
Field-Evolved Sulfoxaflor Resistance of Three Wheat Aphid Species in China





Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects and Insecticides
2.2. Bioassays
2.3. Data Analysis
3. Results
3.1. Susceptibility of S. miscanthi Field Populations to Sulfoxaflor
3.2. Susceptibility of R. padi Field Populations to Sulfoxaflor
3.3. Susceptibility of M. dirhodum Field Populations to Sulfoxaflor
3.4. Differences in the Toxicity of Sulfoxaflor among Various Wheat Aphid Field Populations
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, Y.; Loso, M.R.; Watson, G.B.; Sparks, T.C.; Rogers, R.B.; Huang, J.X.; Gerwick, B.C.; Babcock, J.M.; Kelley, D.; Hegde, V.B.; et al. Discovery and characterization of sulfoxaflor, a novel insecticide targeting sap-feeding pests. J. Agric. Food Chem. 2011, 59, 2950–2957. [Google Scholar] [CrossRef] [PubMed]
- Babcock, J.M.; Gerwick, C.B.; Huang, J.X.; Loso, M.R.; Nakamura, G.; Nolting, S.P.; Rogers, R.B.; Sparks, T.C.; Thomas, J.; Watson, G.B.; et al. Biological characterization of sulfoxaflor, a novel insecticide. Pest Manag. Sci. 2011, 67, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Sparks, T.C.; Watson, G.B.; Loso, M.R.; Geng, C.; Babcock, J.M.; Thomas, J.D. Sulfoxaflor and the sulfoximine insecticides: Chemistry, mode of action and basis for efficacy on resistant insects. Pestic. Biochem. Physiol. 2013, 107, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparks, T.C.; DeBoer, G.J.; Wang, N.X.; Hasler, J.M.; Loso, M.R.; Watson, G.B. Differential metabolism of sulfoximine and neonicotinoid insecticides by Drosophila melanogaster monooxygenase CYP6G1. Pestic. Biochem. Physiol. 2012, 103, 159–165. [Google Scholar] [CrossRef]
- Gore, J.; Cook, D.; Catchot, A.; Leonard, B.R.; Stewart, S.D.; Lorenz, G.; Kerns, D. Cotton aphid (Heteroptera: Aphididae) susceptibility to commercial and experimental insecticides in the southern United States. J. Econ. Entomol. 2013, 106, 1430–1439. [Google Scholar] [CrossRef] [PubMed]
- Longhurst, C.; Babcock, J.M.; Denholm, I.; Gorman, K.; Thomas, J.D.; Sparks, T.C. Cross-resistance relationships of the sulfoximine insecticide sulfoxaflor with neonicotinoids and other insecticides in the whiteflies Bemisia tabaci and Trialeurodes vaporariorum. Pest Manag. Sci. 2013, 69, 809–813. [Google Scholar] [CrossRef] [PubMed]
- Gong, P.; Li, X.; Wang, C.; Zhu, S.; Li, Q.; Zhang, Y.; Li, X.; Li, G.; Liu, E.; Gao, H.; et al. The Sensitivity of Field Populations of Metopolophium dirhodum (Walker) (Hemiptera: Aphididae) to Seven Insecticides in Northern China. Agronomy 2021, 11, 1556. [Google Scholar] [CrossRef]
- Gong, P.; Li, X.; Gao, H.; Wang, C.; Li, M.; Zhang, Y.; Li, X.; Liu, E.; Zhu, X. Field evolved resistance to pyrethroids, neonicotinoids, organophosphates and macrolides in Rhopalosiphum padi (Linnaeus) and Sitobion avenae (Fabricius) from China. Chemosphere 2021, 269, 128747. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Gao, X. Multiple mechanisms responsible for differential susceptibilities of Sitobion avenae (Fabricius) and Rhopalosiphum padi (Linnaeus) to pirimicarb. Bull. Entomol. Res. 2009, 99, 611–617. [Google Scholar] [CrossRef]
- Lu, Y.H.; He, Y.P.; Gao, X.W. Comparative studies on acetylcholinesterase characteristics between the aphids, Sitobion avenae and Rhopalosiphum padi. J. Insect Sci. 2013, 13, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, T.; Chen, H.; Sun, Y.; Yu, X.; Xia, L. RNA Interference of the Ecdysone Receptor Genes EcR and USP in Grain Aphid (Sitobion avenae F.) Affects Its Survival and Fecundity upon Feeding on Wheat Plants. Int. J. Mol. Sci. 2016, 17, 2098. [Google Scholar] [CrossRef] [Green Version]
- Xiao, D.; Yang, T.; Desneux, N.; Han, P.; Gao, X. Assessment of Sublethal and Transgenerational Effects of Pirimicarb on the Wheat Aphids Rhopalosiphum padi and Sitobion avenae. PLoS ONE 2015, 10, e0128936. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.S.; Liu, Y.J.; Wang, Y.H.; Wang, Z.; Yu, X.L.; Wang, B.; Zhang, G.S.; Liu, X.F.; Hu, Z.Q.; Zhao, H.Y.; et al. Resistance of wheat accessions to the English grain aphid Sitobion avenae. PLoS ONE 2016, 11, e0156158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, Y.; Wang, K.; Zhang, M.; Peng, X.; Piñero, J.C.; Chen, M. Regional Susceptibilities of Rhopalosiphum padi(Hemiptera: Aphididae) to Ten Insecticides. Fla. Entomol. 2016, 99, 269–275. [Google Scholar] [CrossRef]
- Zhang, L.; Lu, H.; Guo, K.; Yao, S.; Cui, F. Insecticide resistance status and detoxification enzymes of wheat aphids Sitobion avenae and Rhopalosiphum padi. Sci. China Life Sci. 2017, 60, 927–930. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Mao, K.; Ali, E.; Zhang, X.; Wan, H.; Li, J. Temporal variability and resistance correlation of sulfoxaflor susceptibility among Chinese populations of the brown planthopper Nilaparvata lugens (Stål). Crop Protect. 2017, 102, 141–146. [Google Scholar] [CrossRef]
- Ma, K.; Tang, Q.; Xia, J.; Lv, N.; Gao, X. Fitness costs of sulfoxaflor resistance in the cotton aphid, Aphis gossypii Glover. Pestic. Biochem. Physiol. 2019, 158, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Gong, P.; Chen, D.; Wang, C.; Li, M.; Li, X.; Zhang, Y.; Li, X.; Zhu, X. Susceptibility of Four Species of Aphids in Wheat to Seven Insecticides and Its Relationship to Detoxifying Enzymes. Front. Physiol. 2020, 11, 623612. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, M.W.; Park, R.M.; Bailey, A.J. Comparing median lethal concentration values using confidence interval overlap or ratio tests. Environ. Toxic. Chem. 2006, 25, 1441–1444. [Google Scholar] [CrossRef] [PubMed]
- Deng, J. The Detection and Midgut Intracellular Location of Rickettsia Symbiont in the Camellia Aphid (Aphis aurantii). Appl. Ecol. Environ. Res. 2019, 17, 12203–12212. [Google Scholar] [CrossRef]
- Rosenheim, J.A.; Johnson, M.W.; Mau, R.F.L.; Welter, S.C.; Tabashnik, B.E. Biochemical Peradaptations, Founder Events, and the Evolution of Resistance in Arthropods. J. Econ. Entomol. 1996, 89, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Sun, J.; Qin, Y.; Fan, J.; Zhang, Y.; Tan, X.; Hou, M.; Chen, J. Reduced insecticide susceptibility of the wheat aphid Sitobion miscanthi after infection by the secondary bacterial symbiont Hamiltonella defensa. Pest Manag. Sci. 2021, 77, 1936–1944. [Google Scholar] [CrossRef] [PubMed]
- Cai, T.; Zhang, Y.; Liu, Y.; Deng, X.; He, S.; Li, J.; Wan, H. Wolbachia enhances expression of NlCYP4CE1 in Nilaparvata lugens in response to imidacloprid stress. Insect Sci. 2021, 28, 355–362. [Google Scholar] [CrossRef]
- Zhang, Y.; Cai, T.; Ren, Z.; Liu, Y.; Yuan, M.; Cai, Y.; Yu, C.; Shu, R.; He, S.; Li, J.; et al. Decline in symbiont-dependent host detoxification metabolism contributes to increased insecticide susceptibility of insects under high temperature. ISME. J. 2021. [Google Scholar] [CrossRef] [PubMed]
- Dermauw, W.; Pym, A.; Bass, C.; Van Leeuwen, T.; Feyereisen, R. Does host plant adaptation lead to pesticide resistance in generalist herbivores? Curr. Opin. Insect Sci. 2018, 26, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, A.; Desneux, N.; Monticelli, L.S.; Fan, Y.; Shi, X.; Guedes, R.N.C.; Gao, X. Potential for insecticide-mediated shift in ecological dominance between two competing aphid species. Chemosphere 2019, 226, 651–658. [Google Scholar] [CrossRef]
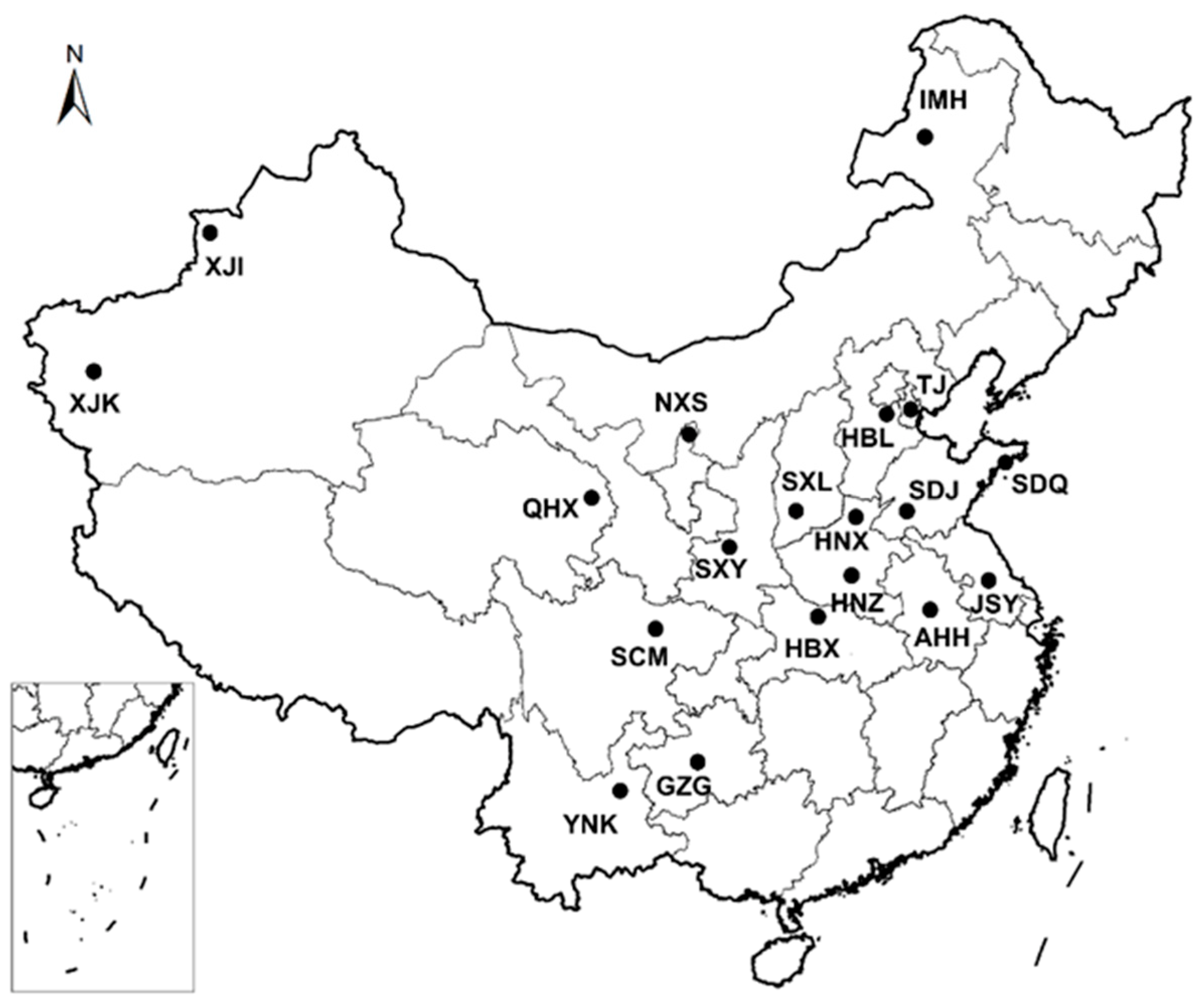

{kind=link}
| No. | Code | N a | Slope ± SE b | LC50 c | 95%CL d | χ2 | RLR e |
|---|---|---|---|---|---|---|---|
| 1 | YNK-2019 | 574 | 0.49 ± 0.09 | 443.05 | 192.13–1717.55 | 1.26 | 194.32 |
| 2 | YNK-2021 | 664 | 0.88 ± 0.08 | 252.72 | 179.64–355.16 | 0.48 | 110.84 |
| 3 | SXL-2019 | 451 | 0.64 ± 0.08 | 128.10 | 76.02–235.77 | 1.52 | 56.18 |
| 4 | SXY-2019 | 582 | 0.64 ± 0.07 | 81.76 | 25.75–289.65 | 5.82 | 35.86 |
| 5 | SXY-2021 | 866 | 1.39 ± 0.09 | 75.81 | 38.34–149.07 | 8.91 | 33.25 |
| 6 | HBX-2021 | 831 | 0.96 ± 0.07 | 60.94 | 46.31–79.29 | 1.61 | 26.73 |
| 7 | SDJ-2019 | 664 | 1.28 ± 0.15 | 58.51 | 44.71–80.62 | 1.16 | 25.66 |
| 8 | AHH-2019 | 607 | 0.79 ± 0.08 | 57.49 | 12.71–158.36 | 7.92 | 25.21 |
| 9 | IMH-2019 | 634 | 0.77 ± 0.07 | 52.17 | 16.18–139.50 | 7.07 | 22.88 |
| 10 | QHX-2019 | 595 | 0.82 ± 0.08 | 44.33 | 16.97–112.30 | 5.74 | 19.44 |
| 11 | HNZ-2019 | 418 | 0.90 ± 0.10 | 37.34 | 10.95–140.16 | 7.02 | 16.38 |
| 12 | XJK-2019 | 759 | 0.81 ± 0.08 | 32.27 | 21.07–46.93 | 1.29 | 14.15 |
| 13 | GZG-2021 | 648 | 0.90 ± 0.07 | 29.18 | 8.93–66.24 | 7.39 | 12.8 |
| 14 | HNX-2019 | 454 | 0.84 ± 0.09 | 28.83 | 18.85–43.35 | 2.42 | 12.64 |
| 15 | NXY-2019 | 535 | 0.76 ± 0.08 | 23.42 | 15.13–34.78 | 2.27 | 10.27 |
| 16 | GZG-2019 | 466 | 0.81 ± 0.08 | 22.24 | 11.27–42.62 | 3.43 | 9.75 |
| 17 | TJ-2019 | 496 | 0.68 ± 0.09 | 16.40 | 9.2–26.70 | 1.19 | 7.19 |
| 18 | SDQ-2019 | 552 | 0.69 ± 0.08 | 16.38 | 9.57–26.64 | 0.87 | 7.18 |
| 19 | HNX-2021 | 805 | 0.66 ± 0.06 | 15.44 | 8.36–24.86 | 1.07 | 6.77 |
| 20 | HBL-2021 | 555 | 0.84 ± 0.08 | 12.91 | 8.73–18.67 | 0.94 | 5.66 |
| 21 | QHX-2021 | 700 | 1.11 ± 0.08 | 12.59 | 7.21–21.72 | 4.62 | 5.52 |
| 22 | HBX-2019 | 575 | 0.56 ± 0.06 | 10.91 | 4.34–23.28 | 3.04 | 4.79 |
| 23 | AHH-2021 | 797 | 0.62 ± 0.06 | 10.73 | 3.58–22.28 | 3.10 | 4.71 |
| 24 | HBL-2019 | 332 | 0.53 ± 0.12 | 2.28 | 0.37–5.65 | 1.54 | 1 |
| No. | Code | N a | Slope ± SE b | LC50 c | 95%CL d | χ2 | RLR e |
|---|---|---|---|---|---|---|---|
| 1 | HNX-2021 | 881 | 0.82 ± 0.06 | 288.24 | 164.09–506.74 | 4.28 | 113.93 |
| 2 | IMH-2019 | 927 | 0.63 ± 0.06 | 103.68 | 44.12–315.22 | 5.88 | 40.98 |
| 3 | SDJ-2019 | 854 | 0.82 ± 0.10 | 99.45 | 68.87–155.84 | 1.64 | 39.31 |
| 4 | HBX-2021 | 964 | 0.54 ± 0.05 | 94.56 | 61.90–153.76 | 1.58 | 37.38 |
| 5 | XJK-2019 | 662 | 0.86 ± 0.08 | 93.98 | 64.19–138.55 | 0.75 | 37.15 |
| 6 | SXY-2019 | 732 | 0.60 ± 0.07 | 75.04 | 46.29–121.92 | 1.76 | 29.66 |
| 7 | QHX-2021 | 890 | 1.40 ± 0.11 | 50.99 | 21.63–93.13 | 8.64 | 20.15 |
| 8 | AHH-2021 | 670 | 1.28 ± 0.10 | 34.42 | 27.64–47.00 | 2.53 | 13.61 |
| 9 | AHH-2019 | 754 | 0.87 ± 0.07 | 30.31 | 15.02–54.19 | 4.11 | 11.98 |
| 10 | GZG-2021 | 807 | 1.054 ± 0.08 | 26.30 | 18.79–35.17 | 2.66 | 10.39 |
| 11 | SXL-2019 | 666 | 0.65 ± 0.07 | 23.74 | 7.35–54.52 | 4.12 | 9.38 |
| 12 | SCM-2019 | 656 | 0.75 ± 0.07 | 20.34 | 13.25–30.21 | 2.66 | 8.04 |
| 13 | SXL-2021 | 935 | 1.62 ± 0.12 | 16.93 | 13.85–20.32 | 1.20 | 6.69 |
| 14 | TJ-2019 | 629 | 0.74 ± 0.07 | 15.50 | 6.45–30.54 | 3.50 | 6.13 |
| 15 | QHX-2019 | 676 | 0.62 ± 0.07 | 15.13 | 9.02–23.45 | 2.05 | 5.98 |
| 16 | SXY-2021 | 820 | 1.13 ± 0.08 | 12.97 | 6.45–22.90 | 6.14 | 5.13 |
| 17 | HNZ-2019 | 909 | 0.90 ± 0.06 | 12.69 | 4.26–30.11 | 10.35 | 5.02 |
| 18 | YNK-2019 | 654 | 0.77 ± 0.07 | 12.22 | 5.07–25.24 | 4.99 | 4.83 |
| 19 | HBL-2021 | 727 | 0.95 ± 0.08 | 9.86 | 6.51–14.03 | 1.71 | 3.9 |
| 20 | NXY-2019 | 579 | 1.15 ± 0.11 | 6.74 | 4.60–9.20 | 1.26 | 2.66 |
| 21 | XJI-2021 | 639 | 1.23 ± 0.12 | 4.98 | 3.31–6.91 | 0.36 | 1.97 |
| 22 | GZG-2019 | 587 | 0.76 ± 0.08 | 4.22 | 0.57–11.52 | 6.48 | 1.67 |
| 23 | HBX-2019 | 590 | 0.79 ± 0.09 | 3.75 | 1.91–6.19 | 1.08 | 1.48 |
| 24 | JSY-2019 | 688 | 0.39 ± 0.07 | 2.53 | 0.44–6.83 | 0.07 | 1 |
| No. | Code | N a | Slope ± SE b | LC50 c | 95%CL d | χ2 | RLR e |
|---|---|---|---|---|---|---|---|
| 1 | XJI-2021 | 652 | 0.60 ± 0.08 | 251.64 | 73.00–251.44 | 1.03 | 206.26 |
| 2 | SXL-2019 | 663 | 0.60 ± 0.08 | 123.77 | 146.00–502.84 | 1.03 | 101.45 |
| 3 | SXY-2021 | 594 | 0.74 ± 0.07 | 83.86 | 38.28–183.91 | 3.90 | 68.74 |
| 4 | HBL-2019 | 864 | 1.36 ± 0.16 | 25.12 | 13.39–42.01 | 4.41 | 20.59 |
| 5 | SXY-2019 | 685 | 0.51 ± 0.06 | 15.72 | 8.02–27.11 | 2.18 | 12.89 |
| 6 | NXY-2019 | 696 | 0.88 ± 0.07 | 14.51 | 10.06–20.33 | 1.60 | 11.89 |
| 7 | GZG-2019 | 586 | 0.78 ± 0.08 | 9.12 | 2.69–1.22 | 10.4 | 7.48 |
| 8 | QHX-2019 | 493 | 0.70 ± 0.10 | 8.04 | 4.45–12.92 | 1.78 | 6.59 |
| 9 | GZG-2021 | 751 | 1.05 ± 0.23 | 1.28 | 0.14–3.18 | 0.09 | 1.05 |
| 10 | XJK-2019 | 579 | 0.44 ± 0.06 | 1.22 | 0.08–4.92 | 4.31 | 1 |
| Species | N a | ns b | + c | − d |
|---|---|---|---|---|
| S. miscanthi/R. padi | 20 | 5 | 9 | 6 |
| S. miscanthi/M. dirhodum | 9 | 3 | 5 | 1 |
| R. padi/M. dirhodum | 9 | 3 | 3 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Wang, C.; Li, Q.; Zhu, S.; Tian, X.; Zhang, Y.; Li, X.; Gao, H.; Liu, E.; Wang, L.; et al. Field-Evolved Sulfoxaflor Resistance of Three Wheat Aphid Species in China. Agronomy 2021, 11, 2325. https://doi.org/10.3390/agronomy11112325
Li X, Wang C, Li Q, Zhu S, Tian X, Zhang Y, Li X, Gao H, Liu E, Wang L, et al. Field-Evolved Sulfoxaflor Resistance of Three Wheat Aphid Species in China. Agronomy. 2021; 11(11):2325. https://doi.org/10.3390/agronomy11112325
Chicago/Turabian StyleLi, Xinan, Chao Wang, Qiuchi Li, Saige Zhu, Xujun Tian, Yunhui Zhang, Xiangrui Li, Haifeng Gao, Enliang Liu, Liande Wang, and et al. 2021. "Field-Evolved Sulfoxaflor Resistance of Three Wheat Aphid Species in China" Agronomy 11, no. 11: 2325. https://doi.org/10.3390/agronomy11112325
APA StyleLi, X., Wang, C., Li, Q., Zhu, S., Tian, X., Zhang, Y., Li, X., Gao, H., Liu, E., Wang, L., & Zhu, X. (2021). Field-Evolved Sulfoxaflor Resistance of Three Wheat Aphid Species in China. Agronomy, 11(11), 2325. https://doi.org/10.3390/agronomy11112325