Release of Medicago truncatula Gaertn. and Pisum sativum subsp. elatius (M. Bieb.) Asch. et Graebn. Seed Dormancy Tested in Soil Conditions

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Soil Deposition Experiment
2.3. Meteorological and Environmental Data Acquisition
2.4. Data Analysis
2.4.1. PY Release of Barrel Medic and Wild Pea Accessions in Natural Soil Condition
2.4.2. Association between PY Release under Laboratory Condition and Soil Burial Environment
2.4.3. Relationship between Environment of Origin and PY Release under in Situ (Soil) Condition
3. Results
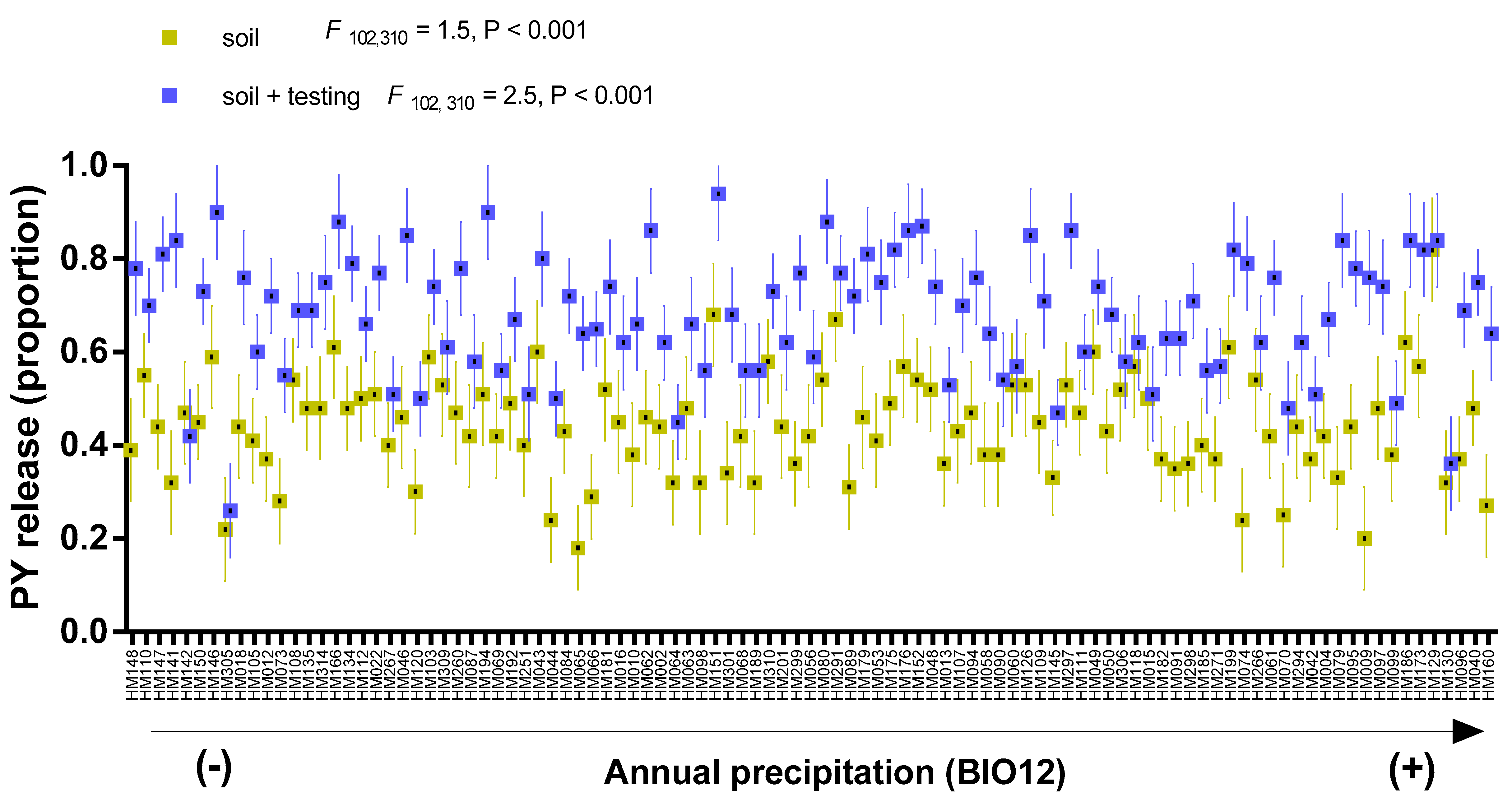
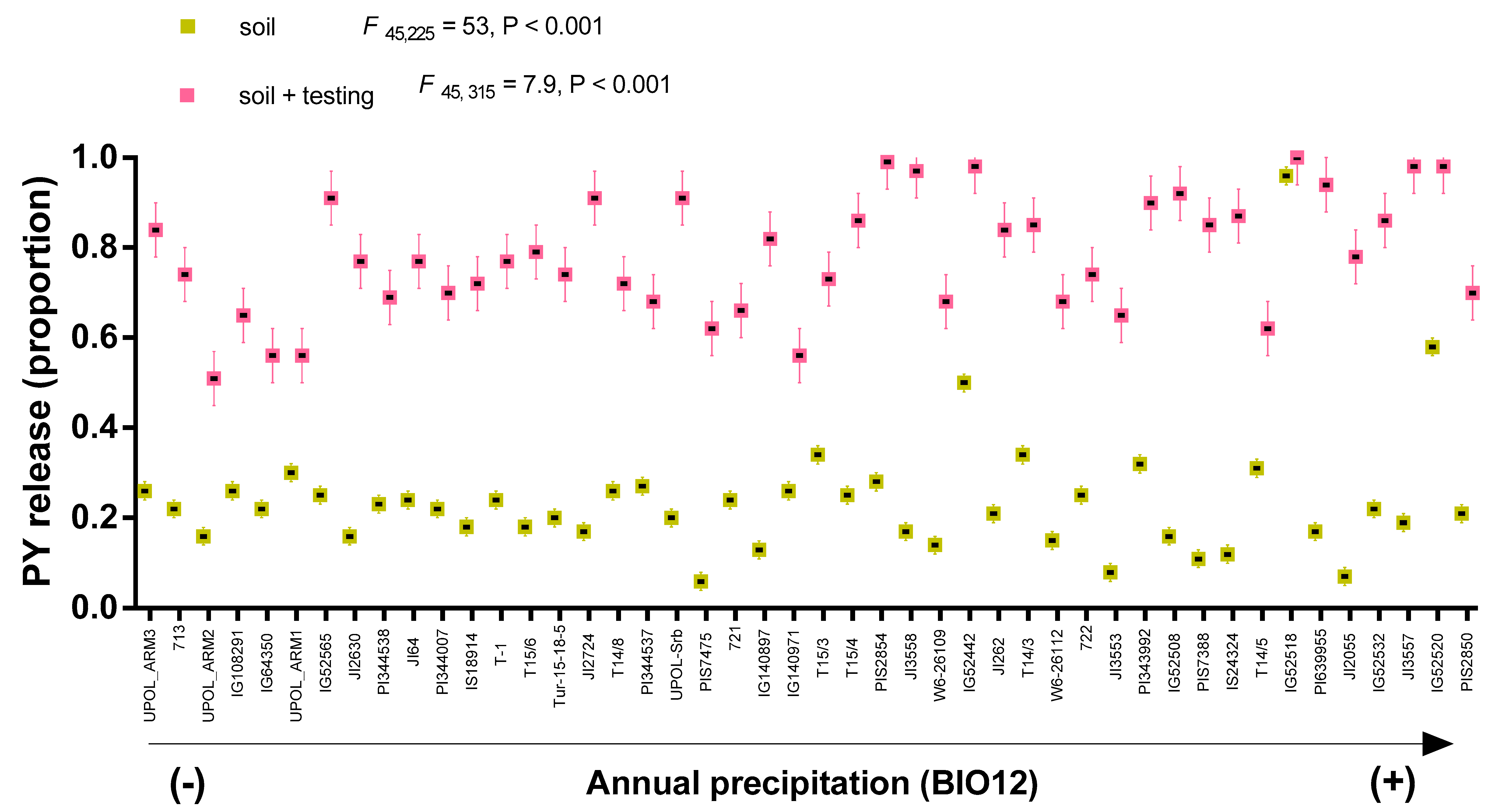
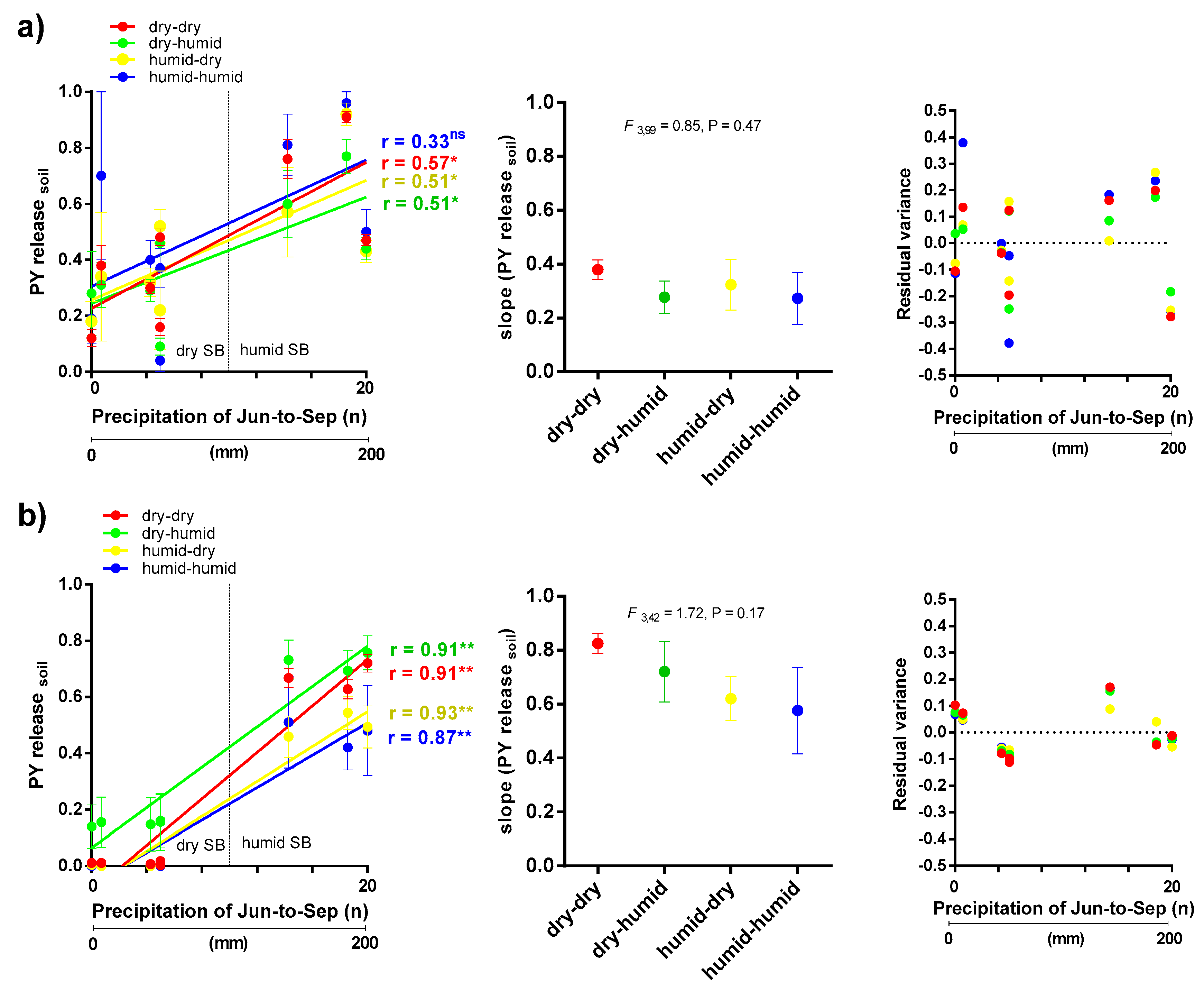
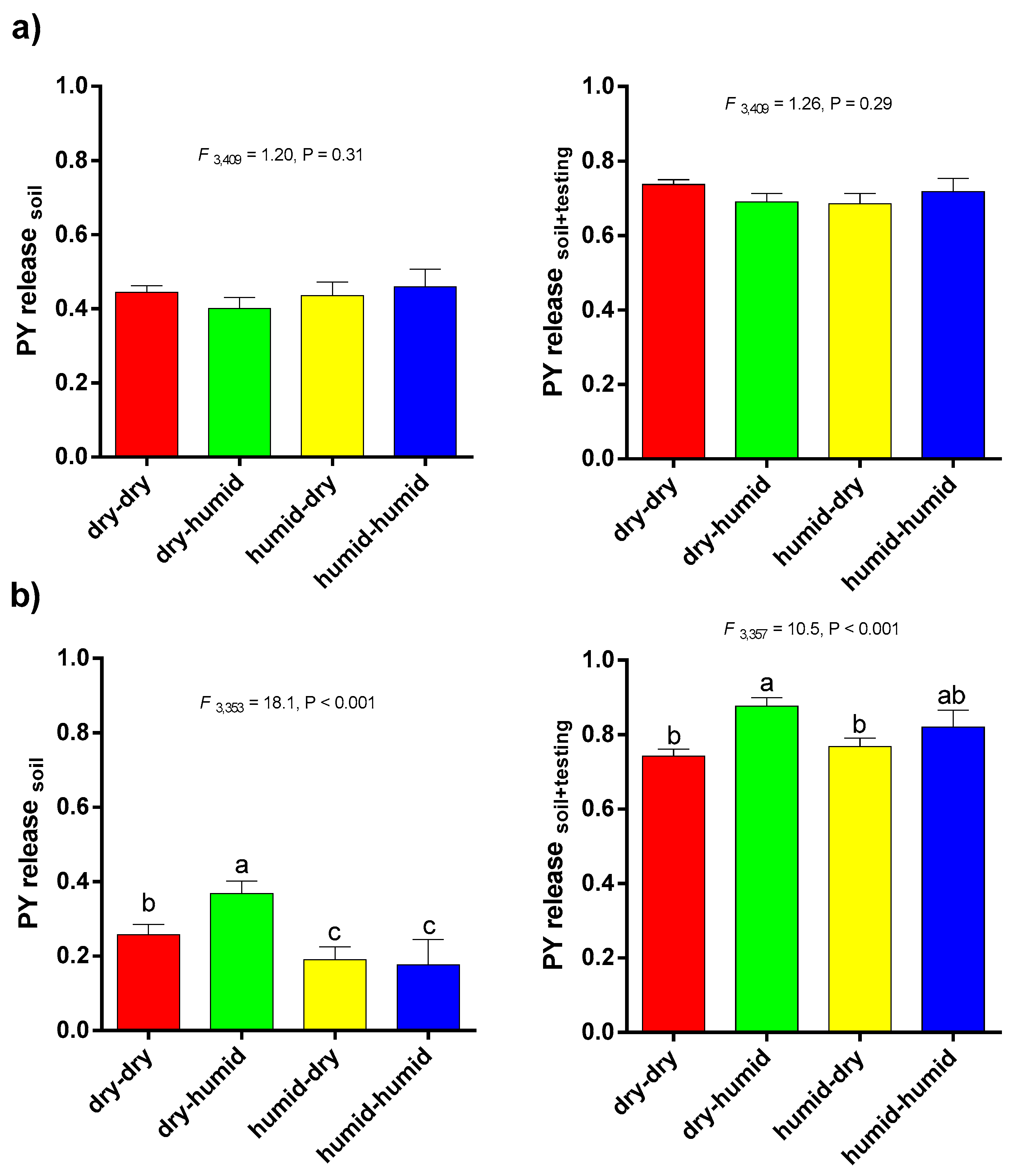
3.1. PY Release of Barrel Medic and Wild Pea Accessions in Natural Soil Condition
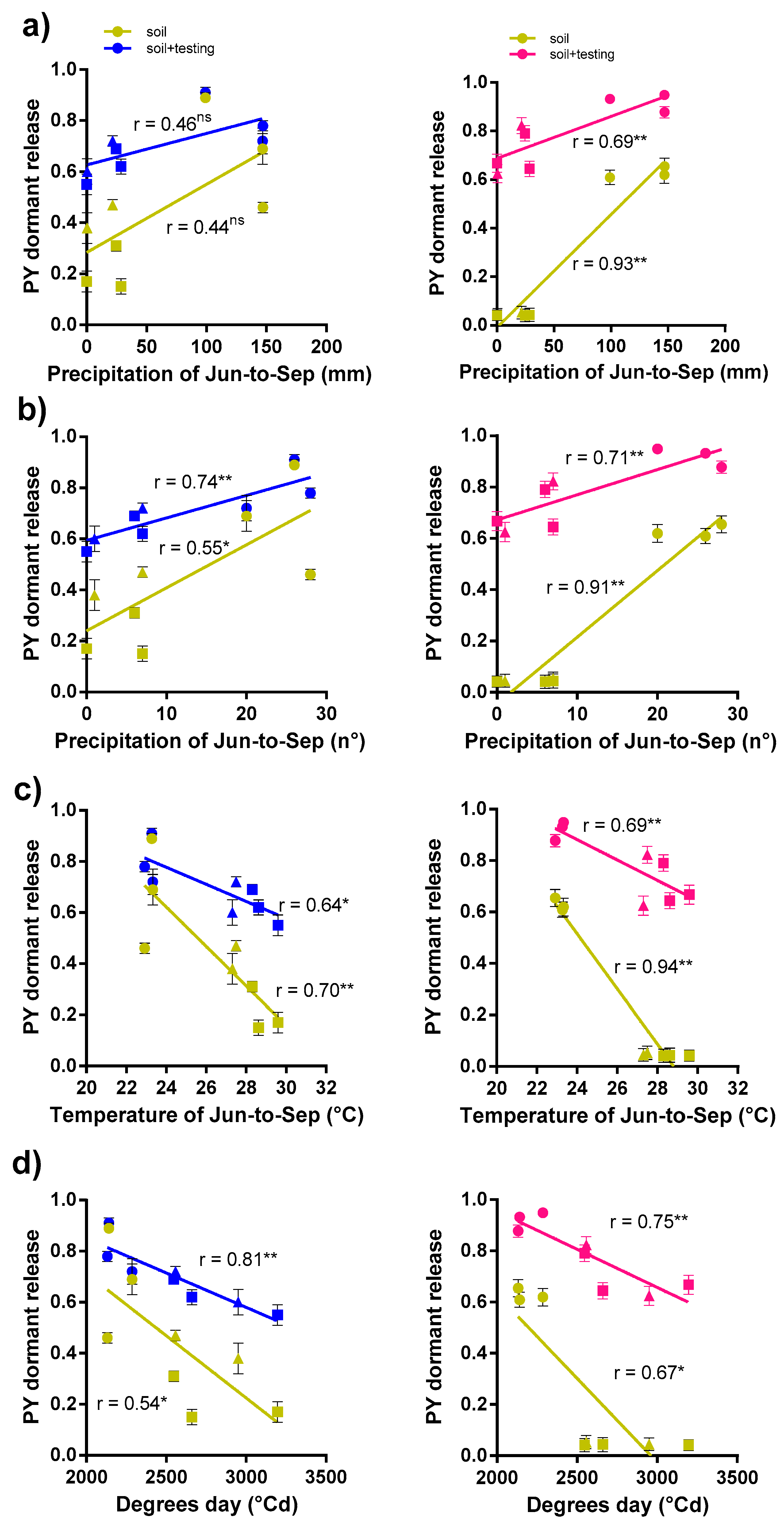
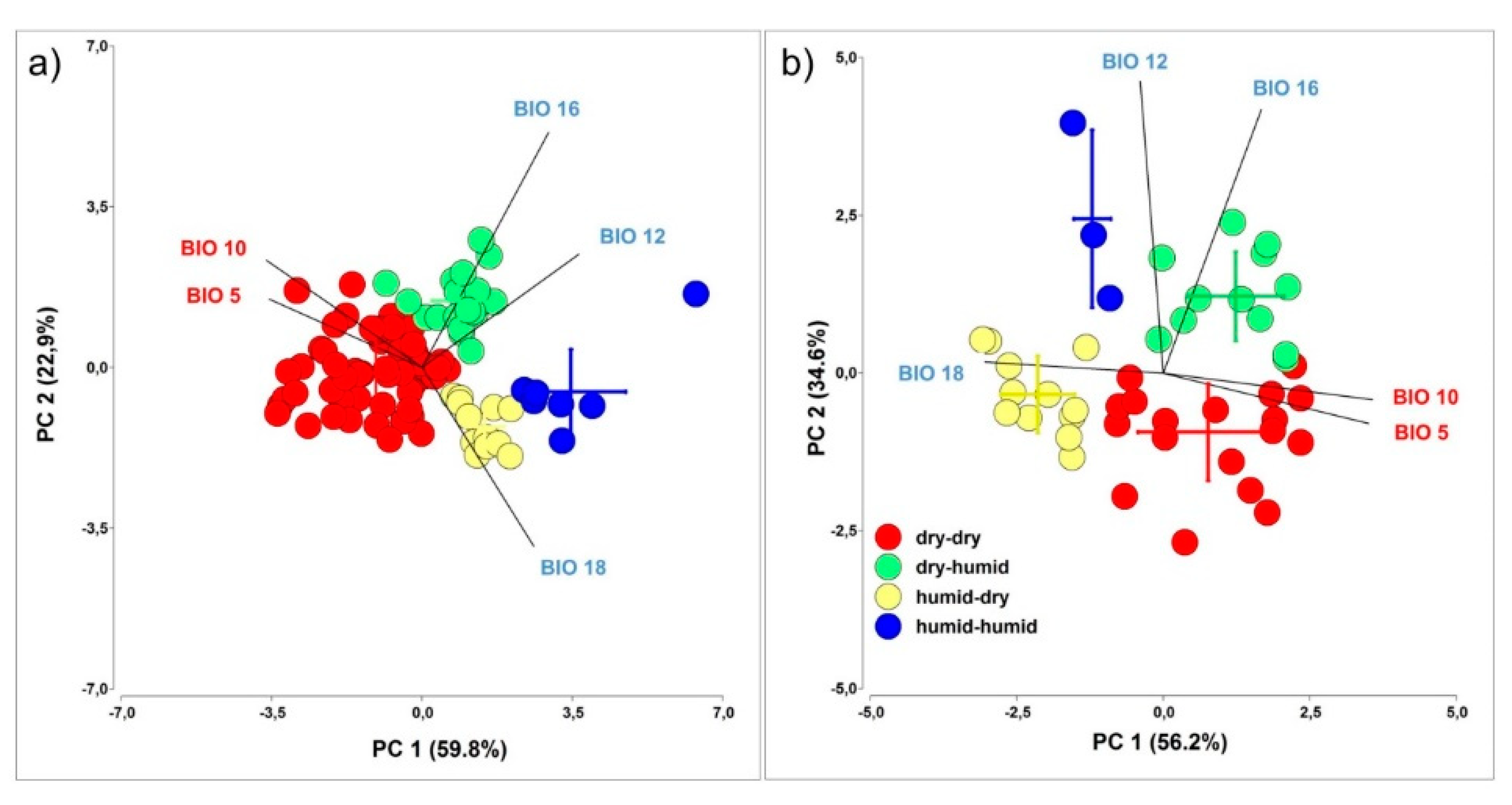
3.2. Correlations between PY Release under Laboratory Condition and Soil Burial Environment
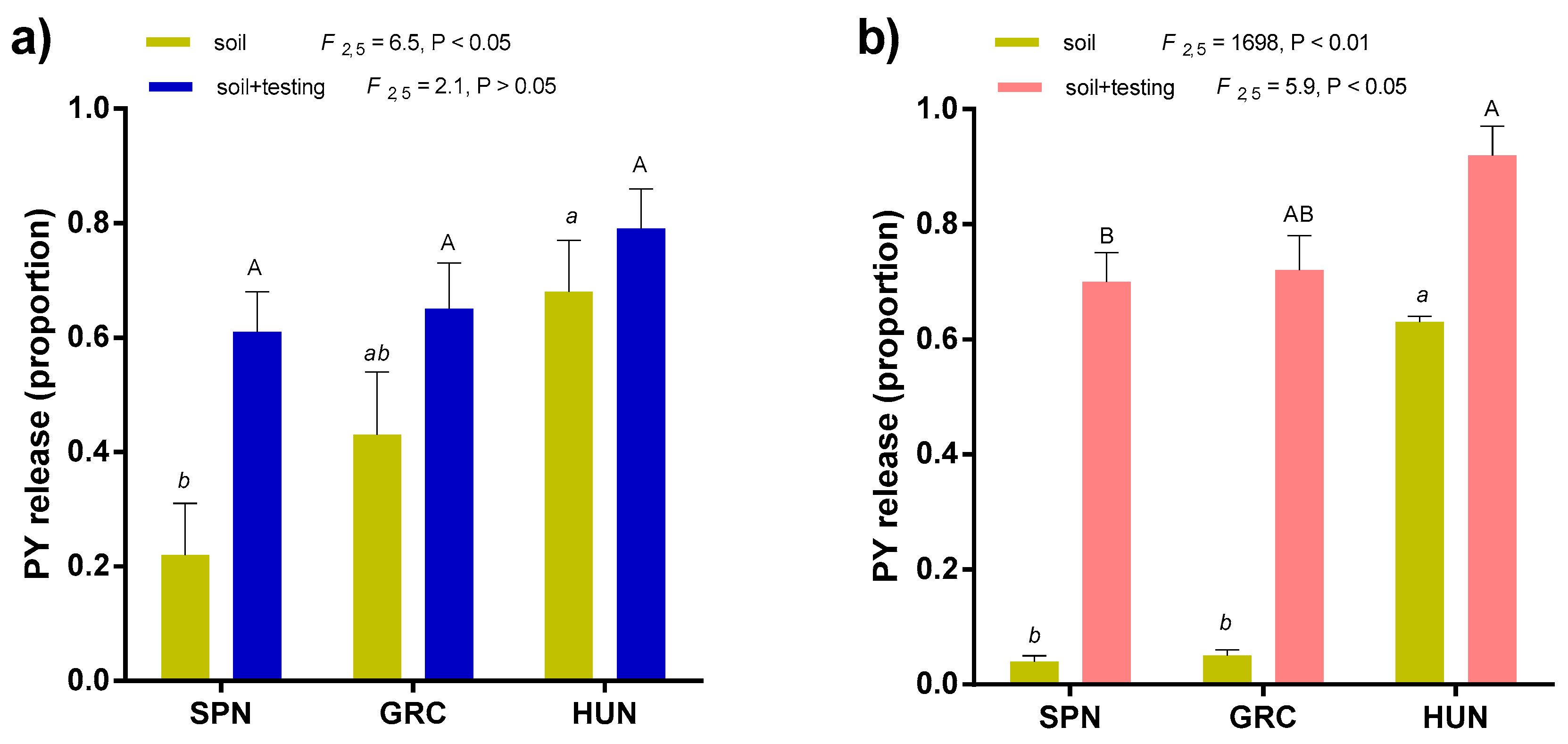
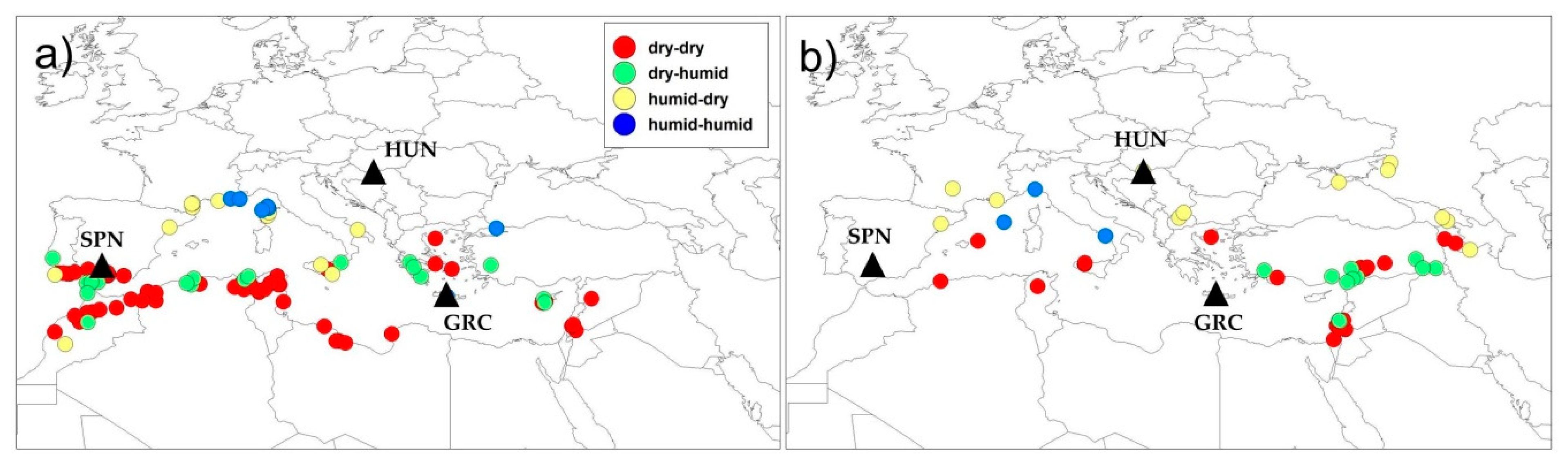
3.3. Relationship between Environment of Origin and PY Dormancy Release Under Soil Conditions
4. Discussion
4.1. Association between PY Release under Laboratory Conditions and Soil Burial Environment
4.2. Plasticity and Bet-Hedging Strategy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Larson, J.E.; Anacker, B.L.; Wanous, S.; Funk, J.L. Ecological strategies begin at germination: Traits, plasticity, and survival in the first four days of plant life. Funct. Ecol. 2020. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. Seeds: Ecology, Biogeography and Evolution of Dormancy and Germination, 2nd ed.; Elsevier, Academic Press: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Finch-Savage, W.E.; Footitt, S. Seed dormancy cycling and the regulation of dormancy mechanisms to time germination in variable field environments. J. Exp. Bot. 2017, 68, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Bentsink, L.; Jowett, J.; Hanhart, C.J.; Koornneef, M. Cloning of DOG1, a quantitative trait locus controlling seed dormancy in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 17042–17047. [Google Scholar] [CrossRef] [PubMed]
- Vidigal, D.S.; Marques, A.C.; Willems, L.A.; Buijs, G.; Méndez-Vigo, B.; Hilhorst, H.W.; Bentsink, L.; Picó, F.X.; Alonso-Blanco, C. Altitudinal and climatic associations of seed dormancy and flowering traits evidence adaptation of annual life cycle timing in Arabidopsis thaliana. Plant Cell Environ. 2016, 39, 1737–1748. [Google Scholar] [CrossRef]
- Smýkal, P.; Vernoud, V.; Blair, M.W.; Soukup, A.; Thompson, R.D. The role of the testa during development and in establishment of dormancy of the legume seed. Front. Plant Sci. 2014, 5, 351. [Google Scholar]
- Willis, C.G.; Baskin, C.C.; Baskin, J.M.; Auld, J.R.; Venable, D.L.; Cavender-Bares, J.; Donohue, K.; de Casas, R.R.; NESCent Germination Working Group. The evolution of seed dormancy: Environmental cues, evolutionary hubs, and diversification of the seed plants. New Phytol. 2014, 203, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Simons, A.M. Playing smart vs. playing safe: The joint expression of phenotypic plasticity and potential bet hedging across and within thermal environments. J. Evol. Biol. 2014, 27, 1047–1056. [Google Scholar] [CrossRef] [PubMed]
- Sultan, S.E. Phenotypic plasticity for plant development, function and life history. Trends Plant Sci. 2000, 5, 537–542. [Google Scholar] [CrossRef]
- Pigliucci, M. Evolution of phenotypic plasticity: Where are we going now? Trends Ecol. Evol. 2005, 20, 481–486. [Google Scholar] [CrossRef]
- Cohen, D. Optimizing reproduction in a randomly varying environment. J. Theor. Biol. 1966, 352, 119–129. [Google Scholar] [CrossRef]
- Kelly, M. Adaptation to climate change through genetic accommodation and assimilation of plastic phenotypes. Philos. Trans. R. Soc. B 2019, 374, 20180176. [Google Scholar] [CrossRef]
- Nicotra, A.B.; Atkin, O.K.; Bonser, S.P.; Davidson, A.M.; Finnegan, J.; Mathesius, U.; Poot, P.; Purugganan, M.D.; Valladares, F.; van Kleunen, M. Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Via, S.; Lande, R. Genotype-environment interaction and the evolution of phenotypic plasticity. Evolution 1985, 39, 505–522. [Google Scholar] [CrossRef] [PubMed]
- Slatkin, M. Hedging one’s evolutionary bets. Nature 1974, 250, 704–705. [Google Scholar] [CrossRef]
- Starrfelt, J.; Kokko, H. Bet-hedging—A triple trade-off between means, variances and correlations. Biol. Rev. 2012, 87, 742–755. [Google Scholar] [CrossRef]
- Donaldson-Matasci, M.C.; Bergstrom, C.T.; Lachmann, M. When unreliable cues are good enough. Am. Nat. 2013, 182, 313–327. [Google Scholar] [CrossRef]
- Donohue, K. Niche construction through phenological plasticity: Life history dynamics and ecological consequences. New Phytol. 2005, 166, 83–92. [Google Scholar] [CrossRef]
- Van Assche, J.A.; Debucquoy, K.L.A.; Rommens, W.A.F. Seasonal cycles in the germination capacity of buried seeds of some Leguminosae (Fabaceae). New Phytol. 2003, 158, 315–323. [Google Scholar] [CrossRef]
- Quinlivan, B.J. The effect of constant and fluctuating temperatures on the permeability of the hard seeds of some legume species. Aust. J. Agric. Res. 1961, 12, 1009–1022. [Google Scholar] [CrossRef]
- Quinlivan, B.J. The relationship between temperature fluctuations and the softening of hard seeds of some legume species. Aust. J. Agric. Res. 1966, 17, 625–631. [Google Scholar] [CrossRef]
- Quinlivan, B.J. Environmental variation in the long term pattern of germination from hard seeds of Lupinus varius. Aust. J. Exp. Agric. Anim. Husb. 1967, 7, 263–265. [Google Scholar] [CrossRef]
- Quinlivan, B.J.; Millington, A.J. The effect of a Mediterranean summer environment on the permeability of hard seeds of subterranean clover. Aust. J. Agric. Res. 1962, 13, 377–387. [Google Scholar] [CrossRef]
- Taylor, G.B. Hardseededness in Mediterranean annual pasture legumes in Australia: A review. Aust. J. Agric. Res. 2005, 56, 645–661. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy; Springer: New York, NY, USA, 2013. [Google Scholar]
- Probert, R. The role of temperature in the regulation of seed dormancy and germination. In Seeds: The Ecology of Regeneration in Plant Communities; Fenner, M., Ed.; CAB International: Wallingford, UK, 2000; pp. 261–292. [Google Scholar]
- de Casas, R.R.; Willis, C.G.; Pearse, W.D.; Baskin, C.C.; Baskin, J.M.; Cavender-Bares, J. Global biogeography of seed dormancy is determined by seasonality and seed size: A case study in the legumes. New Phytol. 2017, 214, 1527–1536. [Google Scholar] [CrossRef] [PubMed]
- Donohue, K.; de Casas, R.R.; Burghardt, L.; Kovach, K.; Willis, C.G. Germination, post-germination adaptation, and species ecological ranges. Ann. Rev. Ecol. Evol. Syst. 2010, 41, 293–319. [Google Scholar] [CrossRef]
- Thompson, J.D. Plant Evolution in the Mediterranean; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Hradilová, I.; Duchoslav, M.; Brus, J.; Pechanec, V.; Hýbl, M.; Kopecký, P.; Smržová, L.; Štefelová, N.; Vaclávek, T.; Bariotakis, M.; et al. Variation in wild pea (Pisum sativum subsp. elatius) seed dormancy and its relationship to the environment and seed coat traits. PeerJ 2019, 7, e6263. [Google Scholar]
- Smýkal, P.; Hradilová, I.; Trněný, O.; Brus, J.; Rathore, A.; Bariotakis, M.; Das, R.R.; Bhattacharyya, D.; Richards, C.; Coyne, C.J.; et al. Genomic diversity and macroecology of the crop wild relatives of domesticated pea. Sci. Rep. 2017, 7, 17384. [Google Scholar] [CrossRef]
- Smýkal, P.; Trněný, O.; Brus, J.; Hanáček, P.; Rathore, A.; Roma, R.D.; Pechanec, V.; Duchoslav, M.; Bhattacharyya, D.; Bariotakis, M.; et al. Genetic structure of wild pea (Pisum sativum subsp. elatius) populations in the northern part of the Fertile Crescent reflects moderate cross-pollination and strong effect of geographic but not environmental distance. PLoS ONE 2018, 13, e0194056. [Google Scholar]
- Renzi, J.P.; Duchoslav, M.; Brus, J.; Hradilová, I.; Pechanec, V.; Václavek, T.; Machalová, J.; Hron, K.; Verdier, J.; Smýkal, P. Physical dormancy release in Medicago truncatula seeds is related to environmental variations. Plants 2020, 9, 503. [Google Scholar] [CrossRef] [PubMed]
- Cochrane, A. Multi-year sampling provides insight into the bet-hedging capacity of the soil-stored seed reserve of a threatened Acacia species from Western Australia. Plant Ecol. 2019, 220, 241–253. [Google Scholar] [CrossRef]
- Hudson, A.R.; Ayre, D.J.; Ooi, M.K.J. Physical dormancy in a changing climate. Seed Sci. Res. 2015, 25, 66–81. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Di Rienzo, J.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C. Infostat Versión 2013; Grupo InfoStat, FCA, Universidad Nacional de Córdoba: Córdoba, Argentina, 2013. [Google Scholar]
- Valladares, F.; Sanchez-Gomez, D.; Zavala, M.A. Quantitative estimation of phenotypic plasticity: Bridging the gap between the evolutionary concept and its ecological applications. J. Ecol. 2006, 94, 1103–1116. [Google Scholar] [CrossRef]
- Liu, K.; Baskin, J.M.; Baskin, C.C.; Bu, H.; Liu, M.; Liu, W.; Du, G. Effect of storage conditions on germination of seeds of 489 species from high elevation grasslands of the eastern Tibet Plateau and some implications for climate change. Am. J. Bot. 2011, 98, 12–19. [Google Scholar] [CrossRef]
- Hu, X.W.; Wu, Y.P.; Wang, Y.R. Different requirements for physical dormancy release in two populations of Sophora alopecuroides relation to burial depth. Ecol. Res. 2009, 24, 1051–1056. [Google Scholar] [CrossRef]
- Long, R.L.; Gorecki, M.J.; Renton, M.; Scott, J.K.; Colville, L.; Goggin, D.E.; Commander, L.E.; Westcott, D.A.; Cherry, H.; Finch-Savage, W.E. The ecophysiology of seed persistence: A mechanistic view of the journey to germination or demise. Biol. Rev. Camb. Philos. Soc. 2015, 90, 31–59. [Google Scholar] [CrossRef]
- Trněný, O.; Brus, J.; Hradilová, I.; Rathore, A.; Das, R.R.; Kopecký, P.; Coyne, C.J.; Reeves, P.; Richards, C.; Smýkal, P. Molecular evidence for two domestication events in the pea crop. Genes 2018, 9, 535. [Google Scholar] [CrossRef]
- Norman, H.C.; Cocks, P.C.; Galwey, N.W. Hardseededness in annual clovers: Variation between populations from wet and dry environments. Aust. J. Agric. Sci. 2002, 53, 821–829. [Google Scholar] [CrossRef]
- Ferreras, A.E.; Marcora, P.I.; Venier, M.P.; Funes, G. Different strategies for breaking physical seed dormancy in field conditions in two fruit morphs of Vachellia caven (Fabaceae). Seed Sci. Res. 2018, 28, 8–15. [Google Scholar] [CrossRef]
- Guerrero, J.; Andrello, M.; Burgarella, C.; Manel, S. Soil environment is a key driver of adaptation in Medicago truncatula: New insights from landscape genomics. New Phytol. 2018, 219, 378–390. [Google Scholar] [CrossRef]
- Friedman, J.; Middleton, T.E.; Rubin, M.J. Environmental heterogeneity generates intrapopulation variation in life-history traits in an annual plant. New Phytol. 2019, 224, 1171–1183. [Google Scholar] [CrossRef] [PubMed]
- Matesanz, S.; Ramos-Muñoz, M.; Moncalvillo, B.; Rubio Teso, M.L.; de Dionisio, S.L.G.; Romero, J.; Iriondo, J.M. Plasticity to drought and ecotypic differentiation in populations of a crop wild relative. AoB Plants 2020. [Google Scholar] [CrossRef] [PubMed]
- Yoder, J.B.; Stanton-Geddes, J.; Zhou, P.; Briskine, R.; Young, N.D.; Tiffin, P. Genomic signature of adaptation to climate in Medicago truncatula. Genetics 2014, 196, 1263–1275. [Google Scholar] [CrossRef] [PubMed]
- Burgarella, C.; Chantret, N.; Gay, L.; Prosperi, J.-M.; Bonhomme, M.; Tiffin, P.; Young, N.D.; Ronfort, J. Adaptation to climate through flowering phenology: A case study in Medicago truncatula. Mol. Ecol. 2016, 25, 3397–3415. [Google Scholar] [CrossRef] [PubMed]
- ten Brink, H.; Gremer, J.R.; Kokko, H. Optimal germination timing in unpredictable environments: The importance of dormancy for both among- and within-season variation. Ecol. Lett. 2020, 23, 620–630. [Google Scholar] [CrossRef] [PubMed]
- Liyanage, G.S.; Ooi, M.K.J. Intra-population level variation in thresholds for physical dormancy-breaking temperature. Ann. Bot. 2015, 116, 123–131. [Google Scholar] [CrossRef]
- Burghardt, L.T.; Metcalf, C.J.E.; Donohue, K. A cline in seed dormancy helps conserve the environment experienced during reproduction across the range of Arabidopsis thaliana. Am. J. Bot. 2016, 103, 47–59. [Google Scholar] [CrossRef]
- Bonser, S. High reproductive efficiency as an adaptive strategy in competitive environments. Funct. Ecol. 2013, 27, 876–885. [Google Scholar] [CrossRef]
- Duncan, C.; Schultz, N.L.; Good, M.K.; Lewandrowski, W.; Cook, S. The risk-takers and -avoiders: Germination sensitivity to water stress in an arid zone with unpredictable rainfall. AoB Plants 2019. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Renzi, J.P.; Brus, J.; Pirintsos, S.; Erdős, L.; Duchoslav, M.; Smýkal, P. Release of Medicago truncatula Gaertn. and Pisum sativum subsp. elatius (M. Bieb.) Asch. et Graebn. Seed Dormancy Tested in Soil Conditions. Agronomy 2020, 10, 1026. https://doi.org/10.3390/agronomy10071026
Renzi JP, Brus J, Pirintsos S, Erdős L, Duchoslav M, Smýkal P. Release of Medicago truncatula Gaertn. and Pisum sativum subsp. elatius (M. Bieb.) Asch. et Graebn. Seed Dormancy Tested in Soil Conditions. Agronomy. 2020; 10(7):1026. https://doi.org/10.3390/agronomy10071026
Chicago/Turabian StyleRenzi, Juan Pablo, Jan Brus, Stergios Pirintsos, László Erdős, Martin Duchoslav, and Petr Smýkal. 2020. "Release of Medicago truncatula Gaertn. and Pisum sativum subsp. elatius (M. Bieb.) Asch. et Graebn. Seed Dormancy Tested in Soil Conditions" Agronomy 10, no. 7: 1026. https://doi.org/10.3390/agronomy10071026
APA StyleRenzi, J. P., Brus, J., Pirintsos, S., Erdős, L., Duchoslav, M., & Smýkal, P. (2020). Release of Medicago truncatula Gaertn. and Pisum sativum subsp. elatius (M. Bieb.) Asch. et Graebn. Seed Dormancy Tested in Soil Conditions. Agronomy, 10(7), 1026. https://doi.org/10.3390/agronomy10071026