
Effects of Nitrogen, Azoxystrobin and a Biostimulant Based on Brown Algae and Yeast on Wild Rocket Features at Harvest and During Storage



 ,
,  ,
,  , , ,
, , ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Experimental Field Trial
2.1.1. Characteristics of the Experimental Site
2.1.2. Treatments, Experimental Design and Crop Management
2.2. Yield Assessment
2.3. Shelf Life Assessment
2.4. Quality Features Measurement
2.5. Statistical Analysis
3. Results
3.1. Effect of Preharvest Treatments on Rocket’s Yield and Quality at Harvest
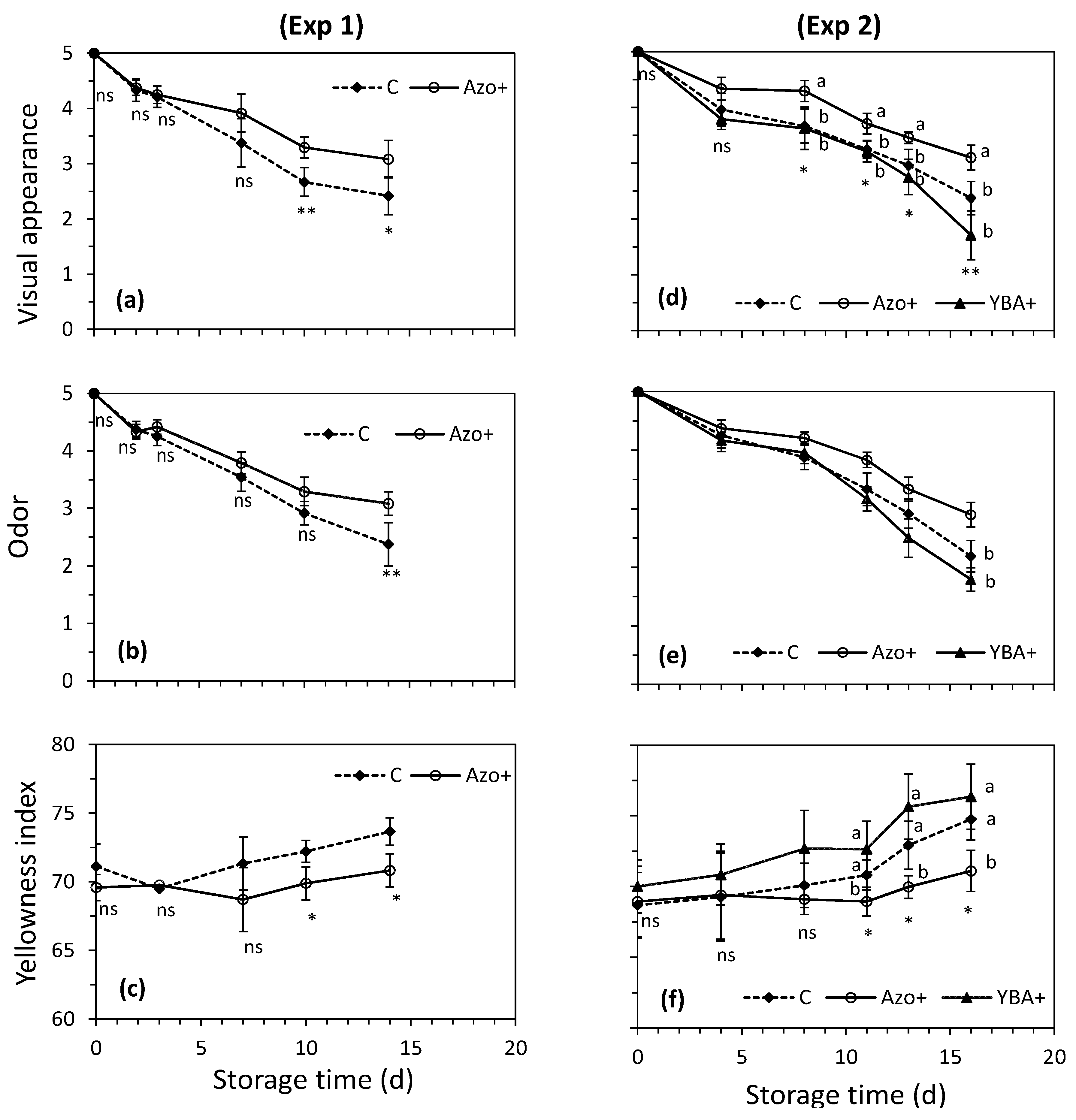
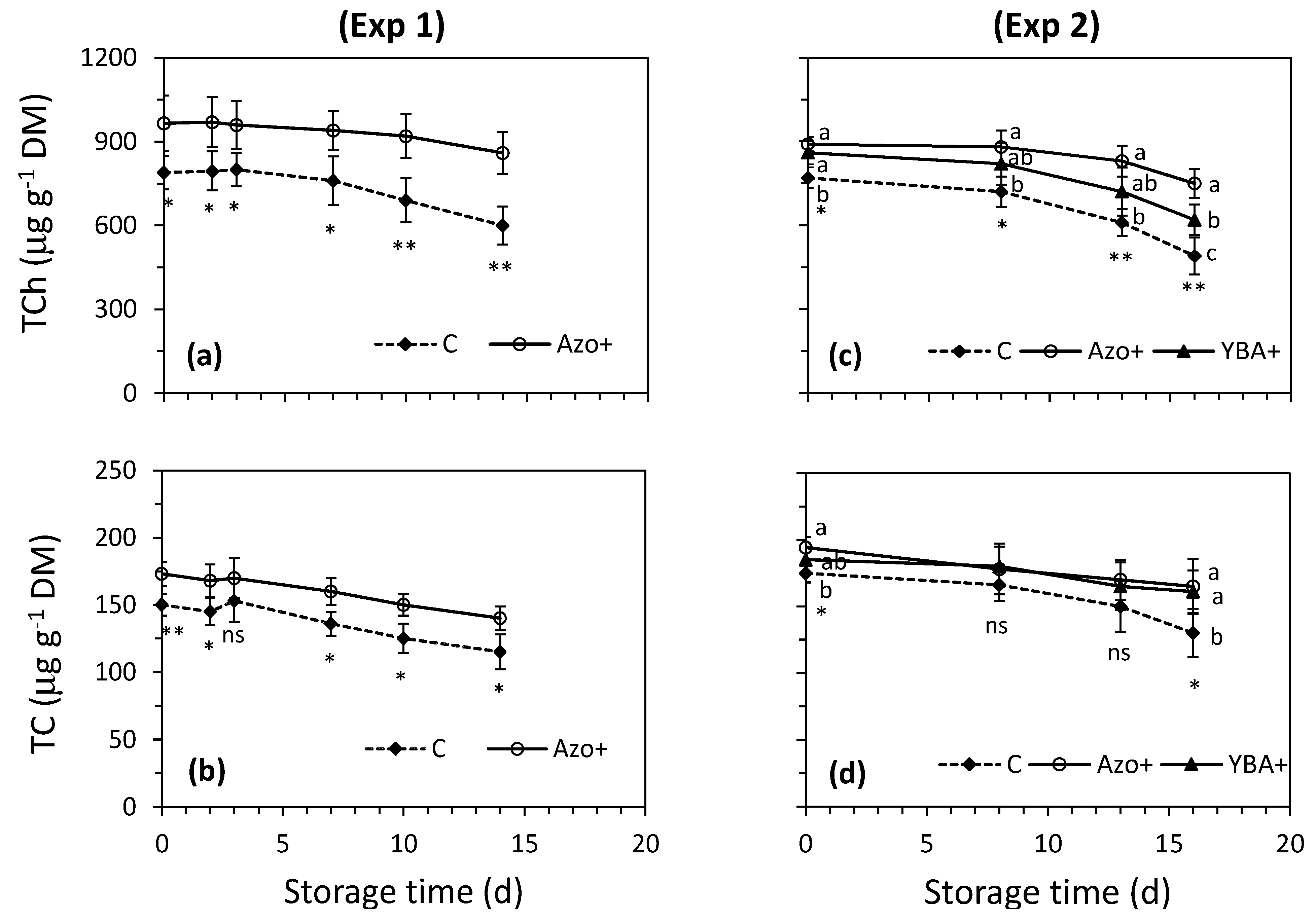
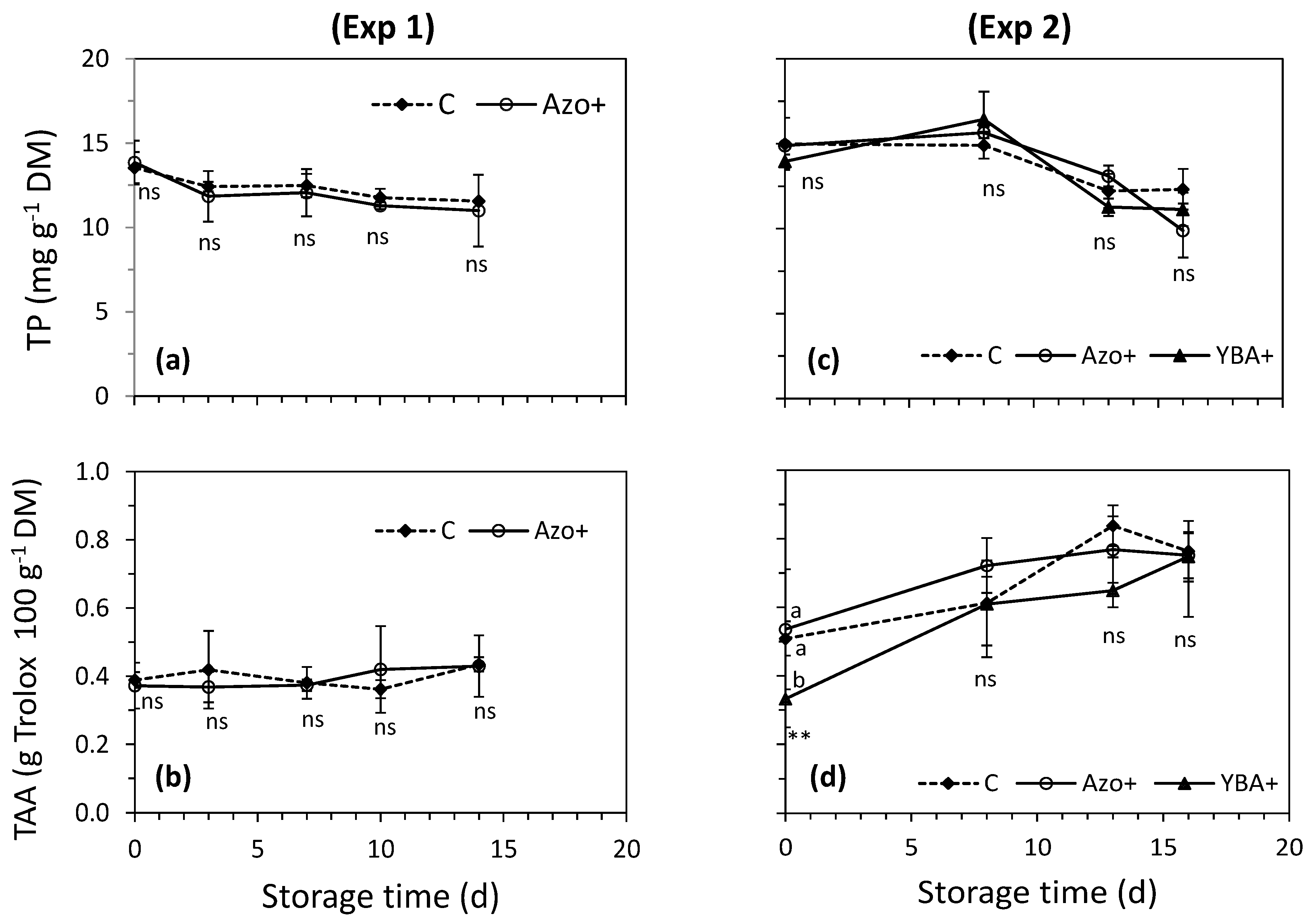
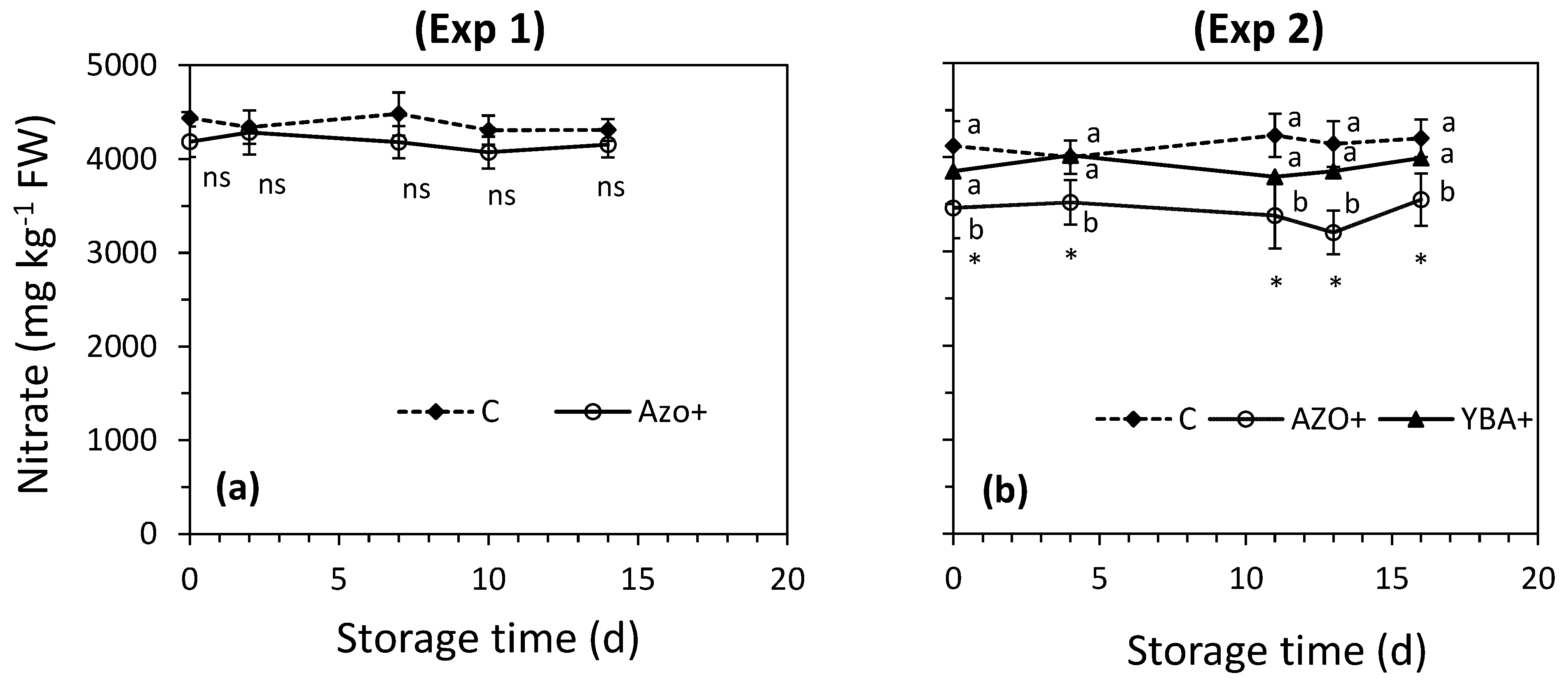
3.2. Effect of Preharvest Treatments on Rocket’s Characteristics during the Shelf-Life
4. Discussion
4.1. Effect of Preharvest Treatments on Wild Rocket’s Yield and Quality at Harvest
4.2. Effect of Preharvest Treatments on Rocket’s Characteristics during the Shelf-Life
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fallovo, C.; Rouphael, Y.; Rea, E.; Battistelli, A.; Colla, G. Nutrient solution concentration and growing season affect yield and quality of Lactuca sativa L. var. acephala in floating raft culture. J. Sci. Food Agric. 2009, 89, 1682–1689. [Google Scholar] [CrossRef]
- Bonasia, A.; Lazzizera, C.; Elia, A.; Conversa, G. Nutritional, biophysical and physiological characteristics of wild rocket genotypes as affected by soilless cultivation system, salinity level of nutrient solution and growing period. Front. Plant Sci. 2017, 8, 300. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.K.D.; Joblling, J.J.; Rogers, G.S. Influence of storage temperature on the seasonal shelf life of perennial wall rocket and annual garden rocket. Int. J. Veg. Sci. 2013, 19, 83–95. [Google Scholar] [CrossRef]
- Martínez-Sánchez, A.; Allende, A.; Cortes-Galera, I.; Gil, M.I. Respiration rate response of four baby leaf Brassica species to cutting at harvest and fresh cut washing. Postharvest Biol. Technol. 2008, 47, 382–388. [Google Scholar] [CrossRef]
- Martínez-Sánchez, A.; Luna, M.C.; Selma, M.V.; Tudela, J.A.; Abad, J.; Gil, M.I. Baby-leaf and multi-leaf of green and red lettuces are suitable raw materials for the fresh-cut industry. Postharvest Biol. Technol. 2012, 63, 1–10. [Google Scholar] [CrossRef]
- Cantwell, M.; Rovelo, J.; Nie, X.; Rubatzky, V. Specialty salad greens: Postharvest physiology and shelf-life. Acta Hortic. 1998, 467, 371–377. [Google Scholar] [CrossRef]
- Candido, V.; Boari, F.; Cantore, V.; Castronuovo, D.; Di Venere, D.; Perniola, M.; Sergio, L.; Viggiani, R.; Schiattone, M.I. In-teractive effect of nitrogen and Azoxystrobin on yield, quality, nitrogen and water use efficiency of wild rocket in Southern Italy. Agronomy 2020, 10, 849. [Google Scholar] [CrossRef]
- EC. Regulation (EU) December 2011 amending Regulation (EC) No. 1881/as regards maximum levels for nitrates in foodstuffs. Off. J. Eur. Union L. 2011, 320, 15–17. [Google Scholar]
- Amaro, A.C.E.; Baron, D.; Ono, E.O.; Rodrigues, J.D. Physiological effects of strobilurin and carboxamides on plants: An overview. Acta Physiol. Plant. 2020, 42, 4. [Google Scholar] [CrossRef]
- Boari, F.; Cantore, V.; Di Venere, D.; Sergio, L.; Candido, V.; Schiattone, M.I. Pyraclostrobin can mitigate salinity stress in tomato crop. Agric. Water Manag. 2019, 222, 254–264. [Google Scholar] [CrossRef]
- Venancio, W.S.; Rodrigues, M.A.T.; Begliomini, E.; de Souza, N.L. Physiological effects of strobilurin fungicides on plants. UEPG Ciência Exatas Terra Ciênc. Agrar. Eng. Ponta Grossa 2003, 9, 59–68. [Google Scholar]
- Bonasia, A.; Conversa, G.; Lazzizzera, C.; Elia, A. Pre-harvest nitrogen and Azoxystrobin application enhances postharvest shelf-life in Butterhead lettuce. Postharvest Biol. Technol. 2013, 85, 67–76. [Google Scholar] [CrossRef]
- Conversa, G.; Bonasia, A.; Lazzizzera, C.; Elia, A. Pre-harvest nitrogen and Azoxystrobin application enhances raw product quality and post-harvest shelf-life of baby spinach (Spinacia oleracea L.). J. Sci. Food Agric. 2014, 94, 3263–6272. [Google Scholar] [CrossRef] [PubMed]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Ronga, D.; Biazzi, E.; Parati, K.; Carminati, D.; Carminati, E.; Tava, A. Microalgal biostimulants and biofertilisers in crop productions. Agronomy 2019, 9, 192. [Google Scholar] [CrossRef] [Green Version]
- Campobenedetto, C.; Agliassa, C.; Mannino, G.; Vigliante, I.; Contartese, V.; Secchi, F.; Bertea, C.M. A biostimulant based on seaweed (Ascophyllum nodosum and Laminaria digitata) and yeast extracts mitigates water stress effects on tomato (Solanum lycopersicum L.). Agriculture 2021, 11, 557. [Google Scholar] [CrossRef]
- Drobek, M.; Fraç, M.; Cybulska, J. Plant Biostimulants: Importance of the Quality and Yield of Horticultural Crops and the Improvement of Plant Tolerance to Abiotic Stress—A Review. Agronomy 2019, 9, 335. [Google Scholar] [CrossRef] [Green Version]
- Cassi, F.; Viviano, L. I Suoli della Basilicata-Carta pedologica della Regione Basilicata in scala 1:250.000. In The Soils of Basilica-ta-Soil Map of the Basilicata Region in Scale 1:250,000; Reg. Basilicata-Dip. Agric. Svilup. Rurale. Dir. Gen.: Potenza, Italy, 2006; 343p. [Google Scholar]
- Cantore, V.; Iovino, F.; Pontecorvo, G. Aspetti climatici e zone fitoclimatiche della Basilicata. In Climatic Aspects and Phytoclimatic Zones of Basilicata; CNR-IEIF: Cosenza, Italy, 1987; Volume 2, 49p. [Google Scholar]
- Sergio, L.; Boari, F.; Pieralice, M.; Linsalata, V.; Cantore, V.; Di Venere, D. Bioactive phenolics and antioxidant capacity of some wild edible greens as affected by different cooking treatments. Foods 2020, 9, 1320. [Google Scholar] [CrossRef]
- Sergio, L.; Gonnella, M.; Renna, M.; Linsalata, V.; Gatto, M.A.; Boari, F.; Di Venere, D. Biochemical traits of asparagus cultivars and quality changes in two differently coloured genotypes during cold storage. LWT 2019, 101, 427–434. [Google Scholar] [CrossRef]
- Bonasia, A.; Conversa, G.; Gonnella, M.; Serio, F.; Santamaria, P. Effects of ammonium and nitrate nutrition on yield and quality in endive. J. Hortic. Sci. Biotechnol. 2008, 83, 64–70. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophyll a and chlorophyll b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Amodio, M.L.; Derossi, A.; Mastrandrea, L.; Colelli, G. A study of the estimated shelf life of fresh rocket using a non-linear model. J. Food Eng. 2015, 150, 19–28. [Google Scholar] [CrossRef]
- CIE. Colorimetry, 2nd ed.; Central Bureau of the Commission Internationale the L’Eclairage: Vienne, France, 1986. [Google Scholar]
- Schiattone, M.I.; Viggiani, R.; Di Venere, D.; Sergio, L.; Cantore, V.; Todorovic, M.; Perniola, M.; Candido, V. Impact of irrigation regime and nitrogen rate on yield, quality and water use efficiency of wild rocket under greenhouse conditions. Sci. Hortic. 2018, 229, 182–192. [Google Scholar] [CrossRef]
- Sharma, L.K.; Bali, S.K. A review of methods to improve nitrogen use efficiency in agriculture. Sustainability 2018, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Li, S. Effects of nitrogen and phosphorus fertilization on plant growth and nitrate accumulation in vegetables. J. Plant Nutr. 2004, 27, 539–556. [Google Scholar] [CrossRef]
- Zhang, J.; Sha, Z.; Zhang, Y.; Bei, Z.; Cao, L. The effects of di_erent water and nitrogen levels on yield, water and nitrogen utilization effciencies of spinach (Spinacia oleracea L.). Can. J. Plant Sci. 2015, 95, 671–679. [Google Scholar] [CrossRef]
- van Dingenen, J.; Antoniou, C.; Filippou, P.; Pollier, J.; Gonzalez, N.; Dhondt, S.; Goossens, A.; Fotopoulos, V.; Inzé, D. Strobilurins as growth-promoting compounds: How Stroby regulates Arabidopsis leaf growth. Plant Cell Environ. 2017, 40, 1748–1760. [Google Scholar] [CrossRef]
- Liang, S.; Xu, X.; Lu, Z. Effect of Azoxystrobin fungicide on the physiological and biochemical indices and ginsenoside contents of ginseng leaves. J. Ginseng Res. 2018, 42, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Taiz, L.; Zeiger, E. Plant Physiology; The Benjamin/Cummings Publishing Company, Inc.: Redwood City, CA, USA; Berlin, Germany, 2002; 675p. [Google Scholar]
- Chbani, A.; Majed, S.; Mawlawi, H.; Kammoun, M. The use of seaweed as a bio-fertilizer: Does it influence Proline and Chlorophyll concentration in plants treated? Arab. J. Med. Aromat. Plants 2015, 1, 67–77. [Google Scholar]
- Sundqvist, C.; Bjorn, L.O.; Virgin, H.I. Factors in chloroplast differentiation. In Results and Problems in Cell Differentiation—Chloroplast; Reinert, J., Heidelberg, D.E., Eds.; Springer: Berlin/Heidelberg, Germany, 1980; pp. 202–224. [Google Scholar]
- Abeles, G.B.; Morgan, P.W.; Saltveit, M.E. Ethylene in Plant Biology; Academic Press: San Diego, CA, USA, 1982. [Google Scholar]
- Koyama, R.; Itoh, H.; Kimura, S.; Morioka, A.; Uno, Y. Augmentation of antioxidant constituents by drought stress to roots in leafy vegetables. HortTecnology 2012, 22, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Luoh, J.W.; Begg, C.B.; Symonds, R.C.; Ledesma, D.; Yang, R.-Y. Nutritional yield of african indigenous vegetables in wa-ter-deficient and water-sufficient conditions. Food Nutr. Sci. 2014, 5, 812–822. [Google Scholar]
- Devienne-Barret, F.; Justes, E.; Machet, J.M.; Mary, B. Integrated control of nitrate uptake by crop growth rate and soil nitrate availability under field conditions. Ann. Bot. 2000, 86, 995–1005. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.-M.; Wang, Z.-H.; Li, S.-X.; Wang, G.-X.; Song, H.-X.; Wang, X.-N. Effects of nitrate supply on plant growth, nitrate accumulation, metabolic nitrate concentration and nitrate reductase activity in three leafy vegetables. Plant Sci. 2004, 167, 635–643. [Google Scholar] [CrossRef]
- Weightman, R.M.; Huckle, A.J.; Roques, S.E.; Ginsburg, D.; Dyer, C.J. Factors influencing tissue nitrate concentration in field-grown wild rocket (Diplotaxis tenuifolia) in southern England. Food Addit. Contam. Part A 2012, 29, 1425–1435. [Google Scholar] [CrossRef]
- Joshi, J.; Sharma, S.; Guruprasad, K.N. Foliar application of Pyraclostrobin fungicide enhances the growth, rhizobial-nodule formation and nitrogenase activity in soybean (var. JS-335). Pestic. Biochem. Physiol. 2014, 114, 61–66. [Google Scholar] [CrossRef]
- Abd El-Baky, H.H.; Hussein, M.M.; El-Baroty, G.S. Algal extracts improve antioxidant defense abilities and salt tolerance of wheat plant irrigated with sea water. Afr. J. Biochem. Res. 2008, 2, 151–164. [Google Scholar]
- Jannin, L.; Arkoun, M.; Etienne, P.; Laîné, P.; Goux, D.; Garnica, M.; Fuentes, M.; Francisco, S.S.; Baigorri, R.; Cruz, F.; et al. Brassica napus growth is promoted by Ascophyllum nodosum (L.) Le Jol. seaweed extract: Microarray analysis and physiological characterization of N, C, and S me-tabolisms. J. Plant Growth Regul. 2013, 32, 31–52. [Google Scholar] [CrossRef]
- Siomos, A.S.; Papadopoulou, P.P.; Dogras, C.C.; Vasiliadis, E.; Dosas, A.; Georgiou, N. Lettuce composition as affected by genotype and leaf position. Acta Hortic. 2002, 579, 635–639. [Google Scholar] [CrossRef]
- Konstantopoulou, E.; Kapotis, G.; Salachas, G.; Petropoulos, S.A.; Karapanos, I.C.; Passam, H.C. Nutritional quality of greenhouse lettuce at harvest and after storage in relation to N application and cultivation season. Sci. Hortic. 2010, 125, 93–94. [Google Scholar] [CrossRef]
- Fukumoto, L.R.; Toivonen, P.M.A.; Delaquis, P.J. Effect of wash water temperature and chlorination on phenolic metabolism and browning of stored iceberg lettuce photosynthetic vascular tissues. J. Agric. Food Chem. 2002, 50, 4503–4511. [Google Scholar] [CrossRef]
- Bulgari, R.; Baldi, A.; Ferrante, A.; Lenzi, A. Yield and quality of basil, Swiss chard, and rocket microgreens grown in a hy-droponic system. New Zeal. J. Crop. Hortic. Sci. 2017, 45, 119–129. [Google Scholar] [CrossRef]
- Hodges, D.M.; Forney, C.F.; Wismer, W. Processing Line Effects on Storage Attributes of Fresh-cut Spinach Leaves. HortScience 2000, 35, 1308–1311. [Google Scholar] [CrossRef] [Green Version]
- Toivonen, P.M.A.; Sweeney, M. Differences in Chlorophyll Loss at 13 °C for Two Broccoli (Brassica oleracea L.) Cultivars As-sociated with Antioxidant Enzyme Activities. J. Agric. Food Chem. 1998, 46, 20–24. [Google Scholar] [CrossRef]
- Aguero, M.V.; Barg, M.V.; Yommi, A.; Camelo, A.; Roura, S.I. Postharvest changes in water status and chlorophyll content of lettuce (Lactuca sativa L.) and their relationship with overall visual quality. J. Food Sci. 2008, 73, S47–S55. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.Y.; Chen, Y.H.; Chao, P.Y.; Chen, C.M.; Hsiehm, L.L.; Hu, S.P. Naturally occurring chlorophyll derivatives inhibit aflatoxin B1-DNA adduct formation in hepatoma cells. Mutat Res.—Genet. Toxicol. Environ. Mutagen. 2008, 657, 98–104. [Google Scholar] [CrossRef]
- Ferruzzi, M.G.; Bohm, V.; Courtney, P.D.; Schwartz, S.J. Antioxidant and antimutagenic activity of dietary chlorophyll deri-vates determined by radical scavenging and bacterial reverse mutagenesis assays. J. Food Sci. 2002, 67, 2589–2595. [Google Scholar] [CrossRef]
- Ferruzzi, M.G.; Blakeslee, J. Digestion, absorption, and cancer preventative activity of dietary chlorophyll derivatives. Nutr. Res. 2007, 27, 1–12. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | MY | DM | Ni | TP | TAA | TCh | TC |
|---|---|---|---|---|---|---|---|
| (kg m−2) | (g 100 g−1 FW) | (mg kg−1 FW) | (mg CAE g−1 DM) | (g Trolox 100 g−1 DM) | (μg g−1 DM) | (μg g−1 DM) | |
| N level (NL) | * | ns | ns | * | * | * | ** |
| NL | 1.47 | 8.6 | 4398.0 | 14.5 | 0.43 | 825.6 | 175.2 |
| NH | 1.64 | 8.6 | 4374.4 | 12.9 | 0.35 | 931.8 | 147.7 |
| Azoxystrobin (Azo) | * | ns | ns | ns | ns | * | ** |
| C | 1.48 | 8.5 | 4438.1 | 13.6 | 0.40 | 790.7 | 149.8 |
| Azo+ | 1.63 | 8.7 | 4334.7 | 13.8 | 0.38 | 966.7 | 173.1 |
| NL x Azo | ns | ns | ns | ns | ns | ns | ns |
| Treatments | MY | DM | Ni | TP | TAA | TCh | TC |
|---|---|---|---|---|---|---|---|
| (kg m−2) | (g 100 g−1 FW) | (mg kg−1 FW) | (mg CAE g−1 DM) | (g Trolox 100 g−1 DM) | (μg g−1 DM) | (μg g−1 DM) | |
| N level (NL) | * | ns | ** | ns | * | * | * |
| NL | 1.36 | 8.7 | 3200.1 | 14.8 | 0.53 | 779.8 | 192.3 |
| NH | 1.71 | 8.2 | 4399.9 | 13.9 | 0.39 | 900.2 | 176.7 |
| Biostimulants (BS) | ** | ns | * | ns | * | * | * |
| C | 1.41 b | 8.7 | 4100.0 a | 14.2 | 0.51 a | 771.2 b | 175.4 b |
| Azo+ | 1.74 a | 8.4 | 3400.5 b | 14.9 | 0.54 a | 889.4 a | 194.0 a |
| YBA+ | 1.46 b | 8.5 | 3899.5 a | 14.0 | 0.33 b | 859.2 a | 184.6 ab |
| NL x BS | ns | ns | ns | ns | ns | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schiattone, M.I.; Boari, F.; Cantore, V.; Castronuovo, D.; Denora, M.; Di Venere, D.; Perniola, M.; Renna, M.; Sergio, L.; Candido, V. Effects of Nitrogen, Azoxystrobin and a Biostimulant Based on Brown Algae and Yeast on Wild Rocket Features at Harvest and During Storage. Agronomy 2021, 11, 2326. https://doi.org/10.3390/agronomy11112326
Schiattone MI, Boari F, Cantore V, Castronuovo D, Denora M, Di Venere D, Perniola M, Renna M, Sergio L, Candido V. Effects of Nitrogen, Azoxystrobin and a Biostimulant Based on Brown Algae and Yeast on Wild Rocket Features at Harvest and During Storage. Agronomy. 2021; 11(11):2326. https://doi.org/10.3390/agronomy11112326
Chicago/Turabian StyleSchiattone, Maria Immacolata, Francesca Boari, Vito Cantore, Donato Castronuovo, Michele Denora, Donato Di Venere, Michele Perniola, Massimiliano Renna, Lucrezia Sergio, and Vincenzo Candido. 2021. "Effects of Nitrogen, Azoxystrobin and a Biostimulant Based on Brown Algae and Yeast on Wild Rocket Features at Harvest and During Storage" Agronomy 11, no. 11: 2326. https://doi.org/10.3390/agronomy11112326
APA StyleSchiattone, M. I., Boari, F., Cantore, V., Castronuovo, D., Denora, M., Di Venere, D., Perniola, M., Renna, M., Sergio, L., & Candido, V. (2021). Effects of Nitrogen, Azoxystrobin and a Biostimulant Based on Brown Algae and Yeast on Wild Rocket Features at Harvest and During Storage. Agronomy, 11(11), 2326. https://doi.org/10.3390/agronomy11112326