
Effectiveness of Multifunctional Margins in Insect Biodiversity Enhancement and RTE Species Conservation in Intensive Agricultural Landscapes

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Areas of Study
2.2. MFM Mixture Plant Selection
2.3. Experimental Design and Sampling
2.4. Statistical Analysis
2.4.1. Model for RTE Species
2.4.2. Models for Abundance of Species and Individuals
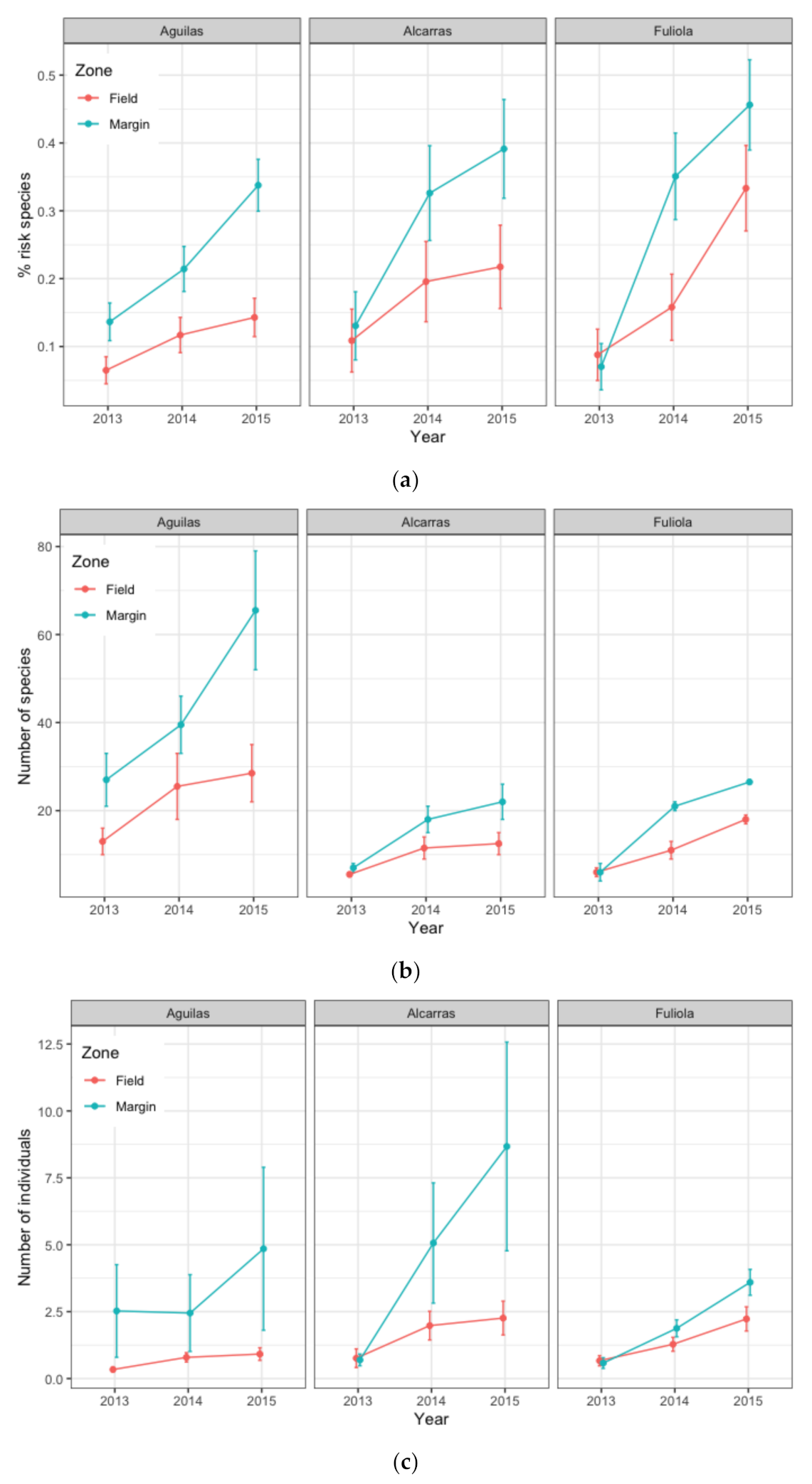
3. Results
3.1. Diversity of Insects
3.2. GLM Modelling
3.2.1. Model for RTE Species
3.2.2. Models for Abundance of Species and Individuals
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Farm | Year | Zone | % RTE Species | Number of Species | Number of Individuals | |||
|---|---|---|---|---|---|---|---|---|
| Mean | Std. Deviation | Mean | Std. Deviation | Mean | Std. Deviation | |||
| Aguilas | 2013 | Field | 0.06 | 0.25 | 13 | 4.2 | 0.3 | 1.2 |
| Margin | 0.14 | 0.34 | 27 | 8.5 | 2.5 | 21.5 | ||
| 2014 | Field | 0.12 | 0.32 | 26 | 10.6 | 0.8 | 2.2 | |
| Margin | 0.21 | 0.41 | 40 | 9.2 | 2.4 | 17.8 | ||
| 2015 | Field | 0.14 | 0.35 | 29 | 9.2 | 0.9 | 2.9 | |
| Margin | 0.34 | 0.47 | 66 | 19.1 | 4.9 | 37.8 | ||
| Alcarras | 2013 | Field | 0.11 | 0.31 | 6 | 0.7 | 0.8 | 2.4 |
| Margin | 0.13 | 0.34 | 7 | 1.4 | 0.7 | 1.5 | ||
| 2014 | Field | 0.20 | 0.40 | 12 | 3.5 | 2.0 | 3.6 | |
| Margin | 0.33 | 0.47 | 18 | 4.2 | 5.1 | 15.2 | ||
| 2015 | Field | 0.22 | 0.42 | 13 | 3.5 | 2.3 | 4.3 | |
| Margin | 0.39 | 0.49 | 22 | 5.7 | 8.7 | 26.5 | ||
| Fuliola | 2013 | Field | 0.09 | 0.29 | 6 | 1.4 | 0.7 | 1.4 |
| Margin | 0.07 | 0.26 | 6 | 2.8 | 0.6 | 1.5 | ||
| 2014 | Field | 0.16 | 0.37 | 11 | 2.8 | 1.3 | 2.0 | |
| Margin | 0.35 | 0.48 | 21 | 1.4 | 1.9 | 2.4 | ||
| 2015 | Field | 0.33 | 0.48 | 18 | 1.4 | 2.2 | 3.4 | |
| Margin | 0.46 | 0.50 | 27 | 0.7 | 3.6 | 3.6 | ||
| Model for the Number of Identified Species | ||||
|---|---|---|---|---|
| Test | Poisson | Negative binomial | ||
| Likelihood ratio (LR) | 322.6 | *** | 94.6 | *** |
| Deviance (D) | 18.1 | 18.1 | ||
| AIC | 198.2 | 200.2 | ||
| BIC | 209.3 | 212.9 | ||
| Model for the Number of Identified Individuals | ||||
| Test | Poisson | Negative binomial | ||
| Likelihood ratio (LR) | 1824.2 | *** | 203.8 | *** |
| Deviance (D) | 13,247.7 | 1316.6 | *** | |
| AIC | 15,205.9 | 5015.2 | ||
| BIC | 15,243.3 | 5057.9 | ||
References
- Gaston, K.J. The magnitude of global insect species richness. Conserv. Biol. 1991, 5, 283–296. [Google Scholar] [CrossRef]
- Mayr, E. A local flora and the biological species concept. Am. J. Botany. 1992, 79, 222–238. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Ritchie, J.M. Biodiversity and Biosystematic Priorities: Microorganisms and Invertebrates; CAB International: Wallingford, England, 1993. [Google Scholar]
- Wilson, P.J. Botanical diversity in arable field margins. In Field margins: Integrating Agriculture and Conservation; Boatman, N.D., Ed.; BCPC Monograph No. 58; British Crop Protection Council: Farnham, UK, 1994; pp. 53–58. [Google Scholar]
- Otto, S.; Vasileiadis, V.P.; Masin, R.; Zanin, G. Evaluating weed diversity with indices of varying complexity in north-eastern Italy. Weed Res. 2012, 52, 373–382. [Google Scholar] [CrossRef]
- Conrad, K.F.; Woiwod, I.P.; Parsons, M.; Fox, R.; Warren, M.S. Long-term population trends in widespread British moths. J. Insect Conserv. 2004, 8, 119–136. [Google Scholar] [CrossRef]
- Wenzel, M.; Schmitt, T.; Weitzel, M.; Seitz, A. The severe decline of butterflies on western German calcareous grasslands during the last 30 years: A conservation problem. Biol. Conserv. 2006, 128, 542–552. [Google Scholar] [CrossRef]
- Fox, R. The decline of moths in Great Britain: A review of possible causes. Insect Conserv. Divers. 2013, 6, 5–19. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Foppen, R.P.; van Turnhout, C.A.; de Kroon, H.; Jongejans, E. Declines in insectivorous birds are associated with high neonicotinoid concentrations. Nature 2014, 511, 641–643. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [Green Version]
- Habel, J.C.; Segerer, A.; Ulrich, W.; Torchyk, O.; Weisser, W.W.; Schmitt, T. Butterfly community shifts over two centuries. Conserv. Biol. 2016, 30, 754–762. [Google Scholar] [CrossRef]
- Samways, M.J.; Barton, P.S.; Birkhofer, K.; Chichorro, F.; Deacon, C.; Fartmann, T.; Fukushima, C.S.; Gaigner, R.; Habel., J.C.; Hallmann, C.; et al. Solutions for humanity on how to conserve insects. Biol. Conserv. 2020, 242, 108427. [Google Scholar] [CrossRef]
- Fanfarillo, E.; Kasperski, A.; Giuliani, A.; Abbate, G. Shifts of arable plant communities after agricultural intensification: A floristic and ecological diachronic analysis in maize fields of Latium (central Italy). Bot. Lett. 2019, 166, 356–365. [Google Scholar] [CrossRef]
- Richner, N.; Holderegger, R.; Linder, H.P.; Walter, T. Reviewing change in the arable flora of Europe: A meta-analysis. Weed Res. 2015, 55, 1–13. [Google Scholar] [CrossRef]
- McKechnie, I.M.; Thomsen, C.J.M.; Sargent, R.D. Forested field edges support a greater diversity of wild pollinators in lowbush blueberry (Vaccinium angustifolium). Agr. Ecosyst. Environ. 2017, 237, 154–161. [Google Scholar] [CrossRef]
- Williams, P.H.; Osborne, J.L. Bumblebee vulnerability and conservation world-wide. Apidologie 2009, 40, 367–387. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.J.F.; Paxton, R.J. The conservation of bees: A global perspective. Apidologie 2009, 40, 410–416. [Google Scholar] [CrossRef] [Green Version]
- Ollerton, J. Pollinator diversity: Distribution, ecological function, and conservation. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 353–376. [Google Scholar] [CrossRef] [Green Version]
- Millard, J.; Outhwaite, C.L.; Kinnersley, R.; Freeman, R.; Gregory, R.D.; Adedoja, O.; Gavini, S.; Kioko, E.; Kuhlmann, M.; Ollerton, J.; et al. Global effects of land-use intensity on local pollinator biodiversity. Nat. Commun. 2021, 12, 2902. [Google Scholar] [CrossRef]
- Saunders, M.E.; Janes, J.K.; O’hanlon, J.C. Moving on from the insect apocalypse narrative: Engaging with evidence-based insect conservation. BioScience 2020, 70, 80–89. [Google Scholar] [CrossRef] [Green Version]
- De Palma, A. Predicting bee community responses to land-use changes: Effects of geographic and taxonomic biases. Sci. Rep. 2016, 6, 31153. [Google Scholar] [CrossRef] [Green Version]
- Richards, A.J. Does low biodiversity resulting from modern agricultural practice affect crop pollination and yield? Ann. Bot. 2001, 88, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemuller, R.; Edwards, M.; Peeters, T.; Schaers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D. Parallel Declines in Pollinators and Insect-Pollinated Plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef]
- Westphal, C.; Bommarco, R.; Carré, G.; Lamborn, E.; Morison, N.; Petanidou, T.; Potts, S.G.; Roberts, S.P.M.; Szentgyörgyi, H.; Tscheulin, T.; et al. Measuring bee diversity in di erent European habitats and biogeographical regions. Ecol. Monogr. 2008, 78, 653–671. [Google Scholar] [CrossRef] [Green Version]
- Ricketts, T.H.; Regetz, J.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Bogdanski, A.; Gemmill-Herren, B.; Greenleaf, S.S.; Klein, A.M.; Mayfield, M.M.; et al. Landscape effects on crop pollination services: Are there general patterns? Ecol. Lett. 2008, 11, 499–515. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Montero-Castaño, A.; Vilà, M. Impact of landscape alteration and invasions on pollinators: A meta-analysis. J. Ecol. 2012, 100, 884–893. [Google Scholar] [CrossRef]
- IPBES (Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services). The Assessment Report on Pollinators; Pollination and Food Production: Bonn, Germany, 2017. [Google Scholar]
- Schumacher, W. Gefährdete Ackerwildkräuter können auf ungespritzten Feldrändern erhalten werden. Mitt. LÖLF 1984, 9, 14–20. [Google Scholar]
- Jörg, E. Field Margin-Strip Programmes; Landeranstalt fur Pflanzenbau und Pflanzenschutz: Mainz, Germany, 1994. [Google Scholar]
- De Snoo, G.R. Unsprayed field margins: Effects on environment, biodiversity and agricultural practice. Landsc. Urban Plan. 1999, 46, 151–160. [Google Scholar] [CrossRef]
- Marshall, E.J.P.; Moonen, A.C. Field margins in northern Europe: Their functions and interactions with agriculture. Agr. Ecosyst. Environ. 2002, 89, 5–21. [Google Scholar] [CrossRef]
- Jacot, K.; Eggenschwiler, L.; Junge, X.; Luka, H.; Bosshard, A. Improved field margins for a higher biodiversity in agricultural landscapes. Asp. Appl. Biol. 2007, 87, 277–283. [Google Scholar]
- Smith, J.; Potts, S.G.; Woodcock, B.A.; Eggleton, P. Can arable field margins be managed to enhance their biodiversity, conservation and functional value for soil macrofauna? J. Appl. Ecol. 2008, 45, 269–278. [Google Scholar] [CrossRef]
- Haddaway, N.R.; Brown, C.; Eggers, S.; Josefsson, J.; Kronvang, B.; Randall, N.; Uusi-Kämppä, J. The multifunctional roles of vegetated strips around and within agricultural fields. A systematic map protocol. Environ. Evid. 2016, 5, 18. [Google Scholar] [CrossRef] [Green Version]
- Nowakowski, M.; Pywell, R. Habitat Creation and Management for Pollinators; Centre for Ecology & Hydrology: Wallingford, UK, 2016. [Google Scholar]
- Karamaouna, F.; Jacques, J.A.; Kati, V. Practices to conserve pollinators and natural enemies. Insects 2021, 12, 31. [Google Scholar] [CrossRef] [PubMed]
- Marshall, E.J.P.; Nowakowski, M. Successional Changes in the Flora of a Sown Field Margin Strip Managed by Cutting and Herbicide Application; Brighton Crop Protection Conference—Weeds; British Crop Protection Council: Farnham, Surrey, 1995; pp. 973–978. [Google Scholar]
- Holland, J.; Fahrig, L. Effect of woody borders on insect density and diversity in crop fields: A landscape-scale analysis. Agr. Ecosyst. Environ. 2000, 78, 115–122. [Google Scholar] [CrossRef]
- Meek, B.; Loxton, D.; Sparks, T.; Pywell, R.; Pickett, H.; Nowakowski, M. The effect of arable field margin composition on invertebrate biodiversity. Biol. Conserv. 2002, 106, 259–271. [Google Scholar] [CrossRef]
- Roy, D.B.; Bohan, D.A.; Haughton, A.J.; Hill, M.O.; Osborne, J.L.; Clark, S.J.; Perry, J.N.; Rothery, P.; Scott, R.J.; Brooks, D.R.; et al. Invertebrates and vegetation of field margins adjacent to crops subject to contrasting herbicide regimes in the Farm Scale Evaluations of genetically modified herbicide-tolerant crops. Philos. Trans. Royal. Soc. B. 2003, 358, 1879–1898. [Google Scholar]
- New, T.R. Invertebrate Conservation in Agricultural Ecosystems; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Morandin, L.A.; Kremen, C. Hedgerow restoration promotes pollinator populations and exports native bees to adjacent fields. Ecol. Appl. 2013, 23, 829–839. [Google Scholar] [CrossRef] [Green Version]
- Tschumi, M.; Albrecht, M.; Entling, M.H.; Jacot, K. High effectiveness of tailored flower strips in reducing pests and crop plant damage. Proc. R. Soc. B 2015, 282, 20151369. [Google Scholar] [CrossRef] [Green Version]
- Holland, J.M.; Bianchi, F.J.J.A.; Entling, M.H.; Moonen, A.C.; Smith, B.M.; Jeanneret, P. Structure, function and management of semi-natural habitats for conservation biological control: A review of European studies. Pest Manag. Sci. 2016, 72, 1638–1651. [Google Scholar] [CrossRef]
- Castle, D.; Grass, I.; Westphal, C. Fruit quantity of strawberries benefit from enhanced pollinator abundance at hedgerows in agricultural landscapes. Agr. Ecosyst. Environ. 2019, 275, 14–22. [Google Scholar] [CrossRef]
- Kremen, C.; Albrech, M.; Ponisio, L.C. Restoring pollinator communities and pollination services in hedgerows in intensively managed agricultural landscapes. In The Ecology of Hedgerows and Field Margins; Dover, J.W., Ed.; Routledge: London, UK, 2019. [Google Scholar]
- Holden, J.; Grayson, R.P.; Berdeni, D.; Bird, S.; Chapman, P.J.; Edmondson, J.L.; Firbank, L.G.; Helgason, T.; Hodosn, M.E.; Hunt, S.F.P.; et al. The role of hedgerows in soil functioning within agricultural landscapes. Agr. Ecosyst. Environ. 2019, 273, 1–12. [Google Scholar] [CrossRef]
- Sánchez, J.A.; Carrasco, A.; La Spina, M.; Pérez-Marcos, M.; Ortíz-Sanchez, F.J. How bees respond differently to field margins of shrubby and herbaceous plants in intensive agricultural crops of the Mediterranean area. Insects 2020, 11, 26. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, M.; Kleijin, D.; Williams, N.M.; Tschumi, M.; Blaauw, B.R.; Bommarco, R.; Campbell, A.J.; Dainese, M.; Drummond, F.A.; Entling, M.H.; et al. Global synthesis of the effectiveness of flowers strips and hedgerows on pest control, pollination services and crop yield: A quantitative synthesis. Ecol. Lett. 2020, 23, 1488–1498. [Google Scholar] [CrossRef]
- Iwasaki, J.M.; Hogendoorn, K. How protection of honey bees can help and hinder bee conservation. Curr. Opin. Insect Sci. 2021, 46, 112–118. [Google Scholar] [CrossRef]
- Pyle, R.; Bentzien, M.; Opler, P. Insect conservation. Annu. Rev. Entomol. 1981, 26, 233–258. [Google Scholar] [CrossRef]
- Pearson, D.L.; Cassola, F. World-wide species richness patterns of tiger-beetles (Coleoptera: Cicindelidae): Indicator taxon for biodiversity and conservation studies. Conserv. Biol. 1992, 6, 376–391. [Google Scholar] [CrossRef]
- Kremen, C.; Colwell, R.K.; Erwin, T.L.; Murphy, D.D.; Noss, R.F.; Saujayan, M.A. Terrestrial arthropod assemblages: Their use as indicators in conservation planning. Conserv. Biol. 1993, 7, 796–808. [Google Scholar] [CrossRef] [Green Version]
- Kremen, C.; Williams, N.M.; Thorp, R.W. Crop pollination from native bees at risk from agricultural intensification. Proc. Natl. Acad. Sci. USA 2002, 99, 16812–16816. [Google Scholar] [CrossRef] [Green Version]
- Klein, A.; Vaissière, B.E.; Cane, J.H.; Stean-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. Royal Soc. B 2007, 274, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Memmott, J.; Craze, P.G.; Waser, N.M.; Price, M.V. Global warming and the disruption of plant–pollinator interactions. Ecol. Lett. 2007, 10, 710–717. [Google Scholar] [CrossRef]
- Goulson, D.; Lye, G.C.; Darvill, B. Decline and Conservation of Bumble Bees. Annu. Rev. Entomol. 2008, 53, 191–208. [Google Scholar] [CrossRef]
- Morrison, J.; Izquierdo, J.; Plaza, E.H.; González-Andújar, J.L.; Hernández, E.; González-Andújar, J.L. The role of field margins in supporting wild bees in Mediterranean cereal agroecosystems: Which biotic and abiotic factors are important? Agr. Ecosyst. Environ. 2017, 247, 216–224. [Google Scholar] [CrossRef]
- AEMET. Agencia Estatal de Meteorología. Available online: http://www.aemet.es/es/portada (accessed on 26 September 2021).
- Aguado, L.O.; Viñuelas, E.; Ferreres, A. Guía de Polinizadores de la Península Ibérica y de los Archipiélagos Balear y Canario; Ediciones Mundiprensa & Syngenta: Madrid, Spain, 2016. [Google Scholar]
- Amiet, F.; Herrmann, M.; Müller, A.; Neumeyer, R. Fauna Helvetica APIDAE 3: Halictus, Lasioglossum; Centre suisse de cartographie de la faune (CSCF Info Fauna) Schweizerische Entomological Gesellschaft (SEG/SES): Bern, Switzerland, 2001. [Google Scholar]
- Amiet, F.; Hermann, M.; Müller, A.; Neumeyer, R. Fauna Helvetica APIDAE 5: Ammobates, Ammobatoides, Anthophora, Biastes, Ceratina, Dasypoda, Epeoloides, Epeolus, Eucera, Macropis, Melecta, Melitta, Nomada, Pasites, Tetralonia, Thyreus, Xylocopa; Centre suisse de cartographie de la faune (CSCF Info Fauna) Schweizerische Entomological Gesellschaft (SEG/SES): Bern, Switzerland, 2007. [Google Scholar]
- Amiet, F.; Hermann, M.; Müller, A.; Neumeyer, R. Fauna Helvetica APIDAE 6: Andrena, Meliturga, Panurginus, Panurgus; Centre suisse de cartographie de la faune (CSCF Info Fauna) Schweizerische Entomological Gesellschaft (SEG/SES): Bern, Switzerland, 2010. [Google Scholar]
- Cobos, A. Fauna Ibérica de Coleópteros Buprestidae; Consejo Superior de Investigaciones Científicas: Madrid, Spain, 1986. [Google Scholar]
- Eizaguirre, S. Coleóptera, Cerambycidae. In Fauna Ibérica; Ramos, M.A., Ed.; Museo Nacional de Ciencias Naturales: Madrid, Spain; CSIC: Madrid, Spain, 2004; Volume 40. [Google Scholar]
- Fernández-Rubio, F. Guía de las Mariposas Diurnas de la Península Ibérica, Baleares, Canarias, Azores y Madeira-1. Libytheidae, Nymphalidae, Riodinidae y Lycaenidae; Ediciones Pirámide, S.A.: Madrid, Spain, 1991. [Google Scholar]
- Fernández-Rubio, F. Guía de las mariposas diurnas de la Península Ibérica, Baleares, Canarias, Azores y Madeira. 2. Papilionidae, Pieridae, Danaidae, Satyridae y Hesperiidae; Ediciones Pirámide, S.A.: Madrid, Spain, 1991. [Google Scholar]
- Fernández-Rubio, F. Clave para la determinación de las especies españolas del género Zygaena Fabricius, 1775 (Insecta: Lepidoptera). Graellsia 2006, 62, 3–12. [Google Scholar] [CrossRef]
- Martín-Piera, F.; López-Colón, J.I. Coleóptera Scarabaeoidea I. In Fauna Ibérica; Ramos, M.A., Ed.; Museo Nacional de Ciencias Naturales: Madrid, Spain; CSIC: Madrid, Spain, 2000; Volume 14. [Google Scholar]
- Oosterbroek, P. The European Families of the Diptera. Identification, Diagnosis, Biology; KNNV Publishing: Utrecht, The Netherlands, 2006. [Google Scholar]
- Scheuchl, E. Illustrierte Bestimmungstabellen der Wildbienen. Band I: Anthophoridae; Preisinger KG: Landhut, Germany, 2000. [Google Scholar]
- Scheuchl, E. Taschenlexikon der Wildbienen Deutschlands und Österreichs. Band I: Megachilidae-Melittidae; Lito Tryk A/S: Berlin, Germany, 2006. [Google Scholar]
- Séméria, Y.; Berland, L. Atlas des Néuroptères de France et d’Europe. Mégaloptères, Raphidioptères, Néuroptères Planipennes, Mécoptères; Société Nouvelle des Éditions Boubée: Paris, France, 1988. [Google Scholar]
- Vives, E. Coleóptera, Cerambycidae. In Fauna Ibérica; Ramos, M.A., Ed.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2000; Volume 12. [Google Scholar]
- The International Union for Conservation of Nature (IUCN) Red List of Threatened Species. Version 2020-1. Available online: https://www.iucnredlist.org (accessed on 8 August 2021).
- Dobson, A.J.; Barnett, A.G. An Introduction to Generalized Linear Models; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Hilbe, J.M. Log negative binomial regression as a generalized linear model. Grad. Coll. Comm. Stat. 1993, 1024, 1–16. [Google Scholar]
- Cameron, A.C.; Trivedi, P.K. Econometric models based on count data. Comparisons and applications on some estimators and tests. J. Appl. Econom. 1986, 1, 29–53. [Google Scholar] [CrossRef]
- R Core Team. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Pungetti, G. Conectividad Ambiental: Las áreas Protegidas en la Cuenca Mediterránea; Consejería de Medio ambiente, Junta de Andalucia: Sevilla, Spain, 2003. [Google Scholar]
- Siemann, E.; Tilman, D.; Haarstad, J.; Ritchie, M. Experimental tests of the dependence of arthropod diversity on plant diversity. Am. Nat. 1998, 152, 738–750. [Google Scholar] [CrossRef]
- Liu, Y.; Duan, M.; Zhang, X.; Zhang, X.; Yu, Z.; Axmacher, J.C.; Stewart, A.; Ewers, R. Effects of plant diversity, habitat and agricultural landscape structure on the functional diversity of carabid assemblages in the North China Plain. Insect Conserv. Diver. 2015, 8, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Danne, A.; Thomson, L.J.; Sharley, D.J.; Penfold, C.M.; Hoffmann, A.A. Effects on native grass cover crops on beneficial and pest invertebrates in Australian vineyards. Pest Manag. 2010, 39, 970–978. [Google Scholar] [CrossRef]
- Eckert, M.; Mathulwe, L.L.; Gaigher, R.; Joubert-van der Merwe, L.; Pryke, J.S. Native cover crops enhance arthropod diversity in vineyards of the Cape Floristic Region. J. Ins. Conser. 2020, 24, 133–149. [Google Scholar] [CrossRef]
- Sáenz-Romo, M.G.; Veas-Bernal, A.; Martínez-García, H.; Campos-Herrera, R.; Ibáñez-Pascual, S.; Martínez-Villar, E.; Pérez-Moreno, I.; Marco-Mancebón, V.S. Ground cover management in a Mediterranean vineyard: Impact on insect abundance and diversity. Agric. Ecosyst. Environ. 2019, 283, 106571. [Google Scholar] [CrossRef]
- Sáenz-Romo, M.G.; Veas-Bernal, A.; Martínez-García, H.; Campos-Herrera, R.; Ibáñez-Pascual, S.; Martínez-Villar, E.; Marco-Mancebón, V.S.; Pérez-Moreno, I. Effects of ground cover management on insect predators and pests in a Mediterranean vineyard. Insects 2019, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Geldenhuys, M.; Gaigher, R.; Pryke, J.S.; Samways, M.J. Diverse herbaceous cover crops promote vineyard arthropod diversity across different management regimes. Agric. Ecosyst. Environ. 2021, 307, 107222. [Google Scholar] [CrossRef]
- Sutter, L.; Jeanneret, P.; Bartual, A.M.; Bocci, G.; Albrecht, M. Enhancing plant diversity in agricultural landscapes promotes both rare bees and dominant crop-pollinating bees through complementary increase in key floral resources. J. Appl. Ecol. 2017, 54, 1856–1864. [Google Scholar] [CrossRef] [Green Version]
- Hannon, L.E.; Sisk, T.D. Hedgerows in an agri-natural landscape: Potential habitat value for native bees. Biol. Conserv. 2009, 142, 2140–2154. [Google Scholar] [CrossRef]
- Miranda-Barroso, L.; Aguado, O.; Falcó-Garí, J.V.; Lopez, D.; Schade, M.; Vasileiadis, V.; Peris-Felipo, F.J. Multifunctional areas as a tool to enhance biodiversity and promote conservation in alfalfa fields. J. Ins. Biodiver. Syst. 2021, 7, 251–261. [Google Scholar]
- Peris-Felipo, F.J.; Santa, F.; Aguado, O.; Falcó-Garí, J.V.; Iborra, A.; Schade, M.; Brittain, C.; Vasileiadis, V.; Miranda-Barroso, L. Enhancement of the diversity of pollinators and beneficial insects in intensively managed vineyards. Insects 2021, 12, 740. [Google Scholar] [CrossRef]
- Carvalheiro, L.G.; Veldtman, R.; Shenkute, A.G.; Tesfay, G.B.; Pirk, C.W.W.; Donaldson, J.S.; Nicolson, S.W. Natural and within farmland biodiversity enhances crop productivity. Ecol. Lett. 2011, 14, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Paiola, A.; Assandri, G.; Brambilla, M.; Zottini, M.; Pedrini, P.; Nascimbene, J. Exploring the potential of vineyards for biodiversity-mediated ecosystem services: A global-scale systematic review. Sci. Total Environ. 2020, 706, 135839. [Google Scholar] [CrossRef]

| Location | 2013 | 2014 | 2015 | ||||
| Field | Margin | Field | Margin | Field | Margin | ||
| Species | Águilas | 26 | 54 | 51 | 79 | 57 | 131 |
| Alcarràs | 11 | 14 | 23 | 36 | 25 | 44 | |
| Fuliola | 12 | 12 | 22 | 42 | 36 | 53 | |
| Individuals | Águilas | 52 | 389 | 122 | 377 | 141 | 747 |
| Alcarràs | 35 | 32 | 91 | 233 | 104 | 399 | |
| Fuliola | 38 | 33 | 73 | 107 | 127 | 205 | |
| Statistics of Goodness of Fit | ||||
|---|---|---|---|---|
| Likelihood ratio (LR) | 104.8 *** | |||
| Deviance (D) | 1420.5 * | |||
| AIC | 1432.5 | |||
| BIC | 1464.5 | |||
| Analysis of Deviance Table | ||||
| Source | LR Chisq | Df | p–value | |
| Farm | 11.42 | 2 | 0.003 | ** |
| Year | 61.62 | 2 | 4.170 × 10−14 | *** |
| Zone | 33.80 | 1 | 6.125 × 10−9 | *** |
| Parameter | Estimate | OR | 2.5% | 97.5% |
|---|---|---|---|---|
| Intercept | −2.85 | 0.06 | 0.04 | 0.08 |
| Location: Alcarras | 0.40 | 1.49 | 1.06 | 2.09 |
| Location: Fuliola | 0.49 | 1.63 | 1.19 | 2.22 |
| Zone: Margin | 0.78 | 2.18 | 1.67 | 2.85 |
| Year: 2014 | 0.85 | 2.35 | 1.64 | 3.41 |
| Year: 2015 | 1.33 | 3.77 | 2.67 | 5.4 |
| Model for the Number of Identified Species | Model for the Number of Identified Individuals | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Source | LR Chisq | Df | p-Value | Source | LR Chisq | Df | p-Value | ||
| Farm | 141.0 | 2 | <2.2 × 10−16 | *** | Farm | 15.1 | 2 | 0.0005293 | *** |
| Zone | 56.8 | 1 | 4.85 × 10−14 | *** | Zone | 128.7 | 1 | <2.2 × 10−16 | *** |
| Year | 103.6 | 2 | <2.2 × 10−16 | *** | Year | 66.3 | 2 | 4.11 × 10−15 | *** |
| Type of species | 21.2 | 1 | 4.09 × 10−6 | *** | Type of species | 1.6 | 1 | 0.2106602 | |
| Model for the Number of Identified Species | Model for the Number of Identified Individuals | ||||||
|---|---|---|---|---|---|---|---|
| Parameter | Estimate | 2.5% | 97.5% | Parameter | Estimate | 2.5% | 97.5% |
| Intercept | 2.70 | 2.48 | 2.92 | Intercept | −0.76 | −1.00 | −0.52 |
| Location: Alcarras | −0.96 | −1.15 | −0.77 | Location: Alcarras | 0.49 | 0.23 | 0.76 |
| Location: Fuliola | −0.81 | −0.99 | −0.64 | Location: Fuliola | 0.01 | −0.24 | 0.27 |
| Zone: Margin | 0.57 | 0.42 | 0.72 | Zone: Margin | 1.20 | 0.99 | 1.40 |
| Year: 2014 | 0.67 | 0.46 | 0.89 | Year: 2014 | 0.56 | 0.31 | 0.81 |
| Year: 2015 | 0.99 | 0.79 | 1.19 | Year: 2015 | 1.03 | 0.78 | 1.27 |
| Type of species: RTE | −0.34 | −0.49 | −0.20 | Type of species: RTE | 0.16 | −0.09 | 0.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santa, F.; Aguado, L.O.; Falcó-Garí, J.V.; Jiménez-Peydró, R.; Schade, M.; Vasileiadis, V.; Miranda-Barroso, L.; Peris-Felipo, F.J. Effectiveness of Multifunctional Margins in Insect Biodiversity Enhancement and RTE Species Conservation in Intensive Agricultural Landscapes. Agronomy 2021, 11, 2093. https://doi.org/10.3390/agronomy11112093
Santa F, Aguado LO, Falcó-Garí JV, Jiménez-Peydró R, Schade M, Vasileiadis V, Miranda-Barroso L, Peris-Felipo FJ. Effectiveness of Multifunctional Margins in Insect Biodiversity Enhancement and RTE Species Conservation in Intensive Agricultural Landscapes. Agronomy. 2021; 11(11):2093. https://doi.org/10.3390/agronomy11112093
Chicago/Turabian StyleSanta, Fernando, Luis Oscar Aguado, José Vicente Falcó-Garí, Ricardo Jiménez-Peydró, Michael Schade, Vasileios Vasileiadis, Luis Miranda-Barroso, and Francisco Javier Peris-Felipo. 2021. "Effectiveness of Multifunctional Margins in Insect Biodiversity Enhancement and RTE Species Conservation in Intensive Agricultural Landscapes" Agronomy 11, no. 11: 2093. https://doi.org/10.3390/agronomy11112093
APA StyleSanta, F., Aguado, L. O., Falcó-Garí, J. V., Jiménez-Peydró, R., Schade, M., Vasileiadis, V., Miranda-Barroso, L., & Peris-Felipo, F. J. (2021). Effectiveness of Multifunctional Margins in Insect Biodiversity Enhancement and RTE Species Conservation in Intensive Agricultural Landscapes. Agronomy, 11(11), 2093. https://doi.org/10.3390/agronomy11112093