Optimization of Light Interception, Leaf Area and Yield in “WA38”: Comparisons among Training Systems, Rootstocks and Pruning Techniques




Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Leaf Area Estimation—Image Acquisition
2.3. Leaf Area Estimation—Image Processing
2.4. Light Interception
2.5. Production
2.6. Statistical Analyses
3. Results
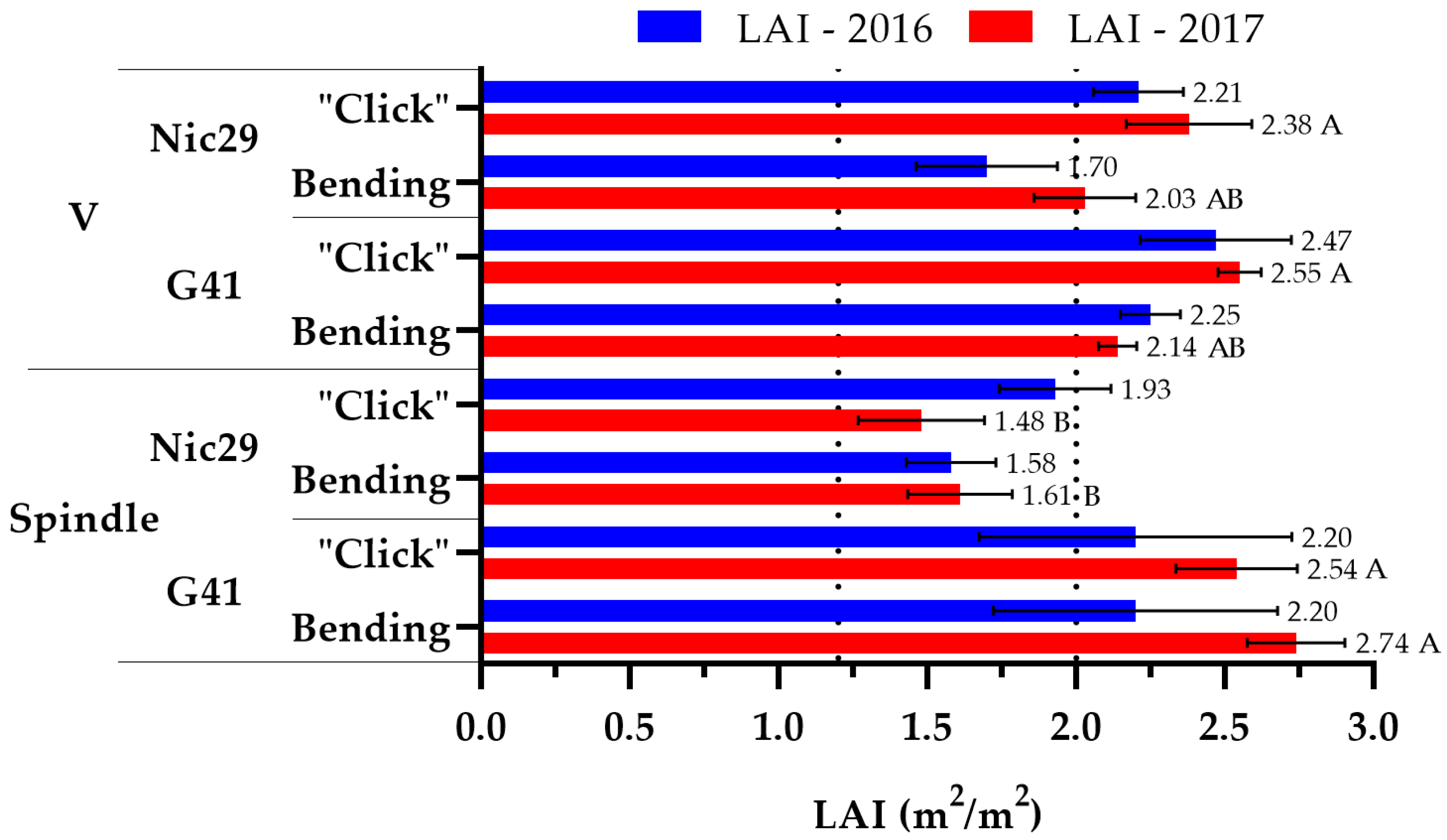
3.1. Leaf Area Estimation
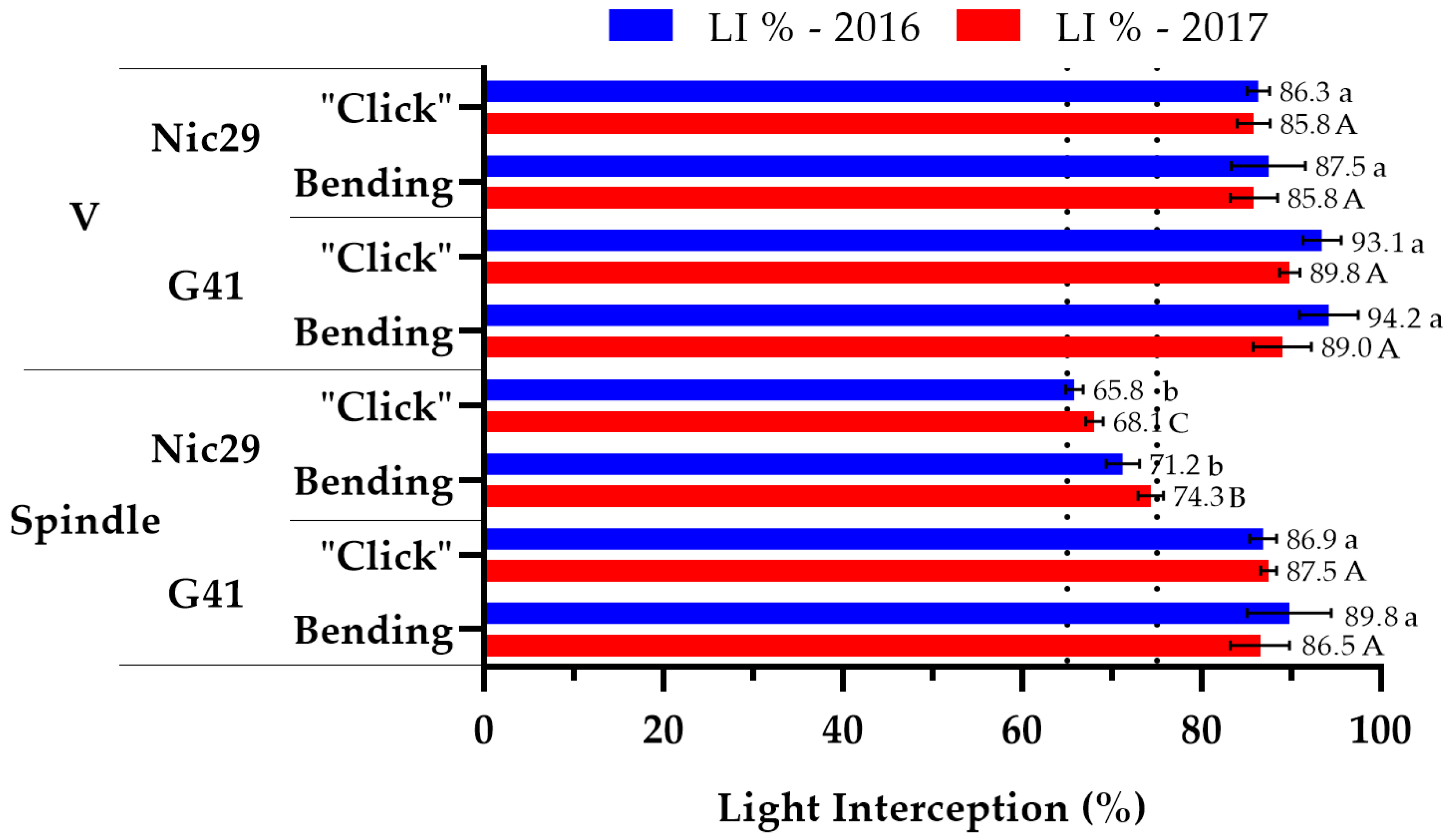
3.2. Light Interception
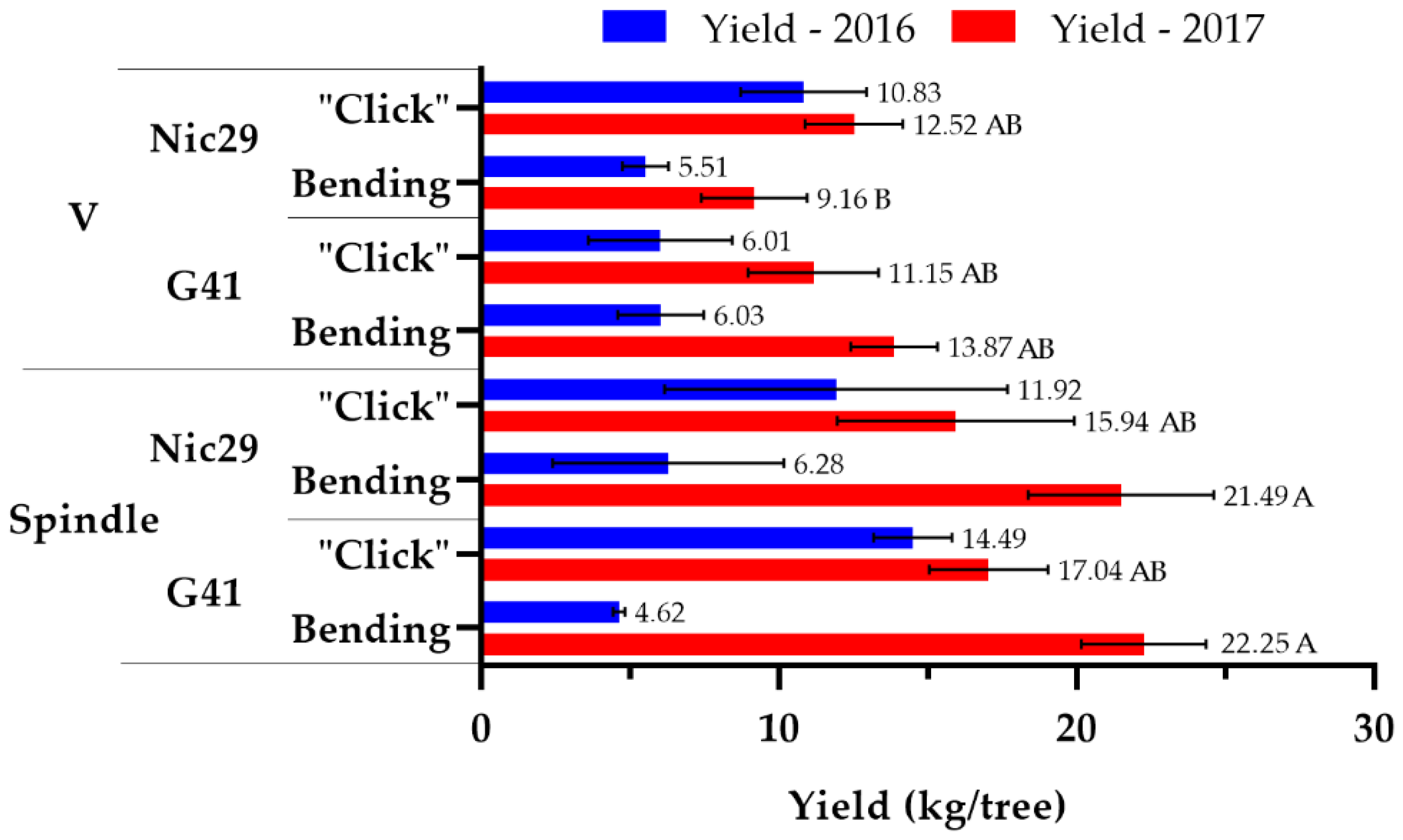
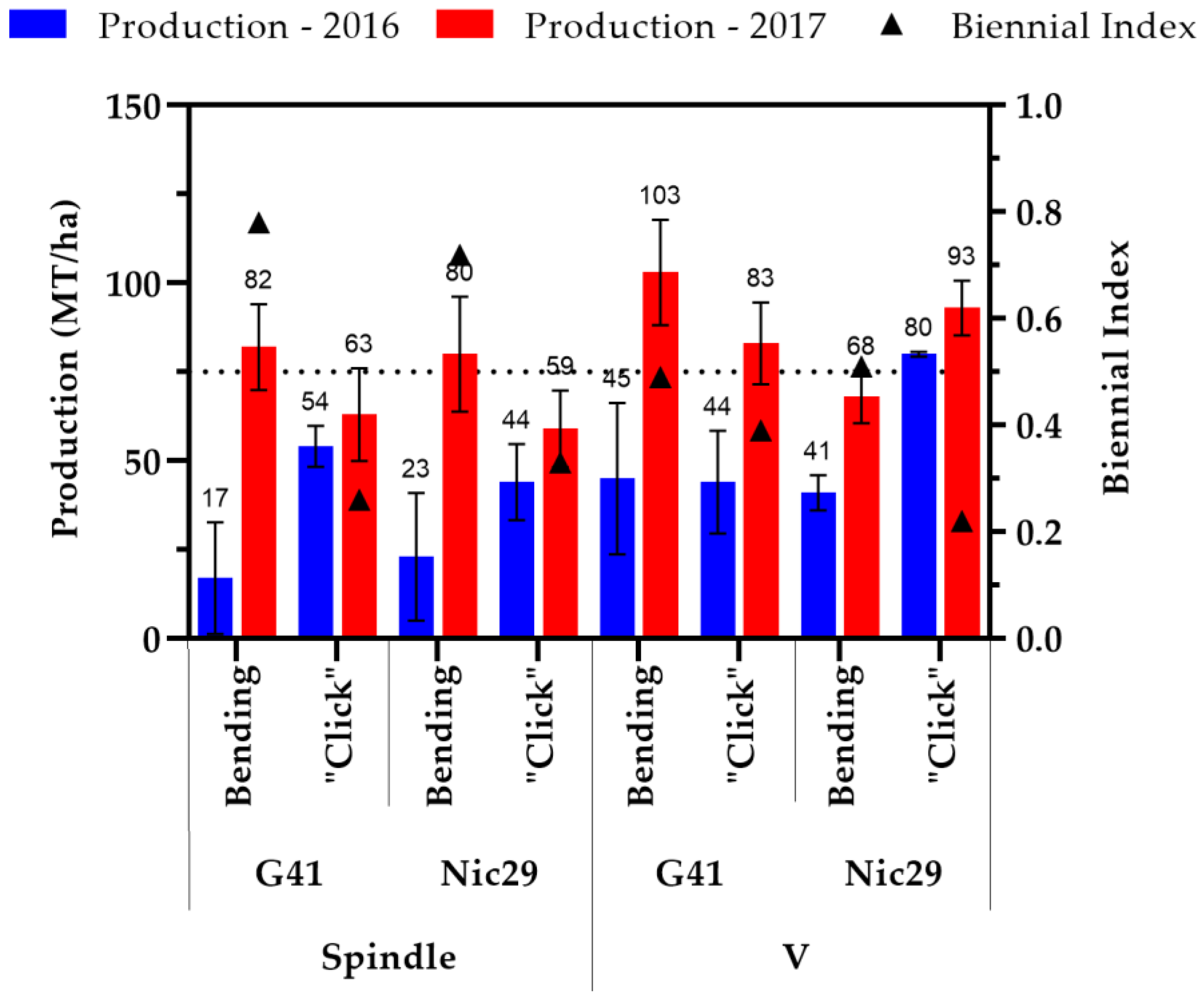
3.3. Production
4. Discussion
4.1. Leaf Area Estimation
4.2. Light Interception
4.3. Production
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Corelli-Grappadelli, L.; Lakso, A.N. Is maximizing orchard light interception always the best choice? In VIII International Symposium on Canopy, Rootstocks and Environmental Physiology in Orchard Systems. Acta Hortic. 2007, 732, 507–518. [Google Scholar] [CrossRef]
- Stephan, J.; Sinoquet, H.; Donès, N.; Haddad, N.; Talhouk, S.; Lauri, P.É. Light interception and partitioning between shoots in apple cultivars influenced by training. Tree Phys. 2008, 28, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Buler, Z.; Mika, A. Evaluation of the ‘Mikado’ tree training system versus the spindle form in apple trees. J. Fruit Orn. Plant Res. 2004, 12, 49–60. [Google Scholar]
- Musacchi, S. Training system and management for a high density orchard of ‘Abbe Fetel’. Acta Hortic. 2011, 909, 225–240. [Google Scholar] [CrossRef]
- Musacchi, S.; Lugli, S. High density planting for sweet cherry orchards. Acta Hortic. 2014, 1020, 489–496. [Google Scholar] [CrossRef]
- Musacchi, S.; Greene, D. Innovations in Apple Tree Cultivation to Manage Crop Load and Ripening. In Achieving Sustainable Cultivation of Apples; Evans, K., Ed.; Burleigh Dodds Science Publishing: Cambridge, UK, 2017; pp. 195–237. [Google Scholar]
- Palmer, J.W. Computed Effects of Spacing on Light Interception and Distribution within Hedgerow Trees in Relation to Productivity. In Symposium on Research and Development on Orchard and Plantation Systems. Acta Hortic. 1980, 114, 80–88. [Google Scholar]
- Robinson, T.L.; Lakso, A.N. Bases of yield and production efficiency in apple orchard systems. J. Am. Soc. Hort. Sci. 1991, 116, 188–194. [Google Scholar] [CrossRef]
- Jackson, J.E.; Palmer, J.W. Interception of light by model hedgerow orchards in relation to latitude, time of year and hedgerow configuration and orientation. J. App. Ecol. 1972, 9, 341–357. [Google Scholar] [CrossRef]
- Palmer, J.W. The effects of row orientation, tree height, time of year and latitude on light interception and distribution in model apple hedgerow canopies. J. Hort. Sci. 1989, 64, 137–145. [Google Scholar] [CrossRef]
- Robinson, T.L.; Wünsche, J.; Lakso, A. The Influence of Orchard System and Pruning Severity on Yield, Light Interception, Conversion Efficiency, Partitioning Index and Leaf Area Index. In V International Symposium on Orchard and Plantation Systems. Acta Hortic. 1992, 349, 123–128. [Google Scholar]
- Wünsche, J.N.; Lakso, A.N. Apple tree physiology: Implications for orchard and tree management. Compact Fruit Tree 2000, 33, 82–88. [Google Scholar]
- Khosravi, S.; Namiranian, M.; Ghazanfariz, H.; Shirvani, A. Estimation of leaf area index and assessment of its allometric equations in oak forests: Northern Zagros, Iran. J. For. Sci. 2012, 58, 116–122. [Google Scholar] [CrossRef]
- Lakso, A.N. Characterizing apple tree canopies by fisheye photography. HortScience 1976, 11, 404–405. [Google Scholar]
- Knerl, A.; Anthony, B.; Serra, S.; Musacchi, S. Optimization of leaf area estimation in a high-density apple orchard using hemispherical photography. HortScience 2018, 53, 799–804. [Google Scholar] [CrossRef]
- Bréda, N.J. Ground-based measurements of leaf area index: A review of methods, instruments and current controversies. J. Exp. Bot. 2003, 54, 2403–2417. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Kang, S.; Li, F.; Li, S.; Du, T. Canopy leaf area index for apple tree using hemispherical photography in arid region. Sci. Hort. 2013, 164, 610–615. [Google Scholar] [CrossRef]
- Chen, J.M.; Rich, P.M.; Gower, S.T.; Norman, J.M.; Plummer, S. Leaf area index of boreal forests: Theory, techniques, and measurements. J. Geophys. Res. Atmos. 1997, 102, 29429–29443. [Google Scholar] [CrossRef]
- Palmer, J.W.; Jackson, J.E. Seasonal light interception and canopy development in hedgerow and bed system apple orchards. J. App. Ecol. 1977, 14, 539–549. [Google Scholar] [CrossRef]
- Barritt, B.H. Influence of orchard system on canopy development, light interception and production of third-year Granny Smith apple trees. In IV International Symposium on Research and Development on Orchard and Plantation Systems. Acta Hortic. 1989, 243, 121–130. [Google Scholar] [CrossRef]
- Lakso, A.N. Correlations of fisheye photography to canopy structure, light climate, and biological responses to light in apple trees. J. Am. Soc. Hort. Sci. 1980, 105, 43–46. [Google Scholar]
- Lakso, A.N. Characterization of apple tree light climate with hemispherical photography. In Symposium on High Density Planting. Acta Hortic. 1978, 65, 71–72. [Google Scholar] [CrossRef]
- Wertheim, S.J. Pruning of slender spindle type trees. Acta Hortic. 1978, 65, 173–180. [Google Scholar] [CrossRef]
- Robinson, T.L. V-Shaped Apple Planting Systems. Acta Hortic. 1998, 513, 337–348. [Google Scholar] [CrossRef]
- Rom, C. Coordination of root and shoot growth: Rootstocks, roots, and rootstocks. In Tree Fruit Physiology: Growth and Development: A Comprehensive Manual for Regulating Deciduous Tree Fruit Growth and Development; Good Fruit Grower: Yakima, WA, USA, 1996; pp. 53–68. [Google Scholar]
- Warner, J. Rootstock affects primary scaffold branch crotch angle of apple trees. HortScience 1991, 26, 1266–1267. [Google Scholar] [CrossRef]
- Miller, S.S.; Hott, C.; Tworkoski, T. Shade effects on growth, flowering and fruit of apple. J. App. Hort. 2015, 17, 101–105. [Google Scholar] [CrossRef]
- Cain, J.C. Hedgerow orchard design for most efficient interception of solar radiation. Effects of tree size, shape, spacing, and row direction. N. Y. St. Agric. Exp. Stn. Search Agric. 1972, 2, 1–14. [Google Scholar]
- Ferree, D.C.; Schupp, J.R. Pruning and training physiology. In Apples: Botany, Production and Uses; CABI Publishing: Cambridge, UK, 2003; pp. 319–344. [Google Scholar]
- He, L.; Schupp, J. Sensing and automation in pruning of apple trees: A review. Agronomy 2018, 8, 211. [Google Scholar] [CrossRef]
- Anthony, B.; Serra, S.; Musacchi, S. Optimizing crop load for new apple cultivar: “WA38”. Agronomy 2019, 9, 107. [Google Scholar] [CrossRef]
- Lespinasse, J.M.; Chol, P.; Dupin, J.; Terenne, E. La conduite du Pommier: Types de Fructification, Incidence sur la conduite de l’arbre; Brochure INVUFLEC: Paris, France, 1977; p. 80. [Google Scholar]
- Giulivo, C. Basic considerations about pruning deciduous fruit trees. Adv. Hort. Sci. 2011, 25, 129–142. [Google Scholar]
- Forshey, C.G.; Elfving, D.C.; Stebbins, R.L. Training and Pruning Apple and Pear Trees; American Society for Horticultural Science: Alexandria, VA, USA, 1992; p. 166. [Google Scholar]
- Robinson, T. Advances in apple culture worldwide. Revista Brasileira de Fruticultura 2011, 33, 37–47. [Google Scholar] [CrossRef]
- Lakso, A.N.; Robinson, T.L. Sunlight, yield and productivity of apples. N. Y. Fruit Q. 2014, 22, 5–7. [Google Scholar]
- Palmer, J.W. Light, canopies, fruit and dollars. Compact Fruit Tree 1999, 32, 119–122. [Google Scholar]
- Robinson, T.L.; Lakso, A.N.; Ren, Z. Modifying apple tree canopies for improved production efficiency. HortScience 1991, 26, 1005–1012. [Google Scholar] [CrossRef]
- Corelli-Grappadelli, L.; Lakso, A.N.; Flore, J.A. Early season patterns of carbohydrate partitioning in exposed and shaded apple branches. J. Am. Soc. Hort. Sci. 1994, 119, 596–603. [Google Scholar] [CrossRef]
- Schechter, I.; Proctor, J.T.A.; Elfving, D.C. Reappraisal of seasonal apple fruit growth. Can. J. Plant Sci. 1993, 73, 549–556. [Google Scholar] [CrossRef]
- Jackson, J.E. Utilization of light resources by HDP systems, In Symposium on High Density Planting 65. Acta Hortic. 1978, 65, 61–70. [Google Scholar] [CrossRef]
- Faust, M. Physiology of Temperate Zone Fruit Trees; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1989. [Google Scholar]
- Heinecke, A.J. The microclimate of fruit trees III the effect of tree size on light penetration and leaf area in red Delicious apple trees. J. Am. Soc. Hort. Sci. 1964, 85, 33–41. [Google Scholar]
- McDonald, M.S. Photobiology of Higher Plants; John Wiley & Sons: Hoboken, NJ, USA, 2003. [Google Scholar]
- Mika, A. Treatments improving illumination of the fruiting zone of the tree. Reg. Photosynth. Fruit Trees 1986, 8, 42–45. [Google Scholar]
- Bepete, M.; Lakso, A.N. Differential effects of shade on early-season fruit and shoot growth rates in ‘Empire’ apple. HortScience 1998, 33, 823–825. [Google Scholar] [CrossRef]
- Hampson, C.; Quamme, H.; Brownlee, R. Tree density or training system-What is important in apple orchard design? Compact. Fruit Tree 2002, 35, 48–50. [Google Scholar]
- Robinson, T.; Hoying, S.; Sazo, M.M.; DeMarree, A.; Dominguez, L. A vision for apple orchard systems of the future. N. Y. Fruit Q. 2013, 21, 11–16. [Google Scholar]
- Olszewski, T.; Mika, A. Planting density effects on mineral composition of leaves and fruits of Macspur apples. Acta Hortic. 1986, 160, 259–260. [Google Scholar]
- Ferree, D.C. Early performance of two apple cultivars in three training systems. HortScience 1994, 29, 1004–1007. [Google Scholar] [CrossRef]
- Serra, S.; Leisso, R.; Giordani, L.; Kalcsits, L.; Musacchi, S. Crop load influences fruit quality, nutritional balance, and return bloom in ‘Honeycrisp’ apple. HortScience 2016, 51, 236–244. [Google Scholar] [CrossRef]
- Zhang, J.; Whiting, M.D.; Zhang, Q. Diurnal pattern in canopy light interception for tree fruit orchard trained to an upright fruiting offshoots (UFO) architecture. Biosyst. Eng. 2015, 129, 1–10. [Google Scholar] [CrossRef]
- Hoblyn, T. Studies in biennial bearing. J. Pomol. Hortic. Sci. 1936, 14, 39–76. [Google Scholar] [CrossRef]
- Laužikė, K.; Samuolienė, G.; Uselis, N. The impact of light penetration into canopy and seasonality on photosynthetic indices in apple tree leaves. AGROFOR Int. J. 2019, 4, 18. [Google Scholar]
- Peper, P.J.; McPherson, E.G. Comparison of five methods for estimating leaf area index of open-grown deciduous trees. J. Arbor. 1998, 24, 98–111. [Google Scholar]
- Chianucci, F.; Cutini, A. Digital hemispherical photography for estimating forest canopy properties: Current controversies and opportunities. IForest 2012, 5, 290–295. [Google Scholar] [CrossRef]
- Anthony, B.; Serra, S.; Musacchi, S. Optimizing “WA38” Management; Tree Fruit Research and Extension Center, Washington State University: Wenatchee, WA, USA, 2017; Unpublished data.
- Verheij, E.W.M.; Verwer, F.L.J.A.W. Light studies in a spacing trial with apple on a dwarfing and a semi-dwarfing rootstock. Sci. Hort. 1973, 1, 25–42. [Google Scholar] [CrossRef]
- Jackson, J.E. Aspects of light climate within apple orchards. J. App. Ecol. 1970, 7, 207–216. [Google Scholar] [CrossRef]
- Wünsche, J.N.; Ferguson, I.B. Crop load interactions in apple. Hort. Rev. 2005, 31, 231–290. [Google Scholar]
- Lespinasse, J.M.; Delort, J.F. Apple Tree Management in Vertical Axis: Appraisal after Ten Years of Experiments. In III International Symposium on Research and Development on Orchard and Plantation Systems. Acta Hortic. 1984, 160, 139–156. [Google Scholar]
- Rom, C.R. Light thresholds for apple tree canopy growth and development. HortScience 1991, 26, 989–992. [Google Scholar] [CrossRef]
- Wertheim, S.J.; Wagenmaker, P.S.; Bootsma, J.H.; Groot, M.J. Orchard systems for apple and pear; conditions for success. Acta Hortic. 2001, 557, 209–227. [Google Scholar] [CrossRef]
- Hrotkó, K. Development in fruit trees production systems. AgroLife Sci. J. 2013, 2, 28–35. [Google Scholar]
- USDA National Agricultural Statistics Service. NASS—Quick Stats. USDA National Agricultural Statistics Service. Available online: https://www.nass.usda.gov/Data_and_Statistics/index.php (accessed on 27 March 2020).
- Warner, G. Taking Yields to the Limit. Good Fruit Grower, 2015. Available online: http://www.goodfruit.com/taking-yields-to-the-limit/ (accessed on 9 September 2017).
- Mupambi, G.; Layne, D.R.; Kalcsits, L.A.; Musacchi, S.; Serra, S.; Schmidt, T.; Hanrahan, I. Use of Protective Netting in Washington State Apple Production; Washington State University Extension: Pullman, WA, USA, 2019. [Google Scholar]
- Fazio, G.; Aldwinckle, H.; Robinson, T. Unique characteristics of Geneva® apple rootstocks. N. Y. Fruit Q. 2013, 21, 25–28. [Google Scholar]
- Wünsche, J.N.; Palmer, J.W. Comparison of non-destructive methods of estimating leaf area in apple tree canopies. Acta Hortic. 1996, 451, 701–708. [Google Scholar] [CrossRef]
- Mohammadi, A.; Mahmoudi, M.J.; Rezaee, R. Vegetative and reproductive responses of some apple cultivars (Malus domestica Borkh.) to heading back pruning. Int. J. Agriscience 2013, 3, 628–635. [Google Scholar]
- Greene, D.W.; Autio, W.R. Notching techniques increase branching of young apple trees. J. Am. Soc. Hort. Sci. 1994, 119, 678–682. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Variables | LAI (m2/m2) | LI (%) | Fruit Count/Tree (No.) | 2016 Yield (kg/Tree) | Average Fruit Weight (g) | 2016 Production (MT/ha) |
|---|---|---|---|---|---|---|---|
| Training system (T) | Spindle | 1.98 | 78.4 B | 28 | 9.33 | 357 | 34.5 |
| V | 2.16 | 90.4 A | 22 | 7.10 | 329 | 52.5 | |
| Significance | ns | *** | ns | ns | ns | ns | |
| Rootstock (R) | Geneva® 41 | 2.28 | 90.9 A | 23 | 7.79 | 352 | 40.0 |
| M9 – Nic29 | 1.85 | 77.3 B | 27 | 8.64 | 333 | 47.1 | |
| Significance | ns | *** | ns | ns | ns | ns | |
| Pruning technique (P) | bending | 1.93 | 85.4 | 16 B | 5.61 B | 347 | 31.4 B |
| “click” | 2.20 | 82.8 | 33 A | 10.81 A | 338 | 55.6 A | |
| Significance | ns | ns | * | * | ns | * |
| Treatment | Variables | LAI (m2/m2) | LI (%) | Fruit Count (No.) | 2017 Yield (kg/Tree) | Average Fruit Weight (g) | 2017 Production (MT/ha) | Total Production (MT/ha) 2016–2017 |
|---|---|---|---|---|---|---|---|---|
| Training system (T) | Spindle | 2.09 | 79.1 B | 86 A | 19.18 A | 236 | 71.0 | 105.6 B |
| V | 2.28 | 87.6 A | 51 B | 11.68 B | 238 | 86.4 | 139.0 A | |
| Significance | ns | *** | ** | ** | ns | ns | * | |
| Rootstock (R) | Geneva® 41 | 2.49 A | 88.1 A | 63 | 16.08 | 261 A | 82.7 | 122.6 |
| M9 – Nic29 | 1.87 B | 78.2 B | 75 | 14.78 | 213 B | 74.8 | 121.9 | |
| Significance | *** | *** | ns | ns | * | ns | ns | |
| Pruning technique (P) | bending | 2.13 | 83.7 | 80 A | 16.69 | 222 | 83.1 | 114.5 |
| “click” | 2.24 | 82.5 | 57 B | 14.16 | 253 | 74.3 | 130.0 | |
| Significance | ns | ns | * | ns | ns | ns | ns |
| Treatment | Variables | Biennial Index (I) | Biennial Classification z |
|---|---|---|---|
| Training system (T) | Spindle | 0.52 | Biennial |
| V | 0.40 | Consistent | |
| Significance | ns | ||
| Rootstock (R) | Geneva® 41 | 0.48 | Consistent |
| M9 – Nic29 | 0.45 | Consistent | |
| Significance | ns | ||
| Pruning technique (P) | bending | 0.62 A | Biennial |
| “click” | 0.30 B | Consistent | |
| Significance | ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anthony, B.; Serra, S.; Musacchi, S. Optimization of Light Interception, Leaf Area and Yield in “WA38”: Comparisons among Training Systems, Rootstocks and Pruning Techniques. Agronomy 2020, 10, 689. https://doi.org/10.3390/agronomy10050689
Anthony B, Serra S, Musacchi S. Optimization of Light Interception, Leaf Area and Yield in “WA38”: Comparisons among Training Systems, Rootstocks and Pruning Techniques. Agronomy. 2020; 10(5):689. https://doi.org/10.3390/agronomy10050689
Chicago/Turabian StyleAnthony, Brendon, Sara Serra, and Stefano Musacchi. 2020. "Optimization of Light Interception, Leaf Area and Yield in “WA38”: Comparisons among Training Systems, Rootstocks and Pruning Techniques" Agronomy 10, no. 5: 689. https://doi.org/10.3390/agronomy10050689
APA StyleAnthony, B., Serra, S., & Musacchi, S. (2020). Optimization of Light Interception, Leaf Area and Yield in “WA38”: Comparisons among Training Systems, Rootstocks and Pruning Techniques. Agronomy, 10(5), 689. https://doi.org/10.3390/agronomy10050689