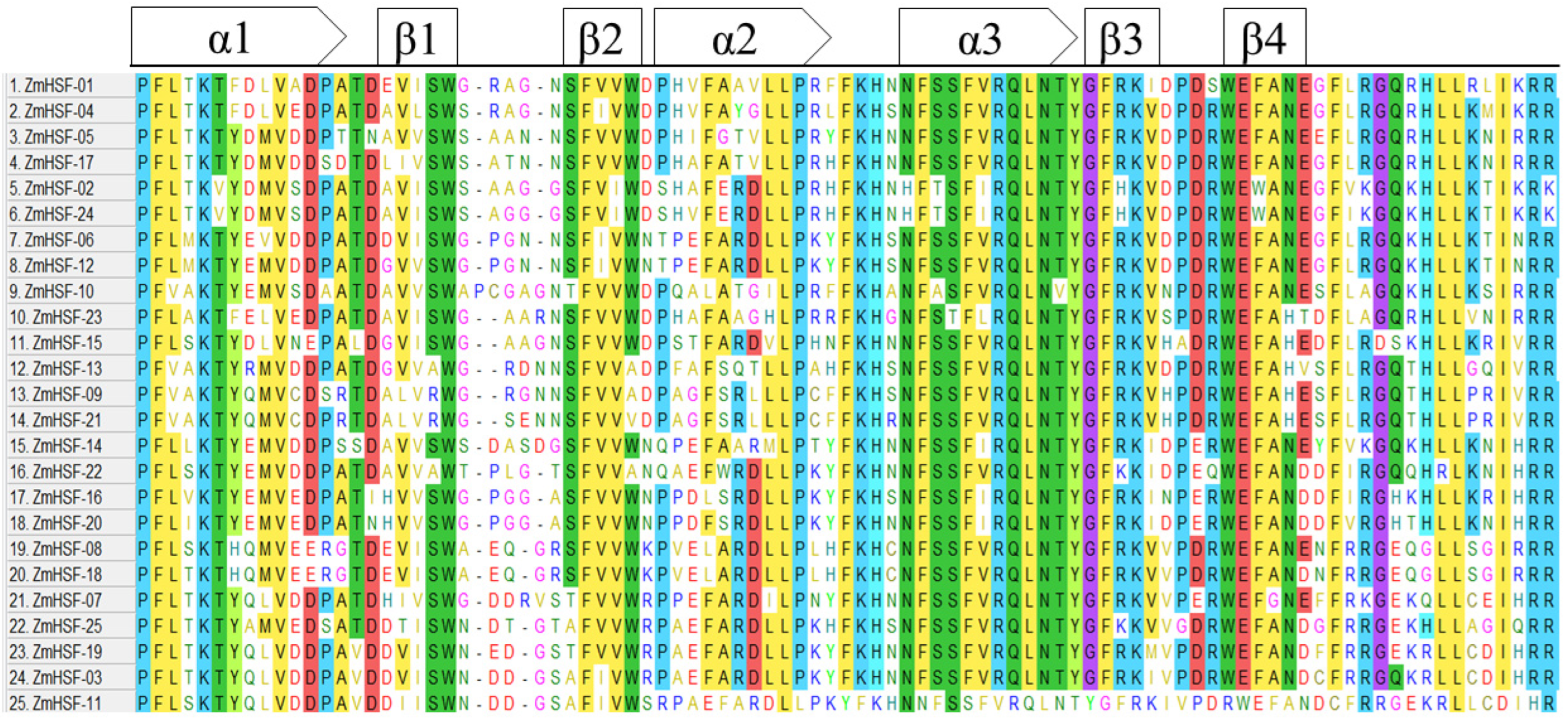
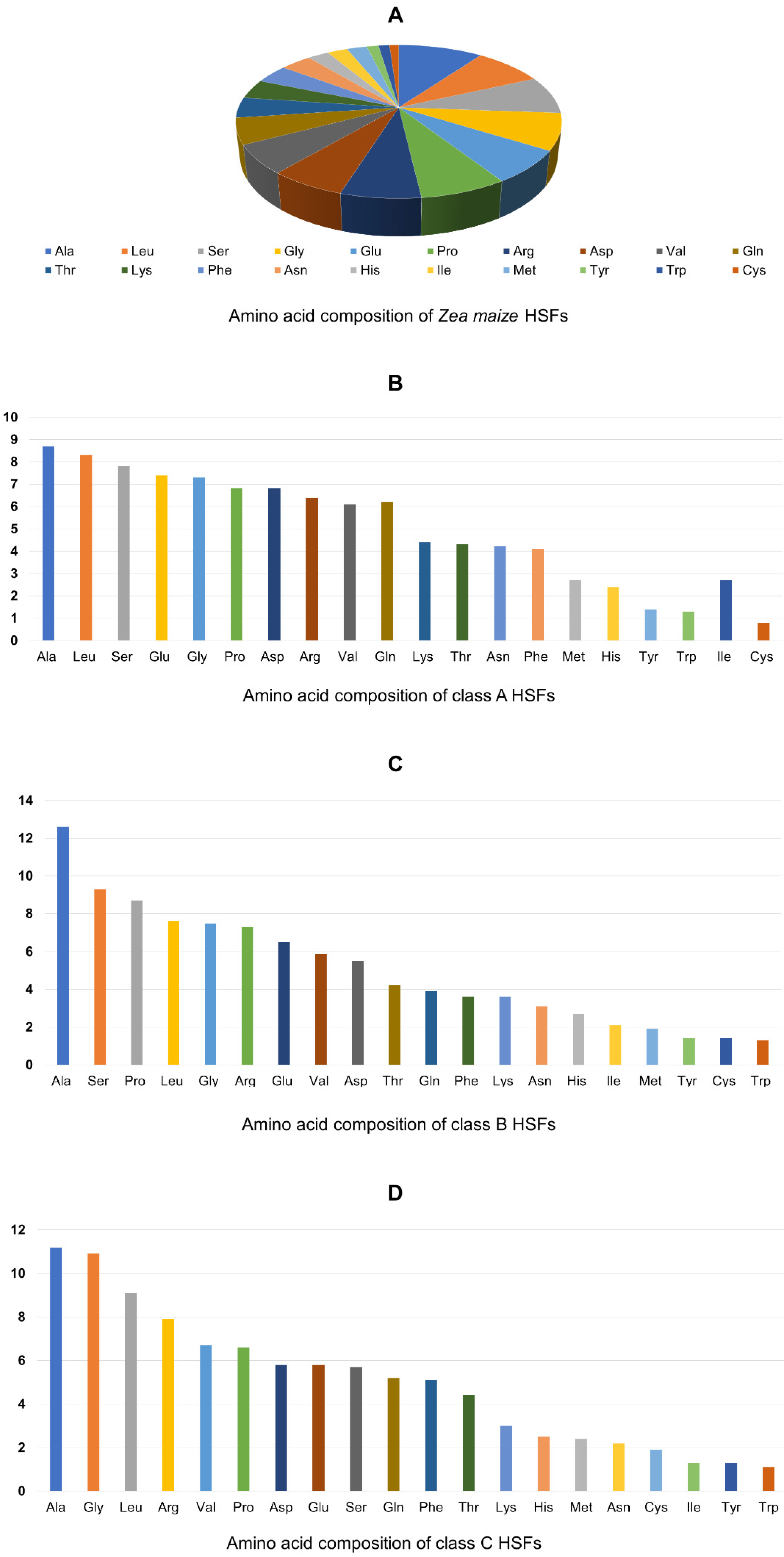
3.5. Domain Analysis and Physio-Chemical Properties
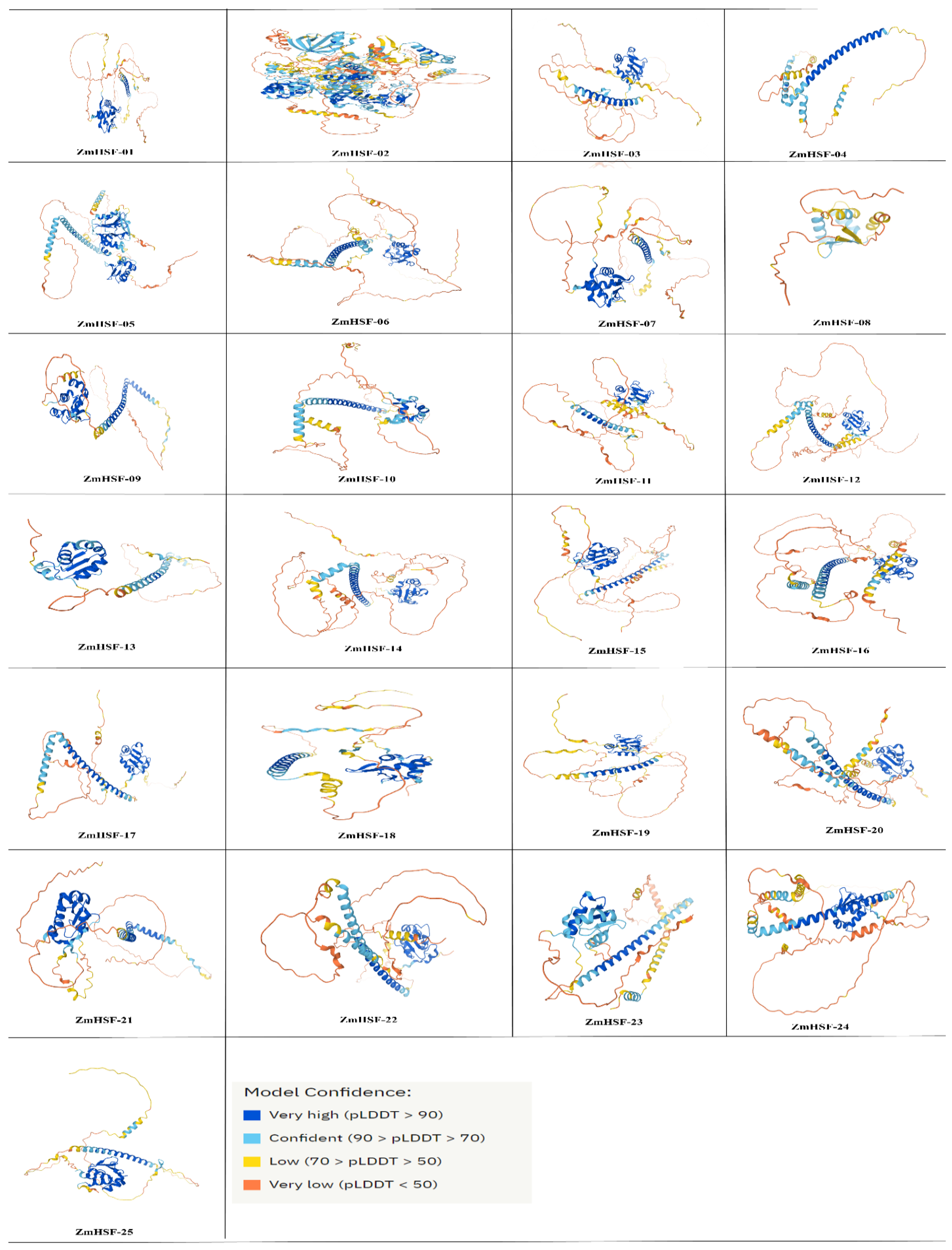
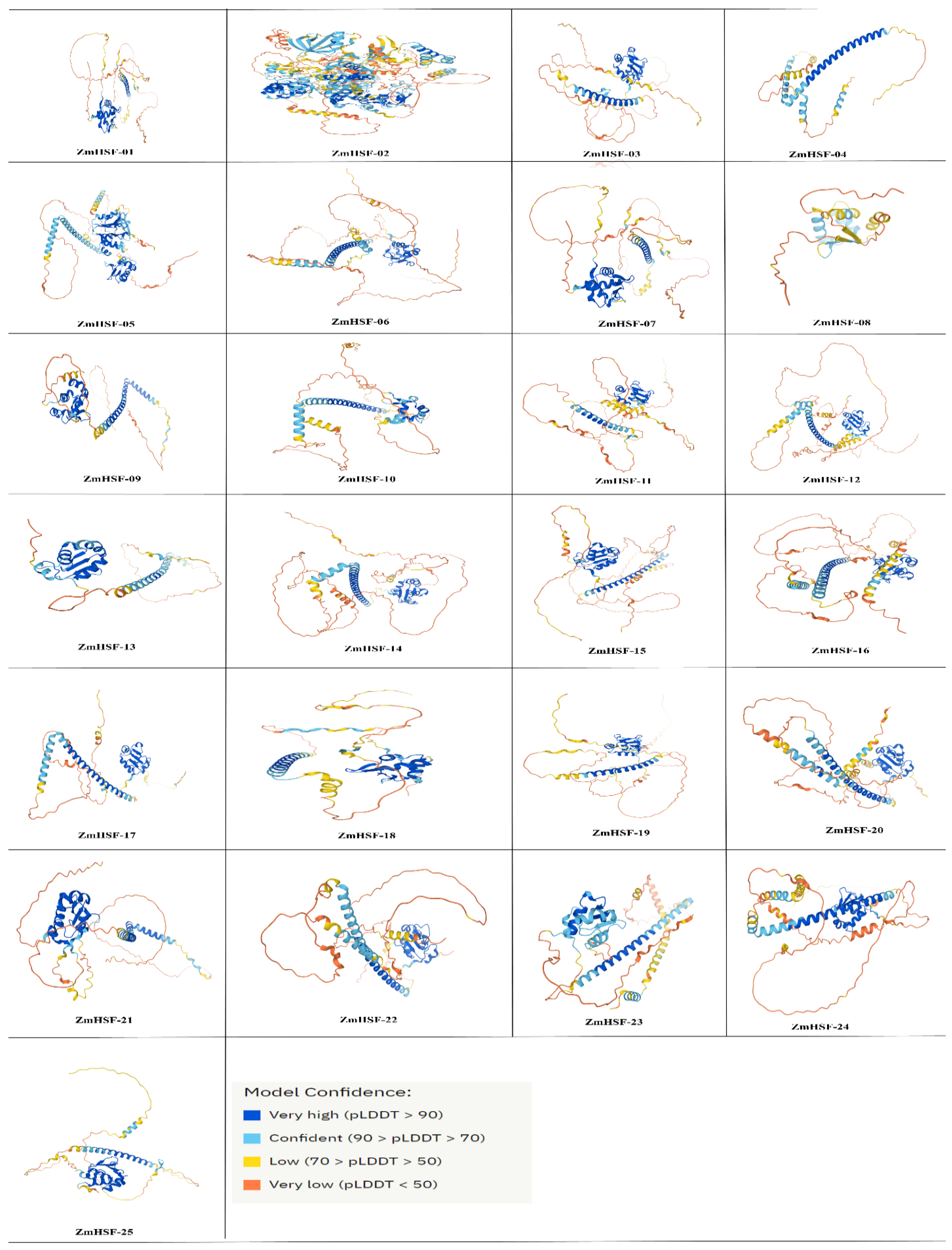
3.6. Proteins Structure and Sub-Cellular Localization of Maize HSFs
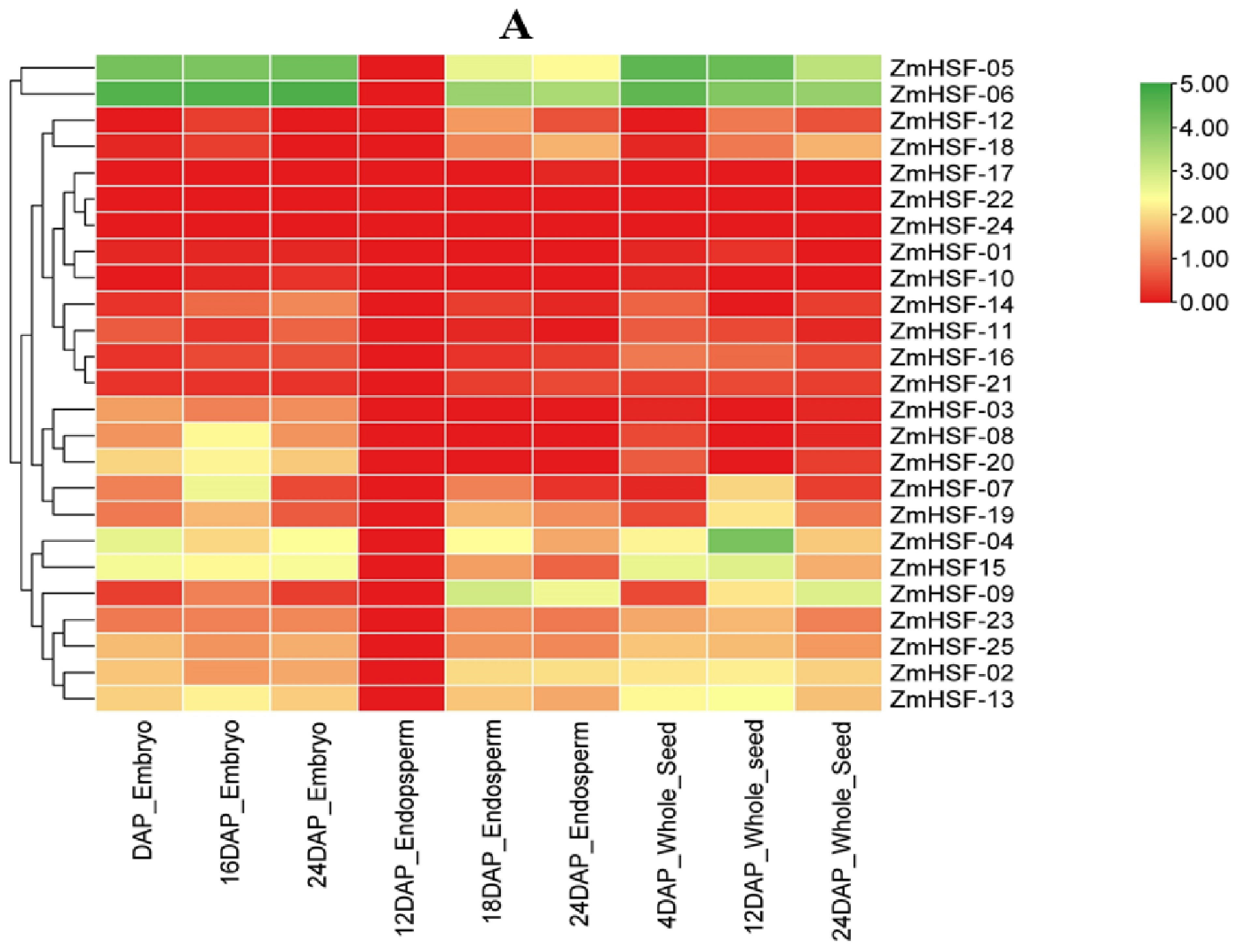
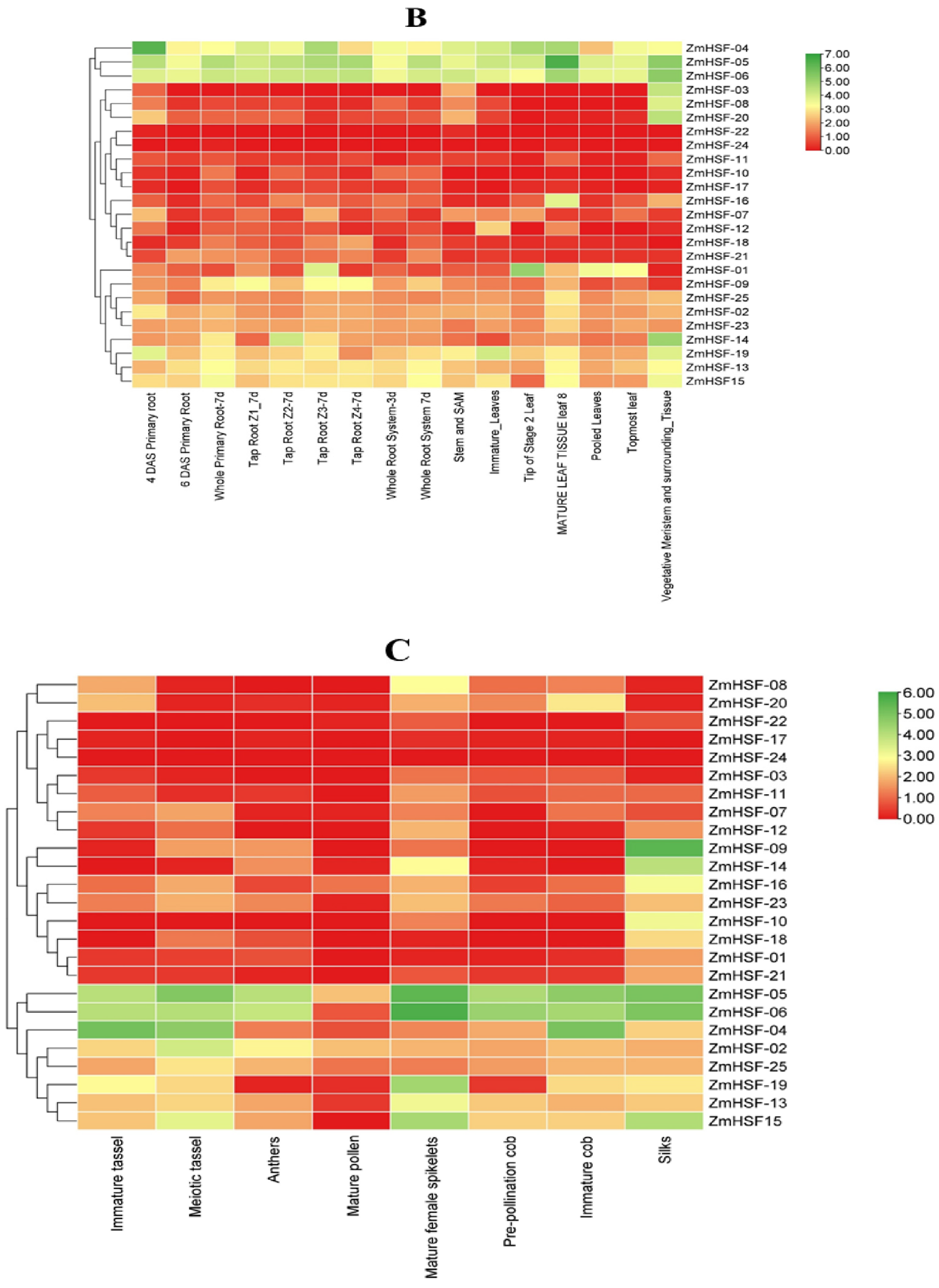
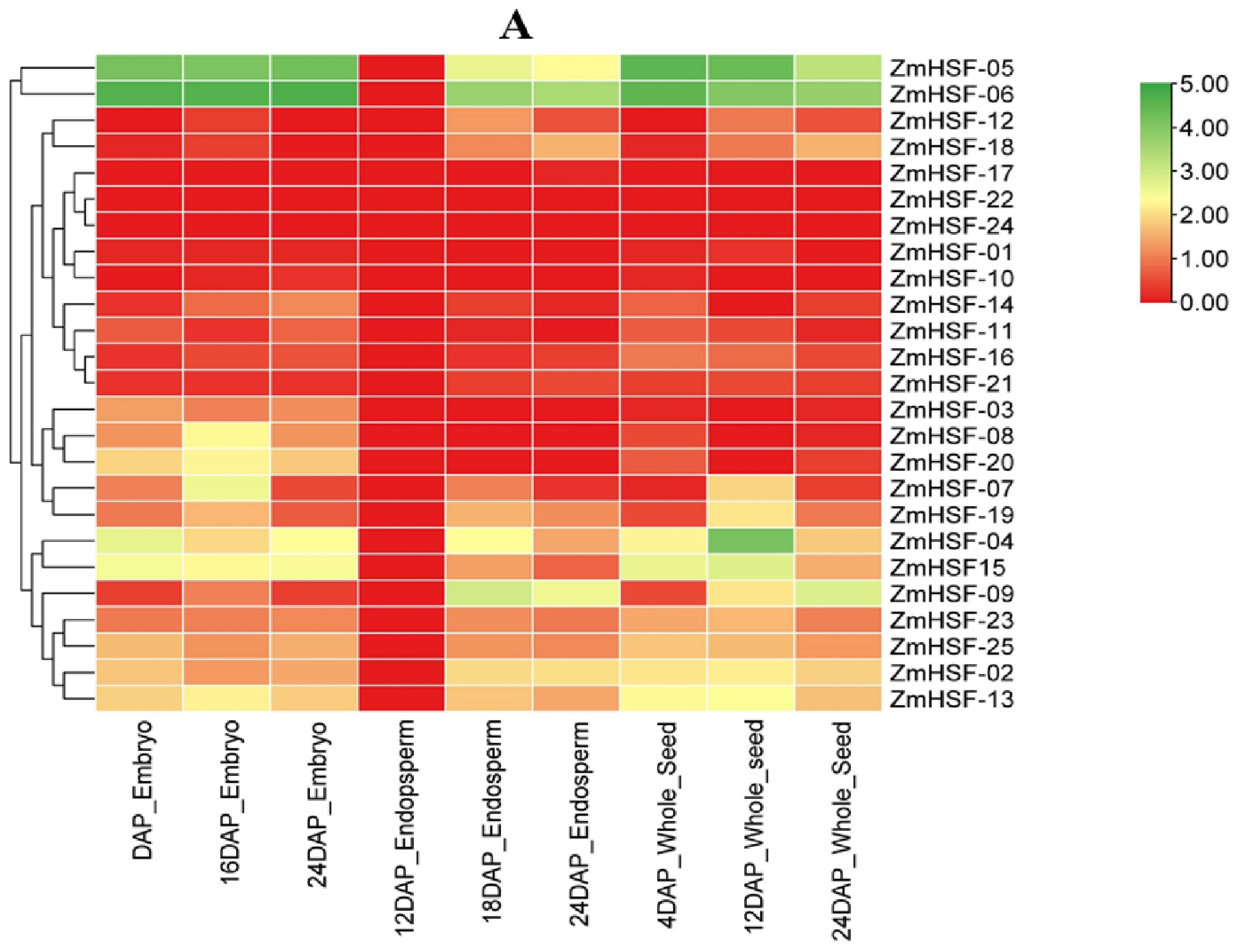
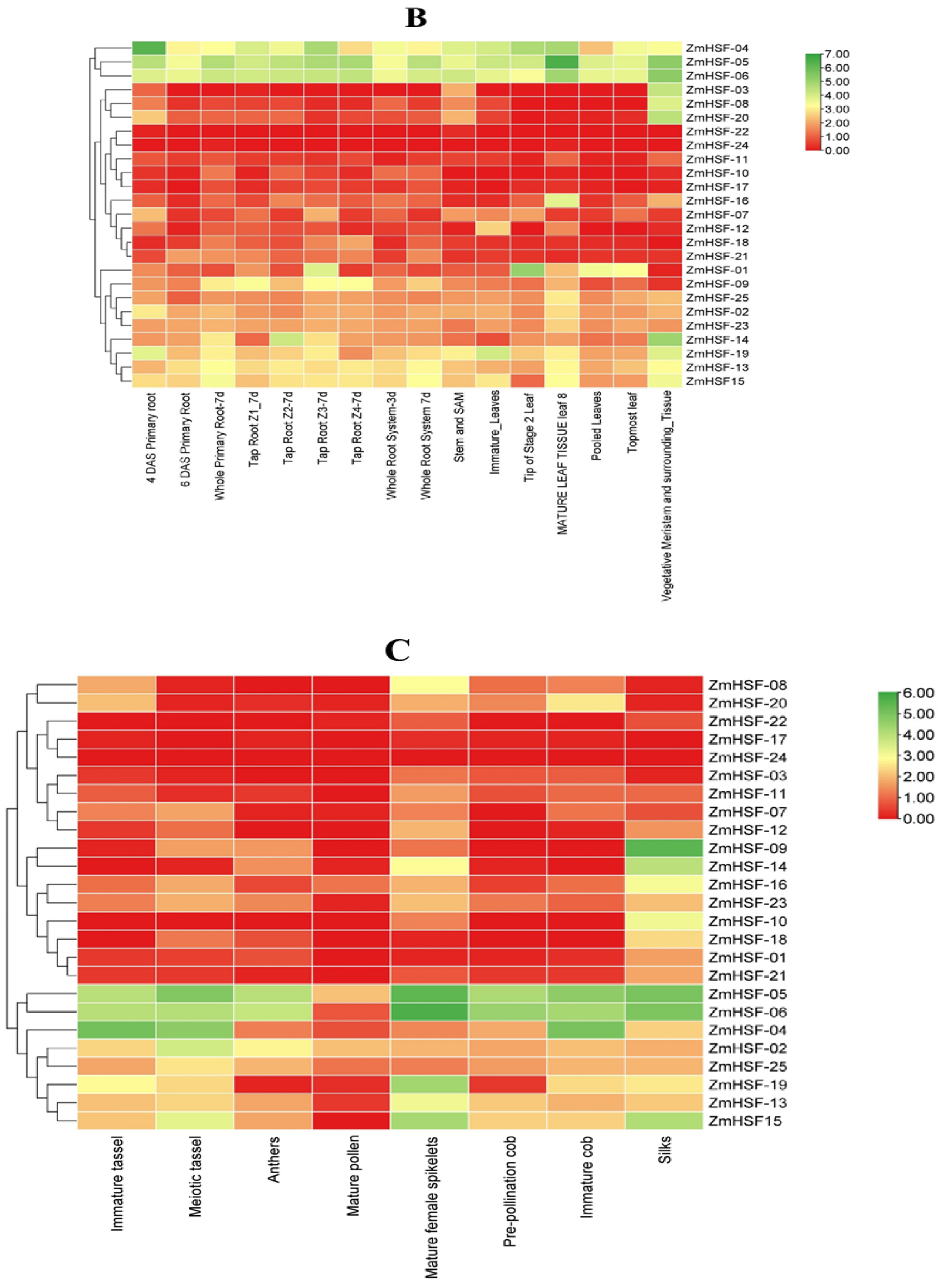
3.7. Expression Profiles of Zea mays HSFs during Different Developmental Stages
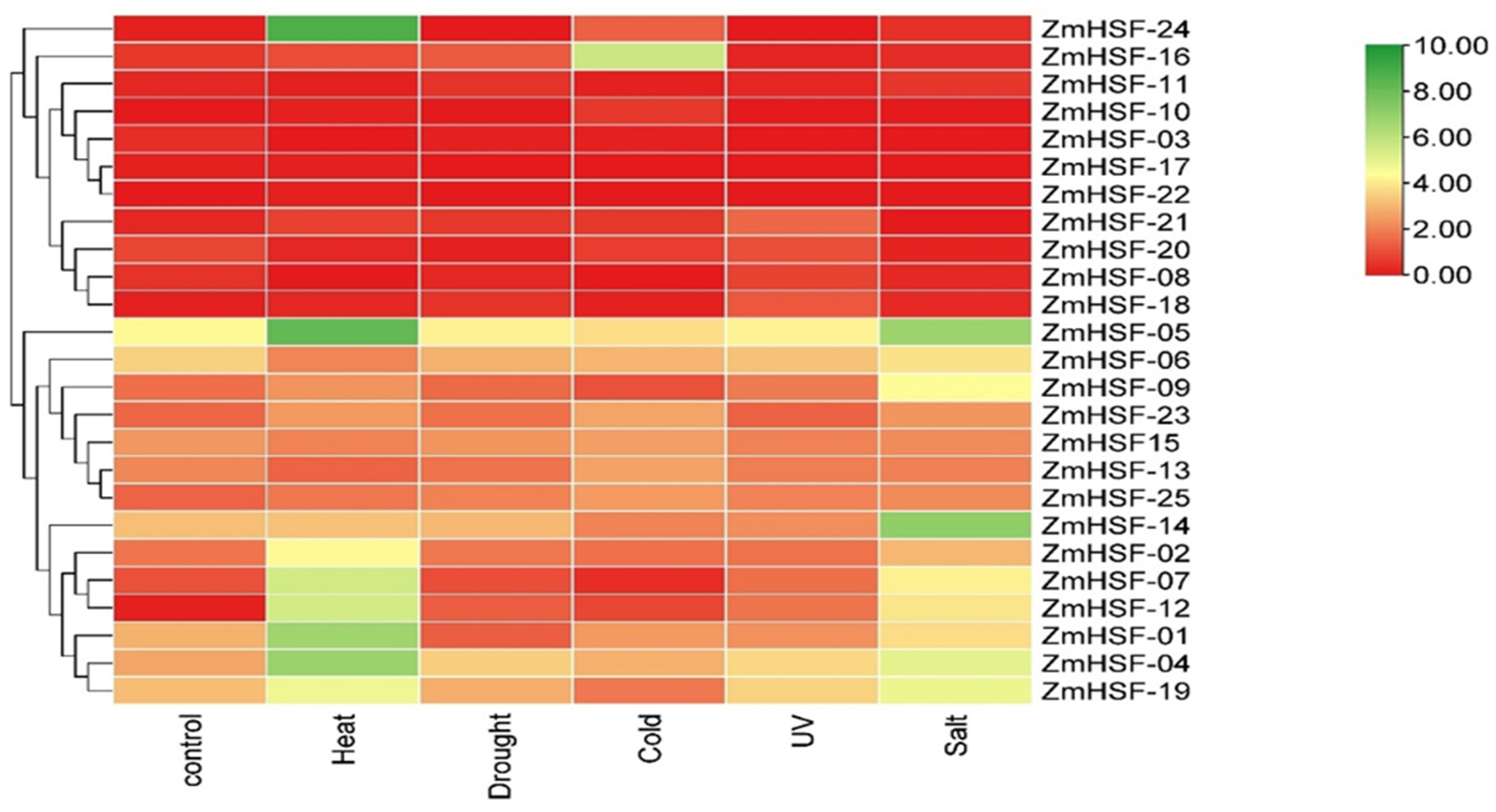
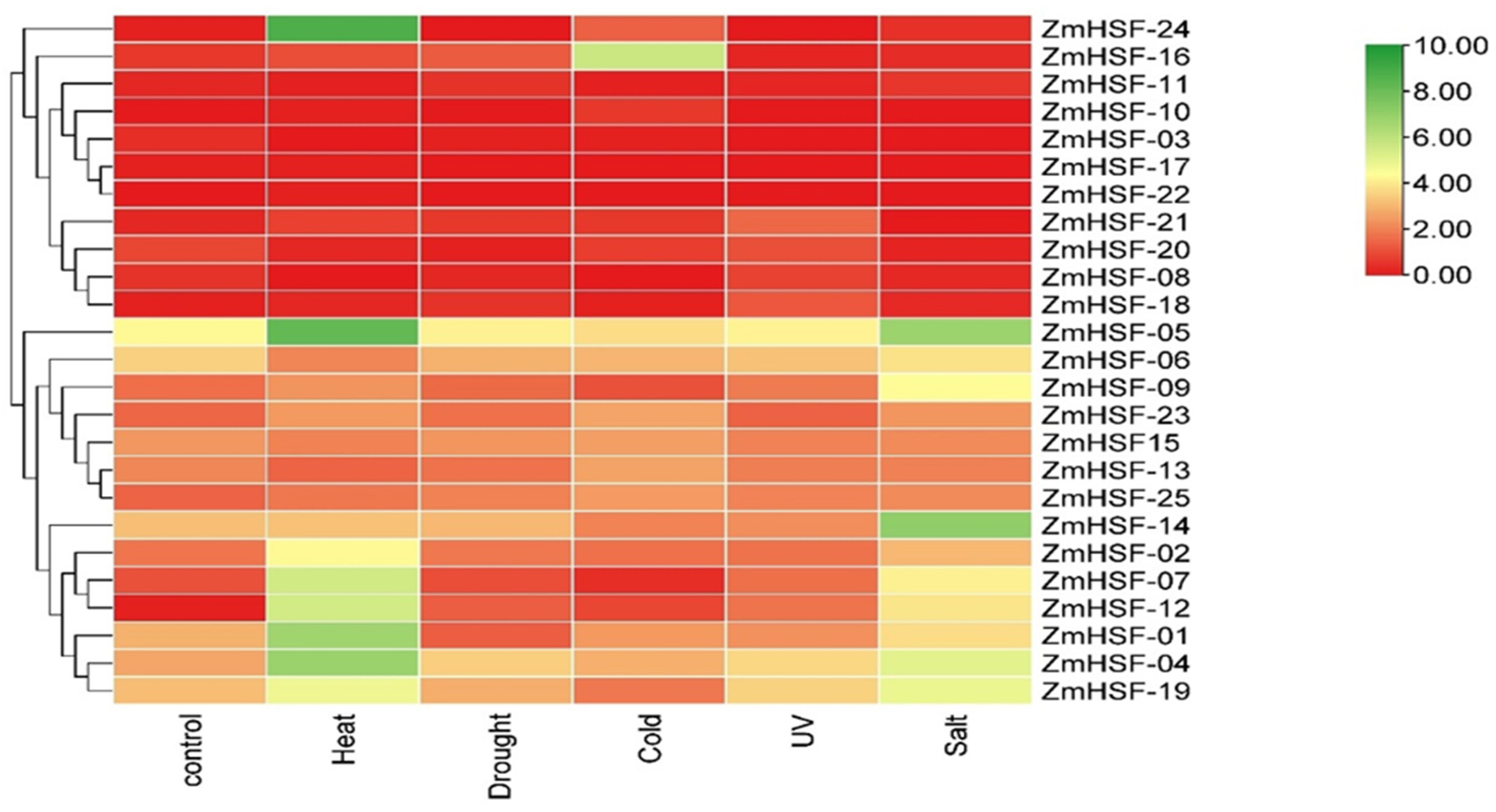
3.8. Expression Pattern of Zea mays HSFs under Abiotic Stresses
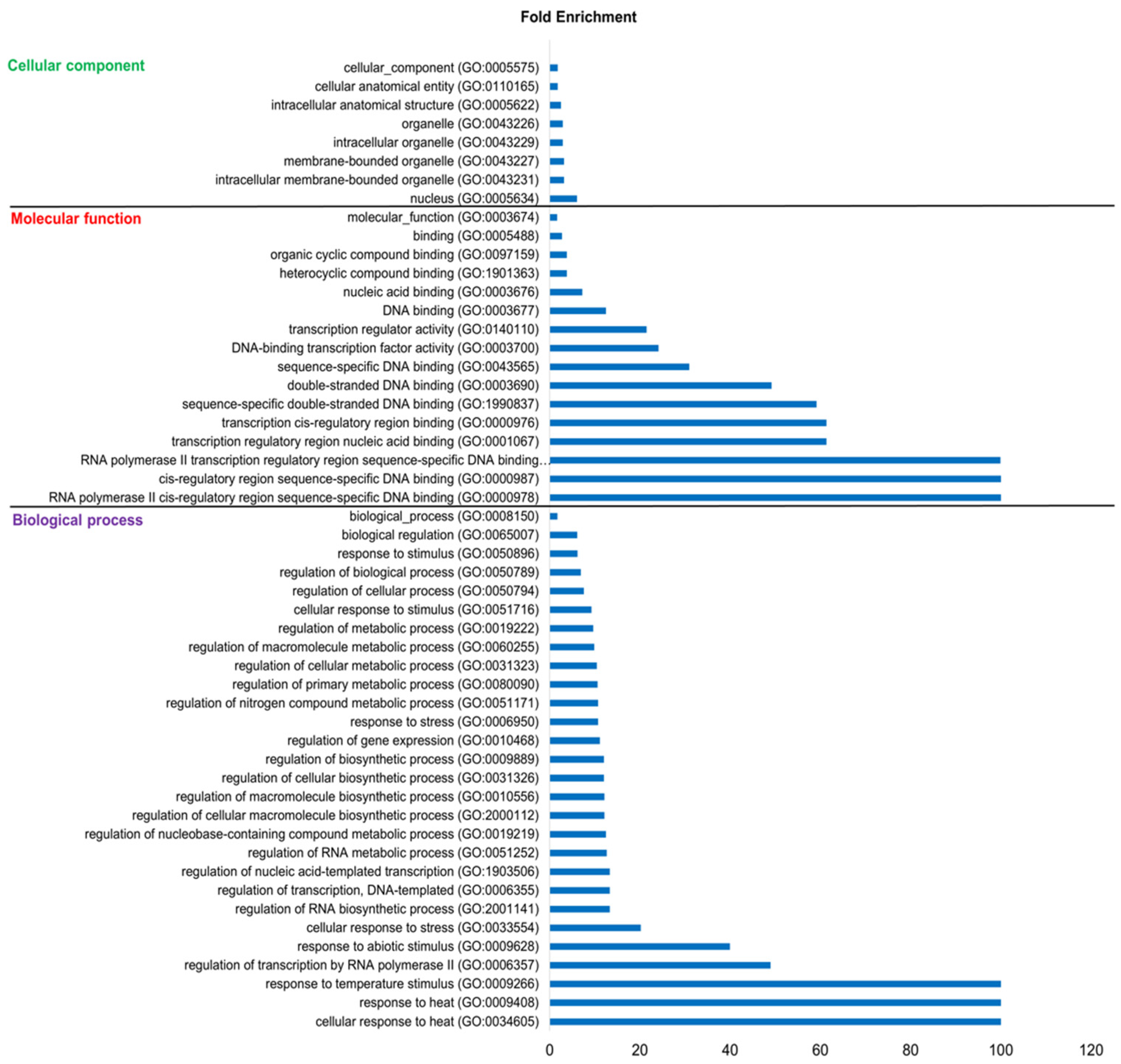
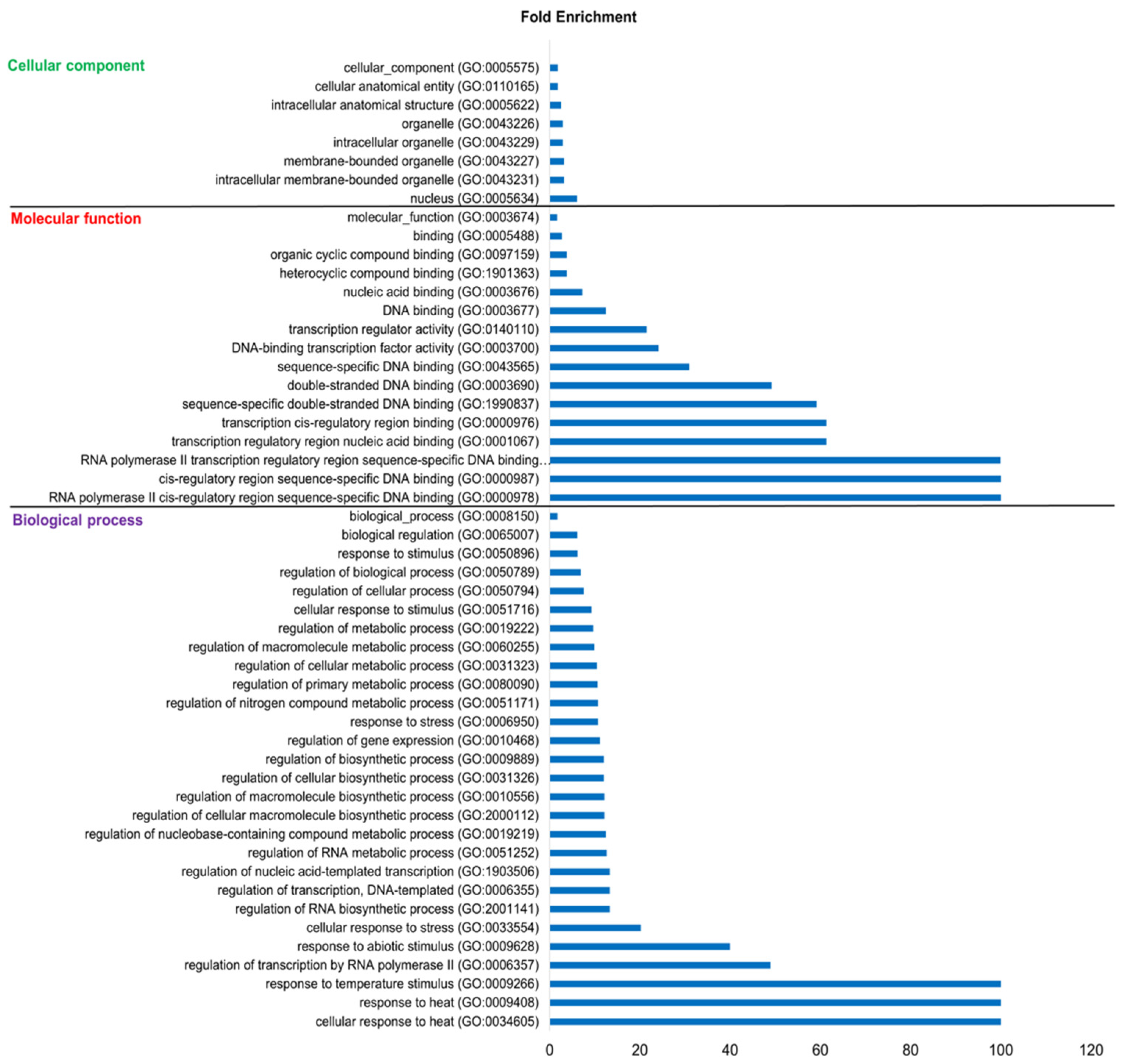
3.9. Functional Annotation of Maize HSFs
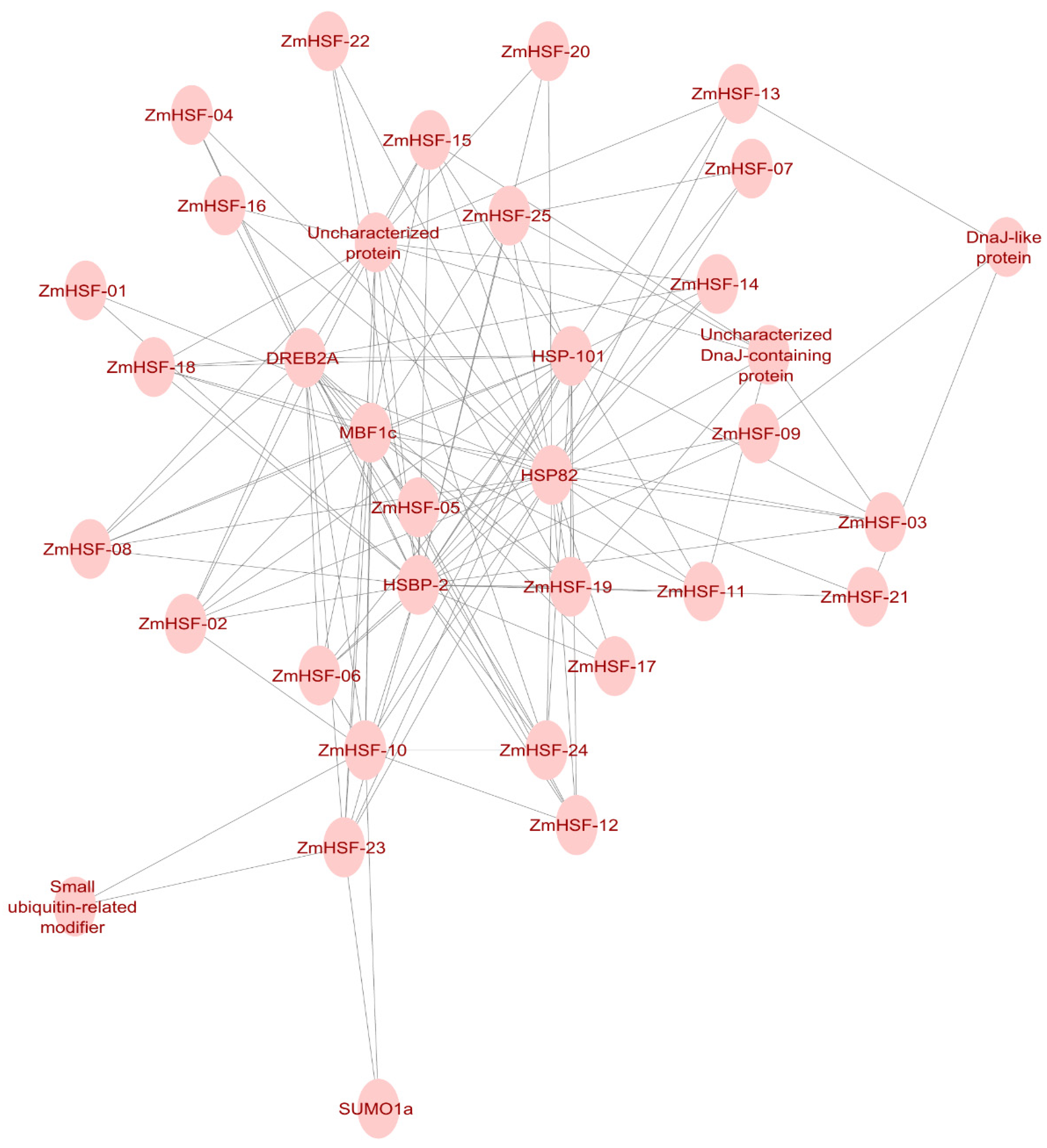
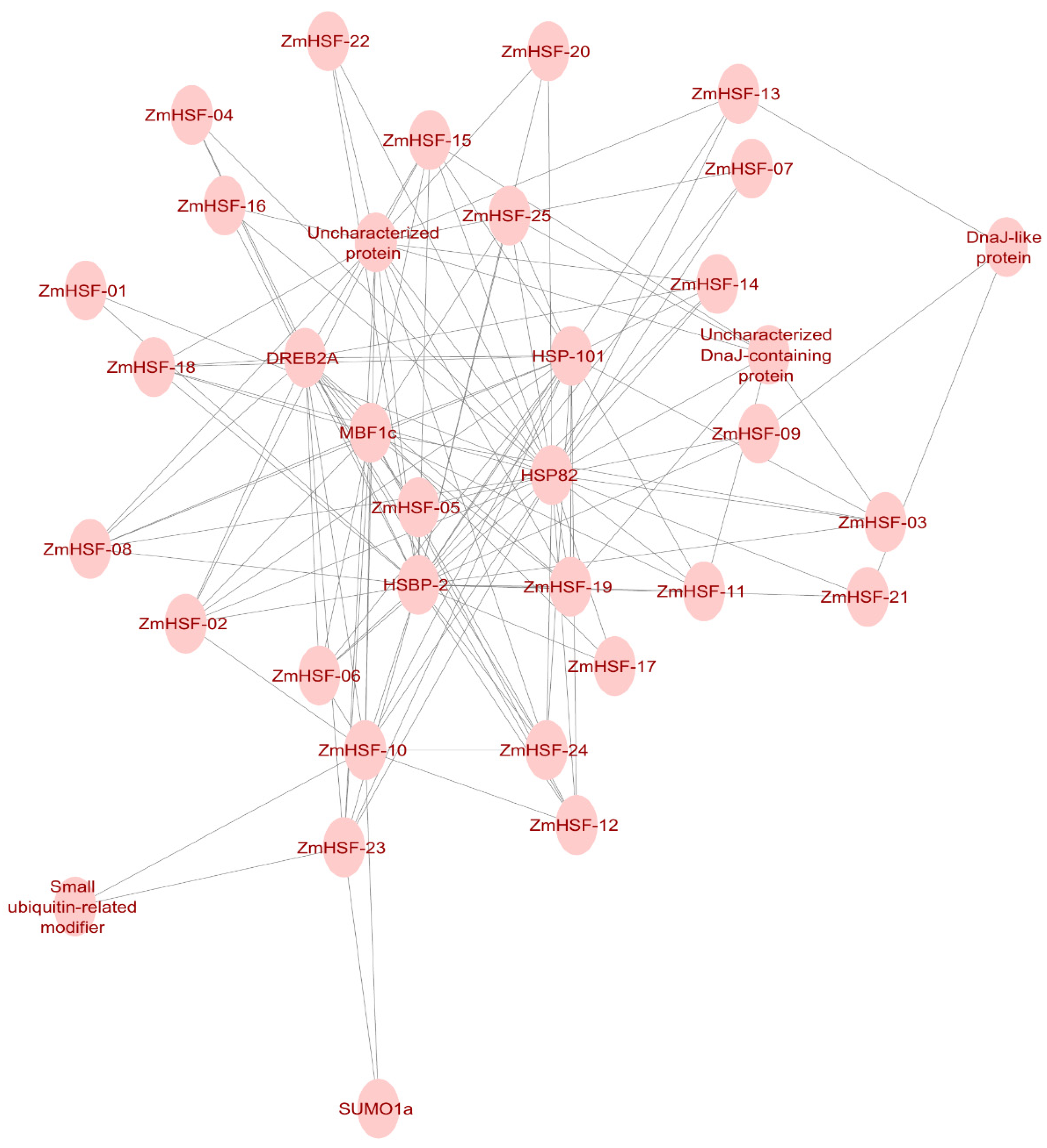
3.10. Protein–Protein Interaction Network Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shabbir, R.; Javed, T.; Afzal, I.; Sabagh, A.E.; Ali, A.; Vicente, O.; Chen, P. Modern Biotechnologies: Innovative and Sustainable Approaches for the Improvement of Sugarcane Tolerance to Environmental Stresses. Agronomy 2021, 11, 1042. [Google Scholar] [CrossRef]
- Lippmann, R.; Babben, S.; Menger, A.; Delker, C.; Quint, M. Development of wild and cultivated plants under global warming conditions. Curr. Biol. 2019, 29, R1326–R1338. [Google Scholar] [CrossRef]
- Haider, S.; Iqbal, J.; Naseer, S.; Yaseen, T.; Shaukat, M.; Bibi, H.; Ahmad, Y.; Daud, H.; Abbasi, N.L.; Mahmood, T. Molecular mechanisms of plant tolerance to heat stress: Current landscape and future perspectives. Plant Cell Rep. 2021, 40, 2247–2271. [Google Scholar] [CrossRef]
- Chaturvedi, P.; Wiese, A.J.; Ghatak, A.; Záveská Drábková, L.; Weckwerth, W.; Honys, D. Heat stress response mechanisms in pollen development. New Phytol. 2021, 231, 571–585. [Google Scholar] [CrossRef]
- Raza, A. Metabolomics: A systems biology approach for enhancing heat stress tolerance in plants. Plant Cell Rep. 2020, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Sharif, R.; Raza, A.; Chen, P.; Li, Y.; El-Ballat, E.M.; Rauf, A.; Hano, C.; El-Esawi, M.A. HD-ZIP gene family: Potential roles in improving plant growth and regulating stress-responsive mechanisms in plants. Genes 2021, 12, 1256. [Google Scholar] [CrossRef]
- Russo, S.; Marchese, A.F.; Sillmann, J.; Immé, G. When will unusual heat waves become normal in a warming Africa? Environ. Res. Lett. 2016, 11, 054016. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Hayes, S.; Schachtschabel, J.; Mishkind, M.; Munnik, T.; Arisz, S.A. Hot topic: Thermosensing in plants. Plant Cell Environ. 2021, 44, 2018–2033. [Google Scholar] [CrossRef]
- Zahra, N.; Shaukat, K.; Hafeez, M.B.; Raza, A.; Hussain, S.; Chaudhary, M.T.; Akram, M.Z.; Kakavand, S.N.; Saddiq, M.S.; Wahid, A. Physiological and molecular responses to high, chilling, and freezing temperature in plant growth and production: Consequences and mitigation possibilities. In Harsh Environment and Plant Resilience: Molecular and Functional Aspects; Springer: New York, NY, USA, 2021; p. 235. [Google Scholar]
- Mittler, R.; Finka, A.; Goloubinoff, P. How do plants feel the heat? Trends Biochem. Sci. 2012, 37, 118–125. [Google Scholar] [CrossRef]
- Ohama, N.; Sato, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional regulatory network of plant heat stress response. Trends Plant Sci. 2017, 22, 53–65. [Google Scholar] [CrossRef]
- Javed, T.; Shabbir, R.; Ali, A.; Afzal, I.; Zaheer, U.; Gao, S.J. Transcription factors in plant stress responses: Challenges and potential for sugarcane improvement. Plants 2020, 9, 491. [Google Scholar] [CrossRef]
- Raza, A.; Tabassum, J.; Kudapa, H.; Varshney, R.K. Can omics deliver temperature resilient ready-to-grow crops? Crit. Rev. Biotechnol. 2021, 41, 1209–1232. [Google Scholar] [CrossRef]
- Haider, S.; Iqbal, J.; Naseer, S.; Shaukat, M.; Abbasi, B.A.; Yaseen, T.; Zahra, S.A.; Mahmood, T. Unfolding molecular switches in plant heat stress resistance: A comprehensive review. Plant Cell Rep. 2021, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Andrási, N.; Pettkó-Szandtner, A.; Szabados, L. Diversity of plant heat shock factors: Regulation, interactions, and functions. J. Exp. Bot. 2021, 72, 1558–1575. [Google Scholar] [CrossRef]
- Sorger, P.K.; Pelham, H.R. Yeast heat shock factor is an essential DNA-binding protein that exhibits temperature-dependent phosphorylation. Cell 1988, 54, 855–864. [Google Scholar] [CrossRef]
- Scharf, K.-D.; Heider, H.; Höhfeld, I.; Lyck, R.; Schmidt, E.; Nover, L. The tomato Hsf system: HsfA2 needs interaction with HsfA1 for efficient nuclear import and may be localized in cytoplasmic heat stress granules. Mol. Cell. Biol. 1998, 18, 2240–2251. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Liu, J.-H.; Ma, X.; Luo, D.-X.; Gong, Z.-H.; Lu, M.-H. The plant heat stress transcription factors (HSFs): Structure, regulation, and function in response to abiotic stresses. Front. Plant Sci. 2016, 7, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nover, L.; Bharti, K.; Döring, P.; Mishra, S.K.; Ganguli, A.; Scharf, K.-D. Arabidopsis and the heat stress transcription factor world: How many heat stress transcription factors do we need? Cell Stress Chaperones 2001, 6, 177. [Google Scholar] [CrossRef]
- von Koskull-Döring, P.; Scharf, K.-D.; Nover, L. The diversity of plant heat stress transcription factors. Trends Plant Sci. 2007, 12, 452–457. [Google Scholar] [CrossRef]
- Scharf, K.-D.; Berberich, T.; Ebersberger, I.; Nover, L. The plant heat stress transcription factor (Hsf) family: Structure, function and evolution. Biochim. Biophys. Acta (BBA) Gene Regul. Mech. 2012, 1819, 104–119. [Google Scholar] [CrossRef]
- Bharti, K.; von Koskull-Döring, P.; Bharti, S.; Kumar, P.; Tintschl-Körbitzer, A.; Treuter, E.; Nover, L. Tomato heat stress transcription factor HsfB1 represents a novel type of general transcription coactivator with a histone-like motif interacting with the plant CREB binding protein ortholog HAC1. Plant Cell 2004, 16, 1521–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fragkostefanakis, S.; Simm, S.; El-Shershaby, A.; Hu, Y.; Bublak, D.; Mesihovic, A.; Darm, K.; Mishra, S.K.; Tschiersch, B.; Theres, K. The repressor and co-activator HsfB1 regulates the major heat stress transcription factors in tomato. Plant Cell Environ. 2019, 42, 874–890. [Google Scholar] [CrossRef] [PubMed]
- Jacob, P.; Hirt, H.; Bendahmane, A. The heat-shock protein/chaperone network and multiple stress resistance. Plant Biotechnol. J. 2017, 15, 405–414. [Google Scholar] [CrossRef]
- Czarnecka-Verner, E.; Yuan, C.-X.; Scharf, K.-D.; Englich, G.; Gurley, W.B. Plants contain a novel multi-member class of heat shock factors without transcriptional activator potential. Plant Mol. Biol. 2000, 43, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Kotak, S.; Larkindale, J.; Lee, U.; von Koskull-Döring, P.; Vierling, E.; Scharf, K.-D. Complexity of the heat stress response in plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef]
- Mishra, S.K.; Tripp, J.; Winkelhaus, S.; Tschiersch, B.; Theres, K.; Nover, L.; Scharf, K.-D. In the complex family of heat stress transcription factors, HsfA1 has a unique role as master regulator of thermotolerance in tomato. Genes Develop. 2002, 16, 1555–1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, T.; Ohama, N.; Nakajima, J.; Kidokoro, S.; Mizoi, J.; Nakashima, K.; Maruyama, K.; Kim, J.-M.; Seki, M.; Todaka, D. Arabidopsis HsfA1 transcription factors function as the main positive regulators in heat shock-responsive gene expression. Mol. Genet. Genom. 2011, 286, 321–332. [Google Scholar] [CrossRef]
- Hahn, A.; Bublak, D.; Schleiff, E.; Scharf, K.-D. Crosstalk between Hsp90 and Hsp70 chaperones and heat stress transcription factors in tomato. Plant Cell 2011, 23, 741–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charng, Y.-Y.; Liu, H.-C.; Liu, N.-Y.; Chi, W.-T.; Wang, C.-N.; Chang, S.-H.; Wang, T.-T. A heat-inducible transcription factor, HsfA2, is required for extension of acquired thermotolerance in Arabidopsis. Plant Physiol. 2007, 143, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Lämke, J.; Brzezinka, K.; Altmann, S.; Bäurle, I. A hit-and-run heat shock factor governs sustained histone methylation and transcriptional stress memory. EMBO J. 2016, 35, 162–175. [Google Scholar] [CrossRef] [Green Version]
- Nishizawa, A.; Yabuta, Y.; Yoshida, E.; Maruta, T.; Yoshimura, K.; Shigeoka, S. Arabidopsis heat shock transcription factor A2 as a key regulator in response to several types of environmental stress. Plant J. 2006, 48, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Sakuma, Y.; Todaka, D.; Maruyama, K.; Qin, F.; Mizoi, J.; Kidokoro, S.; Fujita, Y.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of an Arabidopsis heat-shock transcription factor HsfA3 in the transcriptional cascade downstream of the DREB2A stress-regulatory system. Biochem. Biophys. Res. Commun. 2008, 368, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.-F.; Tsai, M.-Y.; Lu, C.-A.; Wu, S.-J.; Yeh, C.-H. The roles of Arabidopsis HSFA2, HSFA4a, and HSFA7a in the heat shock response and cytosolic protein response. Bot. Stud. 2018, 59, 1–9. [Google Scholar] [CrossRef]
- Shim, D.; Hwang, J.-U.; Lee, J.; Lee, S.; Choi, Y.; An, G.; Martinoia, E.; Lee, Y. Orthologs of the class A4 heat shock transcription factor HsfA4a confer cadmium tolerance in wheat and rice. Plant Cell 2009, 21, 4031–4043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.-L.; Yuan, S.-N.; Zhang, H.-N.; Zhang, Y.-Y.; Zhang, Y.-J.; Wang, G.-Y.; Li, Y.-Q.; Li, G.-L. Heat-response patterns of the heat shock transcription factor family in advanced development stages of wheat (Triticum aestivum L.) and thermotolerance-regulation by TaHsfA2–10. BMC Plant Biol. 2020, 20, 364. [Google Scholar] [CrossRef]
- Kotak, S.; Vierling, E.; Bäumlein, H.; Koskull-0Döring, P.V. A novel transcriptional cascade regulating expression of heat stress proteins during seed development of Arabidopsis. Plant Cell 2007, 19, 182–195. [Google Scholar] [CrossRef] [Green Version]
- Albihlal, W.S.; Obomighie, I.; Blein, T.; Persad, R.; Chernukhin, I.; Crespi, M.; Bechtold, U.; Mullineaux, P.M. Arabidopsis heat shock transcription factora1b regulates multiple developmental genes under benign and stress conditions. J. Exp. Bot. 2018, 69, 2847–2862. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Wu, J.; Ji, Q.; Wang, C.; Luo, L.; Yuan, Y.; Wang, Y.; Wang, J. Genome-wide analysis of heat shock transcription factor families in rice and Arabidopsis. J. Genet. Genom. 2008, 35, 105–118. [Google Scholar] [CrossRef]
- Lin, Y.-X.; Jiang, H.-Y.; Chu, Z.-X.; Tang, X.-L.; Zhu, S.-W.; Cheng, B.-J. Genome-wide identification, classification and analysis of heat shock transcription factor family in maize. BMC Genom. 2011, 12, 76. [Google Scholar] [CrossRef] [Green Version]
- Chung, E.; Kim, K.-M.; Lee, J.-H. Genome-wide analysis and molecular characterization of heat shock transcription factor family in Glycine max. J. Genet. Genom. 2013, 40, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Dong, Q.; Jiang, H.; Zhu, S.; Chen, B.; Xiang, Y. Genome-wide analysis of the heat shock transcription factors in Populus trichocarpa and Medicago truncatula. Mol. Biol. Rep. 2012, 39, 1877–1886. [Google Scholar] [CrossRef]
- Yang, X.; Zhu, W.; Zhang, H.; Liu, N.; Tian, S. Heat shock factors in tomatoes: Genome-wide identification, phylogenetic analysis and expression profiling under development and heat stress. Peer J. 2016, 4, e1961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, F.; Wu, X.; Li, T.; Jia, M.; Liu, X.; Li, P.; Zhou, X.; Ji, X.; Yue, X. Genome-wide survey of heat shock factors and heat shock protein 70s and their regulatory network under abiotic stresses in Brachypodium distachyon. PLoS ONE 2017, 12, e0180352. [Google Scholar] [CrossRef] [Green Version]
- Duan, S.; Liu, B.; Zhang, Y.; Li, G.; Guo, X. Genome-wide identification and abiotic stress-responsive pattern of heat shock transcription factor family in Triticum aestivum L. BMC Genom. 2019, 20, 257. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Copley, R.R.; Schmidt, S.; Ciccarelli, F.D.; Doerks, T.; Schultz, J.; Ponting, C.P.; Bork, P. SMART 4.0: Towards genomic data integration. Nucl. Acids Res. 2004, 32, D142–D144. [Google Scholar] [CrossRef] [Green Version]
- Delorenzi, M.; Speed, T. An HMM model for coiled-coil domains and a comparison with PSSM-based predictions. Bioinformatics 2002, 18, 617–625. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucl. Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.S.; Chen, Y.C.; Lu, C.H.; Hwang, J.K. Prediction of protein subcellular localization. Proteins Struct. Funct. Bioinform. 2006, 64, 643–651. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucl. Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucl. Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Cavalcanti, A.; Chen, F.-C.; Bouman, P.; Li, W.-H. Extent of gene duplication in the genomes of Drosophila, nematode, and yeast. Mol. Biol. Evol. 2002, 19, 256–262. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Zhang, X.; Yue, J.-X.; Tian, D.; Chen, J.-Q. Recent duplications dominate NBS-encoding gene expansion in two woody species. Mol. Genet. Genom. 2008, 280, 187–198. [Google Scholar] [CrossRef]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, R.H.; Frank, H.M.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Stelpflug, S.C.; Sekhon, R.S.; Vaillancourt, B.; Hirsch, C.N.; Buell, C.R.; de Leon, N.; Kaeppler, S.M. An expanded maize gene expression atlas based on RNA sequencing and its use to explore root development. Plant Genome 2016, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P. The STRING database in 2021: Customizable protein–protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Consortium, G.O. The gene ontology resource: 20 years and still GOing strong. Nucl. Acids Res. 2019, 47, D330–D338. [Google Scholar]
- Rehman, S.; Jørgensen, B.; Aziz, E.; Batool, R.; Naseer, S.; Rasmussen, S.K. Genome wide identification and comparative analysis of the serpin gene family in brachypodium and barley. Plants 2020, 9, 1439. [Google Scholar] [CrossRef] [PubMed]
- Gaut, B.S. Evolutionary dynamics of grass genomes. New Phytol. 2002, 154, 15–28. [Google Scholar] [CrossRef] [Green Version]
- Initiative, I.B. Genome sequencing and analysis of the model grass Brachypodium distachyon. Nature 2010, 463, 763–768. [Google Scholar]
- Ali, A.; Javed, T.; Zaheer, U.; Zhou, J.R.; Huang, M.T.; Fu, H.Y.; Gao, S.J. Genome-Wide Identification and Expression Profiling of the bHLH Transcription Factor Gene Family in Saccharum spontaneum Under Bacterial Pathogen Stimuli. Tropical Plant Biol. 2021, 14, 283–294. [Google Scholar] [CrossRef]
- Xu, X.; Yang, Y.; Liu, C.; Sun, Y.; Zhang, T.; Hou, M.; Huang, S.; Yuan, H. The evolutionary history of the sucrose synthase gene family in higher plants. BMC Plant Biol. 2019, 19, 566. [Google Scholar] [CrossRef] [Green Version]
- Fragkostefanakis, S.; Röth, S.; Schleiff, E.; Scharf, K.D. Prospects of engineering thermotolerance in crops through modulation of heat stress transcription factor and heat shock protein networks. Plant Cell Environ. 2015, 38, 1881–1895. [Google Scholar] [CrossRef]
- De Cuyper, C.; Struk, S.; Braem, L.; Gevaert, K.; De Jaeger, G.; Goormachtig, S. Strigolactones, karrikins and beyond. Plant Cell Environ. 2017, 40, 1691–1703. [Google Scholar] [CrossRef]
- Ye, J.; Yang, X.; Hu, G.; Liu, Q.; Li, W.; Zhang, L.; Song, X. Genome-wide investigation of heat shock transcription factor family in wheat (Triticum aestivum L.) and possible roles in anther development. Int. J. Mol. Sci. 2020, 21, 608. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.-Y.; Juan, Y.-T.; Hsu, Y.-H.; Wu, S.-H.; Liao, H.-T.; Fung, R.W.; Charng, Y.-Y. Interplay between heat shock proteins HSP101 and HSA32 prolongs heat acclimation memory posttranscriptionally in Arabidopsis. Plant Physiol. 2013, 161, 2075–2084. [Google Scholar] [CrossRef] [Green Version]
- Borkovich, K.; Farrelly, F.; Finkelstein, D.; Taulien, J.; Lindquist, S. hsp82 is an essential protein that is required in higher concentrations for growth of cells at higher temperatures. Mol. Cell. Biol. 1989, 9, 3919–3930. [Google Scholar]
- Gu, L.; Jiang, T.; Zhang, C.; Li, X.; Wang, C.; Zhang, Y.; Li, T.; Dirk, L.M.; Downie, A.B.; Zhao, T. Maize HSFA2 and HSBP2 antagonistically modulate raffinose biosynthesis and heat tolerance in Arabidopsis. Plant J. 2019, 100, 128–142. [Google Scholar] [CrossRef] [PubMed]
- Pulido, P.; Leister, D. Novel DNAJ-related proteins in Arabidopsis thaliana. New Phytol. 2018, 217, 480–490. [Google Scholar] [CrossRef] [Green Version]
- Kong, F.; Deng, Y.; Wang, G.; Wang, J.; Liang, X.; Meng, Q. LeCDJ1, a chloroplast DnaJ protein, facilitates heat tolerance in transgenic tomatoes. J. Integr. Plant Biol. 2014, 56, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Deng, Y.; Zhou, B.; Wang, G.; Wang, Y.; Meng, Q. A chloroplast-targeted DnaJ protein contributes to maintenance of photosystem II under chilling stress. J. Exp. Bot. 2014, 65, 143–158. [Google Scholar] [CrossRef] [Green Version]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF family transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta (BBA) Gene Regul. Mech. 2012, 1819, 86–96. [Google Scholar] [CrossRef]
- Suzuki, N.; Bajad, S.; Shuman, J.; Shulaev, V.; Mittler, R. The transcriptional co-activator MBF1c is a key regulator of thermotolerance in Arabidopsis thaliana. J. Biol. Chem. 2008, 283, 9269–9275. [Google Scholar] [CrossRef] [Green Version]
- Jaimes-Miranda, F.; Chávez Montes, R.A. The plant MBF1 protein family: A bridge between stress and transcription. J. Exp. Bot. 2020, 71, 1782–1791. [Google Scholar] [CrossRef]
- Hay, R.T. SUMO: A history of modification. Mol. Cell 2005, 18, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rytz, T.C.; Miller, M.J.; McLoughlin, F.; Augustine, R.C.; Marshall, R.S.; Juan, Y.-T.; Charng, Y.-Y.; Scalf, M.; Smith, L.M.; Vierstra, R.D. SUMOylome profiling reveals a diverse array of nuclear targets modified by the SUMO ligase SIZ1 during heat stress. Plant Cell 2018, 30, 1077–1099. [Google Scholar] [CrossRef] [PubMed]
- Strable, J.; Scanlon, M.J. Maize (Zea mays): A model organism for basic and applied research in plant biology. Cold Spring Harb. Protoc. 2009, 2009, emo132. [Google Scholar] [CrossRef] [Green Version]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A. The B73 maize genome: Complexity, diversity, and dynamics. Science 2009, 326, 1112–1115. [Google Scholar] [CrossRef] [Green Version]
- Nagaraju, M.; Sudhakar Reddy, P.; Anil Kumar, S.; Srivastava, R.K.; Kavi Kishor, P.; Rao, D.M. Genome-wide scanning and characterization of Sorghum bicolor L. heat shock transcription factors. Curr. Genom. 2015, 16, 279–291. [Google Scholar] [CrossRef] [Green Version]
- Kaul, S.; Koo, H.L.; Jenkins, J.; Rizzo, M.; Rooney, T.; Tallon, L.J.; Feldblyum, T.; Nierman, W.; Benito, M.-I.; Lin, X. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar]
- Young, N.D.; Debellé, F.; Oldroyd, G.E.; Geurts, R.; Cannon, S.B.; Udvardi, M.K.; Benedito, V.A.; Mayer, K.F.; Gouzy, J.; Schoof, H. The Medicago genome provides insight into the evolution of rhizobial symbioses. Nature 2011, 480, 520–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Hu, S.; Wang, J.; Wong, G.K.-S.; Li, S.; Liu, B.; Deng, Y.; Dai, L.; Zhou, Y.; Zhang, X. A draft sequence of the rice genome (Oryza sativa L. ssp. indica). Science 2002, 296, 79–92. [Google Scholar] [CrossRef]
- Raza, A.; Su, W.; Gao, A.; Mehmood, S.S.; Hussain, M.A.; Nie, W.; Lv, Y.; Zou, X.; Zhang, X. Catalase (CAT) Gene Family in Rapeseed (Brassica napus L.): Genome-Wide Analysis, Identification, and Expression Pattern in Response to Multiple Hormones and Abiotic Stress Conditions. Int. J. Mol. Sci. 2021, 22, 4281. [Google Scholar] [CrossRef] [PubMed]
- Van de Peer, Y.; Maere, S.; Meyer, A. The evolutionary significance of ancient genome duplications. Nat. Rev. Genet. 2009, 10, 725–732. [Google Scholar] [CrossRef] [Green Version]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Jiang, H.; Chen, W.; Qian, Y.; Ma, Q.; Cheng, B.; Zhu, S. Genome-wide analysis of the auxin response factor (ARF) gene family in maize (Zea mays). Plant Growth Regul. 2011, 63, 225–234. [Google Scholar] [CrossRef]
- Solé, R.V.; Fernandez, P.; Kauffman, S.A. Adaptive walks in a gene network model of morphogenesis: Insights into the Cambrian explosion. arXiv 2003, arXiv:q-bio/0311013. [Google Scholar]
- Maere, S.; De Bodt, S.; Raes, J.; Casneuf, T.; Van Montagu, M.; Kuiper, M.; Van de Peer, Y. Modeling gene and genome duplications in eukaryotes. Proc. Natl. Acad. Sci. USA 2005, 102, 5454–5459. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Wickett, N.J.; Ayyampalayam, S.; Chanderbali, A.S.; Landherr, L.; Ralph, P.E.; Tomsho, L.P.; Hu, Y.; Liang, H.; Soltis, P.S. Ancestral polyploidy in seed plants and angiosperms. Nature 2011, 473, 97–100. [Google Scholar] [CrossRef]
- Rose, A. Intron-mediated regulation of gene expression. Nucl. Pre-Mrna Process. Plants 2008, 326, 277–290. [Google Scholar]
- Liu, H.-C.; Charng, Y.-Y. Common and distinct functions of Arabidopsis class A1 and A2 heat shock factors in diverse abiotic stress responses and development. Plant Physiol. 2013, 163, 276–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.R.; Goswami, S.; Singh, K.; Dubey, K.; Rai, G.K.; Singh, B.; Singh, S.; Grover, M.; Mishra, D.; Kumar, S. Characterization of novel heat-responsive transcription factor (TaHSFA6e) gene involved in regulation of heat shock proteins (HSPs)—A key member of heat stress-tolerance network of wheat. J. Biotechnol. 2018, 279, 1–12. [Google Scholar] [CrossRef]
- Yokotani, N.; Ichikawa, T.; Kondou, Y.; Matsui, M.; Hirochika, H.; Iwabuchi, M.; Oda, K. Expression of rice heat stress transcription factor OsHsfA2e enhances tolerance to environmental stresses in transgenic Arabidopsis. Planta 2008, 227, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zheng, Q.; Chen, L.; Liang, Y.; Wu, J. Ectopic overexpression of maize heat shock transcription factor gene ZmHsf04 confers increased thermo and salt-stress tolerance in transgenic Arabidopsis. Acta Physiol. Plant. 2018, 40, 9. [Google Scholar] [CrossRef]
- Li, G.; Zhang, Y.; Zhang, H.; Zhang, Y.; Zhao, L.; Liu, Z.; Guo, X. Characteristics and regulating role in thermotolerance of the heat shock transcription factor ZmHsf12 from Zea mays L. J. Plant Biol. 2019, 62, 329–341. [Google Scholar] [CrossRef]
- Banti, V.; Mafessoni, F.; Loreti, E.; Alpi, A.; Perata, P. The heat-inducible transcription factor HsfA2 enhances anoxia tolerance in Arabidopsis. Plant Physiol. 2010, 152, 1471–1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.J.; Barrett-Wilt, G.A.; Hua, Z.; Vierstra, R.D. Proteomic analyses identify a diverse array of nuclear processes affected by small ubiquitin-like modifier conjugation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2010, 107, 16512–16517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Wang, S.; Lv, J.; Liu, Z.; Wang, Y.; Ma, N.; Meng, Q. SUMO E3 ligase SlSIZ1 facilitates heat tolerance in tomato. Plant Cell Physiol. 2018, 59, 58–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Transcript ID | Gene ID | Chromosome | Location | Strand | Transcript Count | Genomic DNA | CDS Length | Exon | Intron |
|---|---|---|---|---|---|---|---|---|---|---|
| ZmHSF-01 | GRMZM2G165972_T02 | Zm00001d027757 | 1 | 12518410–12520734 | + | 4 | 2325 | 1590 | 2 | 1 |
| ZmHSF-02 | GRMZM2G118485_T02 | Zm00001d028269 | 1 | 28522823–28529222 | + | 3 | 7416 | 1254 | 3 | 2 |
| ZmHSF-03 | GRMZM2G164909_T01 | Zm00001d029270 | 1 | 197781323–197783002 | − | 1 | 1680 | 1245 | 2 | 1 |
| ZmHSF-04 | GRMZM2G010871_T01 | Zm00001d032923 | 1 | 239675070–239678097 | − | 6 | 3628 | 1074 | 2 | 1 |
| ZmHSF-05 | GRMZM2G132971_T01 | Zm00001d034433 | 1 | 287786016–287790222 | + | 3 | 4207 | 1080 | 2 | 1 |
| ZmHSF-06 | GRMZM2G115456_T01 | Zm00001d034886 | 1 | 299272434–299277717 | + | 1 | 5284 | 1584 | 2 | 1 |
| ZmHSF-07 | GRMZM2G088242_T01 | Zm00001d005843 | 2 | 185666362–185668432 | + | 1 | 2071 | 1185 | 2 | 1 |
| ZmHSF-08 | GRMZM2G002131_T01 | Zm00001d005888 | 2 | 186829423–186833487 | + | 1 | 4065 | 1125 | 2 | 1 |
| ZmHSF-09 | GRMZM2G089525_T01 | Zm00001d044168 | 3 | 217438918–217440728 | − | 1 | 1811 | 996 | 2 | 1 |
| ZmHSF-10 | GRMZM2G005815_T01 | Zm00001d044259 | 3 | 219545854–219552097 | + | 2 | 6244 | 1389 | 2 | 1 |
| ZmHSF-11 | GRMZM2G098696_T01 | Zm00001d052738 | 4 | 195298840–195300584 | − | 2 | 1745 | 1113 | 2 | 1 |
| ZmHSF12 | GRMZM2G384339_T01 | Zm00001d016520 | 5 | 889476–893867 | − | 2 | 4392 | 1494 | 2 | 1 |
| ZmHSF-13 | GRMZM2G105348_T01 | Zm00001d016674 | 5 | 150228697–150230232 | − | 1 | 1536 | 1185 | 2 | 1 |
| ZmHSF-14 | GRMZM2G179802_T02 | Zm00001d012823 | 5 | 162047221–162053475 | − | 3 | 6261 | 1920 | 2 | 1 |
| ZmHSF-15 | GRMZM2G059851_T01 | Zm00001d016255 | 5 | 168263116–168266561 | + | 2 | 3446 | 1527 | 2 | 1 |
| ZmHSF-16 | AC206165.3_FGT007 | Zm00001d038746 | 6 | 159812447–159814105 | − | 1 | 1659 | 1410 | 2 | 1 |
| ZmHSF-17 | GRMZM2G125969_T01 | Zm00001d020714 | 7 | 9712577–9716596 | − | 3 | 4020 | 1284 | 2 | 1 |
| ZmHSF-18 | GRMZM2G139535_T01 | Zm00001d021263 | 7 | 125896425–125900126 | + | 2 | 3702 | 897 | 2 | 1 |
| ZmHSF-19 | GRMZM2G165272_T01 | Zm00001d022295 | 7 | 142270318- 142272208 | + | 1 | 1891 | 1185 | 2 | 1 |
| ZmHSF-20 | AC205471.4_FGT003 | Zm00001d010812 | 8 | 124710390–124712153 | + | 1 | 1764 | 1341 | 2 | 1 |
| ZmHSF-21 | GRMZM2G086880_T01 | Zm00001d011406 | 8 | 144887947–144889308 | + | 1 | 1364 | 1248 | 2 | 1 |
| ZmHSF-22 | GRMZM2G118453_T01 | Zm00001d012749 | 8 | 173979796–173982203 | − | 1 | 2407 | 1870 | 2 | 1 |
| ZmHSF-23 | GRMZM2G173090_T02 | Zm00001d046204 | 9 | 59460696–59462007 | + | 2 | 1473 | 1357 | 2 | 1 |
| ZmHSF-24 | GRMZM2G026742_T01 | Zm00001d048041 | 9 | 146029209–146032662 | − | 4 | 4461 | 1840 | 5 | 4 |
| ZmHSF-25 | GRMZM2G301485_T01 | Zm00001d026094 | 10 | 137124859–137126235 | − | 2 | 1376 | 1300 | 2 | 1 |
| Sub-Class | Arabidopsis AtHSFs | Zea mays ZmHSFs | Sorghum bicolor SbHSFs | Oryza sativa OsHSFs | Brachypodium distachyon BdHSFs |
|---|---|---|---|---|---|
| A1 | 4 | 2 | 1 | 1 | 1 |
| A2 | 1 | 2 | 3 | 3 | 3 |
| A3 | 1 | 1 | 1 | 1 | 1 |
| A4 | 2 | 3 | 1 | 2 | 2 |
| A5 | 1 | 1 | 1 | 1 | 1 |
| A6 | 2 | 2 | 2 | 2 | 2 |
| A7 | 2 | 2 | 2 | 2 | 2 |
| A8 | 1 | 2 | 1 | 1 | 1 |
| A9 | 1 | 0 | 0 | 0 | 0 |
| B1 | 1 | 2 | 1 | 1 | 1 |
| B2 | 2 | 4 | 3 | 3 | 3 |
| B3 | 1 | 0 | 0 | 0 | 0 |
| B4 | 1 | 1 | 3 | 4 | 3 |
| C1 | 1 | 2 | 2 | 2 | 2 |
| C2 | 0 | 1 | 2 | 2 | 2 |
| Total | 21 | 25 | 23 | 25 | 24 |
| Duplicated Genes | Ka | Ks | Ka/Ks | Estimated Time (MYA) | Mode of Duplication |
|---|---|---|---|---|---|
| ZmHSF-01/ZmHSF-04 | 0.1837 | 0.3733 | 0.4921 | 30.59 | Tandem |
| ZmHSF-02/ZmHSF-24 | 0.0248 | 0.0727 | 0.3415 | 5.96 | Segmental |
| ZmHSF-03/ZmHSF-11 | 0.2497 | 0.3726 | 0.6702 | 30.54 | Segmental |
| ZmHSF-05/ZmHSF-17 | 0.1810 | 0.3250 | 0.5570 | 26.63 | Segmental |
| ZmHSF-06/ZmHSF-12 | 0.2772 | 0.4600 | 0.6026 | 37.70 | Segmental |
| ZmHSF-07/ZmHSF-19 | 0.1538 | 2.2641 | 0.5143 | 21.64 | Segmental |
| ZmHSF-08/ZmHSF-18 | 0.1814 | 0.2395 | 0.7572 | 19.63 | Segmental |
| ZmHSF-09/ZmHSF-21 | 0.0389 | 0.0839 | 0.4644 | 6.87 | Segmental |
| ZmHSF-16/ZmHSF20 | 0.0733 | 0.1040 | 0.7049 | 8.52 | Segmental |
| Motif | Consensus Sequence |
|---|---|
| 1 | LPKYFKHNNFSSFVRQLNTYGFRKVDPDRWEFANEGFLRGQKHLLKNIHR |
| 2 | PFLTKTYEMVDDPATDAVVSWGAAGNSFV |
| 3 | LLAELVRLRQZQQSTSEQLQALERRLQGMEQRQQQMMAFLA |
| 4 | VWBPAEFARDL |
| 5 | GLEEEIERLKRDKAL |
| 6 | MQNPDFLRQLVQQQEKSKELEDAINKKRR |
| 7 | LARELAQMRKLCNNILLLMSKYADTQQPD |
| 8 | VNDDFWEZFLTE |
| 9 | DGGPVDDSEAAGGGGQIIKYQPPIPEAAKQPLPKNLAFDSS |
| 10 | MPMDVEMASNNVGTFDSTGNDFTDTSALCEWDDMDIFGGELEHILQQPEQ |
| 11 | QSWPIYRPRPVYHPLRACNG |
| 12 | KPSQDGPSDPQQPPVKTAPGPENIEIGKY |
| 13 | TIEDYGYDRPWLEQDCQMEAQQNCKNPQY |
| 14 | KSVKLFGVLLKDAARKRGRCEEAAASERPIKMIRIGEPWIGVPSSGPGRC |
| 15 | RLFGVSIGRKRMRD |
| 16 | DNVBQLTEQMGYLSSANH |
| 17 | RKPIHSHSPQTQ |
| 18 | VDMCSDTTTGDTSQDETTSETGGSHGPAK |
| 19 | CCISMGGEDHR |
| 20 | PRPMEGLHDVGPP |
| Gene | Group | Protein ID | DBD | OD | Amino Acid | pI | Molecular Weight | Localization CELLO | Localization WoLFPSORT |
|---|---|---|---|---|---|---|---|---|---|
| ZmHSF-01 | A-6b | GRMZM2G165972_P02 | 44–133 | 164–184 | 384 | 5.30 | 43,268.46 | Nucleus | Nucleus |
| ZmHSF-02 | A-8a | GRMZM2G118485_P02 | 51–140 | 170–190 | 417 | 5.09 | 46,816.21 | Nucleus | Nucleus |
| ZmHSF-03 | B-2d | GRMZM2G164909_P01 | 72–161 | 231–260 | 414 | 7.14 | 44,381.82 | Nucleus | Nucleus |
| ZmHSF-04 | A-6a | GRMZM2G010871_P01 | 46–135 | 165–206 | 357 | 5.03 | 40,550.58 | Nucleus | Nucleus |
| ZmHSF-05 | A-2a | GRMZM2G132971_P01 | 40–129 | 149–204 | 359 | 5.57 | 40,587.23 | Nucleus | Nucleus |
| ZmHSF-06 | A-1a | GRMZM2G115456_P01 | 59–148 | 176–203 | 527 | 5.11 | 56,724.50 | Nucleus | Nucleus |
| ZmHSF-07 | B-4a | GRMZM2G088242_P01 | 22–112 | 216–243 | 394 | 7.81 | 41,742.83 | Nucleus | Nucleus |
| ZmHSF-08 | B-1a | GRMZM2G002131_P01 | 18–107 | 165–192 | 298 | 9.13 | 32,270.31 | Nucleus | Nucleus |
| ZmHSF-09 | C-1c | GRMZM2G089525_P01 | 27–116 | 168–188 | 331 | 5.94 | 35,883.69 | Nucleus | Nucleus |
| ZmHSF-10 | A-7a | GRMZM2G005815_P01 | 140–231 | 260–280 | 462 | 8.87 | 50,970.67 | Chloroplast | Nucleus |
| ZmHSF-11 | B-2b | GRMZM2G098696_P01 | 37–126 | 196–216 | 370 | 5.89 | 39,562.03 | Nucleus | Nucleus |
| ZmHSF-12 | A-1b | GRMZM2G059851_P01 | 76–165 | 190–231 | 508 | 4.96 | 56,061.66 | Nucleus | Nucleus |
| ZmHSF-13 | C-2a | GRMZM2G105348_P01 | 15–104 | 135–171 | 257 | 5.85 | 27,836.98 | Nucleus | Nucleus |
| ZmHSF-14 | A-5 | GRMZM2G179802_P02 | 69–159 | 177–225 | 528 | 5.57 | 58,138.65 | Nucleus | Nucleus |
| ZmHSF-15 | A-3 | GRMZM2G384339_P01 | 32–121 | 149–176 | 497 | 5.06 | 54,104.58 | Nucleus | Nucleus |
| ZmHSF-16 | A-4b | AC206165.3_FGP007 | 25–114 | 135–155 | 469 | 5.41 | 51,623.79 | Nucleus | Nucleus |
| ZmHSF-17 | A-2b | GRMZM2G125969_P01 | 52–141 | 169–210 | 375 | 4.70 | 42,043.73 | Nucleus | Nucleus |
| ZmHSF-18 | B-1b | GRMZM2G139535_P01 | 20–109 | 169–189 | 298 | 9.53 | 32,258.35 | Nucleus | Nucleus |
| ZmHSF-19 | B-2c | GRMZM2G165272_P01 | 45–134 | 207–234 | 394 | 5.00 | 41,468.10 | Nucleus | Nucleus |
| ZmHSF-20 | A-4d | AC205471.4_FGP003 | 13–102 | 123–143 | 446 | 5.15 | 49,718.46 | Nucleus | Nucleus |
| ZmHSF-21 | C-1a | GRMZM2G086880_P01 | 35–124 | 173–193 | 348 | 8.09 | 37,409.50 | Nucleus | Nucleus |
| ZmHSF-22 | A-4a | GRMZM2G118453_P01 | 10–99 | 120–175 | 433 | 5.30 | 48,829.92 | Nucleus | Nucleus |
| ZmHSF-23 | A-7b | GRMZM2G173090_P02 | 51–140 | 164–205 | 350 | 4.95 | 38,154.66 | Cytoplasm | Cytoplasm |
| ZmHSF-24 | A-8b | GRMZM2G026742_P01 | 51–140 | 170–190 | 407 | 4.97 | 45,317.63 | Nucleus | Nucleus |
| ZmHSF-25 | B-2a | GRMZM2G301485_P01 | 51–140 | 170–190 | 318 | 5.89 | 39,562.03 | Nucleus | Nucleus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haider, S.; Rehman, S.; Ahmad, Y.; Raza, A.; Tabassum, J.; Javed, T.; Osman, H.S.; Mahmood, T. In Silico Characterization and Expression Profiles of Heat Shock Transcription Factors (HSFs) in Maize (Zea mays L.). Agronomy 2021, 11, 2335. https://doi.org/10.3390/agronomy11112335
Haider S, Rehman S, Ahmad Y, Raza A, Tabassum J, Javed T, Osman HS, Mahmood T. In Silico Characterization and Expression Profiles of Heat Shock Transcription Factors (HSFs) in Maize (Zea mays L.). Agronomy. 2021; 11(11):2335. https://doi.org/10.3390/agronomy11112335
Chicago/Turabian StyleHaider, Saqlain, Shazia Rehman, Yumna Ahmad, Ali Raza, Javaria Tabassum, Talha Javed, Hany S. Osman, and Tariq Mahmood. 2021. "In Silico Characterization and Expression Profiles of Heat Shock Transcription Factors (HSFs) in Maize (Zea mays L.)" Agronomy 11, no. 11: 2335. https://doi.org/10.3390/agronomy11112335
APA StyleHaider, S., Rehman, S., Ahmad, Y., Raza, A., Tabassum, J., Javed, T., Osman, H. S., & Mahmood, T. (2021). In Silico Characterization and Expression Profiles of Heat Shock Transcription Factors (HSFs) in Maize (Zea mays L.). Agronomy, 11(11), 2335. https://doi.org/10.3390/agronomy11112335