Sheep Dung Composition and Phosphorus and Potassium Release Affected by Grazing Intensity and Pasture Development Stage in an Integrated Crop-Livestock System

 ,
, 
 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. History of the Experimental Area
2.2. Experimental Design and Conduction
2.3. Dung Sampling and Composition Analysis
2.4. Dung Decomposition and Nutrient Release over Time
2.5. Statistical Analysis
3. Results
3.1. Dung Composition and Daily Nutrient Return to the System
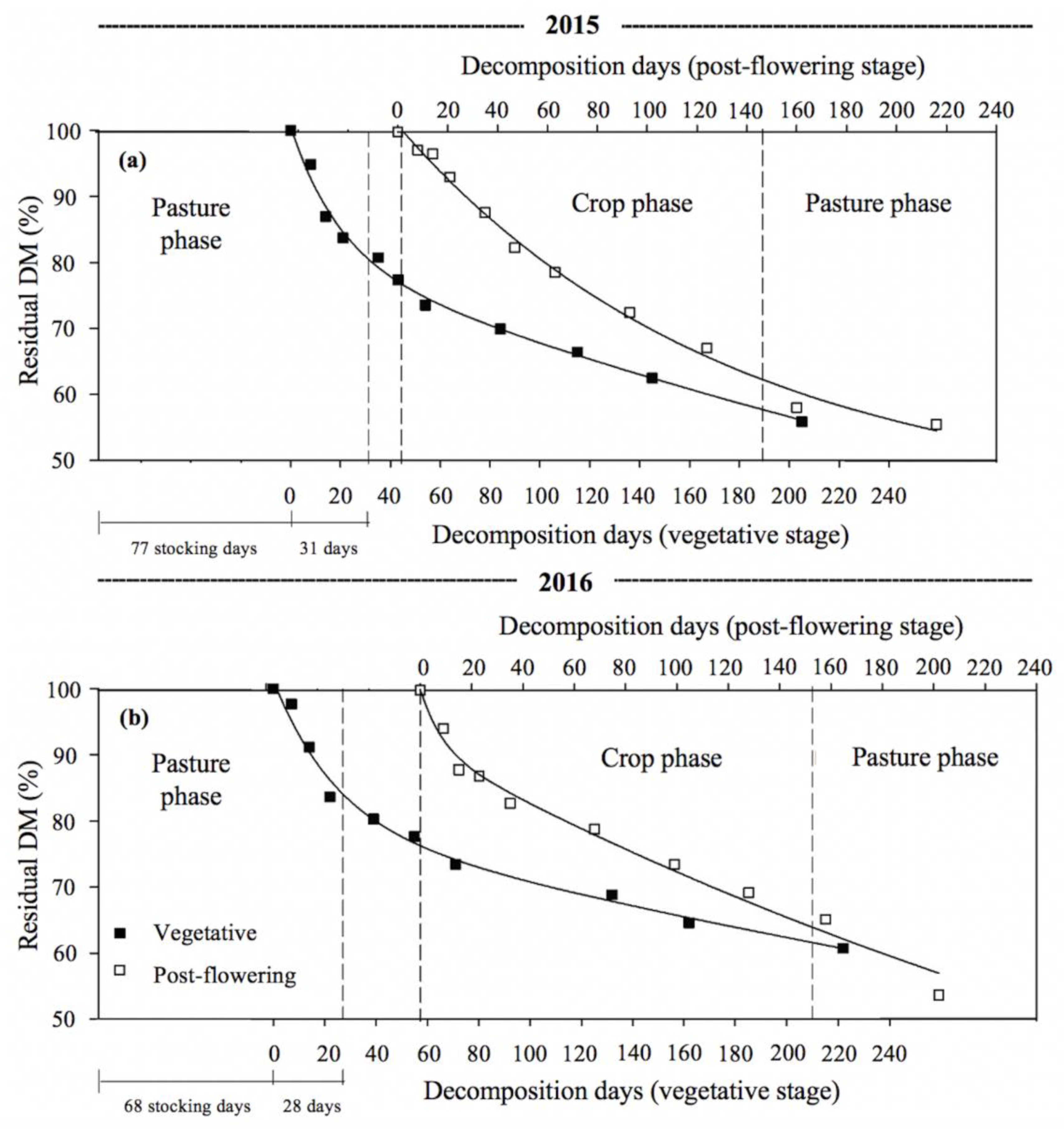
3.2. Dung Decomposition
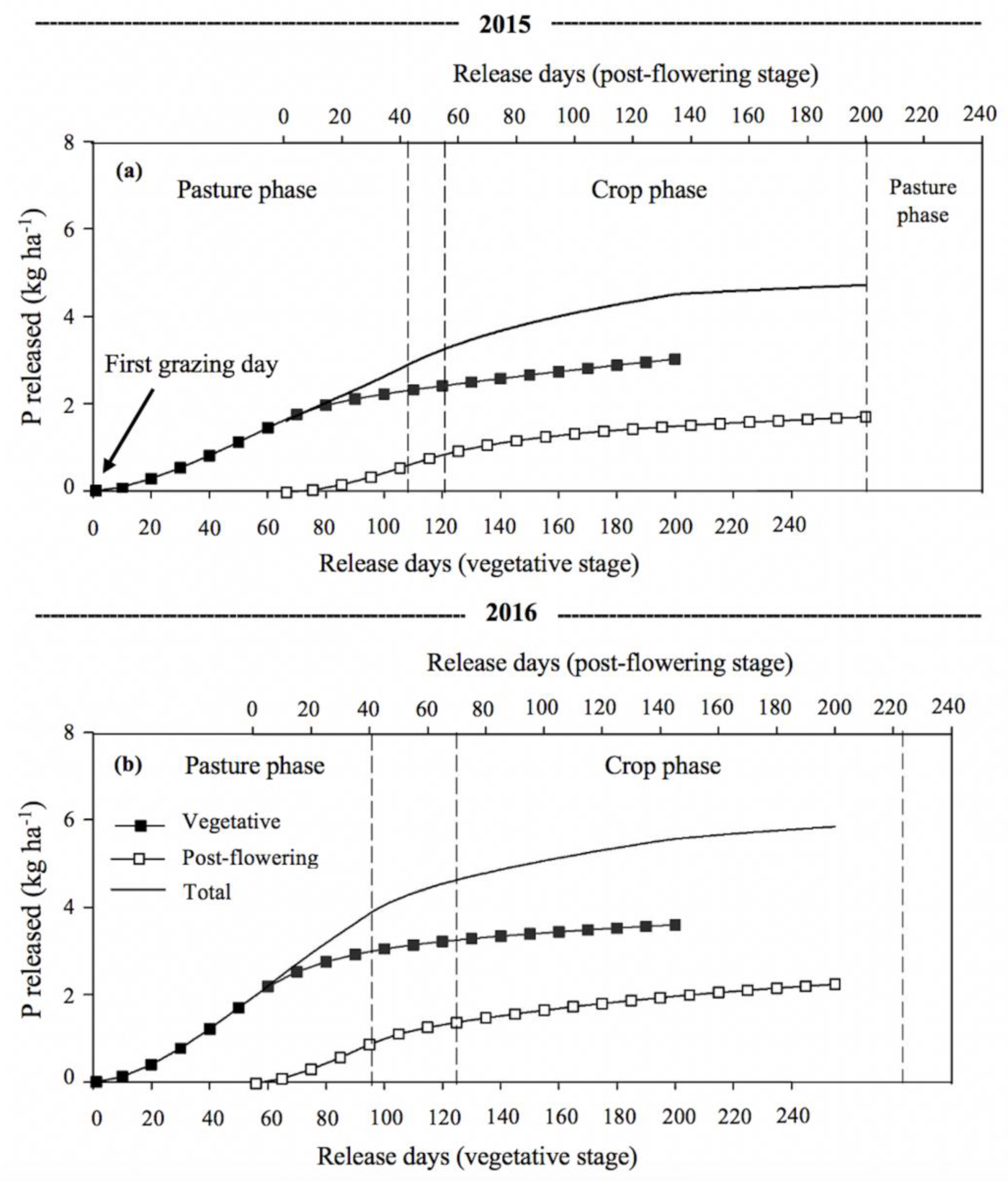
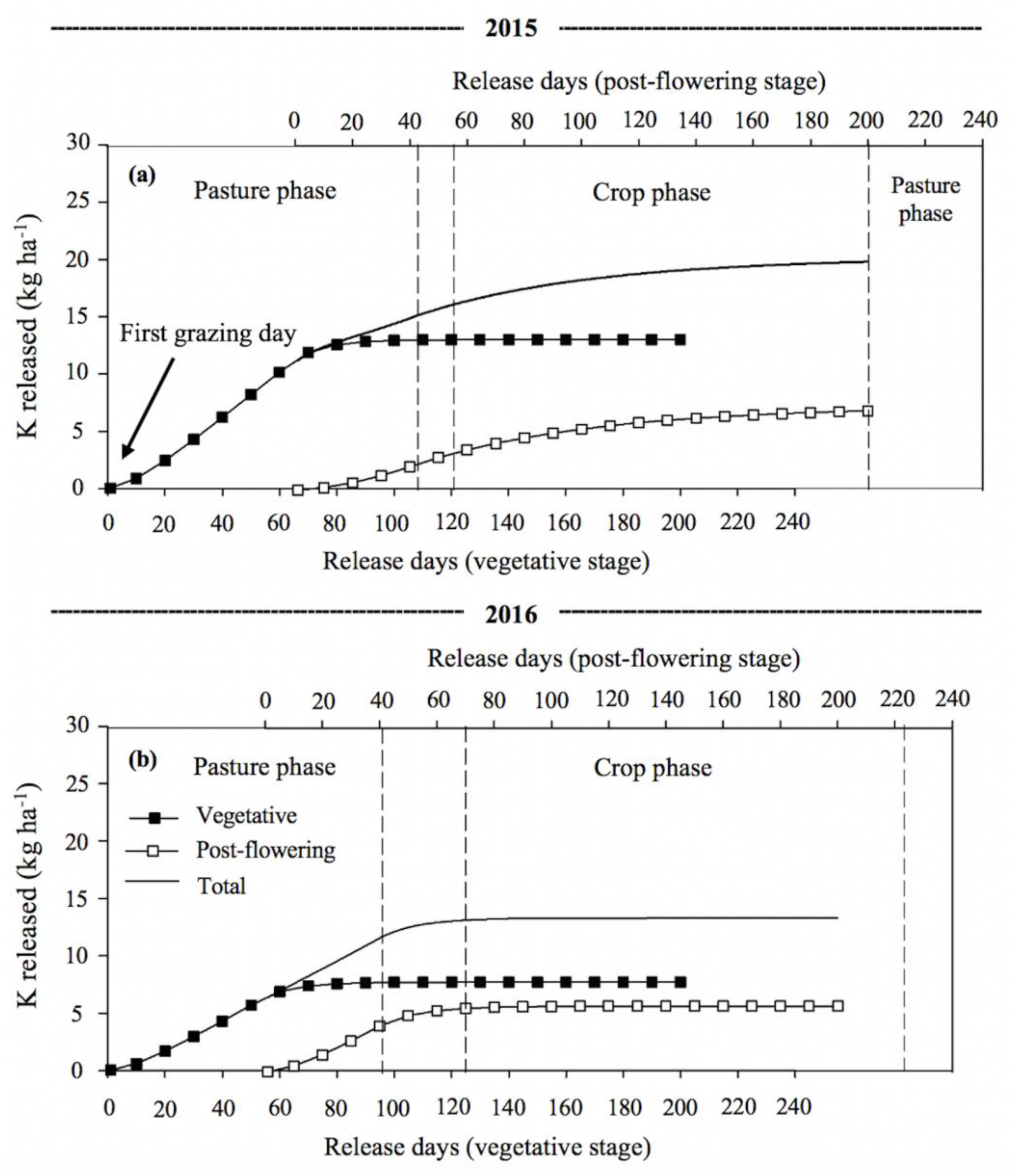
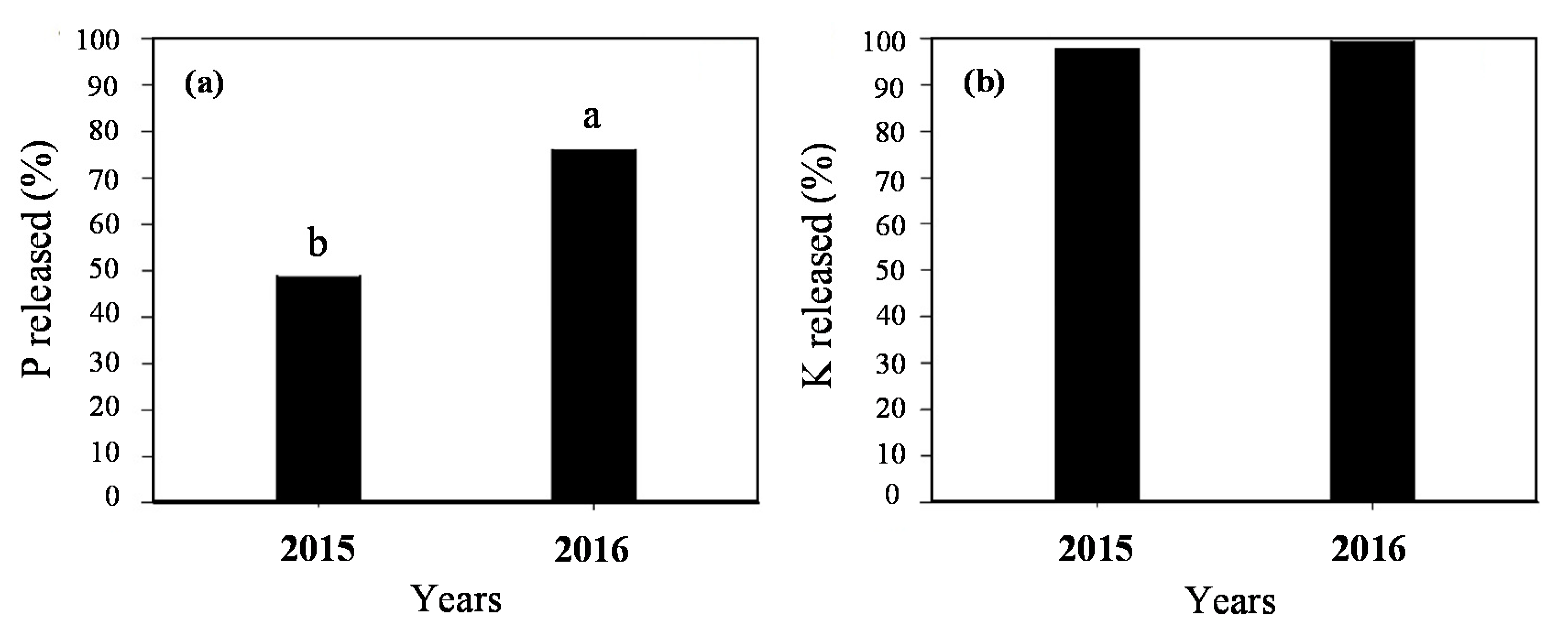
3.3. Phosphorus and Potassium Release
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sulc, R.M.; Franzluebbers, A.J. Exploring integrated crop-livestock systems in different ecoregions ofthe United States. Eur. J. Agron. 2004, 57, 21–30. [Google Scholar] [CrossRef]
- Peterson, C.A.; Pedro, A.D.A.; Martins, A.P.; Bergamaschi, H.; Anghinoni, I.; Carvalho, P.C.F.; Gaudin, A.C. Winter grazing does not affect soybean yield despite lower soil water content in a subtropical crop-livestock system. Agron. Sustain. Dev. 2019, 39, 26. [Google Scholar] [CrossRef]
- Lemaire, G.; Franzluebbers, A.J.; Carvalho, P.C.F.; Dedieu, B. Integrated crop-livestock systems: Strategies to achieve synergybetween agricultural production and environmental quality. Agric. Ecosyst. Environ. 2014, 190, 4–8. [Google Scholar] [CrossRef]
- Martens, J.R.T.; Entz, M.H. Integrating green manure and grazing systems: A review. Can. J. Plant. Sci. 2011, 91, 811–824. [Google Scholar] [CrossRef]
- Alves, L.A.; Denardin, L.G.O.; Martins, A.P.; Anghinoni, I.; Carvalho, P.C.F.; Tiecher, T. Soil acidification and P, K, Ca and Mg budget as affected by sheep grazing and crop rotation in a long-term integrated crop-livestock system in southern Brazil. Geoderma 2019, 351, 197–208. [Google Scholar] [CrossRef]
- Whitehead, D.C. Nutrient Elements in Grassland: Soil-Plant-Animal Relationships; CABI Publishing: Wallingford, UK, 2020. [Google Scholar]
- Chávez, L.F.; Escobar, L.F.; Anghinoni, I.; Carvalho, P.F.C.; Meurer, E.J. Diversidade metabólica e atividade microbiana no solo em sistema de integração lavoura-pecuária sob intensidades de pastejo. Pesq Agropec. Bras. 2011, 46, 1254–1261. [Google Scholar] [CrossRef]
- Denardin, L.G.O.; Martins, A.P.; Carmona, F.C.; Veloso, M.G.; Carmona, G.I.; Carvalho, P.C.F.; Anghinoni, I. Integrated crop-livestock systems in paddy fields: New strategies for flooded rice nutrition. Agron. J. 2020, 112, 2219–2229. [Google Scholar] [CrossRef]
- Kunrath, T.R.; Nunes, P.A.A.; Filho, W.S.; Cadenazzi, M.; Bremm, C.; Martins, A.P.; Carvalho, P.C.F. Sward height determines pasture production and animal performance in a long-term soybean-beef cattle integrated system. Agric. Syst. 2020, 177, 102716. [Google Scholar] [CrossRef]
- Matthew, C.; Assuero, S.; Black, C.K.; Sackville Hamilton, N.R. Tiller dynamics of grazed swards. In Grassland Ecophysiology and Grazing Ecology; Lemaire, G., Hodgson, J., Moraes, A., Carvalho, P.C.F., Nabinger, C., Eds.; CABI Publishing, Lusignan: Wallingford, UK, 2020; pp. 127–150. [Google Scholar] [CrossRef]
- Filho, W.S.; Nunes, P.A.A.; Barro, R.S.; Kunrath, T.R.; Almeida, G.M.; Genro, T.C.M.; Bayer, C.; Carvalho, P.C.F. Mitigation of enteric methane emissions through pasture management in integrated crop-livestock systems: Trade-offs between animal performance and environmental impacts. J. Clean Prod. 2019, 213, 968–975. [Google Scholar] [CrossRef]
- Semmartin, M.; Garibaldi, L.A.; Chaneton, E.J. Grazing history effects on above- and below-ground litter decomposition and nutrient cycling in two co-occurring grasses. Plant. Soil 2008, 303, 177–189. [Google Scholar] [CrossRef]
- Shand, C.A.; Coutts, G. The effects of sheep faeces on soil solution composition. Plant. Soil 2006, 285, 135–148. [Google Scholar] [CrossRef]
- Briske, D.D.; Derner, J.D.; Brown, J.R.; Fuhlendorf, S.D.; Teague, W.R.; Havstad, K.M.; Gillen, R.L.; Ash, A.J.; Willms, W.D. Rotational grazing on rangelands: Reconciliation of perception and experimental evidence. Rangel. Ecol. Manag. 2008, 61, 3–17. [Google Scholar] [CrossRef]
- Cherney, D.J.R.; Cherney, J.H.; Lucey, R.F. In vitro digestion kinetics and quality of perennial grasses as influenced by forage maturity. J. Dairy Sci. 1993, 76, 790–797. [Google Scholar] [CrossRef]
- Van Soest, P.J. Nutritional Ecology of the Ruminant, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 1994. [Google Scholar]
- Haynes, R.J.; Williams, P.H. Nutrient cycling and soil fertility in the grazed pasture ecosystem. Adv. Agron. 1993, 49, 119–199. [Google Scholar] [CrossRef]
- Deiss, L.; Moraes, A.; Dieckow, J.; Franzluebbers, A.J.; Gatiboni, L.C.; Sassaki, L.G.; Carvalho, P.C.F. Soil phosphorus compounds in integrated crop-livestock systems of subtropical Brazil. Geoderma 2016, 274, 88–96. [Google Scholar] [CrossRef]
- Braz, S.P.; Junior, D.N.; Cantarutti, R.B.; Regazzi, A.J.; Martins, C.E.; Fonseca, D.M. Disponibilização dos nutrientes das fezes de bovinos em pastejo para a Forragem. R. Bras. Zootec. 2002, 31, 1614–1623. [Google Scholar] [CrossRef][Green Version]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Gonçalves, J.L.M.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef]
- FAO. World Reference Base for Soil Resources 2014, International Soil Classification System for Naming Soils and Creating Legends for Soil Maps. 2015. Available online: http://www.fao.org/3/i3794en/I3794en.pdf (accessed on 15 May 2020).
- CFS-RS/SC–Comissão de Fertilidade Do Solo-RS/SC. Recomendações de Adubação e de Calagem Para os Estados do Rio Grande do Sul e de Santa Catarina, 3rd ed.; SBCS-Núcleo Regional Sul/EMBRAPA-CNPT: Passo Fundo, Brazil, 1995; pp. 1–224. [Google Scholar]
- NRC−National Research Council. Nutrient Requirements of Sheep; National Academic of Science: Washington, DC, USA, 1985. [Google Scholar]
- Barthram, G.T. Experimental Techniques: The HFRO Sward Stick; The Hill Farming Research Organization/Biennial Report; HFRO: Penicuik, Scotland, 1985; pp. 29–30. [Google Scholar]
- Mott, G.O.; Lucas, H.L. The design, conduct, and interpretation of grazing trials on cultivated and improved pastures. In Proceedings of the 6th International Grassland Congress; Wagner, R.E., Myers, W.M., Gaines, S.H., Lucas, H.L., Eds.; State College Press: Philadelphia, PA, USA, 1952; pp. 1380–1385. [Google Scholar]
- Wieder, R.K.; Lang, G.E. A critique of the analytical methods used in examining decomposition data obtained from litter bags. Ecology 1982, 63, 1636–1642. [Google Scholar] [CrossRef]
- Tedesco, M.J.; Gianello, C.; Bissani, C.A.; Bohnen, H.; Volkweiss, S.J. Análise de Solo, Plantas e Outros Materiais; Departamento de Solos/UFRGS: Porto Alegre, UK, 1995. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B. Analysis of Forages and Fibrous Foods—A Laboratory Manual for Animal Science; Cornell University Press: Ithaca, NY, USA, 1985. [Google Scholar]
- Assmann, J.M.; Martins, A.P.; Anghinoni, I.; Denardin, L.G.O.; Nichel, G.H.; Costa, S.E.V.G.A.; Silva, R.A.P.; Balerini, F.; Carvalho, P.C.F.; Franzluebbers, A.J. Phosphorus and potassium cycling in a long-term no-till integrated soybean-beef cattle production system under different grazing intensities in subtropics. Nutr. Cycl. Agroecosys 2017, 108, 21–33. [Google Scholar] [CrossRef]
- Plante, A.F.; Parton, W.J. The dynamics of soil organic matter and nutrient cycling. In Soil Microbiology, Ecology and Biochemistry; Paul, E.A., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 433–467. [Google Scholar]
- Savian, J.V.; Neto, A.B.; de David, D.B.; Bremm, C.; Schons, R.M.T.; Genro, T.C.M.; do Amaral, G.A.; Gere, J.; McManus, C.M.; Bayer, C.; et al. Grazing intensity and stocking methods on animal production and methane emission by grazing sheep: Implications for integrated crop-livestock system. Agric. Ecosyst. Environ. 2014, 190, 112–119. [Google Scholar] [CrossRef]
- Rook, A.J.; Dumont, B.; Isselstein, J.; Osoro, K.; WallisDeVries, M.F.; Parente, G.; Mills, J. Matching type of livestock to desired biodiversity outcomes in pastures—A review. Biol. Cons. 2014, 119, 137–150. [Google Scholar] [CrossRef]
- Mowat, D.N.; Fulkerson, R.S.; Tossell, W.E.; Winch, J.E. The in vitro digestibility and protein content of leaf and stem proportions of forages. Can. J. Plant. Sci. 1965, 45, 321–331. [Google Scholar] [CrossRef]
- Doreau, M.; Diawara, A. Effect of level of intake on digestion in cows: Influence of animal genotype and nature of hay. Livest. Prod. Sci. 2003, 81, 35–45. [Google Scholar] [CrossRef]
- Freitas, M.; Araujo, C.A.S.; Silva, D.J. Decomposição e liberação de nutrientes de estercos em função da profundidade e do tempo de incorporação. Rev. Semiárido Visu. 2012, 2, 150–161. [Google Scholar]
- Bahamonde, H.A.; Gargaglione, V.; Peri, P.L. Sheep faeces decomposition and nutrient release across an environmental gradient in Southern Patagonia. Ecol. Austral. 2017, 27, 18–28. [Google Scholar]
- CQFS-RS/SC. Manual de Calagem e Adubação Para os Estados do Rio Grande do Sul e de Santa Catarina, 11th ed.; Sociedade Brasileira de Ciência do Solo–Núcleo Regional Sul.: Viçosa, Brazil, 2016; pp. 1–376. [Google Scholar]
- Weeda, W.C. Effect of cattle dung patches on soil tests and botanical and chemical composition of herbage. Nz. J. Agric. Res. 1977, 20, 471–478. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Pasture Development Stage | No. Animals Per ha | Dung Excreted per Animal (1) | Total Dung Return | Dung Nutrient Content | Total Nutrient Return | ||
|---|---|---|---|---|---|---|---|---|
| P | K | P | K | |||||
| g day−1 | kg ha−1 day−1 | g kg−1 | g ha−1 day−1 | |||||
| 2015 | Vegetative | 28 A | 290 ns | 8.1 A | 11.7 a | 24.8 a | 95.0 Aa | 201.4 Aa |
| Post-flowering | 28 A | 290 | 8.1 A | 10.6 b | 21.6 b | 86.1 Ab | 175.4 Ab | |
| 2016 | Vegetative | 19 B | 290 | 5.5 B | 16.4 a | 25.7 a | 90.4 Ba | 141.6 Ba |
| Post-flowering | 19 B | 290 | 5.5 B | 12.9 b | 25.5 b | 71.1 Bb | 140.5 Bb | |
| Year | Pasture Development Stage | Cellulose | Hemicellulose | Lignin |
|---|---|---|---|---|
| g kg−1 | ||||
| 2015 | Vegetative | 192 a | 139 a | 149 ns |
| Post-flowering | 203 a | 153 a | 141 | |
| 2016 | Vegetative | 180 b | 134 b | 105 |
| Post-flowering | 227 a | 179 a | 112 | |
| Year | Dung Decomposition Phase | Comp. A (1) | k1 (2) | k2 (3) | t1/2 | R2 | |
|---|---|---|---|---|---|---|---|
| A (4) | (100-A) (5) | ||||||
| % | days−1 | days | |||||
| 2015 | Pasture | 22 a | 0.0442 a | 0.0016 a | 16 | 433 | 0.99 |
| Crop | 36 a | 0.0128 b | 0.0015 a | 54 | 462 | 0.99 | |
| 2016 | Pasture | 21 a | 0.0413 a | 0.0012 a | 17 | 594 | 0.99 |
| Crop | 13 b | 0.0623 a | 0.0018 a | 11 | 385 | 0.99 | |
| Year | Dung Decomposition Phase | Comp. A (1) | k1 (2) | k2 (3) | t1/2 | R2 | |
|---|---|---|---|---|---|---|---|
| A (4) | (100-A) (5) | ||||||
| % | days−1 | days | |||||
| 2015 | Pasture | 28 a | 0.0738 a | 0.0022 a | 9 | 319 | 0.94 |
| Crop | 33 a | 0.0298 a | 0.0013 a | 23 | 554 | 0.95 | |
| 2016 | Pasture | 57 a | 0.0448 a | 0.0027 b | 15 | 260 | 0.97 |
| Crop | 28 b | 0.1027 a | 0.0066 a | 7 | 105 | 0.95 | |
| Year | Dung Decomposition Phase | Comp. A (1) | k1 (2) | t1/2 | R2 |
|---|---|---|---|---|---|
| % | days−1 | days | |||
| 2015 | Pasture | 99.4 a | 0.0943 a | 7 | 0.95 |
| Crop | 97.7 a | 0.0163 b | 43 | 0.92 | |
| 2016 | Pasture | 100.0 a | 0.0922 a | 8 | 0.97 |
| Crop | 99.3 a | 0.0740 a | 9 | 0.97 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arnuti, F.; Denardin, L.G.d.O.; Nunes, P.A.d.A.; Alves, L.A.; Cecagno, D.; de Assis, J.; Schaidhauer, W.d.S.; Anghinoni, I.; Chabbi, A.; César de F. Carvalho, P. Sheep Dung Composition and Phosphorus and Potassium Release Affected by Grazing Intensity and Pasture Development Stage in an Integrated Crop-Livestock System. Agronomy 2020, 10, 1162. https://doi.org/10.3390/agronomy10081162
Arnuti F, Denardin LGdO, Nunes PAdA, Alves LA, Cecagno D, de Assis J, Schaidhauer WdS, Anghinoni I, Chabbi A, César de F. Carvalho P. Sheep Dung Composition and Phosphorus and Potassium Release Affected by Grazing Intensity and Pasture Development Stage in an Integrated Crop-Livestock System. Agronomy. 2020; 10(8):1162. https://doi.org/10.3390/agronomy10081162
Chicago/Turabian StyleArnuti, Fernando, Luiz Gustavo de O. Denardin, Pedro Arthur de A. Nunes, Lucas A. Alves, Diego Cecagno, Júlia de Assis, Walker da S. Schaidhauer, Ibanor Anghinoni, Abad Chabbi, and Paulo César de F. Carvalho. 2020. "Sheep Dung Composition and Phosphorus and Potassium Release Affected by Grazing Intensity and Pasture Development Stage in an Integrated Crop-Livestock System" Agronomy 10, no. 8: 1162. https://doi.org/10.3390/agronomy10081162
APA StyleArnuti, F., Denardin, L. G. d. O., Nunes, P. A. d. A., Alves, L. A., Cecagno, D., de Assis, J., Schaidhauer, W. d. S., Anghinoni, I., Chabbi, A., & César de F. Carvalho, P. (2020). Sheep Dung Composition and Phosphorus and Potassium Release Affected by Grazing Intensity and Pasture Development Stage in an Integrated Crop-Livestock System. Agronomy, 10(8), 1162. https://doi.org/10.3390/agronomy10081162