1. Introduction
Vitamin A deficiency (VAD) is a major health concern in sub-Saharan Africa (SSA) and many developing countries. Over 190 million pre-school children and 19 million pregnant women in Africa and South Asia are affected by VAD [
1]. In addition, VAD is a risk factor for blindness and mortality from measles and diarrhoea in children aged 6–59 months [
2]. The primary sources of vitamin A are animal-based foods, fresh fruits and vegetables [
3] that are not readily available to poor people who constitute 41% of the population in developing countries [
4], many of whom are rural families. Increasing the concentration of provitamin A carotenoids (PVA) in staple crops, such as maize, is an affordable and durable solution to the problem of VAD. HarvestPlus has developed an online Biofortification Priority Index (BPI) tool that shows that enriching maize with PVA can reduce the prevalence of VAD in developing nations where maize is consumed as staple crop (
www.harvestplus.org/knowledgemarket/BPI, accessed on 9 September 2020).
Considering the loss of up to 50% of carotenoids during storage and processing [
5] and the conversion factor of maize β-carotene to retinol [
6], HarvestPlus has set a biofortification breeding target of 15 μg/g PVA in maize. Pixley et al. [
7] reported maize breeding lines with up to 30 μg/g PVA. However, the most common maize varieties cultivated and consumed globally accumulate less than 2 μg/g PVA [
8]. Considerable efforts have been made to increase PVA concentration in maize cultivars grown in SSA with more than 40 PVA varieties released [
9]. However, the PVA concentrations in these varieties fall short of the target 15 μg/g. There is, therefore, a need to develop hybrid and synthetic varieties with a higher level of PVA concentrations and desirable agronomic performance.
Maize breeders working on PVA biofortification confronted with the challenge of high cost of carotenoid quantification in maize endosperm. High-performance liquid chromatography (HPLC) can cost up to
$100 per sample. Ultra-performance liquid chromatography (UPLC) could be used as an alternative, but this technique is also not affordable given the thousands of samples breeding programs may need to analyse each year. The use of visible yellow to orange kernel color to select genotypes with a high concentration of total carotenoids is limited by the weak correlation with PVA concentration [
8]. DNA markers linked to target loci are now affordable and could accurately screen a large number of genotypes in breeding programs.
The PVA carotenoids accumulated in maize are α-carotene, β-carotene, and β-cryptoxanthin. Biofortified maize varieties also contain non-provitamin A carotenoids such as lutein and zeaxanthin, which are also beneficial to human health [
10]. The carotenoid biosynthesis pathway in maize kernels is well elaborated, and the genes controlling each step have been identified [
11]. Harjes et al. [
8], Yan et al. [
12] and Fu et al. [
13] identified three genes (
LCYE crtRB1 and
PSY1) underlying the critical steps in carotenoid biosynthesis. Gel-based markers associated with both the favorable and unfavorable alleles of the three genes have been developed and their effects on accumulation of PVA and non-PVA carotenoids were validated in tropical maize [
14,
15]. The markers were linked to insertions/deletions (InDels) and single nucleotide polymorphism (SNPs) in different regions of the genes. Sequence analysis of 3′-untranslated region (UTR) of
crtRB1 [
16] and 5′-UTR of
LCYE gene [
17] also detected SNPs and InDels associated with PVA accumulation in maize. Although the gel-based markers have been used for developing maize genotypes with high levels of PVA [
18,
19], the assay is slow and amenable to genotyping a limited number of samples at a time. Furthermore, it is often difficult to visualize the difference between DNA fragments with very small differences in weight. This may require repeating genotyping several times, resulting in increases in the assay cost and delays in the selection process.
To reduce the cost of genotyping and accelerate the rate of genetic gain in carotenoid concentrations in maize, seven Kompetitive Allele-Specific PCR (KASP) SNPs markers associated with the favorable alleles of
crtRB1 gene on chromosome 10 were developed at the International Maize and Wheat Improvement Center (CIMMYT) to select maize with high PVA [
20]. The KASP genotyping is easy to run, accurate and offers flexibility in terms of number of SNPs markers and samples for screening [
21]. Though the KASP genotyping assay was developed for replacing the gel-based genotyping to breed maize for increased carotenoids levels, there are no published reports about the effectiveness of the seven
crtRB1 KASP SNPs markers relative to the gel-based markers to screen maize germplasm for PVA content. Obeng-Bio et al. [
22] used only one of the seven PVA KASP SNPs markers along with
crtRB1 to characterize PVA content in early maturing maize inbred lines. Assessing the effectiveness of the seven
crtRB1 KASP SNPs markers relative to the gel-based markers can validate their usefulness for optimizing selection for high PVA carotenoids in maize. This study was therefore conducted to (i) investigate the comparative effectiveness of PVA KASP SNPs markers relative to the gel-based functional markers for selecting lines with high PVA content and (ii) sequence the PCR products of
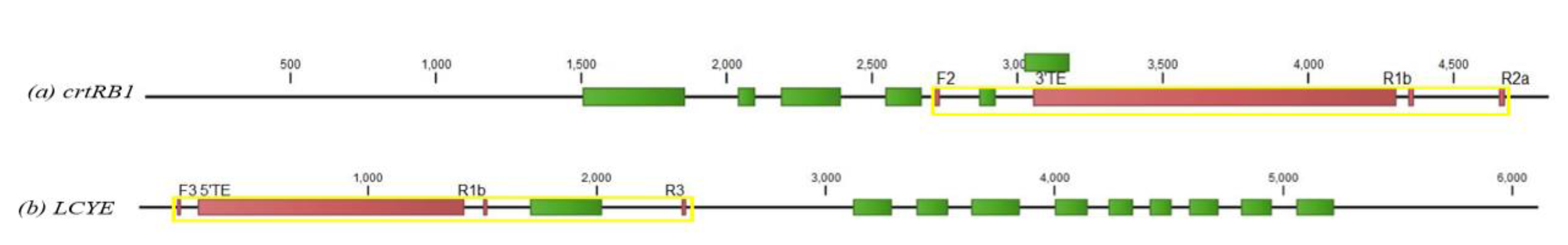
LCYE 5′TE and
crtRB1 3′TE to identify sequence variations separating inbred lines with high and low PVA content.
4. Discussion
The wide ranges in concentrations of the PVA and non-PVA carotenoids detected among inbred lines in our study indicate the suitability of the lines to compare the two types of marker assays. The high repeatability values (0.78 to 0.95) obtained for all carotenoids indicate the high level of accuracy and reliability of the results obtained from carotenoid analyses. These findings are consistent with the results of Egesel et al. [
27], Kurilich and Juvik [
28], Menkir and Maziya-Dixon [
29] and Menkir et al. [
30] reported on maize.
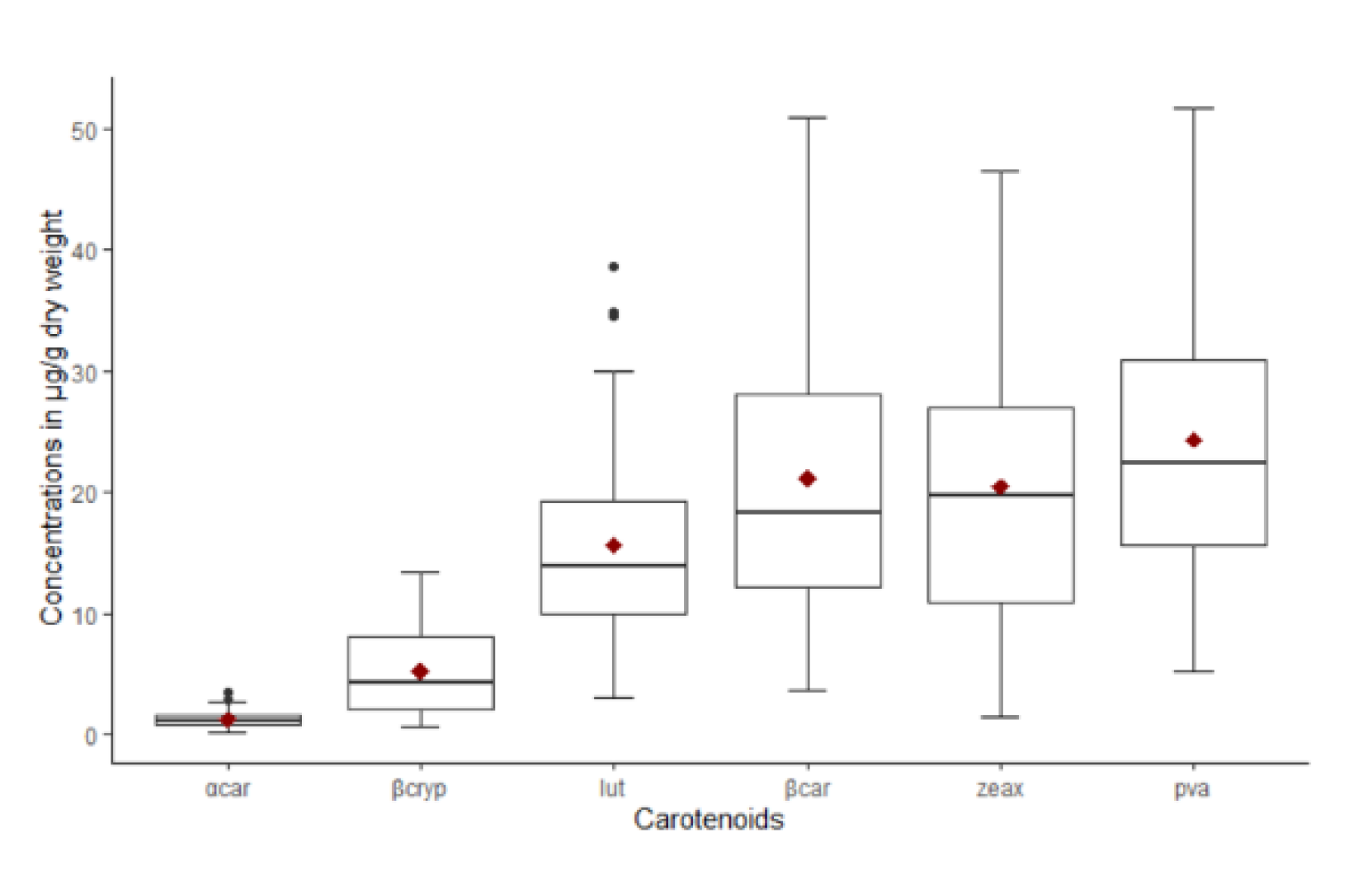
Lutein and zeaxanthine were the predominant non-PVA carotenoids while β-carotene was the dominant one among the PVA carotenoids. The inbred line with the highest PVA concentration (IITATZI1653, 51 µg/g β-carotene) and many other inbreds identified in this study had considerably higher PVA content than those reported in other studies involving tropical inbred lines [
14,
22,
31]. The present study has also identified several inbred lines that have high levels of lutein and zeaxanthin, in addition to high PVA carotenoid content. These inbred lines can be used as promising parents for increasing the concentrations of all beneficial carotenoids for human health.
Of the eight functional gel-based markers of
LCYE [
8],
crtRB1 [
12],
PSY1 [
13], and seven
crtRB1-KASP SNPs markers used to investigate the effect of favorable alleles on carotenoids, only the markers of
LCYE and
crtRB1 were polymorphic while the
PSY markers of were monomorphic in the 64 inbred lines. There are reports of fixation of the
PSY1 gene within and across species [
13,
14]. We found 26 inbred lines carrying the favorable alleles of
crtRB1 that also had high concentrations of β-carotene, consistent with the results in other studies [
14,
22]. These favorable alleles have been found to be the major contributors to high PVA content in maize [
12,
15]. The results obtained using the KASP SNPs markers assay were similar to the results obtained from the gel-based
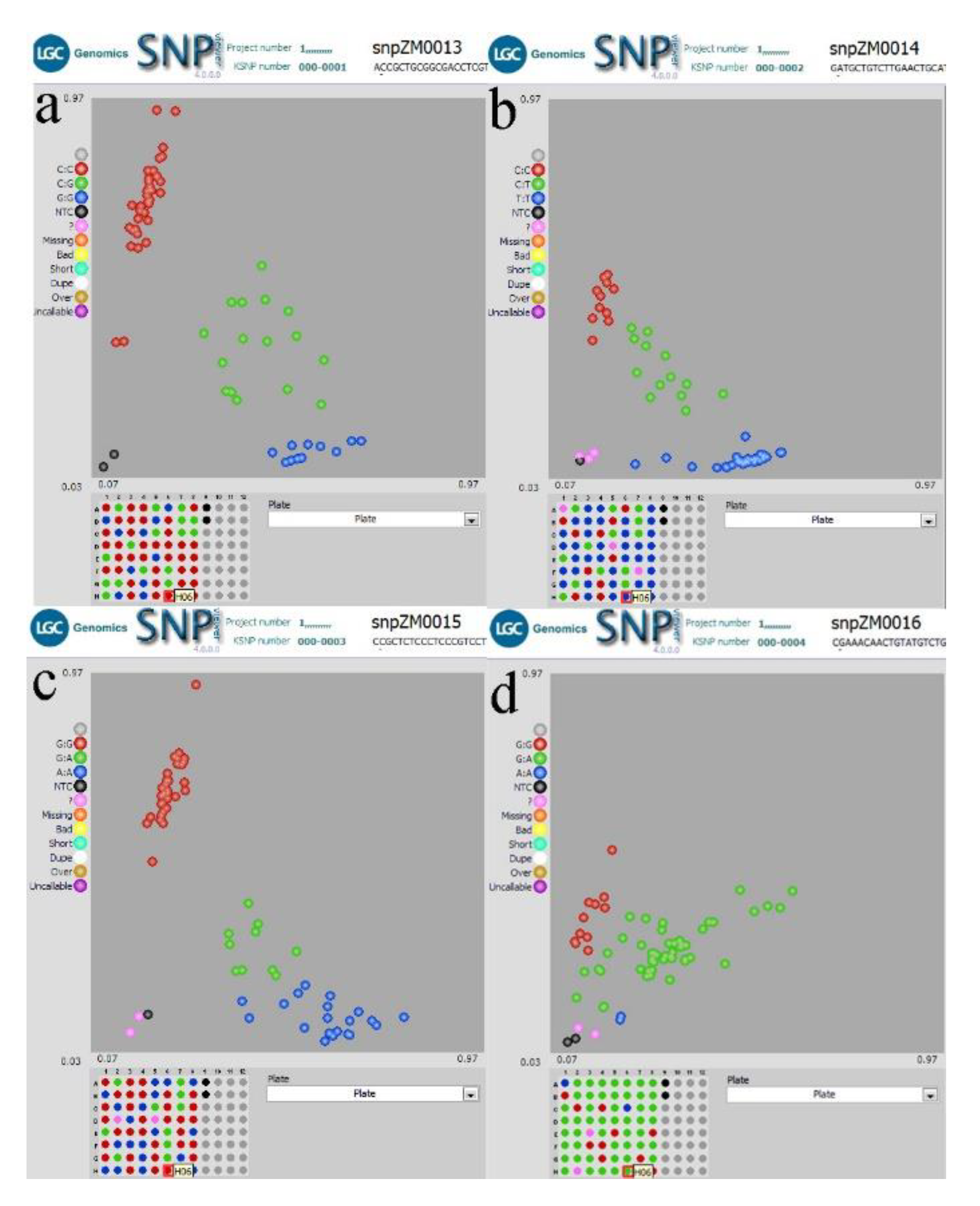
crtRB1 markers in identifying inbred lines with favorable alleles of this gene. Amongst the seven KASP SNP markers, marker snpZM0016 was found to be the most reliable in identifying the largest number of inbred lines carrying favorable alleles. All the inbred lines carrying the favorable alleles of the gel-based
crtRB1-3′TE and
crtRB1-5′TE markers also harboured the favorable allele of snpZM0016. However, three inbred lines, namely IITATZI2015, IITATZI2068 and IITATZI2025, did not carry favorable alleles of almost all the KASP markers while they had favorable alleles for the gel-based
crtRB1-3′TE and
crtRB1-5′TE markers. This contradicts the findings of Obeng-Bio et al. [
22] who reported an agreement between the results obtained using snpZM0015 and the gel-based
crtRB1-3′TE and
crtRB1-5′TE markers. Studies involving a large number of inbred lines with diverse carotenoid composition and content need to be conducted for better understanding of concordance of the gel-based and KASP assays.
The similarity of association of the gel-based and KASP SNPs markers with individual and total carotenoids indicates the effectiveness of the two assays in identifying inbreds with high levels of PVA carotenoids. The favorable alleles of
LCYE gene significantly increased the level of non-PVA carotenoids in the inbred lines included in our study, consistent with the results obtained by Gebremeskel et al. [
31]. In general, the combination of several favorable alleles of
crtRB1 and
LCYE resulted in higher levels of PVA carotenoids. It is reasonable to assume that the favorable allele of
LCYE-3′indel having a significant effect on β-branch carotenoids [
8], in combination with the favorable alleles of
crtRB1, can have a beneficial effect on the accumulation of PVA carotenoid. Yan et al. [
12] evaluated the independent effect of 3′TE alleles on
crtRB1 expression in the endosperm and found that lines with favorable
crtRB1 alleles (1250 bp deletion) had the lowest expression while lines with unfavorable alleles (1250 bp insertion) had the highest expression. The deletion of the last 124 base pairs in exon 6 of the crtRB1 allele present in high beta-carotene maize genotypes could have led to a functional loss of the gene. Moreover, the expression profiling experiment by Harjes et al. [
8] also revealed that lines with insertion of the transposon near the
LCYE transcription start site had a much lower expression of the gene leading to alteration in the ratio of an α- to β-branch carotenoid.
It is noteworthy that some inbred lines that did not carry any of the favorable alleles of
LCYE and
crtRB1 had relatively high PVA carotenoids. These results indicate that genes other than
LCYE and
crtRB1 such as
zep1 and
lut1 [
32] could be associated with the accumulation of PVA carotenoids in these inbred lines. Another possibility is that
SNPs/InDels present in the 5′- and 3′-UTR of
LCYE and
crtRB1 may play a regulatory role in the expression of the genes [
33,
34]. We attempted to find sequence variations in the
LCYE-5′TE and
crtRB1-3′TE genes from 14 maize inbreds having contrasting levels of β-carotene. The sequence variations found in the
LCYE-5′UTR and
crtRB1-3′UTR could not be correlated with the β-carotene accumulation while the
SNP1 found in the intronic region of
LCYE clearly separated the high and low β-carotene genotypes. In general, increases in β-carotene and provitamin A content were associated with decreases in lutein and zeaxanthin in many inbred lines included in our study. Consequently, further research is needed to develop high throughput markers with other genes to complement the KASP assay for accurate screening and identification of inbred lines with high levels of provitamin A and other beneficial carotenoids.


 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}