
Maceration Time Affects the Efficacy of Borage Extracts as Potential Biostimulant on Rocket Salad




1
Department of Agricultural and Environmental Sciences, DiSAA, University of Milan, 20133 Milano, Italy
2
Department of Agricultural, Forest, and Food Sciences, DISAFA, Vegetable Crops and Medicinal and Aromatic Plants VEGMAP, University of Turin, 10095 Grugliasco, Italy
*
Author to whom correspondence should be addressed.
Agronomy 2021, 11(11), 2182; https://doi.org/10.3390/agronomy11112182
Submission received: 10 October 2021
/
Revised: 23 October 2021
/
Accepted: 27 October 2021
/
Published: 28 October 2021
(This article belongs to the Special Issue Toward Sustainability in Horticultural Crop Production Systems)
Abstract
:The efficacy of plant-based biostimulants lays on the extraction of bioactive compounds that can trigger positive biological responses in plants. Their use has been increasing for lowering the production costs, enhance the yield and the stress tolerance, and improve the cropping systems’ sustainability. The aim of this work was to investigate the effect of maceration time on borage extracts, subsequently sprayed on rocket plants hydroponically grown. A preliminary test was also assessed to evaluate the extracts’ effect on seeds germination. Borage leaves and flowers were separated and macerated in the same conditions for different times. The obtained borage extracts were applied two times during the cultivation of rocket plants. The yield was not affected by the treatments, while the quality of leafy vegetables was improved. The results indicated that the flower extracts (FE) were more active than leaf extracts (LE) in stimulating the crop response. Promising results were observed on the nitrate assimilation, with reduced accumulation in the edible and commercial parts. Non-destructive analysis revealed that electron transport rates were significantly higher in treated plants. Biostimulant effects were also confirmed by the lowering of sucrose and sugars content in the treated plants and by a general increment of secondary metabolites.
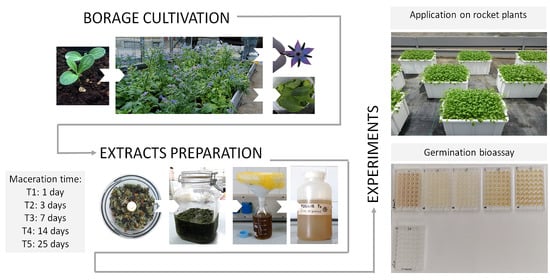
1. Introduction
Plants are a unique resource of bioactive compounds due to their structural and biological diversity, and they have been used since the ancient time for different purposes. The interest for their application showed a decline in the past years due to the synthetic approaches to drug discovery [1]. The plants’ composition might vary both in the qualitative and quantitative traits since it is influenced by environmental conditions. This aspect makes them a peculiar resource of interesting compounds. Nowadays, plant extracts are becoming increasingly popular in several areas due to their potential and because of the change in consumers’ behaviour. Indeed, in addition to the medical or pharmaceutical use, the request for plant extracts is increasing as food supplement, to preserve raw and processed foods, as beverage additive, and as cosmetics. Moreover, their market also includes the application in the agricultural sector. The interest in the application of plant extracts in this field goes along with the need to find alternative and sustainable solutions to increase crop productivity while preserving natural resources, together with the decreasing application of mineral fertilizers and pesticides. Thus, plants may become raw materials for biostimulant products or bioherbicides [2]. In plants, secondary metabolites are one of the most investigated groups of natural products and they might be responsible for the biostimulant activity. Secondary metabolites are low molecular weight organic compounds including groups like phenolics, terpenes, and nitrogen-containing compounds. They are accumulated especially in response to stresses, playing an adaptive role in plants and exerting pivotal functions in ecological interactions between plants and the surrounding environment, improving the tolerance of plants to abiotic stresses, facilitating the uptake of nutrients from the soil, and acting as signalling molecules [2,3,4]. Phenolic compounds showed a biostimulant activity on different plant species by alleviating the negative effects of stress. Probably, phenolics compounds exert positive effects in counteracting several stressful conditions through the activation of the antioxidant defence system. Similarly, terpenes and phytohormones have a positive effect both by foliar spray and seed priming. Besides the biostimulant activity on plant growth and seed germination, a large group of secondary metabolites is phytotoxic and exerts an allelopathic effect [5,6]. Allelochemicals are generally divided into classes on the base of the structure and the properties, and several of them are synthetized by the shikimate pathway. They act mainly by affecting the activity of the enzymes involved in physiological processes causing the inhibition of the germination or the plant growth. Compounds such as glucosinolates, isothiocyanates, sarmentine, artemisin, sorgoleone, leptospermonen, and momilacetone have been isolated from different plant species and showed an inhibitory potential on a wide range of sensitive weeds [6]. Due to the selection of weed species resistant to synthetic herbicides, biological molecules can be considered as a promising tool for weed management. Moreover, many compounds of natural origin with a phytotoxic effect remain unexplored. The separation of the bioactive compounds from the inactive or inert fraction can be implemented using several extraction methods and technologies [7,8,9,10]. Most of these techniques exploit the different extracting power of solvents and the application of heat, pressure, and/or mixing. Infusion, maceration, digestion, decoction, percolation, and Soxhlet extraction are the conventional extraction techniques. These methods use organic solvents such as hexane, acetone, methanol, ethanol, or water and are generally carried out under atmospheric pressure. Alternative approaches have emerged in order to mitigate limitations of the traditional ones, by reducing the extraction time and the volume of organic solvents used and to improve the yield and the bioactivities of the extracts. The choice of the method, the solvent, and the temperature depends on different factors. Moreover, the time of the extraction affects both the yield and the composition of the extract. Thus, a standardized extraction method which takes into consideration all these parameters is required in order to obtain stable and high-quality extracts. However, even if the newest extraction techniques promote the efficiency of the extraction of specific components, sometimes, the high costs make them not affordable. For example, maceration and decoction extract methods are more applicable, convenient, and economically accessible for small and medium enterprises or in developing countries, compared with the modern techniques [11,12]. Plants extracts obtained through these techniques are commonly applied in small-scale agriculture or in organic farming, given the ease of preparation or the banning using synthetic alternatives. Green approaches for promoting plant performance using natural supplementations are highly requested. Several plant-based preparations showed interesting effects in stimulating plant growth, improving quality traits, or enhancing tolerance to abiotic stresses [13,14,15,16,17,18,19]. Among them, recent studies focused on the effect of extracts from borage (Borago officinalis L.) on different leafy vegetables grown under optimal and stressful conditions [20,21,22]. Borage is an annual crop which possesses several beneficial properties and it is particularly interesting due to the high level and quality of gamma-linolenic acid (GLA) contained in the seeds. Besides, several studies have been conducted to identify different classes of compounds from borage flowers and leaves [23]. Borage leaves represent more than 60% of the plant matter and they are considered also as a low-cost crop by-product by some food processing industries [24]. Considering all these premises, it seems interesting to deepen the use of borage extracts on vegetables. Fatty acids, phospholipids, and glycolipids were found in borage leaves. (E)-(E),2-4 decadienal has been identified as the main compound of the essential oils in the borage stem and flower [25,26]. Particular attention has been paid also to the phenolic profile and antioxidant activity of borage extracts. Tannins and anthocyanins were present in low amounts in leaf extracts compared with total flavonoids and total flavonols. Rosmarinic, syringic, and sinapic acids were found both in borage leaves and seeds [27,28]. Phenolic and flavonoid were found also in borage flowers and among them, the salicylic acid and myricetin were the most abundant compounds [29]. According to several authors [30,31], small amounts of toxic pyrrolizidine alkaloids were identified in leaves, seeds, and flowers of borage.
The purpose of the present study was to evaluate the influence of different maceration times on the preparation and efficacy of aqueous extracts from borage leaves and flowers. In particular, two separate experiments focused on the different effects of the extracts when applied on leaves or seeds of rocket salad (Diplotaxis tenuifolia, L.) were performed. Rocket is a fast-growing crop, belonging to the Brassicaceae family, widely cultivated as baby leaf and popular throughout the world [32]. It is a low calories vegetable and it is considered a potential health promoting produce with several beneficial properties for human health. In fact, it is a good source of ascorbic acid, flavonoids, carotenoids, and glucosinolates [21,33]. In our study, borage extracts were applied to rocket plants for biostimulating purposes, while the allelopathic effect was evaluated on rocket seeds. The rocket plant responses have been investigated by measuring several physiological variables such as the chlorophyll content and chlorophyll a fluorescence related parameters, phenols and anthocyanin, sucrose, and total sugars. Moreover, the percentage of seed germination was evaluated in order to discriminate a biostimulant or inhibitory effect of the extracts on the same species.
2. Materials and Methods
2.1. Plant Material, Growth Conditions, and Experimental Design
2.1.1. Preparation of Borage Extracts
Borage plants were cultivated starting from January 2017 in a greenhouse at the Faculty of Agricultural and Food Sciences of Milan, under controlled conditions (natural ambient light condition, 600 µmol m−2 s−1 plus supplementary light 400 W m−2 per 12 h day−1), in plastic pots containing a peaty substrate (pH (H2O) 5.8–6.8, EC 0.30–0.50 dS/m). Specifically, the substrate is a commercial substrate containing peat (Baltic peat, superfine Irish peat, OTB Irish peat) as major component, plus bark humus and silica sand. The plants have been cultivated in order to obtain a homogeneous and standardized starting raw material. Borage plants received 7 g of a (14:7:17) N:P:K fertilizer, one time during the growing cycle after 10 weeks from seedling. Sampling was carried out starting from flowering stage (April–May) and leaves and flowers were collected separately, weighed, and stored at −20 °C until use. Borage extracts were prepared following the method used by Bulgari et al. [20] with a slight modification in the period of maceration. Plant material was roughly minced, transferred into clean glass jars (3 L) with lid, and soaked in water. The volume of the water was added in proportion to the fresh weight of plant material in a ratio of 1:2 (w/v). The jars were kept in a dark place, at room temperature (25 °C) and stirred once in a while. Four different maceration times were assayed: 1 (T1), 3 (T2), 7 (T3) and 14 (T4) days. At the end of each period, the solid residue was separated from the liquid and discarded. The aqueous phase was filtered using a syringe filter with a 0.45-µm pore size and then diluted with water (10 mL/L) to be used as plant treatments. This dilution was chosen on the basis of our previous experiments. Final extracts from leaves (LE) and from flowers (FE) were stored at −20 °C until use.
2.1.2. Chemical Characterization of the Extracts
For chemical characterization, total phenolic compounds in borage extracts were determined by the Folin–Ciocalteu’s procedure (as reported in Bulgari et al. [20]). A 100-μL aliquot of extracts was diluted with 3.90 mL of double-distilled water and combined with 250 μL of 50% (v/v) Folin–Ciocalteu’s reagent and 750 μL of saturated (20% w/v) Na2CO3. Samples were vigorously shaken and incubated for 2 h, at RT in the dark before absorbance measurement at 765 nm. Total phenolics were expressed as gallic acid equivalents (GAE; mg L–1) upon comparison with a freshly prepared gallic acid standard curve. The pH values of aqueous extracts were measured by a Crison pH-Meter GLP 21+. The electrical conductivity (EC) was determined using a conductivity meter (Delta Ohm, Padova, Italy). The chemical characterization of extracts is reported as Supplementary Table S1.
2.1.3. Rocket Cultivation
The trial was carried out at the Faculty of Agricultural and Food Science of Milan in 2018. Wild rocket plants (Diplotaxis tenuifolia, L.; ISI Sementi S.p.A., Parma, Italy) were grown hydroponically into plastic tanks (35 × 25 × 20 cm) with 10 L of a modified Hoagland medium and the concentration [mM] of nutrients in the solution used was composed by 10.6 N-NO3, 3.9 N-NH4, 1.4 P, 7 K, 2.19 Ca, 0.8 Mg and 1.8 S, and Hoagland’s concentration for micronutrients. The pH (H2O) of the nutrient solution used for soilless cultivation was between 5.5 and 6.5 and the EC was 1.3–1.5 dS m−1. Seeds of rocket were manually sown into 10 polystyrene trays filled with agri-perlite substrate in December. Each tray had 77 holes and 5 seeds were put in each hole. Plants were grown in an experimental greenhouse under controlled conditions with a range of 18–24 °C, RH 50–60%, and 1200 µmol m−2 s−1 (natural ambient light condition). Rocket plants were treated with 20 mL of water (control) and 20 mL of each borage extract (10 mL L−1). Treatments were applied as foliar spray onto leaves until run-off using a trigger pump sprayer, with a pressure of 1–3 bar, between 09:00 and 10:00 a.m. in a randomized experimental design. The application was performed twice during the growing periods: the first one, 23 days after sowing (DAS) and the second one 28 DAS (one day before the harvest). The timing of the treatments was established on the basis of several tests performed on leafy vegetables (rocket and lettuce) [20,21,22,34]. Harvesting was performed when the baby leaf commercial stage was reached. After harvest, leaves were gently rinsed with distilled water, blotted with paper towels, and stored at −20 °C.
2.2. Non-Destructive Analyses
2.2.1. Chlorophyll
Leaf chlorophyll content was estimated in vivo using a chlorophyll content meter (CL-01 Chlorophyll Content Meter, Hansatech Instruments, King’s Lynn, UK). Measurements were performed at the end of the trial, before harvesting. The instrument estimates the chlorophyll content on the basis of the absorbance at 620 and 940 nm. The results are expressed as chlorophyll index (relative units, r.u.).
2.2.2. Chlorophyll a Fluorescence
Chlorophyll a fluorescence was measured in vivo using two different instruments: a hand-portable fluorometer (Handy-PEA, Hansatech Instruments, King’s Lynn, UK) and a field portable pulse modulated chlorophyll fluorometer (FMS2, Hansatech Instruments, UK). Before all measurement with Handy-PEA, leaves were dark-adapted with leaf clips (4 mm diameter) for 30 min. Then, leaf sections were exposed to a saturating light (3000 μmol m−2 s−1) provided by an array of three high-intensity light-emitting diodes for 1 s. Chlorophyll a fluorescence was measured, and analyses was performed applying the JIP-test equations. The fluorescence parameters were calculated automatically by the used device: in particular, we considered the maximum quantum efficiency of photosystem II (Fv/Fm) or the performance index (PI). Information about the structural and functional status of photosynthetic apparatus was provided by the parameters measured (Supplementary Table S2). Modulated chlorophyll fluorescence under ambient light regime was measured using the FMS2, fluorimeter (Hansatech Instruments, UK). In order to calculate the electron transport rate (ETR), PAR value was recorded by a light sensor on the leaf-clip. The steady-state fluorescence (Fs) was measured with the measuring radiation. Subsequently, a pulse of saturating light was imposed to obtain the maximum fluorescence level in light adapted leaves (Fm’). The effective PSII quantum efficiency (ΦPSII) and the ETR were calculated by the FMS software. Measurements were carried out at the end of the trial, before harvesting.
2.3. Destructive Analyses
2.3.1. Total Fresh Biomass and Dry Matter
Yield as fresh above ground biomass was measured for one tank each condition at the end of the experiment by cutting plants at collar. The leaf dry weight was determined by oven-drying samples in 105 °C until a constant weight was reached.
2.3.2. Chlorophylls and Carotenoids
Chlorophyll a+b and carotenoids were determined in rocket leaves at harvest. Pigments were extracted using 5 mL of 99.9% (v/v) methanol. Leaf disc samples (30 mg), obtained with a 5-mm diameter cork borer, were kept in dark room for 24 h at 4 °C. Later, absorbance readings were measured at 665.2 and 652.4 nm for chlorophylls and 470 nm for total carotenoids. Pigments levels were calculated by Lichtenthaler’s formula and expressed on a fresh weight basis [35].
2.3.3. Spectrophotometric Phenol and Anthocyanin Determination
Total phenols and anthocyanin were extracted in 3 mL of methanol acidified with hydrochloric acid. Leaf disc samples (30 mg), obtained with a 5 mm diameter cork borer, were kept in dark room for 24 h at 4 °C. Then, absorbance readings were measured at 320 nm for total phenols, and at 535 nm for anthocyanin [36]. Phenolic index was expressed as Abs320 nm g−1 FW. Anthocyanin concentrations were expressed in cyanidin-3-glucoside equivalents using a molar extinction coefficient (ε) of 29,600 L M−1 cm−1.
2.3.4. Nitrate Concentration
Nitrate concentration was determined by the method of Cataldo et al. [37]. Fresh leaf tissue was homogenized in distilled water (1 g fresh tissue per 4 mL distilled water). The homogenate was centrifuged at 4000 rpm for 15 min at RT (ALC centrifuge-model PK130R) and the recovered supernatant was used for the colorimetric analysis. Twenty microliters of the extract were added to 80 mL of 5% (w/v) salicylic acid in concentrated H2SO4 (SA + H2SO4). Afterward, 3 mL of 1.5 N NaOH were added. The samples were cooled to RT and absorbance at 410 nm was measured with a spectrophotometer. Nitrate content was calculated referring to a KNO3 standard calibration curve. Nitrate concentration was expressed as mg of NO3-N per kg of fresh weight.
2.3.5. Sucrose and Total Sugars
Sucrose content was measured using the resorcinol method [38]. Approximately 1 g of leaf tissue was homogenized in a mortar with 3 mL of water. The mixture was centrifuged at 4000 rpm for 15 min at RT. Sucrose assay was performed by mixing 0.2 mL of supernatant with 0.2 mL of 2 N NaOH and incubated in a water bath at 100 °C for 10 min, then 1.5 mL of resorcinol buffer (containing 30% hydrochloric acid, 1.2 mM resorcinol, 4.1 mM thiourea 1.5 M acetic acid) was added to samples and incubated in a water bath at 80 °C for 10 min. After cooling at RT, the optical density was determined spectrophotometrically at 500 nm and the sucrose concentration was calculated using a standard curve. The total sugars were determined on the same extract using the anthrone method with slight modifications [39]. The anthrone reagent (10.3 mM) was prepared dissolving anthrone in 95% H2SO4. The reagent was left to stand for 30–40 min before use, 0.5 mL extract was placed on top of 2.5 mL of the anthrone reagent incubated in ice for 5 min, and then vortexed vigorously. The tubes were heated to 95 °C for 10 min and left to cool in ice. Readings were performed at 620 nm. Calibration curve was carried out using a glucose standard solution.
2.4. Germination Bioassay
Seeds of wild rocket (Diplotaxis tenuifolia L.) were selected as test plant species and the experiment was performed in microtiter plates, positioning a single seed per well for a total of 24 seeds each treatment. Borage flowers and leaves extracts, obtained after different maceration times, were applied and their effect were compared with the borage extracts used in previous studies obtained with 25 days (T5) of maceration [20,21,22] and with distilled water as control. A volume of 250 µL of each treatment was added to each well and the plates were incubated in the dark at 20 °C. After 10 days, plates were examined and the percentage of seeds germinated was recorded.
2.5. Statistical Analysis
Statistical analysis was performed with GraphPad Prism 6 (GraphPad Software Inc.; San Diego, CA, USA) and data were subjected to one-way ANOVA (except for the experiment on germination bioassay). Differences among means were determined by Tuckey post-test (p < 0.05). If normality assumption for ANOVA was violated, the Kruskal–Wallis test was applied.
3. Results
3.1. Total Fresh Biomass and Dry Matter Percentage
Fresh biomass was measured at harvest and yield was calculated as grams of fresh weight per square meter. Highest and lowest values were reached by the plants treated with the LE (T1) and with the FE (T4), respectively. The values were 3027 g m−2 and 1947 g m−2, respectively (Table 1). Regarding the dry matter percentage, the FE (T4) treatment showed the highest percentage (18%) while the LE (T1) reached the lowest value of 14% (Table 1).
3.2. Chlorophyll
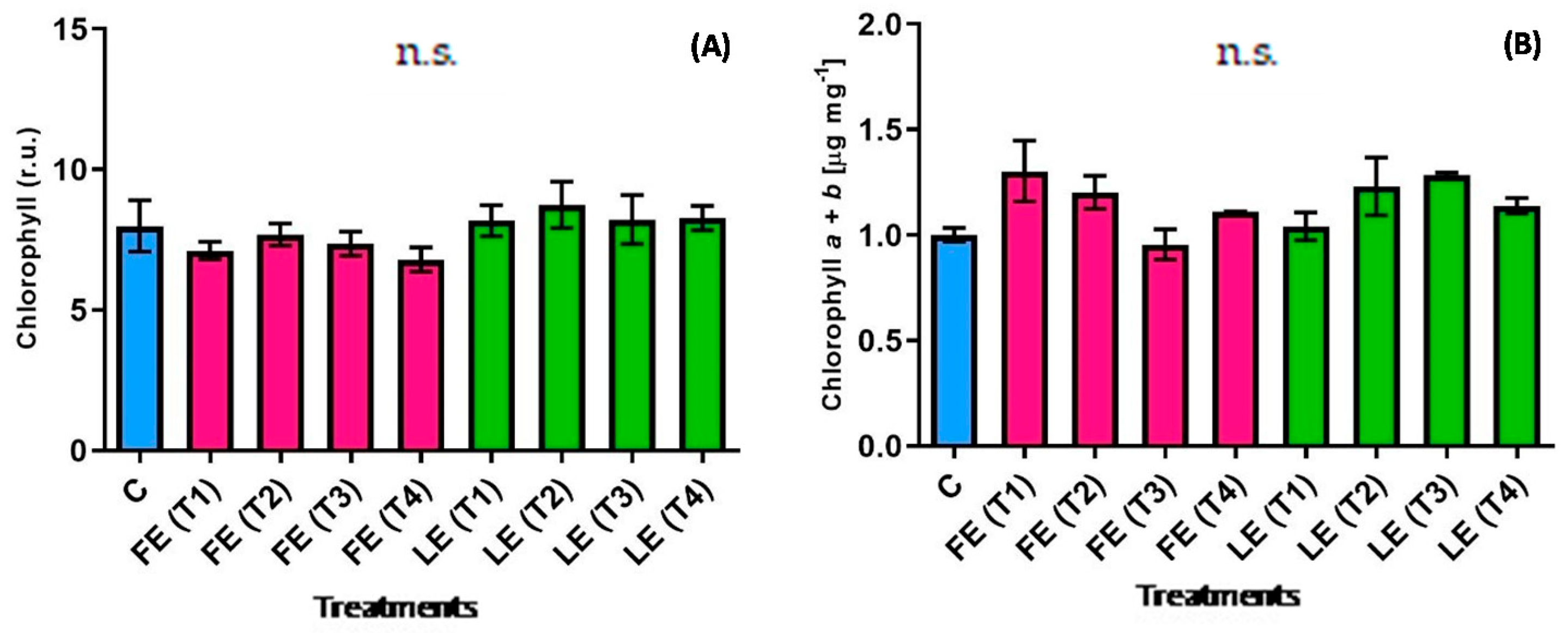
Leaf chlorophyll concentration is an important quality parameter since it has a direct effect on the vegetable visual appearance. Figure 1A,B show the chlorophyll content estimated in vivo and the concentration of chlorophyll a+b measured by the destructive method. Although results were obtained from the two different determinations, the leaf extract (LE) induced a higher accumulation in T2 and T3, even if not in a significant way (Figure 1). Indeed, in LE-and FE-treated plants all chlorophyll values expressed as relative units were similar regardless of period of maceration. Instead, the destructive analysis highlighted two different patterns between the samples treated with FE and LE. FE treatments led to an initial decrease from (T1) to (T3), followed by a final increase at (T4). An opposite trend resulted in plants treated with LE. A significant interaction between the maceration time and the plant organ used for the preparation resulted in the chlorophyll a + b; however, statistical analysis did not show any significant difference among treatments (Figure 1).
3.3. Total Carotenoids
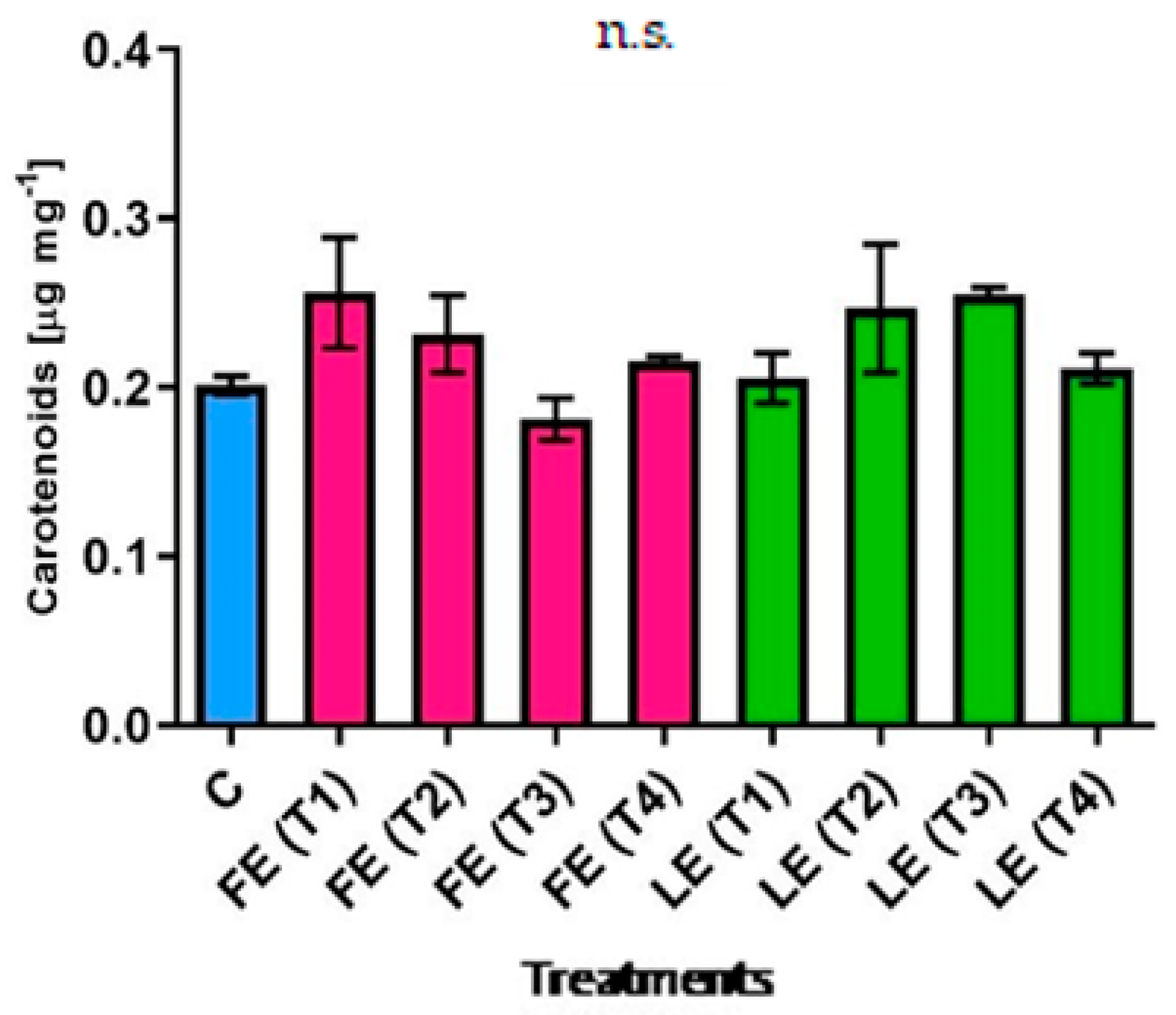
The concentration of carotenoids (Figure 2) in rocket leaves followed the trend of chlorophyll a+b levels. Different patterns were observed comparing the effect of FE and LE. The highest (0.256 µg mg−1 FW) and the lowest (0.181 µg mg−1 FW) average values were observed in plants treated with FE (T1) and FE (T2) respectively, but no significant difference resulted between these two treatments.
3.4. Phenols and Anthocyanin
Phenols and anthocyanins represent antioxidant compounds that can have positive effects on plant ecophysiology under stress conditions and also important parameters as nutritional value in vegetables. Table 2 lists the phenols and anthocyanin concentrations measured in leaf tissues at harvest. The phenol index (expressed as Abs320 nm g−1) in control plants was around 19.7 and all borage extracts generally induced an increase in this parameter, mostly at (T2) maceration time. In fact, the highest averages were observed in samples treated with FE (T2) and LE (T2); these values were statistically relevant compared to the control values (Table 2). The concentration of anthocyanin in rocket leaves showed a similar trend.
3.5. Chlorophyll a Fluorescence
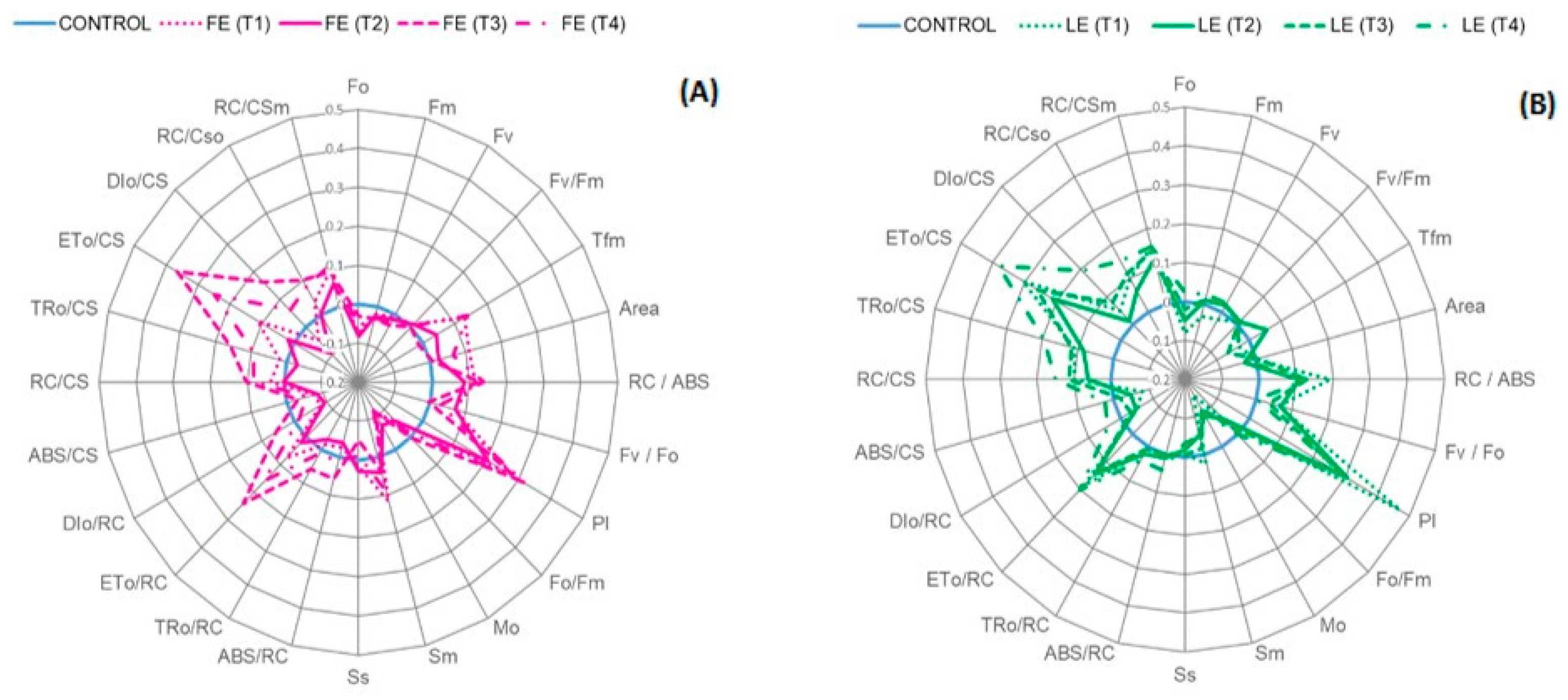
Figure 3A,B represents the effect of the FE and LE on some of the chlorophyll a fluorescence parameters. The application of borage treatments induced a modification in some of these parameters, as shown by the distance of the pink and green lines from the reference blue line. In particular, both the flower and leaf extracts slightly induced the performance index (PI). The control value was around 1.99 and the highest point (2.94) was reached in plants treated with the LE (T1). The maximum quantum efficiency of PSII (Fv/Fm ratio) was not altered by the treatments and values were always higher than 0.83, usually considered as the optimal value of a healthy plant. The electron transport flux per cross section (ETo/CS) was positively affected by the borage treatments, regardless of the nature of the raw material. It was generally higher in plants treated with the FE and LE obtained with a longer maceration time (T3) and (T4). In particular, the value increased (+38%) in plants treated with LE (T4). Furthermore, the density of PSII active reaction centres at tmax, (RC/CSm), increased by +9% in FE treated plants and by +14% in LE treated plants when compared to the control. Moreover, the trapped energy flux per cross section (TRo/CS) increased (+19.4%) in response to LE (T4) treatment. TFm represents the time needed to reach the maximum fluorescence intensity. The average value of control leaves was around 230 milliseconds; FE treatments induced a slight decrease from (T1) to (T3) and a final pick at (T4). In particular, (T1) and (T4) time points were higher than control by +13% and +19.6%, respectively. The highest value (275 ms) was observed in plants treated with FE (T4), while the lowest value (215 ms) resulted in plants treated with LE (T3).
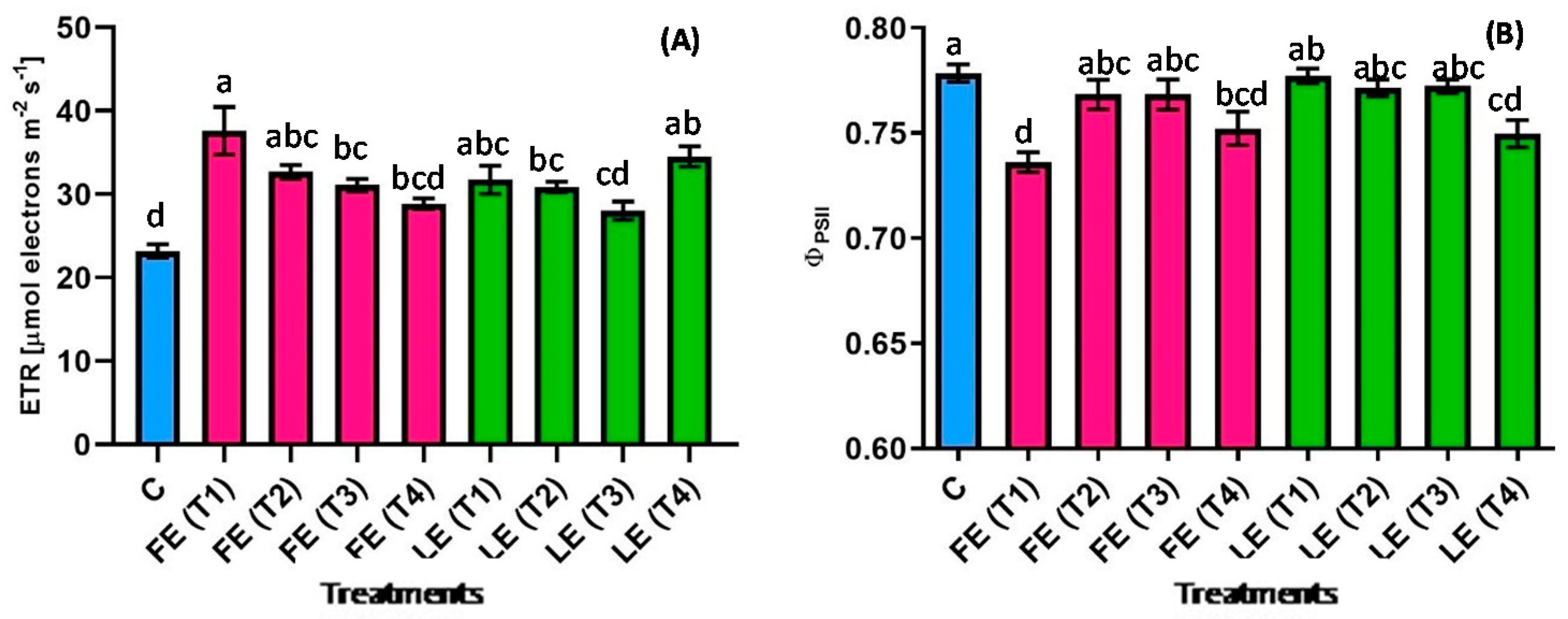
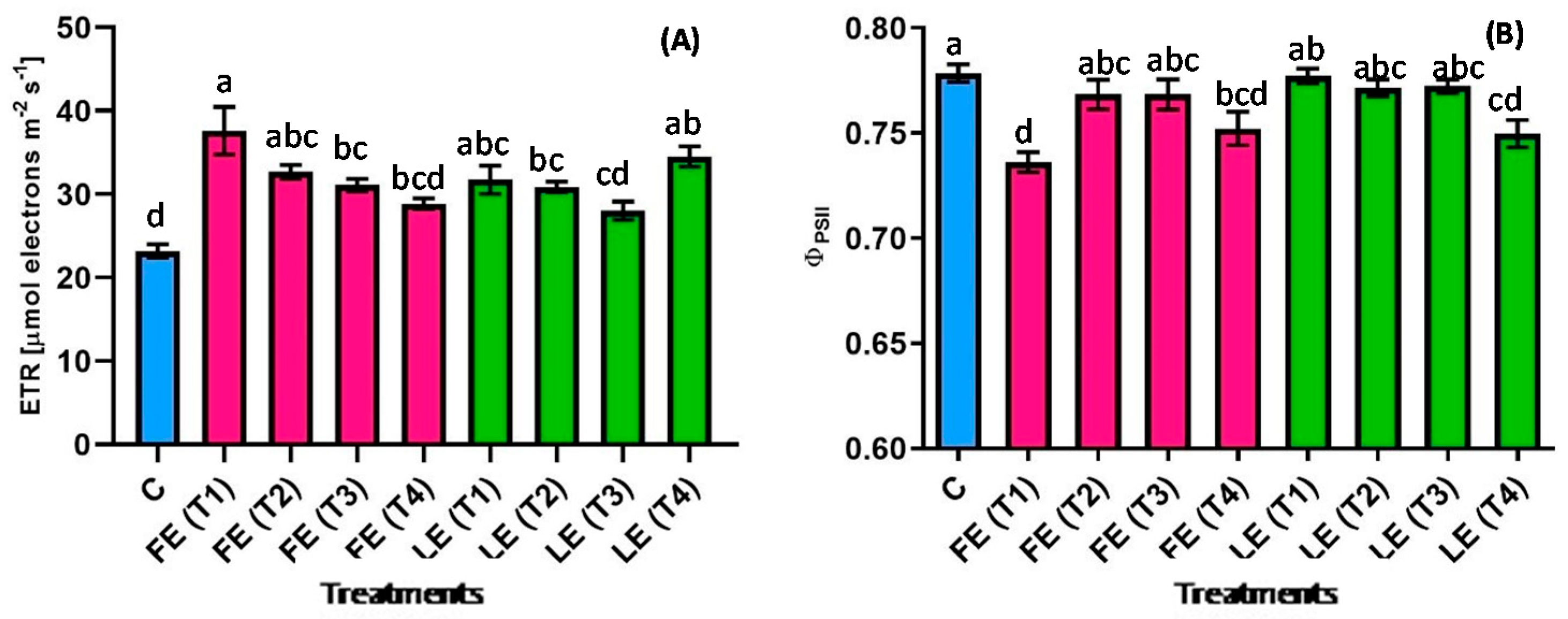
All borage extracts induced a general increase in the ETR (Figure 4A) of rocket leaves compared with the control. The lowest (23.2 µmol m−2 s−1) and the highest (37.6 µmol m−2 s−1) values were reached in control plants and in plants treated with FE (T1). The ETR value of control plants was significantly (p < 0.05) lower than FE (T1, T2, T3) and LE (T1, T2, T4) treatments by −38.3%, −29.1%, −25.4%, −27%, −24.9% and −33%, respectively. Moreover, plants treated with FE (T1) were significantly (p < 0.05) higher than plants treated with FE (T3), FE (T4), LE (T2) and LE (T4). FE-treated plants showed a decrease in ETR levels over the maceration time. A different effect was observed in the effective quantum yield of photosystem II in the light (ΦPSII) (Figure 4B). Most of the plants treated with borage extracts showed similar values as the control plants (0.78) while a significant (p < 0.05) decrease resulted after the treatments with FE (T1), FE (T2) and LE (T4).
3.6. Nitrate Concentration
Nitrate is the main nitrogen form used by the plants, but its concentration can have implication on human health. Table 3 lists the nitrate concentration measured in rocket leaves at harvest. In general, plants treated with borage extracts showed a slight but not significative decrease of nitrate levels compared with the control (6056.3 mg kg−1). Only FE (T4) treatment induced an opposite effect and the average value of nitrate content was 7550.1 mg kg−1 FW, exceeding the maximum level of nitrate concentration in rocket leaves imposed by the EU regulation N. 1258/2011. The statistical analysis revealed significant (p < 0.05) differences among this treatment and FE (T1, T3) and LE (T1, T2, T3) treatments. Short maceration time had a positive effect on nitrate assimilation in both flower and leaf extracts. LE induced the highest assimilation with the lowest concentration of nitrate, although a high variability was observed (Table 3).
3.7. Sucrose and Total Sugar Concentration
The levels of sucrose (Table 3) in rocket plants treated with borage extracts were generally lower than in the control (414.3 mg kg−1 FW). In particular, values recorded in response to FE (T1, T2, T4) and LE (T1, T2) treatments were significantly (p < 0.05) decreased by −26.6%, −21%, −23%, −19.8%, and −22.5%, respectively. The lowest sucrose concentration was observed in samples treated with FE (T1) and the value was around 303.49 mg kg−1 FW. Borage extracts application also induced a general decrease in the concentration of total sugars (Table 3). The highest value was around 2901.4 mg kg−1 FW and was found in the leaves of control plants. Samples treated with FE showed an increase of the concentration of total sugars in relation with maceration time. A similar trend was observed as well in LE-treated plants, after an initial decrease from (T1) to (T2). Nonetheless, no significant (p < 0.05) differences among treatments resulted from the statistical analysis.
3.8. Seed Germination
Table 4 shows the germination percentage (GP), an index that is commonly used to measure the effects of phytotoxic substances, of wild rocket seeds exposed to borage extracts. The tested extracts showed a general inhibitory effect on rocked seeds. The FE totally inhibited the germination, regardless of the time of maceration, whereas the rocket seeds treated with the FE obtained with 25 days of maceration (T5) had a GP of 33.3%. A different trend resulted from the application of extracts obtained by the leaves. In particular, no seeds germinated when soaked with LE (T3) and LE (T4), whereas a GP of 25% and 4.2% was recorded following treatment with LE (T1) and LE (T2), respectively.
4. Discussion
In the recent years, the phytochemicals and antioxidant activity of several plant species, as natural sources, have been studied to find application in different sectors. The interest in plant extracts with biostimulant properties is increasing as they have been shown to improve plant growth and quality [17,18,40,41]. The European Union is going to promote with “Green Deal” action all activities that lead to improve sustainability. In the framework of plant nutrition, the production of plant-based biostimulants could reduce the use of fertilizers and plant growth regulators. The application of plant extracts as a potential alternative or additional products to the currently used agrochemicals fits the need for a more sustainable agriculture [42,43,44]. Plants are a rich source of diverse bioactive compounds; however, the high variability in their composition, the difficulties in the choice of the best extraction method, and the stability of the final product are only some of the problems in the formulation of new plant-based products [8,45,46]. Hence, several experiments need to be carried out to assess their effects. Based on the results obtained from previous experiments on Borago officinalis L. [20,21,22], the aim of this study was to investigate the effect of the maceration time on the properties of the extracts and their effectiveness on rocket plants. The present study is the first report regarding maceration time of aqueous borage extracts. Extracts were obtained starting from plants grown under controlled conditions, instead of collecting the raw material from wild plants in open fields, since most of the bioactive compounds found in plants are secondary metabolites and they are closely related to the environmental conditions [47]. The purpose was to avoid any unexpected change in the external conditions that might affect the composition of the plants, trying to standardize the raw/starting material. In order to screen the effect of borage extracts, and their effectiveness in response to the different maceration times studied, several in vivo and biochemical analyses were performed on wild rocket leaves, at harvest, and a germination test was done to evaluate their effect also on seeds. In general, plants treated with different extracts showed a similar response to the control treated with water, regardless of the maceration time or the part of plant of the raw material. One of the characteristic responses of biostimulant products is an increase in chlorophyll content in treated plants. This effect/behavior has been observed in a wide range of crop species, environmental conditions, and in the response to different biostimulant formulations [17,48,49,50,51]. Chlorophyll is an important pigment and a quality parameter giving some indication of the physiological status of the plant. Indeed, it is closely related to photosynthetic potential and plant productivity; it can be used as index to have an idea of the nutrient status of the plant since most of the nitrogen is incorporated in chlorophyll molecules, and it is also responsible for the visual appearance. This aspect is particularly important in leafy vegetables as a quality trait and affecting consumer preferences [52]. In this study, the level of chlorophyll in plants treated with borage extracts did not change significantly. The content of chlorophyll a+b slightly decreased in plants treated with flower extracts as the time of extraction increases, while an opposite trend has been observed in plants treated with leaves extracts, similar to those observed by Bulgari et al. [20]. Likewise, carotenoids concentration exhibited a similar trend; this result was expected since these molecules serve as accessory pigments in light harvesting and have an important role in the protection of chlorophylls from photooxidation [53,54]. Moreover, they increase the nutritional value of the product and its health benefit for humans [55]. Nevertheless, the differences observed in chlorophylls were more emphasized in the analysis of total carotenoids. Besides their importance in photosynthetic tissues, carotenoids also serve as precursors for two phytohormones (abscisic acid and strigolactones) and several apocarotenoids and some of them also exhibit strong aroma properties. The accumulation of carotenoids and their stability in plant tissues are affected by several factors such as light, temperature, water stress, and pH [56,57] Even if we observed a decrease of carotenoids concentration after borage treatments, no damage in photosynthetic apparatus resulted from the analysis of chlorophyll fluorescence indexes. Indeed, the performance index (PI) and the quantum maximum efficiency of PSII (Fv/Fm) were similar to those of control plants and Fv/Fm values were always higher than 0.83, commonly considered as a threshold between stressed and non-stressed leaves. Thus, borage treatments might not have caused any damage in photosynthetic tissues of rocket leaves and a different cause was responsible of the reduction in carotenoids content. Moreover, all borage treatments stimulated the photosynthetic activity by enhancing the ETR of rocket plants. The increase in the value of ETR has been observed by several authors after the application of different biostimulant products, both in stressful and non-stressful conditions [58,59]. However, at the same time, rocket plants treated with some of the borage extracts showed a decrease in the effective quantum yield of photosystem II in the light (ΦPSII). One of the main effects of plant biostimulants is to trigger the accumulation of secondary metabolites leading to a better plant response to stresses [60] and a highest nutritional value of the final product [61]. This effect was observed in our experiment in response to all borage extracts application. In particular, data highlighted that the maceration time (3 days) induced a significant effect on rocket plants treated with the LE (T2) and FE (T2): the phenolic index and the anthocyanin levels were significantly higher when compared to control plants. Vegetables and in particular leafy vegetables are considered a high source of nitrate in human diet. Nitrate is found in nature as part of the nitrogen cycle, and play an important role in plant growth, nutrition, and development [62,63]. Its accumulation in plant tissues is a complex trait and is influenced by many internal and external factors such as the plant species/cultivar, the amount of fertilizer used, the light intensity, and temperature. Nitrate is relatively non-toxic by itself, but it can be dangerous for humans and increase the risk of several diseases [64]., The European Union imposed limits in nitrate concentration for the commercialization of several leafy products [65]. Thus, the determination of nitrate content has received an increasing attention and much research has been conducted to minimize the accumulation of nitrate in vegetables. Recently, it was proven that different biostimulant products reduce the nitrate levels in several plant species [48,49,66,67,68,69]. Among them, previous experiments showed that borage extracts also are able to decrease the nitrate accumulation in rocket by incrementing the activity of nitrate reductase and, in general, by stimulating N and C metabolisms [21], which are strictly linked. In the present study, only a slight but not significant decrease of nitrate levels was observed in plants treated with almost all borage extracts, except for FE (T4). This treatment induced an opposite effect and the nitrate content was 7550.1 mg kg−1 FW, exceeding also the maximum level of nitrate concentration imposed by EU regulation. At the same time, a decrease in sucrose levels was observed in plants treated with borage extracts, while the total sugars’ amount was not significantly affected. These results are in contrast to previous experiments on borage extracts where sucrose levels increased in rocket leaves after flower extract applications. Sucrose is the metabolic link between photosynthetic carbon production and carbon utilisation. It is the end product of photosynthesis and it functions as carbon skeleton for the production of several molecules. It is also considered as a signal molecule, involved in the regulation of different metabolic pathways. In present study, the lower amount of sucrose observed in rocket leaves might be attributed to a faster metabolization or transport and not to a decrease in its biosynthesis. Indeed, as confirmed by the levels of photosynthetic pigment or by the analyses of chlorophyll fluorescence indexes, borage extracts did not negatively affect the photosynthetic apparatus. The reduction of sugars after biostimulant applications is a common result. Lower sucrose and reducing sugars were found in endive and lettuce vegetables treated with ONE® [50]. The application of borage extracts also reduced the sucrose and the total sugars in lettuce [20]. Overall, based on the current results, even if different trends resulted from the application of borage extracts, neither the time of maceration nor the part of the plants used showed a clear effect on the efficacy of borage treatments. Moreover, the high variability observed, did not help us to understand the effective properties and the mechanisms of action of these extracts. The high variability and the unmarked effects of the treatments are probably related to the borage cultivation system. Indeed, it is known that environmental stress such as drought, salinity, soil strength, and nutrient status have a great influence on the accumulation of the active compounds in plants. Moreover, the longer time (25 days) of maceration chosen in the previous experiment might have led the extraction of different molecules with a biostimulant activity. Plant aqueous extracts are also studied for their potential allelopathic properties [70,71,72,73] deriving from substances therein present. In the last years, there has been a growing interest in allelopathy, because it may provide alternative tools to synthetic herbicides [74,75] and thus can be widely employed in sustainable and organic agriculture [76]. Promising results were obtained with several allelopathic crop types that actively influenced the germination and growth of surrounding plants [77,78]. Due to the multitude of properties attributed to borage, we decided to investigate this possible activity of extracts, under laboratory conditions, as a preliminary trial. The germination test showed that borage extracts exert an inhibitory effect on the seeds of rocket. In particular, the new FE totally blocked the germination, regardless of the time of maceration. A different trend resulted in response to LE. Indeed, the inhibition of seed germination was stronger after the application of LE obtained with longer maceration times. On the contrary, the FE and LE prepared from wild plants collected in open field and obtained after 25 days of macerations had a less inhibitory effect when compared with the new ones. Phenolic acids are often considered as putative allelochemicals and they are the most commonly investigated [79]. The observed inhibition effect could be ascribed to the high phenolics content of borage extracts, as suggested in the literature [28,80]. The results highlighted that FE is more effective than LE, confirming the higher amount of bioactive compounds in flowers compared to leaves.
5. Conclusions
In the recent years, one of the main purposes of agriculture has been to obtain a production taking care of the environment, increasing the sustainability of cropping systems. In light of this, an increasing number of studies has been focused on the exploitation of plant extracts as tools able to affect crops performance. Considering the complex origin of these types of products and the recent regulation related to plant extracts or biostimulants in general, in-depth studies are necessary to obtain a standardized product and to clarify its mode of action. Our trial highlighted a different effect when the extracts are applied on rocket plants or seeds. Borage extracts showed a biostimulant activity on the leafy vegetable, and the present study reveals that time of maceration could affect their efficacy. On the contrary, when applied on rocket seeds, borage extracts exerted an inhibitory activity. This result suggests that the concentrations for stimulating the germination should be further investigated. In addition, the choice of aqueous extracts represents an interesting easy and cheap solution due to their inexpensive and rapid preparation without negative implications for the environment.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/agronomy11112182/s1, Table S1: Chemical characterization of borage extracts. Table S2: Terms and formulae used in the analysis of the fast chlorophyll a fluorescence.
Author Contributions
Conceptualization A.F. and G.F.; methodology R.B., A.F. and G.F.; validation R.B., A.F. and G.F.; formal analysis G.F. and R.B. writing—original draft preparation R.B. and G.F.; writing—review and editing R.B., G.F. and A.F.; supervision A.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data reported in the manuscript can be provided upon request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schmidt, B.; Ribnicky, D.M.; Poulev, A.; Logendra, S.; Cefalu, W.T.; Raskin, I. A natural history of botanical therapeutics. Metabolism 2008, 57, S3–S9. [Google Scholar] [CrossRef] [Green Version]
- Ben Mrid, R.; Benmrid, B.; Hafsa, J.; Boukcim, H.; Sobeh, M.; Yasri, A. Secondary metabolites as biostimulant and bioprotectant agents: A review. Sci. Total Environ. 2021, 777, 146204. [Google Scholar] [CrossRef]
- Erb, M.; Kliebenstein, D.J. Plant Secondary Metabolites as Defenses, Regulators, and Primary Metabolites: The Blurred Functional Trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Franzoni, G.; Trivellini, A.; Bulgari, R.; Cocetta, G.; Ferrante, A. Bioactive Molecules as Regulatory Signals in Plant Responses to Abiotic Stresses. In Plant Signaling Molecules; Elsevier: Amsterdam, The Netherlands, 2019; pp. 169–182. [Google Scholar]
- Dias, L.S.; Dias, A.S. Secondary metabolites as sources of bioherbicides: Present situation and perspectives. Rev. Ciênc. Agrár. 2007, 30, 510–517. [Google Scholar]
- Soltys, D.; Krasuska, U.; Bogatek, R.; Gniazdowsk, A. Allelochemicals as Bioherbicides—Present and Perspectives. In Herbicides—Current Research and Case Studies in Use; Price, A.J., Kelton, J.A., Eds.; InTech: Rijeka, Croatia, 2013. [Google Scholar]
- Handa, S.S.; Khanuja, S.P.S.; Longo, G.; Rakesh, D.D. Extraction Technologies for Medicinal and Aromatic Plants; Earth, Environmental and Marine Sciences and Technologies: Trieste, Italy, 2008. [Google Scholar]
- Azmir, J.; Zaidul, I.S.M.; Rahman, M.M.; Sharif, K.M.; Mohamed, A.; Sahena, F.; Jahurul, M.H.A.; Ghafoor, K.; Norulaini, N.A.N.; Omar, A.K.M. Techniques for extraction of bioactive compounds from plant materials: A review. J. Food Eng. 2013, 117, 426–436. [Google Scholar] [CrossRef]
- Azwanida, N. A Review on the Extraction Methods Use in Medicinal Plants, Principle, Strength and Limitation. Med. Aromat. Plants 2015, 4, 3–8. [Google Scholar] [CrossRef]
- Raynie, D.E. Extraction. In Encyclopedia of Separation Science; Elsevier: Amsterdam, The Netherlands, 2000; pp. 118–128. [Google Scholar]
- Vongsak, B.; Sithisarn, P.; Mangmool, S.; Thongpraditchote, S.; Wongkrajang, Y.; Gritsanapan, W. Maximizing total phenolics, total flavonoids contents and antioxidant activity of Moringa oleifera leaf extract by the appropriate extraction method. Ind. Crop. Prod. 2013, 44, 566–571. [Google Scholar] [CrossRef]
- Sithisarn, P.; Supabphol, R.; Gritsanapan, W. Comparison of free radical scavenging activity of Siamese neem tree (Azadirachta indica A. Juss var. siamensis Valeton) leaf extracts prepared by different methods of extraction. Med. Princ. Pract. 2006, 15, 219–222. [Google Scholar] [CrossRef]
- Abdel-Rahman, S.S.A.; Abdel-Kader, A.A.S. Response of Fennel (Foeniculum vulgare, Mill) plants to foliar application of moringa leaf extract and benzyladenine (BA). S. Afr. J. Bot. 2020, 129, 113–122. [Google Scholar] [CrossRef]
- Abou-Sreea, A.I.; Matter, F.M. Using moringa leaf extract as biostimulant and giberrellic acid for enhancing fennel (Foeniculum vulgare var. azoricum Mill.) growth and oil yield. Acta Sci. Intellectus 2016, 2, 7–20. [Google Scholar]
- Abd El-Mageed, T.A.; Semida, W.M.; Rady, M.M. Moringa leaf extract as biostimulant improves water use efficiency, physio-biochemical attributes of squash plants under deficit irrigation. Agric. Water Manag. 2017, 193, 46–54. [Google Scholar] [CrossRef]
- Howladar, S.M. A novel Moringa oleifera leaf extract can mitigate the stress effects of salinity and cadmium in bean (Phaseolus vulgaris L.) plants. Ecotoxicol. Environ. Saf. 2014, 100, 69–75. [Google Scholar] [CrossRef]
- Abdalla, M.M. The potential of Moringa oleifera extract as a biostimulant in enhancing the growth, biochemical and hormonal contents in rocket (Eruca vesicaria subsp. sativa) plants. Int. J. Plant Physiol. Biochem. 2013, 5, 42–49. [Google Scholar] [CrossRef]
- Culver, M.; Fanuel, T.; Chiteka, A.Z. Effect of Moringa Extract on Growth and Yield of Tomato. Greener J. Agric. Sci. 2012, 2, 207–211. [Google Scholar]
- Rady, M.M.; Desoky, E.S.M.; Elrys, A.S.; Boghdady, M.S. Can licorice root extract be used as an effective natural biostimulant for salt-stressed common bean plants? S. Afr. J. Bot. 2019, 121, 294–305. [Google Scholar] [CrossRef]
- Bulgari, R.; Morgutti, S.; Cocetta, G.; Negrini, N.; Farris, S.; Calcante, A.; Spinardi, A.; Ferrari, E.; Mignani, I.; Oberti, R.; et al. Evaluation of Borage Extracts as Potential Biostimulant Using a Phenomic, Agronomic, Physiological, and Biochemical Approach. Front. Plant Sci. 2017, 8, 1–16. [Google Scholar] [CrossRef]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Ferrante, A. Borage extracts affect wild rocket quality and influence nitrate and carbon metabolism. Physiol. Mol. Biol. Plants 2020, 26, 649–660. [Google Scholar] [CrossRef]
- Franzoni, G.; Cocetta, G.; Trivellini, A.; Ferrante, A. Transcriptional Regulation in Rocket Leaves as Affected by Salinity. Plants 2019, 9, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramezani, M.; Amiri, M.S.; Zibaee, E.; Boghrati, Z.; Ayati, Z.; Sahebkar, A.; Emami, S.A. A Review on the Phytochemistry, Ethnobotanical Uses and Pharmacology of Borago Species. Curr. Pharm. Des. 2019, 26, 110–128. [Google Scholar] [CrossRef]
- Garcia-Herreros, C.; Garcia-Iñiguez-de-Ciriano, M.; Astiasarán, I.; Ansorena-Artieda, D. Antioxidant activity and phenolic content of water extracts of Borago officinalis L: Influence of plant part and cooking procedures. Ital. J. Food Sci. 2010, 22, 156–164. [Google Scholar]
- Salem, N.; Msaada, K.; Hammami, M.; Jday, A.; Salem, S.; Limam, F.; Marzouk, B. Variations in Tunisian borage essential oil profiles and their antioxidant activities during flowering. Nat. Prod. Res. 2014, 28, 1919–1922. [Google Scholar] [CrossRef]
- Salem, N.; Msaada, K.; Hammami, M.; Limam, F.; Vasapollo, G.; Marzouk, B. Variation in anthocyanin and essential oil composition and their antioxidant potentialities during flower development of Borage (Borago officinalis L.). Plant Biosyst. 2014, 148, 444–459. [Google Scholar] [CrossRef]
- Bandoniene, D.; Murkovic, M.; Venskutonis, P.R. Determination of rosmarinic acid in sage and borage leaves by high-performance liquid chromatography with different detection methods. J. Chromatogr. Sci. 2005, 43, 372–376. [Google Scholar] [CrossRef] [Green Version]
- Wettasinghe, M.; Shahidi, F.; Amarowicz, R.; Abou-Zaid, M.M. Phenolic acids in defatted seeds of borage (Borago officinalis L.). Food Chem. 2001, 75, 49–56. [Google Scholar] [CrossRef]
- Karimi, E.; Oskoueian, E.; Karimi, A.; Noura, R.; Ebrahimi, M. Borago officinalis L. flower: A comprehensive study on bioactive compounds and its health-promoting properties. J. Food Meas. Charact. 2018, 12, 826–838. [Google Scholar] [CrossRef]
- Herrmann, M.; Joppe, H.; Schmaus, G. Thesinine-4′-O-β-D-glucoside the first glycosylated plant pyrrolizidine alkaloid from Borago officinalis. Phytochemistry 2002, 60, 399–402. [Google Scholar] [CrossRef]
- Vacillotto, G.; Favretto, D.; Seraglia, R.; Pagiotti, R.; Traldi, P.; Mattoli, L. A rapid and highly specific method to evaluate the presence of pyrrolizidine alkaloids in Borago officinalis seed oil. J. Mass Spectrom. 2013, 48, 1078–1082. [Google Scholar] [CrossRef] [PubMed]
- Bell, L. Rocket Science: Phytochemical, Postharvest, Shelf-Life & Sensory Attributes of Rocket Species. Ph.D. Thesis, University of Reading, Reading, UK, 2016. [Google Scholar]
- Bell, L.; Lignou, S.; Wagstaff, C. High Glucosinolate Content in Rocket Leaves (Diplotaxis tenuifolia and Eruca sativa) after Multiple Harvests Is Associated with Increased Bitterness, Pungency, and Reduced Consumer Liking. Foods 2020, 9, 1799. [Google Scholar] [CrossRef]
- Franzoni, G.; Cocetta, G.; Ferrante, A. Effect of glutamic acid foliar applications on lettuce under water stress. Physiol. Mol. Biol. Plants 2021, 27, 1059–1072. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Plant Cell Membranes; Academic Press: Cambridge, MA, USA, 1987; pp. 350–382. [Google Scholar]
- Klein, A.O.; Hagen, C.W. Anthocyanin production in detached petals of impatiens balsamina L. Plant Physiol. 1961, 36, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Rorem, E.S.; Walker, H.G.; McCready, R.M. Biosynthesis of Sucrose and Sucrose-Phosphate by Sugar Beet Leaf Extracts. Plant Physiol. 1960, 35, 269–272. [Google Scholar] [CrossRef] [Green Version]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508. [Google Scholar] [CrossRef] [Green Version]
- Pardo-García, A.I.; Martínez-Gil, A.M.; Cadahía, E.; Pardo, F.; Alonso, G.L.; Salinas, M.R. Oak extract application to grapevines as a plant biostimulant to increase wine polyphenols. Food Res. Int. 2014, 55, 150–160. [Google Scholar] [CrossRef]
- Szparaga, A.; Czerwińska, E.; Piskier, T. The effect of treating the seeds of Brassica oleracea L. with aqueous extracts on the germination capacity and seed healthiness. J. Res. Appl. Agric. Eng. 2017, 62, 162–167. [Google Scholar]
- Toscano, S.; Romano, D.; Massa, D.; Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulant applications in low input horticultural cultivation systems. Italus Hortus 2019, 25, 27–36. [Google Scholar] [CrossRef]
- Xu, L.; Geelen, D. Developing Biostimulants from Agro-Food and Industrial By-Products. Front. Plant Sci. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chojnacka, K. Innovative bio-products for agriculture. Open Chem. 2015, 13, 932–937. [Google Scholar] [CrossRef]
- Lapornik, B.; Prošek, M.; Wondra, A.G. Comparison of extracts prepared from plant by-products using different solvents and extraction time. J. Food Eng. 2005, 71, 214–222. [Google Scholar] [CrossRef]
- Naboulsi, I.; Aboulmouhajir, A.; Kouisni, L.; Bekkaoui, F.; Yasri, A. Plants extracts and secondary metabolites, their extraction methods and use in agriculture for controlling crop stresses and improving productivity: A review. Acad. J. Med. Plants 2018, 6, 223–240. [Google Scholar] [CrossRef]
- Mazid, M.; Khan, T.; Mohammad, F. Role of Secondary Metabolites in Chemical Defence Mechanisms in Plants. Front. Life Sci. 2011, 3, 232–249. [Google Scholar] [CrossRef]
- Vernieri, P.; Borghesi, E.; Tognoni, F.; Serra, G.; Ferrante, A.; Piaggesi, A. Use of biostimulants for reducing nutrient solution concentration in floating system. Acta Hortic. 2006, 718, 477–484. [Google Scholar] [CrossRef]
- Vernieri, P.; Borghesi, E.; Ferrante, A. Application of Biostimulants in Floating System for Improving Rocket Quality. J. Food Agric. Environ. 2005, 3, 86–88. [Google Scholar]
- Bulgari, R.; Podetta, N.; Cocetta, G.; Piaggesi, A.; Ferrante, A. The effect of a complete fertilizer for leafy vegetables production in family and urban gardens. Bulg. J. Agric. Sci. 2014, 20, 1361–1367. [Google Scholar]
- Abbas, S.M.; Akladious, S.A. Application of carrot root extract induced salinity tolerance in cowpea (Vigna sinensis L.) seedlings. Pak. J. Bot. 2013, 45, 795–806. [Google Scholar]
- Ferrante, A.; Incrocci, L.; Maggini, R.; Serra, G.; Tognoni, F. Colour changes of fresh-cut leafy vegetables during storage. J. Food. Agric. Environ. 2004, 22, 40–44. [Google Scholar]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid metabolism in plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Zhang, J.; Nageswaran, D.; Li, L. Carotenoid metabolism and regulation in horticultural crops. Hortic. Res. 2015, 2, 15036. [Google Scholar] [CrossRef] [Green Version]
- Rao, A.V.; Rao, L.G. Carotenoids and human health. Pharmacol. Res. 2007, 55, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Othman, R.; Hassan, N.M.; Hatta, F.A.M. Influence of Environmental Stress toward Carotenogenesis Regulatory Mechanism through In Vitro Model System. In Carotenoids; InTech: Rijeka, Croatia, 2017; Volume i, p. 13. [Google Scholar]
- Morosinotto, T.; Caffarri, S.; Dall’Osto, L.; Bassi, R. Mechanistic aspects of the xanthophyll dynamics in higher plant thylakoids. Physiol. Plant. 2003, 119, 347–354. [Google Scholar] [CrossRef]
- Kocira, S.; Sujak, A.; Oniszczuk, T.; Szparaga, A.; Szymanek, M.; Karakuła-Juchnowicz, H.; Krawczuk, A.; Kupryaniuk, K. Improvement of the photosynthetic activity of Moldavian dragonhead (Dracocephalum moldavica L.) through foliar application of a nitrophenolate–based biostimulant. BIO Web Conf. 2018, 10, 01009. [Google Scholar] [CrossRef] [Green Version]
- Bulgari, R.; Trivellini, A.; Ferrante, A. Effects of Two Doses of Organic Extract-Based Biostimulant on Greenhouse Lettuce Grown Under Increasing NaCl Concentrations. Front. Plant Sci. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants Application in Horticultural Crops under Abiotic Stress Conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and crop responses: A review. Biol. Agric. Hortic. 2015, 31, 1–17. [Google Scholar] [CrossRef]
- Qiu, W.; Wang, Z.; Huang, C.; Chen, B.; Yang, R. Nitrate accumulation in leafy vegetables and its relationship with water. J. Soil Sci. Plant Nutr. 2014, 14, 761–768. [Google Scholar] [CrossRef]
- Bosnir, D.B.J.; Bevardi, M.; Boskovic, A.G.; Lasic, S.M.D.; Krivohlavek, A.; Racs, A.; Mojosovic-Cuic, A.; Trstenjak, N.U. Nitrate in Leafy Green Vegetables and Estimated Intake. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Cavaiuolo, M.; Ferrante, A. Nitrates and glucosinolates as strong determinants of the nutritional quality in rocket leafy salads. Nutrients 2014, 6, 1519–1538. [Google Scholar] [CrossRef] [Green Version]
- Dudaš, S.; Šola, I.; Sladonja, B.; Erhatić, R.; Ban, D.; Poljuha, D. The Effect of Biostimulant and Fertilizer on “Low Input” Lettuce Production. Acta Bot. Croat. 2016, 75, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Kunicki, E.; Grabowska, A.; Sękara, A.; Wojciechowska, R. The effect of cultivar type, time of cultivation, and biostimulant treatment on the yield of spinach (Spinacia oleracea L.). Folia Hortic. 2010, 22, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Tarantino, E.; Disciglio, G.; Frabboni, L.; Libutti, A.; Gatta, G.; Gagliaridi, A.; Tarantino, A. Effects of Biostimulant Application on Quali-Quantitative Characteristics of Cauliflower, Pepper and Fennel Crops under Organic and Conventional Fertilization. WASET Int. J. Agric. Biosyst. Eng. 2015, 9, 712–716. [Google Scholar]
- Amanda, A.; Ferrante, A.; Valagussa, M.; Piaggesi, A. Effect of biostimulants on quality of baby leaf lettuce grown under plastic tunnel. Acta Hortic. 2009, 807, 407–412. [Google Scholar] [CrossRef]
- Turk, M.A.; Tawaha, A.M. Allelopathic effect of black mustard (Brassica nigra L.) on germination and growth of wild oat (Avena fatua L.). Crop Prot. 2003, 22, 673–677. [Google Scholar] [CrossRef]
- Bogatek, R.; Gniazdowsk, A.; Zakrzewska, W.; Oracz, K.; Gawroński, S.W. Allelopathic effects of sunflower extracts on mustard seed germination and seedling growth. Biol. Plant. 2006, 50, 156–158. [Google Scholar] [CrossRef]
- Islam, A.; KatoNoguchi, H. Plant growth inhibitory activity of medicinal plant Hyptis suaveolens: Could allelopathy be a cause? Emirates J. Food Agric. 2013, 25, 692. [Google Scholar] [CrossRef] [Green Version]
- Baziar, M.R.; Farahvash, F.; Mirshekari, B.; Rashidi, V. Allelopathic effect of ryegrass (Lolium persicum) and wild mustard (Sinapis arvensis) on barley. Pak. J. Bot. 2014, 46, 2069–2075. [Google Scholar]
- Bhowmik, P.C. Inderjit Challenges and opportunities in implementing allelopathy for natural weed management. Crop Prot. 2003, 22, 661–671. [Google Scholar] [CrossRef]
- Jabran, K.; Mahajan, G.; Sardana, V.; Chauhan, B.S. Allelopathy for weed control in agricultural systems. Crop Prot. 2015, 72, 57–65. [Google Scholar] [CrossRef]
- Findura, P.; Hara, P.; Szparaga, A.; Kocira, S.; Czerwińska, E.; Bartoš, P.; Nowak, J.; Treder, K. Evaluation of the Effects of Allelopathic Aqueous Plant Extracts, as Potential Preparations for Seed Dressing, on the Modulation of Cauliflower Seed Germination. Agriculture 2020, 10, 122. [Google Scholar] [CrossRef] [Green Version]
- Azania, A.A.P.M.; Azania, C.A.M.; Alves, P.L.C.A.; Palaniraj, R.; Kadian, H.S.; Sati, S.C.; Rawat, L.S.; Dahiya, D.S.; Narwal, S.S. Allelopathic plants. 7. Sunflower (Helianthus annuus L.). Allelopath. J. 2003, 11, 1–20. [Google Scholar]
- Weston, L.A.; Duke, S.O. Weed and Crop Allelopathy. CRC Crit. Rev. Plant Sci. 2003, 22, 367–389. [Google Scholar] [CrossRef]
- Chon, S.-U.; Jang, H.-G.; Kim, D.-K.; Kim, Y.-M.; Boo, H.-O.; Kim, Y.-J. Allelopathic potential in lettuce (Lactuca sativa L.) plants. Sci. Hortic. 2005, 106, 309–317. [Google Scholar] [CrossRef]
- Aliakbarlu, J.; Tajik, H. Antioxidant and antibacterial activities of various extracts of borago officinalis flowers. J. Food Process. Preserv. 2012, 36, 539–544. [Google Scholar] [CrossRef]
Figure 1.
Chlorophyll content determined in vivo (A) and chlorophyll a+b concentration (B) in rocket leaves treated with water (control = C) and with borage extracts (flowers extract = FE or leaves extract = LE) obtained with different maceration times: 1 day (T1), 3 days (T2), 7 days (T3), and 14 days (T4). Values are means ± SE (n = 5) (A); (n = 3) (B). Data were subjected to one-way ANOVA. “n.s.” means not significant.
Figure 1.
Chlorophyll content determined in vivo (A) and chlorophyll a+b concentration (B) in rocket leaves treated with water (control = C) and with borage extracts (flowers extract = FE or leaves extract = LE) obtained with different maceration times: 1 day (T1), 3 days (T2), 7 days (T3), and 14 days (T4). Values are means ± SE (n = 5) (A); (n = 3) (B). Data were subjected to one-way ANOVA. “n.s.” means not significant.
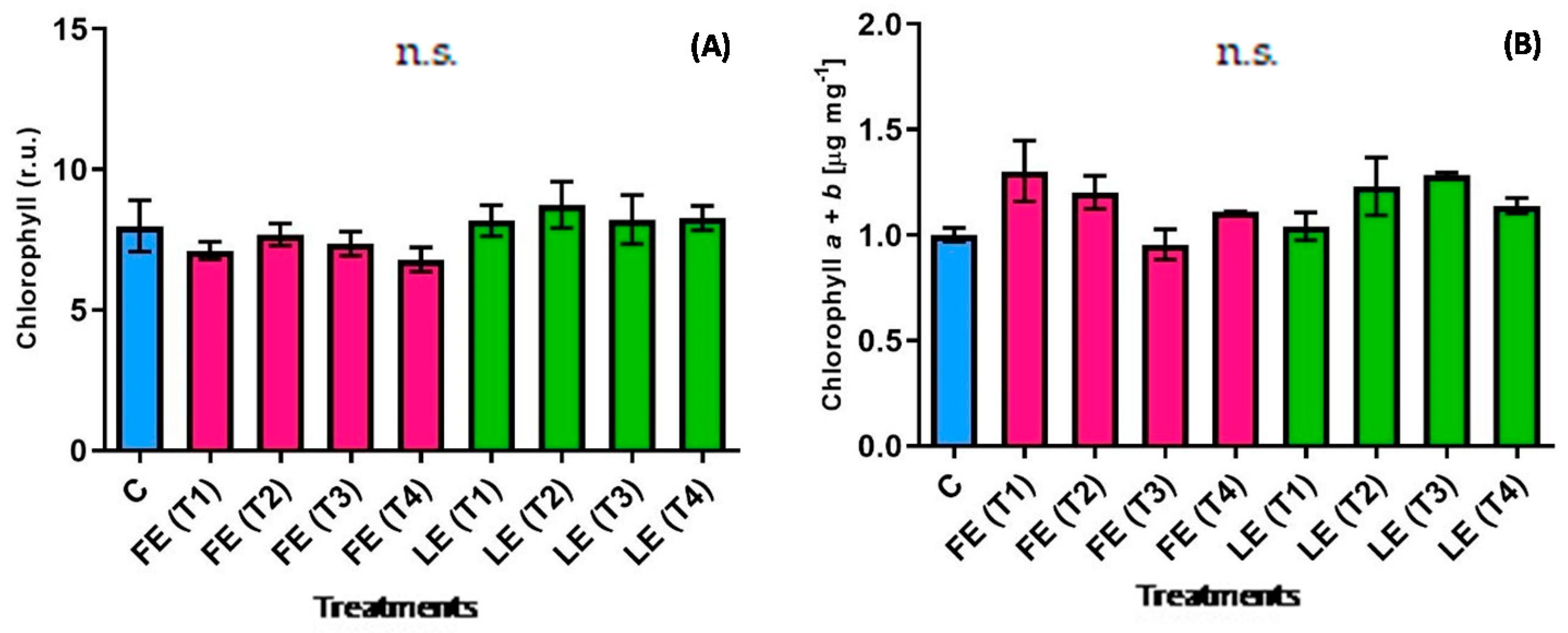
Figure 2.
Carotenoids’ concentration in rocket leaves treated with water (control = C) and with borage extracts (flowers extract = FE or leaves extract = LE) obtained with different maceration times: 1 day (T1), 3 days (T2), 7 days (T3), and 14 days (T4). Values are means ± SE (n = 3). Data were subjected to one-way ANOVA. “n.s.” means not significant.
Figure 2.
Carotenoids’ concentration in rocket leaves treated with water (control = C) and with borage extracts (flowers extract = FE or leaves extract = LE) obtained with different maceration times: 1 day (T1), 3 days (T2), 7 days (T3), and 14 days (T4). Values are means ± SE (n = 3). Data were subjected to one-way ANOVA. “n.s.” means not significant.
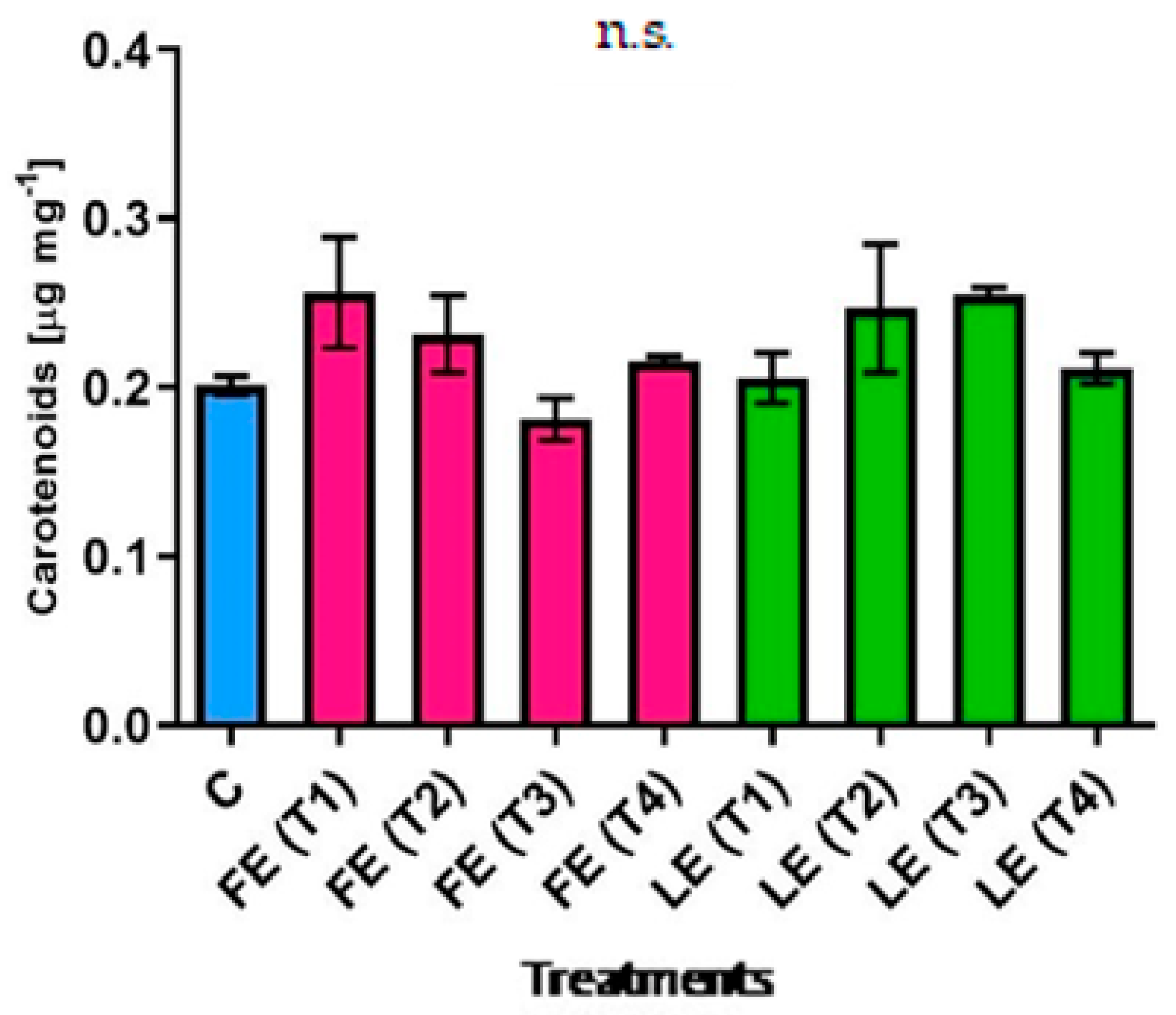
Figure 3.
Chlorophyll a fluorescence parameters of rocket leaves treated with water (control) and with borage extracts: (A) flowers extract = FE or (B) leaves extract = LE, obtained with different maceration times: 1 day (T1), 3 days (T2), 7 days (T3), and 14 days (T4). Values are means ± SE (n = 4). Data were subjected to one-way ANOVA. Data plotted are fluorescence parameters normalized by formulae: (Ft−Fcw)/Fcw, where “Ft” and “Fcw” represent the parameter values of the treated plants and control plants treated with water, respectively. Values of “Fcw” plants were normalized to 0 (control plants treated with water, blue circle = 0).
Figure 3.
Chlorophyll a fluorescence parameters of rocket leaves treated with water (control) and with borage extracts: (A) flowers extract = FE or (B) leaves extract = LE, obtained with different maceration times: 1 day (T1), 3 days (T2), 7 days (T3), and 14 days (T4). Values are means ± SE (n = 4). Data were subjected to one-way ANOVA. Data plotted are fluorescence parameters normalized by formulae: (Ft−Fcw)/Fcw, where “Ft” and “Fcw” represent the parameter values of the treated plants and control plants treated with water, respectively. Values of “Fcw” plants were normalized to 0 (control plants treated with water, blue circle = 0).

Figure 4.
The electron transport rate (ETR) (A) and the effective quantum yield of photosystem II in the light (ΦPSII) (B) in rocket leaves treated with water (control = C) and with borage extracts (flowers extract = FE or leaves extract = LE) obtained with different maceration times: 1 day (T1), 3 days (T2), 7 days (T3), and 14 days (T4). Values are means ± SE (n = 5). Data were subjected to one-way ANOVA. Different letters, where present, represent significant differences among treatments.
Figure 4.
The electron transport rate (ETR) (A) and the effective quantum yield of photosystem II in the light (ΦPSII) (B) in rocket leaves treated with water (control = C) and with borage extracts (flowers extract = FE or leaves extract = LE) obtained with different maceration times: 1 day (T1), 3 days (T2), 7 days (T3), and 14 days (T4). Values are means ± SE (n = 5). Data were subjected to one-way ANOVA. Different letters, where present, represent significant differences among treatments.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Yield and dry matter (%) of rocket plants treated with water (Control) and with borage extracts (flowers extract = FE or leaves extract = LE) obtained with different maceration times: 1 day (T1), 3 days (T2), 7 days (T3), and 14 days (T4). Yield was measured for one tank of each condition at the end of the experiment by cutting plants at collar.
Table 1.
Yield and dry matter (%) of rocket plants treated with water (Control) and with borage extracts (flowers extract = FE or leaves extract = LE) obtained with different maceration times: 1 day (T1), 3 days (T2), 7 days (T3), and 14 days (T4). Yield was measured for one tank of each condition at the end of the experiment by cutting plants at collar.
| Treatment | Yield (g m−2) | Dry Matter (%) |
|---|---|---|
| Control | 2771 | 15 |
| FE (T1) | 2343 | 16 |
| FE (T2) | 2760 | 14 |
| FE (T3) | 2386 | 16 |
| FE (T4) | 1947 | 18 |
| LE (T1) | 3027 | 14 |
| LE (T2) | 2559 | 15 |
| LE (T3) | 2662 | 15 |
| LE (T4) | 2749 | 15 |
Table 2.
Phenol index and anthocyanin content in rocket leaves treated with water (control = C) and with borage extracts (flowers extract = FE or leaves extract = LE) obtained with different maceration times: 1 day (T1), 3 days (T2), 7 days (T3), and 14 days (T4). Values are means ± SE (n = 3). Data were subjected to one-way ANOVA. Different letters, where present, represent significant differences among treatments.
Table 2.
Phenol index and anthocyanin content in rocket leaves treated with water (control = C) and with borage extracts (flowers extract = FE or leaves extract = LE) obtained with different maceration times: 1 day (T1), 3 days (T2), 7 days (T3), and 14 days (T4). Values are means ± SE (n = 3). Data were subjected to one-way ANOVA. Different letters, where present, represent significant differences among treatments.
| Treatment | Phenol Index [Abs320 nm g−1] | Anthocyanin [Cyanidin eq. mg/100 g] |
|---|---|---|
| Control | 19.7 ± 0.66 b | 16.6 ± 0.35 b |
| FE (T1) | 24 ± 0.83 ab | 20.3 ± 0.77 ab |
| FE (T2) | 26.9 ± 0.17 a | 21.7 ± 0.25 a |
| FE (T3) | 20.2 ± 2.24 b | 18.2 ± 1.18 b |
| FE (T4) | 22.9 ± 1.20 ab | 19.7 ± 0.67 ab |
| LE (T1) | 22.6 ± 1.57 ab | 20 ± 1.25 ab |
| LE (T2) | 27.3 ± 1.88 a | 23.6 ± 1.64 a |
| LE (T3) | 21.6 ± 0.07 ab | 18.7 ± 0.16 b |
| LE (T4) | 22.5 ± 0.86 ab | 19.5 ± 0.35 ab |
Table 3.
Nitrate and sugar concentrations of rocket leaves treated with water (control = C) and with borage extracts (flowers extract = FE or leaves extract = LE) obtained with different maceration times: 1 day (T1), 3 days (T2), 7 days (T3), and 14 days (T4). Values are means ± SE (n = 3). Data were subjected to one-way ANOVA. Different letters, where present, represent significant differences among treatments.
Table 3.
Nitrate and sugar concentrations of rocket leaves treated with water (control = C) and with borage extracts (flowers extract = FE or leaves extract = LE) obtained with different maceration times: 1 day (T1), 3 days (T2), 7 days (T3), and 14 days (T4). Values are means ± SE (n = 3). Data were subjected to one-way ANOVA. Different letters, where present, represent significant differences among treatments.
| Treatment | Nitrate [mg kg−1 FW] | Sucrose [mg kg−1 FW] | Total Sugars [mg kg−1 FW] |
|---|---|---|---|
| Control | 6056.3 ± 673.67 ab | 414.3 ± 35.43 a | 2901.4 ± 199.14 |
| FE (T1) | 4509.9 ± 246.38 b | 303.5 ± 9.60 bcd | 1227.9 ± 345.42 |
| FE (T2) | 4842.8 ± 407.35 ab | 326.5 ± 14.38 bcd | 1787.4 ± 460.18 |
| FE (T3) | 4577.9 ± 207.07 b | 354.3 ± 13.73 ad | 1891.6 ± 485.37 |
| FE (T4) | 7550.8 ± 728.49 a | 318.8 ± 1.26 bcd | 2641.1 ± 288.75 |
| LE (T1) | 2764.0 ± 1335.32 b | 332.5 ± 4.68 bcd | 2151.3 ± 255.18 |
| LE (T2) | 4316.2 ± 159.04 b | 321.3 ± 3.15 bcd | 1372.2 ± 261.03 |
| LE (T3) | 4151.0 ± 976.44 b | 364.5 ± 11.22 ab | 1642.1 ± 114.20 |
| LE (T4) | 5236.6 ± 800.75 ab | 358.7 ± 5.31 ac | 2749.5 ± 703.27 |
Table 4.
Germination percentage after 10 days of rocket seeds treated with water (control = C) and with borage extracts (flowers extract = FE or leaves extract = LE) obtained with different maceration times: 1 day (T1), 3 days (T2), 7 days (T3), and 14 days (T4) and compared with FE and LE obtained after 25 days of maceration (T5).
Table 4.
Germination percentage after 10 days of rocket seeds treated with water (control = C) and with borage extracts (flowers extract = FE or leaves extract = LE) obtained with different maceration times: 1 day (T1), 3 days (T2), 7 days (T3), and 14 days (T4) and compared with FE and LE obtained after 25 days of maceration (T5).
| Treatment | Germination Percentage (%) |
|---|---|
| Control | 58.3 |
| FE (T1) | 0.0 |
| FE (T2) | 0.0 |
| FE (T3) | 0.0 |
| FE (T4) | 0.0 |
| LE (T1) | 25.0 |
| LE (T2) | 4.2 |
| LE (T3) | 0.0 |
| LE (T4) | 0.0 |
| FE (T5) | 33.3 |
| LE (T5) | 8.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Franzoni, G.; Bulgari, R.; Ferrante, A. Maceration Time Affects the Efficacy of Borage Extracts as Potential Biostimulant on Rocket Salad. Agronomy 2021, 11, 2182. https://doi.org/10.3390/agronomy11112182
AMA Style
Franzoni G, Bulgari R, Ferrante A. Maceration Time Affects the Efficacy of Borage Extracts as Potential Biostimulant on Rocket Salad. Agronomy. 2021; 11(11):2182. https://doi.org/10.3390/agronomy11112182
Chicago/Turabian StyleFranzoni, Giulia, Roberta Bulgari, and Antonio Ferrante. 2021. "Maceration Time Affects the Efficacy of Borage Extracts as Potential Biostimulant on Rocket Salad" Agronomy 11, no. 11: 2182. https://doi.org/10.3390/agronomy11112182
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.