Silicon Alleviates Temperature Stresses in Poinsettia by Regulating Stomata, Photosynthesis, and Oxidative Damages




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Supplementary Si and Temperature Stress Treatments
2.3. Scanning Electron Microscopy
2.4. Chlorophyll Fluorescence
2.5. H2O2 and MDA Concentrations
2.6. Activities of Antioxidant Enzymes
2.7. Statistical Analysis
3. Results
3.1. Silicon Affected Stomata and Epicuticular Wax of Poinsettia
3.2. Silicon Maintained Photosynthesis in Poinsettia
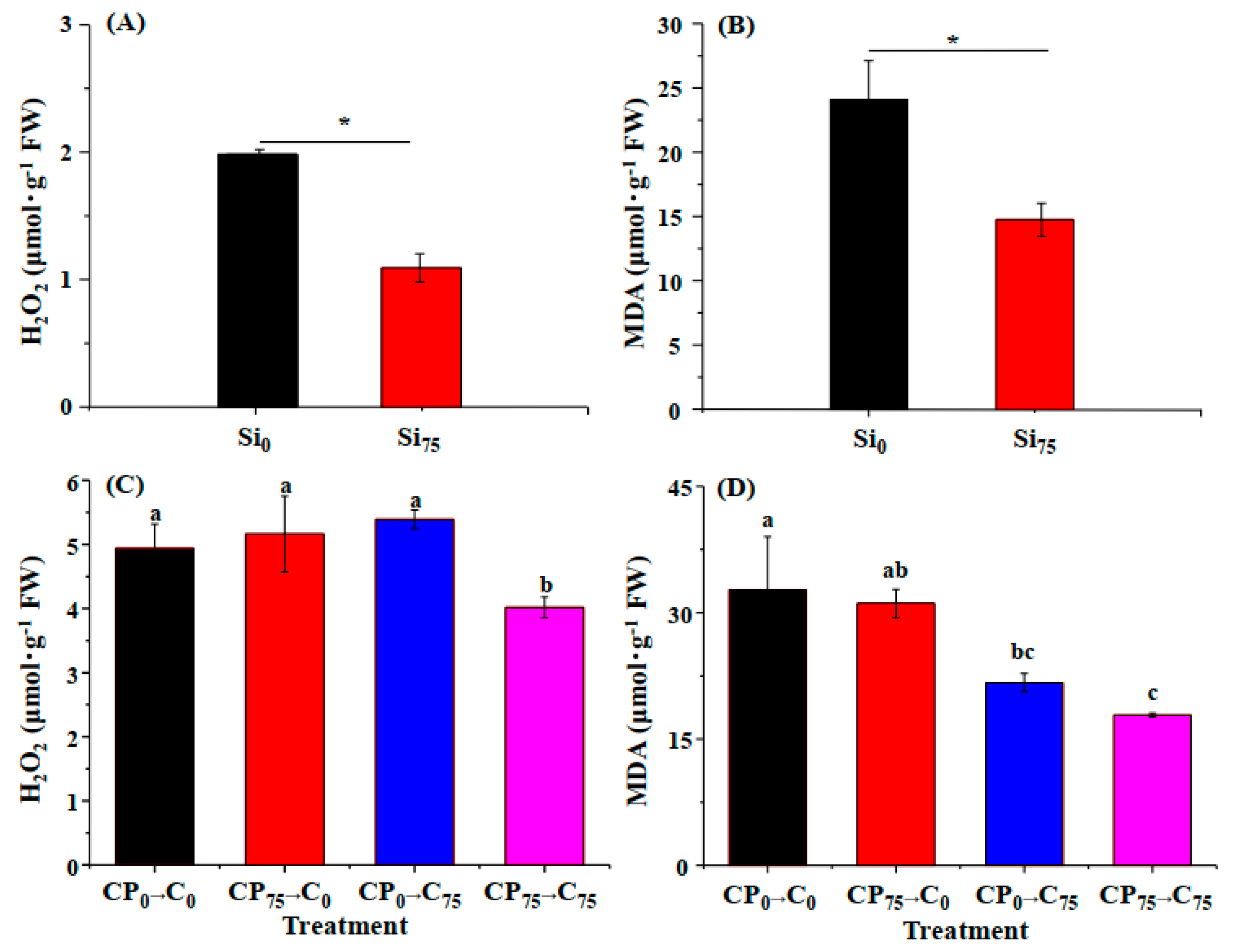
3.3. Concentration of H2O2 and MDA Was Low in Silicon-Treated Cuttings
3.4. APX Activity Was Low in Silicon-Treated Seedlings
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Le, B.; Nadeem, M.; Yang, S.-H.; Shin, J.A.; Kang, M.G.; Chung, G.; Sun, S. Effect of silicon in Pyropia yezoensis under temperature and irradiance stresses through antioxidant gene expression. J. Appl. Phycol. 2019, 31, 1297–1302. [Google Scholar] [CrossRef]
- Kim, Y.H.; Khan, A.L.; Waqas, M.; Lee, I.J. Silicon regulates antioxidant activities of crop plants under abiotic-induced oxidative stress: A review. Front. Plant Sci. 2017, 8, 510. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Park, Y.G.; Kim, S.; Jeong, B.R. Foliar or subirrigation silicon supply mitigates high temperature stress in strawberry by maintaining photosynthetic and stress-responsive proteins. J. Plant Growth Regul. 2017, 36, 836–845. [Google Scholar] [CrossRef]
- Malhotra, C.; Kapoor, R.T. Silicon: A sustainable tool in abiotic stress tolerance in plants. In Plant Abiotic Stress Tolerance; Springer: Cham, Switzerland, 2019; pp. 333–356. [Google Scholar]
- Moradtalab, N.; Weinmann, M.; Walker, F.; Hoglinger, B.; Ludewig, U.; Neumann, G. Silicon improves chilling tolerance during early growth of maize by effects on micronutrient homeostasis and hormonal balances. Front. Plant Sci. 2018, 9, 420. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.P.; Chao, D.Y.; Lin, H.X. Understanding abiotic stress tolerance mechanisms: Recent studies on stress response in rice. J. Integr. Plant Biol. 2007, 49, 742–750. [Google Scholar] [CrossRef]
- Nakashima, K.; Takasaki, H.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. NAC transcription factors in plant abiotic stress responses. BBA-Gene Regul. Mech. 2012, 1819, 97–103. [Google Scholar] [CrossRef]
- Liang, Y.; Nikolic, M.; Bélanger, R.; Gong, H.; Song, A. Silicon in Agriculture; Springer: Dordrecht, The Netherlands, 2015; pp. 978–994. [Google Scholar]
- Park, C.S. Past and future advances in silicon research in the Republic of Korea. In Studies in Plant Science; Elsevier: Amsterdam, The Netherlands, 2001; pp. 359–371. [Google Scholar]
- Dallagnol, L.J.; Rodrigues, F.A.; DaMatta, F.M.; Mielli, M.V.; Pereira, S.C. Deficiency in silicon uptake affects cytological, physiological, and biochemical events in the rice–Bipolaris oryzae interaction. Phytopathology 2011, 101, 92–104. [Google Scholar] [CrossRef]
- Miyake, Y.; Takahashi, E. Silicon deficiency of tomato plant. Soil Sci. Plant Nutr. 1978, 24, 175–189. [Google Scholar] [CrossRef]
- Schaller, J.; Brackhage, C.; Gessner, M.O.; Bäuker, E.; Gert Dudel, E. Silicon supply modifies C:N:P stoichiometry and growth of Phragmites australis. Plant Biol. 2012, 14, 392–396. [Google Scholar] [CrossRef]
- Haldimann, P.; Feller, U. Inhibition of photosynthesis by high temperature in oak (Quercus pubescens L.) leaves grown under natural conditions closely correlates with a reversible heat-dependent reduction of the activation state of ribulose-1, 5-bisphosphate carboxylase/oxygenase. Plant Cell Environ. 2004, 27, 1169–1183. [Google Scholar] [CrossRef]
- Reynolds-Henne, C.E.; Langenegger, A.; Mani, J.; Schenk, N.; Zumsteg, A.; Feller, U. Interactions between temperature, drought and stomatal opening in legumes. Environ. Exp. Bot. 2010, 68, 37–43. [Google Scholar] [CrossRef]
- Ranney, T.G.; Peet, M.M. Heat tolerance of five taxa of birch (Betula): Physiological responses to supraoptimal leaf temperatures. J. Am. Soc. Hortic. Sci. 1994, 119, 243–248. [Google Scholar] [CrossRef]
- Bassow, S.; McConnaughay, K.; Bazzaz, F. The response of temperate tree seedlings grown in elevated CO2 to extreme temperature events. Ecol. Appl. 1994, 4, 593–603. [Google Scholar] [CrossRef]
- Samuelson, L.J.; Teskey, R.O. Net photosynthesis and leaf conductance of loblolly pine seedlings in 2 and 21% oxygen as influenced by irradiance, temperature and provenance. Tree Physiol. 1991, 8, 205–211. [Google Scholar] [CrossRef]
- Al-Khatib, K.; Paulsen, G.M. Enhancement of thermal injury to photosynthesis in wheat plants and thylakoids by high light intensity. Plant Physiol. 1989, 90, 1041–1048. [Google Scholar] [CrossRef]
- Dufrene, E.; Saugier, B. Gas exchange of oil palm in relation to light, vapour pressure deficit, temperature and leaf age. Funct. Ecol. 1993, 7, 97–104. [Google Scholar] [CrossRef]
- Kozlowski, T.; Pallardy, S. Effects of low temperature on leaf diffusion resistance of Ulmus americana and Fraxinus pennsylvanica seedlings. Can. J. Bot. 1979, 57, 2466–2470. [Google Scholar] [CrossRef]
- Lösch, R. Responses of stomata to environmental factors-experiments with isolated epidermal strips of Polypodium vulgare. Oecologia 1977, 29, 85–97. [Google Scholar] [CrossRef]
- Wilkinson, S.; Clephan, A.L.; Davies, W.J. Rapid low temperature-induced stomatal closure occurs in cold-tolerant Commelina communis leaves but not in cold-sensitive tobacco leaves, via a mechanism that involves apoplastic calcium but not abscisic acid. Plant Physiol. 2001, 126, 1566–1578. [Google Scholar] [CrossRef]
- Wise, R.; Olson, A.; Schrader, S.; Sharkey, T. Electron transport is the functional limitation of photosynthesis in field-grown Pima cotton plants at high temperature. Plant Cell Environ. 2004, 27, 717–724. [Google Scholar] [CrossRef]
- Havaux, M. Characterization of thermal damage to the photosynthetic electron transport system in potato leaves. Plant Sci. 1993, 94, 19–33. [Google Scholar] [CrossRef]
- Sinsawat, V.; Leipner, J.; Stamp, P.; Fracheboud, Y. Effect of heat stress on the photosynthetic apparatus in maize (Zea mays L.) grown at control or high temperature. Environ. Exp. Bot. 2004, 52, 123–129. [Google Scholar] [CrossRef]
- Fracheboud, Y.; Haldimann, P.; Leipner, J.; Stamp, P. Chlorophyll fluorescence as a selection tool for cold tolerance of photosynthesis in maize (Zea mays L.). J. Exp. Bot. 1999, 50, 1533–1540. [Google Scholar] [CrossRef]
- Kościelniak, J.; Biesaga-Kościelniak, J. Photosynthesis and non-photochemical excitation quenching components of chlorophyll excitation in maize and field bean during chilling at different photon flux density. Photosynthetica 2006, 44, 174–180. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, L.; He, J.; Qian, L.; Wu, L.; Wang, R. Characteristics of chlorophyll fluorescence and antioxidative system in super-hybrid rice and its parental cultivars under chilling stress. Biol. Plant. 2010, 54, 164–168. [Google Scholar] [CrossRef]
- Wang, G.L.; Guo, Z.F. Effects of chilling stress on photosynthetic rate and chlorophyll fluorescence parameter in seedlings of two rice cultivars differing in cold tolerance. Rice Sci. 2005, 12, 187–191. [Google Scholar]
- Melkonian, J.; Owens, T.G.; Wolfe, D.W. Gas exchange and co-regulation of photochemical and nonphotochemical quenching in bean during chilling at ambient and elevated carbon dioxide. Photosynth. Res. 2004, 79, 71–82. [Google Scholar] [CrossRef]
- Lin, K.; Hwang, W.; Lo, H. Chilling stress and chilling tolerance of sweet potato as sensed by chlorophyll fluorescence. Photosynthetica 2007, 45, 628–632. [Google Scholar] [CrossRef]
- Islam, S.; Izekor, E.; Garner, J. Effect of chilling stress on the chlorophyll fluorescence, peroxidase activity and other physiological activities in Ipomoea batatas L. genotypes. Am. J. Plant Physiol. 2011, 6, 72–82. [Google Scholar] [CrossRef]
- Frolec, J.; Ilík, P.; Krchňák, P.; Sušila, P.; Nauš, J. Irreversible changes in barley leaf chlorophyll fluorescence detected by the fluorescence temperature curve in a linear heating/cooling regime. Photosynthetica 2008, 46, 537–546. [Google Scholar] [CrossRef]
- Richards, G.; Hall, D. Photoinhibition at chilling temperatures in intact leaves and isolated chloroplasts of barley. In Progress in Photosynthesis Research; Springer: Dordrecht, The Netherlands, 1987; pp. 39–42. [Google Scholar]
- Dai, Y.L.; Xu, C.H.; Zhao, F.H. Comparative studies on effects of low temperature on chlorophyll fluorescence induction kinetics and photochemcal activities of cucumber and winter wheat. In Progress in Photosynthesis Research; Springer: Dordrecht, The Netherlands, 1987; pp. 99–102. [Google Scholar]
- Dong, Z.; Men, Y.; Li, Z.; Zou, Q.; Ji, J. Chlorophyll fluorescence imaging as a tool for analyzing the effects of chilling injury on tomato seedlings. Sci. Hortic. 2019, 246, 490–497. [Google Scholar] [CrossRef]
- Zushi, K.; Kajiwara, S.; Matsuzoe, N. Chlorophyll a fluorescence OJIP transient as a tool to characterize and evaluate response to heat and chilling stress in tomato leaf and fruit. Sci. Hortic. 2012, 148, 39–46. [Google Scholar] [CrossRef]
- Brestic, M.; Zivcak, M. PSII fluorescence techniques for measurement of drought and high temperature stress signal in crop plants: Protocols and applications. In Molecular Stress Physiology of Plants; Springer: New Delhi, India, 2013; pp. 87–131. [Google Scholar]
- Janka, E.; Körner, O.; Rosenqvist, E.; Ottosen, C.-O. High temperature stress monitoring and detection using chlorophyll a fluorescence and infrared thermography in chrysanthemum (Dendranthema grandiflora). Plant Physiol. Bioch. 2013, 67, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Bussotti, F.; Desotgiu, R.; Pollastrini, M.; Cascio, C. The JIP test: A tool to screen the capacity of plant adaptation to climate change. Scand. J. Forest Res. 2010, 25, 43–50. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Bosa, K.; Kościelniak, J.; Hossain, Z. Chlorophyll a fluorescence—A useful tool for the early detection of temperature stress in spring barley (Hordeum vulgare L.). Omics 2011, 15, 925–934. [Google Scholar] [CrossRef]
- Malenčić, D.; Vasić, D.; Popović, M.; Dević, D. Antioxidant systems in sunflower as affected by oxalic acid. Biol. Plant. 2004, 48, 243–247. [Google Scholar] [CrossRef]
- Gülen, H.; Çetinkaya, C.; Kadıoğlu, M.; Kesici, M.; Cansev, A.; Eriş, A. Peroxidase activity and lipid peroxidation in strawberry (Fragaria x ananassa) plants under low temperature. J. Biol. Environ. Sci. 2008, 2, 95–100. [Google Scholar]
- Latef, A.A.H.A.; He, C. Arbuscular mycorrhizal influence on growth, photosynthetic pigments, osmotic adjustment and oxidative stress in tomato plants subjected to low temperature stress. Acta Physiol. Plant. 2011, 33, 1217–1225. [Google Scholar] [CrossRef]
- Li, H.; Qiang, S.; Qian, Y. Physiological response of different croftonweed (Eupatorium adenophorum) populations to low temperature. Weed Sci. 2008, 56, 196–202. [Google Scholar] [CrossRef]
- Davey, M.; Stals, E.; Panis, B.; Keulemans, J.; Swennen, R. High-throughput determination of malondialdehyde in plant tissues. Anal. Biochem. 2005, 347, 201–207. [Google Scholar] [CrossRef]
- Yamauchi, Y.; Furutera, A.; Seki, K.; Toyoda, Y.; Tanaka, K.; Sugimoto, Y. Malondialdehyde generated from peroxidized linolenic acid causes protein modification in heat-stressed plants. Plant Physiol. Biochem. 2008, 46, 786–793. [Google Scholar] [CrossRef] [PubMed]
- da-Silva, C.J.; Rodrigues, A.C.; Modolo, L.V. H2O2, NO, and H2S: Tailoring in suiting plants against abiotic stresses. In Reactive Oxygen, Nitrogen and Sulfur Species in Plants: Production, Metabolism, Signaling and Defense Mechanisms; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2019; pp. 841–856. [Google Scholar]
- Hossain, M.A.; Bhattacharjee, S.; Armin, S.M.; Qian, P.; Xin, W.; Li, H.Y.; Tran, L.S.P. Hydrogen peroxide priming modulates abiotic oxidative stress tolerance: Insights from ROS detoxification and scavenging. Front. Plant Sci. 2015, 6, 420. [Google Scholar] [CrossRef] [PubMed]
- Chaitanya, K.; Sundar, D.; Masilamani, S.; Reddy, A.R. Variation in heat stress-induced antioxidant enzyme activities among three mulberry cultivars. Plant Growth Regul. 2002, 36, 175–180. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, L.; Xin, Y.; Wang, W.N.; He, W.Y.; Wang, A.L.; Liu, Y. Effect of temperature on antioxidant enzyme gene expression and stress protein response in white shrimp, Litopenaeus vannamei. J. Therm. Biol. 2010, 35, 284–289. [Google Scholar] [CrossRef]
- Almeselmani, M.; Deshmukh, P.; Sairam, R.; Kushwaha, S.; Singh, T. Protective role of antioxidant enzymes under high temperature stress. Plant Sci. 2006, 171, 382–388. [Google Scholar] [CrossRef]
- Hu, J.; Cai, X.; Jeong, B.R. Silicon affects root development, tissue mineral content, and expression of silicon transporter genes in poinsettia (Euphorbia pulcherrima Willd.) cultivars. Plants 2019, 8, 180. [Google Scholar] [CrossRef]
- Manivannan, A.; Soundararajan, P.; Halimah, N.; Ko, C.H.; Jeong, B.R. Blue led light enhances growth, phytochemical contents, and antioxidant enzyme activities of Rehmannia glutinosa cultured in vitro. Hortic. Environ. Biotechnol. 2015, 56, 105–113. [Google Scholar] [CrossRef]
- Morikawa, C.; Saigusa, M. Mineral composition and accumulation of silicon in tissues of blueberry (Vaccinum corymbosus cv. Bluecrop) cuttings. Plant Soil 2004, 258, 1–8. [Google Scholar] [CrossRef]
- Blanke, M.M.; Prung, R.J.; Baker, E.A. Structure and elemental compositon of grape berry stomata. J. Plant Physiol. 1999, 154, 477–481. [Google Scholar] [CrossRef]
- Agarie, S.; Uchida, H.; Agata, W.; Kubota, F.; Kaufman, P.B. Effects of silicon on transpiration and leaf conductance in rice plants (Oryza sativa L.). Plant Prod. Sci. 1998, 1, 89–95. [Google Scholar] [CrossRef]
- Gao, X.; Zou, C.; Wang, L.; Zhang, F. Silicon improves water use efficiency in maize plants. J. Plant Nutr. 2005, 27, 1457–1470. [Google Scholar] [CrossRef]
- Gao, X.; Zou, C.; Wang, L.; Zhang, F. Silicon decreases transpiration rate and conductance from stomata of maize plants. J. Plant Nutr. 2006, 29, 1637–1647. [Google Scholar] [CrossRef]
- Crawford, A.J.; McLachlan, D.H.; Hetherington, A.M.; Franklin, K.A. High temperature exposure increases plant cooling capacity. Curr. Biol. 2012, 22, R396–R397. [Google Scholar] [CrossRef] [PubMed]
- Huggins, T.; Mohammed, S.; Sengodon, P.; Ibrahim, A.; Tilley, M.; Hays, D. Changes in leaf epicuticular wax load and its effect on leaf temperature and physiological traits in wheat cultivars (Triticum aestivum L.) exposed to high temperatures during anthesis. J. Agron. Crop Sci. 2018, 204, 49–61. [Google Scholar] [CrossRef]
- Asmar, S.A.; Pasqual, M.; de Araujo, A.G.; Silva, R.A.L.; Rodrigues, F.A.; Pio, L.A.S. Morphophysiological characteristics of acclimatized ‘Grande Naine’ banana plants in response to in vitro use of silicon. Semin. Ciênc. Agrár. 2013, 34, 73–82. [Google Scholar] [CrossRef][Green Version]
- Braga, F.T.; Nunes, C.F.; Favero, A.C.; Pasqual, M.; de Carvalho, J.G.; de Castro, E.M. Anatomical characteristics of the strawberry seedlings micropropagated using different sources of silicon. Pesqui. Agropecu. Bras. 2009, 44, 128–132. [Google Scholar] [CrossRef]
- Ahmed, M.; Qadeer, U.; Ahmed, Z.I.; Hassan, F.-U. Improvement of wheat (Triticum aestivum) drought tolerance by seed priming with silicon. Arch. Agron. Soil Sci. 2016, 62, 299–315. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar]
- Ortega-Villasante, C.; Rellan-Alvarez, R.; Del Campo, F.F.; Carpena-Ruiz, R.O.; Hernandez, L.E. Cellular damage induced by cadmium and mercury in Medicago sativa. J. Exp. Bot. 2005, 56, 2239–2251. [Google Scholar] [CrossRef]
- Sun, W.H.; Duan, M.; Li, F.; Shu, D.F.; Yang, S.; Meng, Q.W. Overexpression of tomato tAPX gene in tobacco improves tolerance to high or low temperature stress. Biol. Plant. 2010, 54, 614–620. [Google Scholar] [CrossRef]
- Park, M.R.; Kim, Y.J. Enhanced antioxidant enzymes are associated with reduced hydrogen peroxide in barley roots under saline stress. J. Biochem. Mol. Biol. 2005, 38, 218–224. [Google Scholar]
- Yong, Z.; Hao-Ru, T.; Ya, L. Variation in antioxidant enzyme activities of two strawberry cultivars with short-term low temperature stress. World J. Agric. Sci. 2008, 4, 458–462. [Google Scholar]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Li, Y.; Jeong, B.R. Silicon Alleviates Temperature Stresses in Poinsettia by Regulating Stomata, Photosynthesis, and Oxidative Damages. Agronomy 2020, 10, 1419. https://doi.org/10.3390/agronomy10091419
Hu J, Li Y, Jeong BR. Silicon Alleviates Temperature Stresses in Poinsettia by Regulating Stomata, Photosynthesis, and Oxidative Damages. Agronomy. 2020; 10(9):1419. https://doi.org/10.3390/agronomy10091419
Chicago/Turabian StyleHu, Jiangtao, Yali Li, and Byoung Ryong Jeong. 2020. "Silicon Alleviates Temperature Stresses in Poinsettia by Regulating Stomata, Photosynthesis, and Oxidative Damages" Agronomy 10, no. 9: 1419. https://doi.org/10.3390/agronomy10091419
APA StyleHu, J., Li, Y., & Jeong, B. R. (2020). Silicon Alleviates Temperature Stresses in Poinsettia by Regulating Stomata, Photosynthesis, and Oxidative Damages. Agronomy, 10(9), 1419. https://doi.org/10.3390/agronomy10091419