Identification and Genetic Diversity Analysis of Edible and Medicinal Malva Species Using Flow Cytometry and ISSR Molecular Markers



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
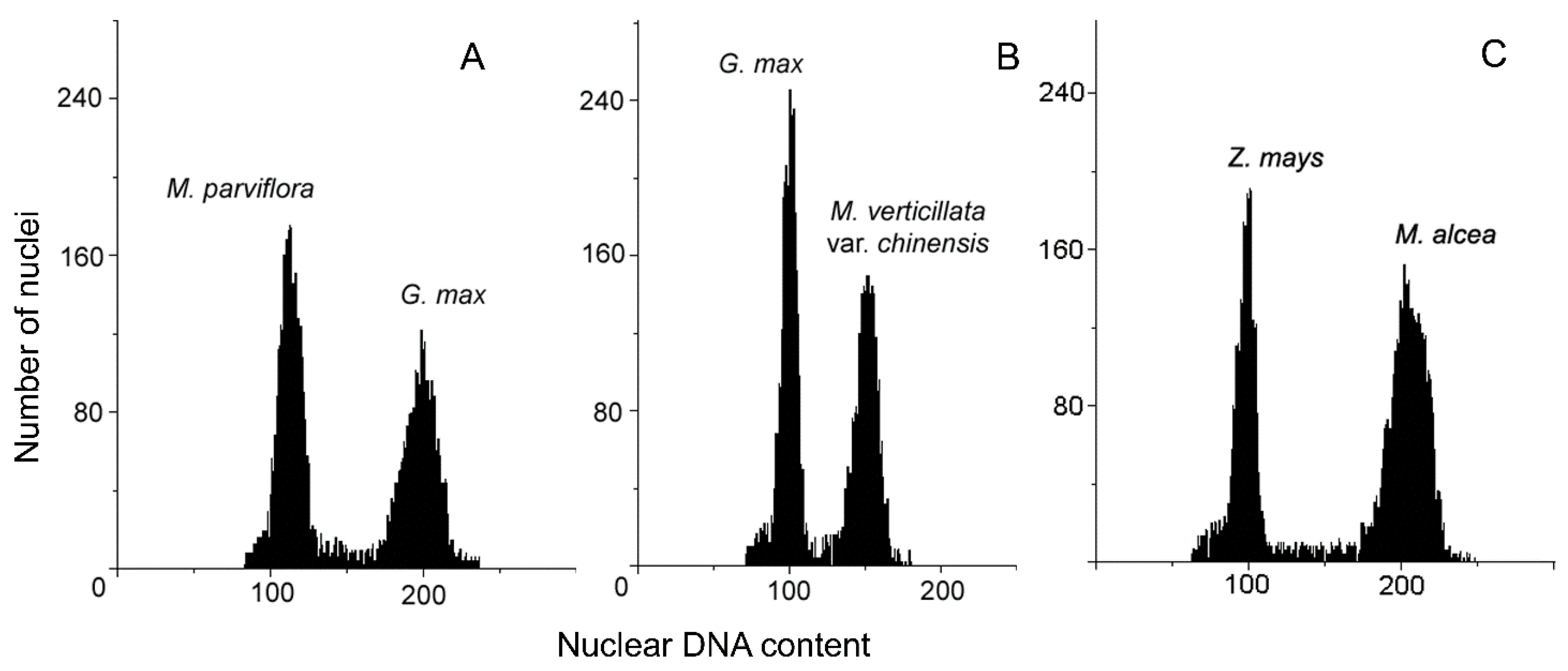
2.2. Genome Size Estimation
2.3. Genomic DNA Extraction
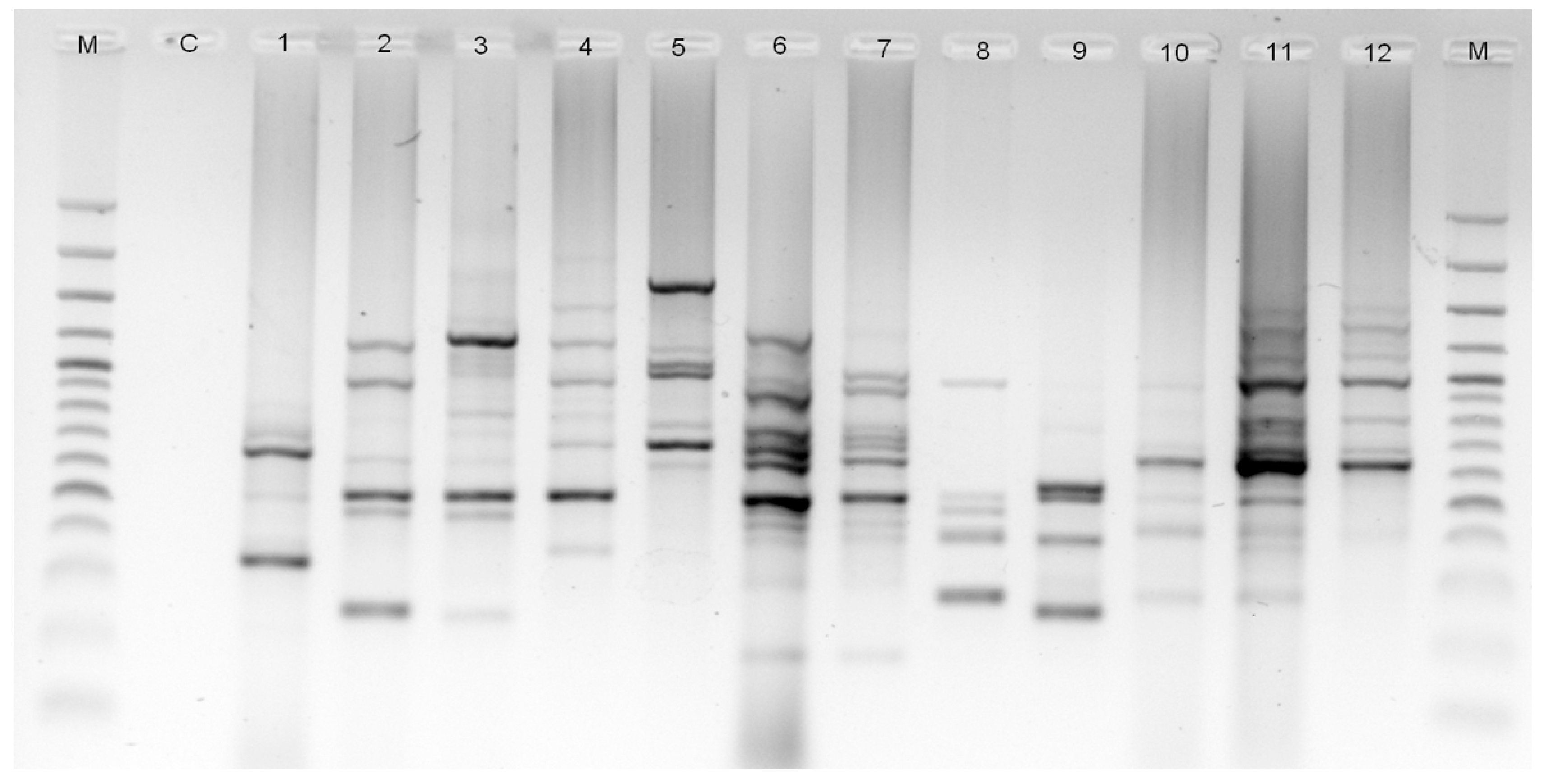
2.4. ISSR-PCR Amplification
2.5. ISSR Marker Analysis
3. Results
3.1. Genome Size
3.2. ISSR Markers
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Dalby, D.H.; Malva, L. Flora Europea. Rosaceae to Umbelliferae; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Weeb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1968; Volume 2, pp. 249–251. [Google Scholar]
- Abedin, S. Malvaceae. In Flora of West Pakistan 130; Nasir, E., Ali, S.I., Eds.; University of Karachi: Karachi, Pakistan, 1979; pp. 1–98. [Google Scholar]
- Ray, M.F. Systematics of Lavatera and Malva (Malvaceae, Malveae)—A new perspective. Plant Syst. Evol. 1995, 198, 29–53. [Google Scholar] [CrossRef]
- El Naggar, S.M. Pollen Morphology of Egyptian Malvaceae: An assessment of taxonomic value. Turk. J. Bot. 2004, 28, 227–240. [Google Scholar]
- Shaheen, N.; Khan, M.A.; Yasmin, G.; Hayat, M.Q.; Ali, S. Taxonomic implication of palynological characters in the genus Malva, L., family Malvaceae from Pakistan. Am. Eurasian J. Agric. Environ. Sci. 2009, 6, 716–722. [Google Scholar]
- Akçin, Ö.E.; Özbucak, T.B. Morphological, anatomical and ecological studies on medicinal and edible plant Malva neglecta Wallr. (Malvaceae). Pak. J. Biol. Sci. 2006, 9, 2716–2719. [Google Scholar] [CrossRef][Green Version]
- DellaGreca, M.; Cutillo, F.; D’Abrosca, B.; Fiorentino, A.; Pacifico, S.; Zarrelli, A. Antioxidant and radical scavenging properties of Malva sylvestris. Nat. Prod. Commun. 2009, 4, 893–896. [Google Scholar] [CrossRef]
- Boual, Z.; Kemassi, A.; Khelil, A.O.E.H.; Michaud, P.; Hadj, M.D.O.E. Partial characterization of water soluble polysaccharides extracted from one Saharian medicinal plant: Malva aegyptiaca L. Proceedings International Conference on Biology. Environ. Chem. 2011, 24, 420–424. [Google Scholar]
- Azab, A. Malva: Food, medicine and chemistry. Eur. Chem. Bull. 2017, 6, 295–320. [Google Scholar] [CrossRef]
- Fakhfakh, N.; Abdelhedi, O.; Jdir, H.; Nasri, M.; Zouari, N. Isolation of polysaccharides from Malva aegyptiaca and evaluation of their antioxidant and antibacterial properties. Int. J. Biol. Macromol. 2017, 105, 1519–1525. [Google Scholar] [CrossRef]
- Badawi, M.A.; El-Sahhar, K.F. Effect of Gibberellin on Some Vegetative Characters of Malva parviflora; Research Bulletin, Faculty of Agriculture Ain Shams University: Cairo, Egypt, 1978; p. 16. [Google Scholar]
- Baytop, T. Dictionary of Turkish Plant Names; Ankara Ataturk Culture, Language and History Institution Press: Ankara, Turkey, 1994; p. 578. [Google Scholar]
- Tanji, A.; Nassif, F. Edible weeds in Morocco. Weed Technol. 1995, 9, 617–620. [Google Scholar] [CrossRef]
- Ranhotra, G.S.; Gelroth, J.A.; Leinen, S.D.; Vinas, M.A.; Lorenz, K.J. Nutritional profile of some edible plants from México. J. Food Compos. Anal. 1998, 11, 298–304. [Google Scholar] [CrossRef]
- Doan, D.; Baolar, S.; Ay, G.; Mert, H.H. The use of wild edible plants in Western and Central Anatolia (Turkey). Econ. Bot. 2004, 58, 684–690. [Google Scholar]
- Baytop, T. The Medicinal and Poisonous Plants of Turkey; Istanbul University Press: Istanbul, Turkey, 1963; p. 1039. [Google Scholar]
- Asimgil, A. Medicinal Plants; Times Press: Istanbul, Turkey, 1993. [Google Scholar]
- Çakilcioğlu, U.; Sengun, M.T.; Türkoglu, I. An ethnobotanical survey of medicinal plants of Yazıkonak and Yurtbaşı districts of Elazığ province, Turkey. J. Med. Plants Res. 2010, 4, 567–572. [Google Scholar]
- Gasparetto, J.C.; Ferreira Martins, C.A.; Hayashi, S.S.; Otuky, M.F.; Pontarolo, R. Ethnobotanical and scientific aspects of Malva sylvestris L.: A millennial herbal medicine. J. Pharm. Pharmacol. 2012, 64, 172–189. [Google Scholar] [CrossRef] [PubMed]
- Hussain, L.; Ikram, J.; Rehman, K.; Tariq, M.; Ibrahim, M.; Akash, M.S.H. Hepatoprotective effects of Malva sylvestris L. against paracetamol-induced hepatotoxicity. Turk. J. Biol. 2014, 38, 396–402. [Google Scholar] [CrossRef]
- Mirghiasi, S.M.; Akhzari, M.; Vassaf, M.; Akbari, A.; Baghi, S.M. The effect of Malva neglecta on the reduction of inflammatory agents in patients with osteoarthritis. Mol. Biol. 2015, 4, 135. [Google Scholar] [CrossRef]
- Keyrouz, E.; El Feghali, P.A.R.; Jaafar, M.; Nawas, T. Malva neglecta: A natural inhibitor of bacterial growth and biofilm formation. J. Med. Plants Res. 2017, 11, 380–386. [Google Scholar] [CrossRef]
- Del Rio, M.; Font, R.; Almela, C.; Velez, D.; Montoro, R.; De Haro Bailon, A. Heavy metals and arsenic uptake by wild vegetation in the Guadiamar river area after the toxic spill of the Aznalcóllar mine. J. Biotechnol. 2002, 98, 125–137. [Google Scholar] [CrossRef]
- Tlustos, P.; Szakova, J.; Hruby, J.; Hartman, I.; Najmanova, J.; Nedelnik, J.; Pavlikova, D.; Batysta, M. Removal of As, Cd, Pb, and Zn from contaminated soil by high biomass producing plants. Plant Soil Environ. 2006, 52, 413–423. [Google Scholar] [CrossRef]
- Ibrahim, M.M.; Alsahli, A.A.; El-Gaaly, G. Evaluation of phytoremediation potential of six wild plants for metal in a site polluted by industrial wastes: A field study in Riyadh, Saudi Arabia. Pak. J. Bot. 2013, 42, 571–576. [Google Scholar]
- Ahmed, Y.M.; Mahmood, A.B.; Ibrahim, H.J. Measuring the accumulation of copper and cadmium in the vegetative parts of the plant and the root of Malva parviflora as a result of irrigation with sewage in city of Kirkuk. Int. J. Curr. Res. Aca. Rev. 2016, 4, 149–154. [Google Scholar] [CrossRef]
- Rahbar, A.; Farjadfard, S.; Leili, M.; Kafaei, R.; Haghshenas, V.; Ramavandi, B. Experimental data of biomaterial derived from Malva sylvestris and charcoal tablet powder for Hg2+ removal from aqueous solutions. Data Brief 2016, 8, 132–135. [Google Scholar] [CrossRef] [PubMed]
- Salahandish, R.; Ghaffarinejad, A.; Norouzbeigi, R. Rapid and efficient lead (II) ion removal from aqueous solutions using Malva sylvestris flower as a green biosorbent. Anal. Methods 2016, 8, 2515–2525. [Google Scholar] [CrossRef]
- Escobar García, P.; Schönswetter, P.; Fuertes Aguilar, J.; Nieto Feliner, G.; Schneeweiss, G.M. Five molecular markers reveal extensive morphological homoplasy and reticulate evolution in the Malva alliance (Malvaceae). Mol. Phylogenet. Evol. 2009, 50, 226–239. [Google Scholar] [CrossRef] [PubMed]
- Ray, M.F. New combinations in Malva (Malvaceae: Malveae). Novon 1998, 3, 288–295. [Google Scholar] [CrossRef]
- Celka, Z.; Szczecińska, M.; Sawicki, J. Genetic relationships between some of Malva species as determined with ISSR and ISJ markers. Biodivers. Res. Conserv. 2010, 19, 23–32. [Google Scholar] [CrossRef]
- Celka, Z.; Szczecińska, M.; Sawicki, J.; Shevera, M.V. Molecular studies did not support the distinctiveness of Malva alcea and M. excise (Malvaceae) in Central and Eastern Europe. Biologia 2012, 67, 1088–1098. [Google Scholar] [CrossRef]
- El Naggar, S.M. Systematic implications of seed coat morphology in Malvaceae. Pak. J. Biol. Sci. 2001, 4, 822–828. [Google Scholar] [CrossRef]
- Celka, Z.; Drapikowska, M.; Buczkowska, K.; Bączkiewicz, A.; Marciniak, J. Morphological variability of Malva alcea L. populations from Poland. In Environmental Changes and Biological Assessment; Kočáarek, P., Plášek, V., Malachová, K., Eds.; Scripta Facultatis Rerum Naturalium Universitatis Ostraviensis: Ostrava, Czech Republic, 2006; Volume 163, pp. 159–165. [Google Scholar]
- Celka, Z.; Szkudlarz, P.; Biereżnoj, U. Morphological variation of hairs in Malva alcea L. (Malvaceae). Biodivers. Res. Conserv. 2006, 3, 258–261. [Google Scholar]
- Celka, Z.; Drapikowska, M.; Jusik, S.; Olejnik, N.; Shevera, M.V.; Szkudlarz, P. Morphological variability of hairs in Malva alcea L. (Malvaceae) populations from Central and Eastern Europe, and consideration of the status of Malva excisa Rchb. Pak. J. Bot. 2015, 47, 467–476. [Google Scholar]
- Celka, Z.; Drapikowska, M.; Ogrodowicz, K.; Shevera, M.V.; Szkudlarz, P. Differentiation of petals in the Malva alcea populations from the region of Central and Eastern Europe. Biodivers. Res. Conserv. 2007, 5, 17–24. [Google Scholar]
- Michael, P.J.; Steadman, K.J.; Plummer, J.A. The biology of Australian weeds 52. Malva parviflora L. Plant Prot. Q. 2009, 24, 2–9. [Google Scholar]
- Makowski, R.M.D.; Morrison, I.N. The biology of Canadian weeds. 91. Malva pusilla Sm. (= M. rotundifolia L.). Can. J. Plant Sci. 1989, 69, 861–879. [Google Scholar] [CrossRef]
- Kristofferson, K.B. Species crossing in Malva. Hereditas 1926, 7, 233–354. [Google Scholar] [CrossRef]
- Olyanitskaya, L.G.; Tzvelev, N.N. Malvaceae. In Flora Europae Orientalis; Tzvelev, N.N., Ed.; ‘Mir i Semia-XCV’: St. Petersburg, Russia, 1996; Volume 9, pp. 231–255. [Google Scholar]
- Lo Bianco, M.; Grillo, O.; Escobar Garcia, P.; Mascia, F.; Venora, G.; Bacchetta, G. Morpho-colorimetric characterization of Malva alliance taxa by seed image analysis. Plant Biol. 2017, 19, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Adamkiewicz, E.; Bijok, K. Badania kariologiczne nad trzema gatunkami rodzaju Malva, L. Acta Soc. Bot. Pol. 1971, 2, 395–400. [Google Scholar]
- Góralski, G.; Judasz, A.; Gacek, P.; Grabowska-Joachimiak, A.; Joachimiak, A.J. Polyploidy, alien species and invasiveness in Polish angiosperms. Plant Syst. Evol. 2014, 300, 225–238. [Google Scholar] [CrossRef][Green Version]
- Jedrzejczyk, I.; Rewers, M. Genome size and ISSR markers for Mentha, L. (Lamiaceae) genetic diversity assessment and species identification. Ind. Crops Prod. 2018, 120, 171–179. [Google Scholar] [CrossRef]
- Lysak, M.A.; Dolezel, J. Estimation of nuclear DNA content in Sesleria (Poaceae). Caryologia 1998, 52, 123–132. [Google Scholar] [CrossRef]
- Doležel, J.; Dolezelova, M.; Novak, F.J. Flow cytometric estimation of nuclear DNA amount in diploid bananas (Musa acuminata and M. balbisiana). Biol. Plant. 1994, 36, 351–357. [Google Scholar] [CrossRef]
- Doležel, J.; Bartoš, J. Plant DNA flow cytometry and estimation of nuclear genome size. Ann. Bot. 2005, 95, 99–110. [Google Scholar] [CrossRef]
- Soltis, D.E.; Soltis, P.S.; Bennett, M.D.; Leitch, I.J. Evolution of genome size in the Angiosperms. Am. J. Bot. 2003, 90, 1596–1603. [Google Scholar] [CrossRef] [PubMed]
- Ghislain, M.; Zhang, D.; Fajardo, D.; Huamán, Z.; Hijmans, R.J. Marker-assisted sampling of the cultivated Andean potato Solanum phureja collection using RAPD markers. Genet. Resour. Crop Evol. 1999, 46, 547–555. [Google Scholar] [CrossRef]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [PubMed]
- Van de Peer, Y.; De Wachter, Y. TREECON for Windows: A software package for the construction and drawing of evolutionary trees for the Microsoft Windows environment. Comput. Appl. Biosci. 1994, 10, 569–570. [Google Scholar] [CrossRef] [PubMed]
- Bidak, L.; Brandham, P.E. Intraspecific uniformity of chromosome number and nuclear DNA quantity in two Egyptian weedy species, Malva parviflora (Malvaceae) and Trigonella stellata (Leguminosae). Kew Bull. 1995, 50, 595–599. [Google Scholar] [CrossRef]
- Bou Dagher-Kharrat, M.; Abdel-Samad, N.; Douaihy, B.; Bourge, M.; Fridlender, A.; Siljak-Yakovlev, S.; Brown, S.C. Nuclear DNA C-values for biodiversity screening: Case of the Lebanese flora. Plant Biosyst. 2013, 147, 1228–1237. [Google Scholar] [CrossRef]
- Bai, C.; Alverson, W.S.; Follansbee, A.; Waller, D.M. New reports of nuclear DNA content for 407 U.S. plant species. Ann. Bot. 2012, 110, 1623–1629. [Google Scholar] [CrossRef]
- Siljak-Yakovlev, S.; Pustahija, F.; Solic, E.M.; Bogunic, F.; Muratovic, E.; Basic, N.; Catrice, O.; Brown, S.C. Towards a genome size and chromosome number database of Balkan flora: C-values in 343 taxa with novel values for 242. Adv. Sci. Lett. 2010, 3, 190–213. [Google Scholar] [CrossRef]
- Ceccarelli, M.; Morosi, L.; Cionini, P.G. Chromocenter association in plant cell nuclei: Determinants, functional significance, and evolutionary implications. Genome 1998, 41, 96–103. [Google Scholar] [CrossRef]
- Wendel, J.F.; Cronn, R.C.; Johnston, J.S.; Price, H.J. Feast and famine in plant genomes. Genetica 2002, 115, 37–47. [Google Scholar] [CrossRef]
- Pustahija, F.; Brown, S.C.; Bogunic, F.; Bašic, N.; Muratovic, E.; Ollier, S.; Hidalgo, O.; Bourge, M.; Stevanovic, V.; Siljak-Yakovlev, S. Small genomes dominate in plants growing on serpentine soils in West Balkans, an exhaustive study of 8 habitats covering 308 taxa. Plant Soil 2013, 373, 427–453. [Google Scholar] [CrossRef]
- Dangi, R.S.; Lagu, M.D.; Choudhary, L.B.; Ranjekar, P.K.; Gupta, V.S. Assessment of genetic diversity in Trigonella foenum–graecum and Trigonella caerulea using ISSR and RAPD markers. BMC Plant Biol. 2004, 4, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Rahimmalek, M.; Bahreininejad, B.; Khorrami, M.; Tabatabaei, B.E. Genetic variability and geographic differentiation in Thymus daenensis subsp. daenensis, an endangered medicinal plant, as revealed by inter simple sequence repeat (ISSR) markers. Biochem. Genet. 2009, 47, 831–842. [Google Scholar] [CrossRef] [PubMed]
- Rewers, M.; Jedrzejczyk, I. Genetic characterization of Ocimum genus using flow cytometry and inter-simple sequence repeat markers. Ind. Crops Prod. 2016, 91, 142–151. [Google Scholar] [CrossRef]
- Jedrzejczyk, I. Study on genetic diversity between Origanum, L. species based on genome size and ISSR markers. Ind. Crops Prod. 2018, 126, 201–207. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Taxon | Accession no. | Origin |
|---|---|---|---|
| M. aegyptia | PI 641694 | Israel | |
| M. alcea | PI 597609 | Poland | |
| M. cretica | ssp. althaeoides | PI 607434 | Spain |
| M. moschata | PI 586620 | Germany | |
| M. neglecta | PI 564836 | Germany | |
| M. nicaeensis | PI 560155 | Germany | |
| M. parviflora | PI 641695 | Portugal | |
| M. sylvestris | var. mauritiana | - | Poland |
| M. tournefortiana | PI 613029 | Portugal | |
| M. verticillata | var. chinensis | PI 631339 | Mongolia |
| M. verticillata | var. crispa | PI 631332 | Georgia |
| M. verticillata | var. verticillata | PI 631333 | South Korea |
| Species | Taxon | DNA Content (2C) | Genome Size Classification b | Internal Standard c | ||
|---|---|---|---|---|---|---|
| pg ± SD | Mbp | |||||
| M. aegyptia | 1.84 ± 0.04 | f a | 1780 | very small | Z | |
| M. alcea | 11.06 ± 0.13 | a | 10,817 | intermediate | Z | |
| M. cretica | ssp. althaeoides | 2.77 ± 0.16 | e | 2709 | very small | Z |
| M. moschata | 2.99 ± 0.03 | d | 2924 | small | Z | |
| M. neglecta | 1.48 ± 0.02 | ij | 1447 | very small | G | |
| M. nicaeensis | 1.55 ± 0.02 | h | 1496 | very small | G | |
| M. parviflora | 1.45 ± 0.02 | j | 1418 | very small | G | |
| M. sylvestris | var. mauritiana | 1.63 ± 0.02 | g | 1594 | very small | G |
| M. tournefortiana | 2.81 ± 0.02 | e | 2748 | small | Z | |
| M. verticillata | var. chinensis | 3.80 ± 0.04 | b | 3716 | small | G |
| M. verticillata | var. crispa | 3.67 ± 0.02 | c | 3589 | small | G |
| M. verticillata | var. verticillata | 3.71 ± 0.05 | c | 3628 | small | G |
| Mean | 3.23 | 3156 | ||||
| Primer Code | Primer Sequence (5’–3’) | Annealing Temperature (˚C) | No. of Total Alleles | No. of Polymorphic Alleles | Percentage of Polymorphism | PIC |
|---|---|---|---|---|---|---|
| ISSR-1 | (GACA)4 | 49.0 | 17 | 17 | 100 | 0.390 |
| ISSR-4 | (GTG)6A | 58.4 | 20 | 20 | 100 | 0.393 |
| ISSR-5 | (GACA)4T | 49.0 | 15 | 15 | 100 | 0.396 |
| ISSR-6 | (GACA)4A | 49.0 | 25 | 25 | 100 | 0.388 |
| ISSR-7 | (GTG)6T | 58.4 | 20 | 20 | 100 | 0.386 |
| ISSR-20 | (TGAG)4 | 52.5 | 19 | 19 | 100 | 0.392 |
| ISSR-38 | (GTC)3GC | 50.6 | 17 | 17 | 100 | 0.401 |
| ISSR-39 | (CT)8GC | 50.0 | 16 | 16 | 100 | 0.434 |
| ISSR-42 | (AG)8YC | 51.0 | 17 | 17 | 100 | 0.466 |
| ISSR-44 | (CA)8GC | 51.0 | 15 | 15 | 100 | 0.364 |
| ISSR-47 | (GA)8YT | 51.0 | 24 | 24 | 100 | 0.392 |
| ISSR-49 | (AC)8YA | 51.0 | 20 | 20 | 100 | 0.380 |
| ISSR-50 | (GT)8YC | 51.0 | 18 | 18 | 100 | 0.350 |
| Average | 19 | 19 | 100 | 0.395 | ||
| Species | Lp. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M. aegyptia | 1 | 0.00 | |||||||||||
| M. alcea | 2 | 0.69 | 0.00 | ||||||||||
| M. cretica ssp. althaeoides | 3 | 0.69 | 0.70 | 0.00 | |||||||||
| M. moschata | 4 | 0.69 | 0.54 | 0.65 | 0.00 | ||||||||
| M. neglecta | 5 | 0.72 | 0.77 | 0.75 | 0.73 | 0.00 | |||||||
| M. nicaeensis | 6 | 0.73 | 0.73 | 0.67 | 0.74 | 0.56 | 0.00 | ||||||
| M. parviflora | 7 | 0.73 | 0.77 | 0.67 | 0.78 | 0.61 | 0.22 | 0.00 | |||||
| M. sylvestris var. mauritiana | 8 | 0.76 | 0.81 | 0.75 | 0.77 | 0.70 | 0.60 | 0.53 | 0.00 | ||||
| M. tournefortiana | 9 | 0.63 | 0.68 | 0.66 | 0.47 | 0.68 | 0.74 | 0.75 | 0.75 | 0.00 | |||
| M. verticillata var. chinensis | 10 | 0.75 | 0.74 | 0.66 | 0.70 | 0.68 | 0.55 | 0.56 | 0.69 | 0.69 | 0.00 | ||
| M. verticillata var. crispa | 11 | 0.67 | 0.66 | 0.63 | 0.69 | 0.72 | 0.61 | 0.58 | 0.65 | 0.68 | 0.29 | 0.00 | |
| M. verticillata var. verticillata | 12 | 0.66 | 0.71 | 0.63 | 0.69 | 0.73 | 0.58 | 0.59 | 0.70 | 0.65 | 0.29 | 0.28 | 0.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jedrzejczyk, I.; Rewers, M. Identification and Genetic Diversity Analysis of Edible and Medicinal Malva Species Using Flow Cytometry and ISSR Molecular Markers. Agronomy 2020, 10, 650. https://doi.org/10.3390/agronomy10050650
Jedrzejczyk I, Rewers M. Identification and Genetic Diversity Analysis of Edible and Medicinal Malva Species Using Flow Cytometry and ISSR Molecular Markers. Agronomy. 2020; 10(5):650. https://doi.org/10.3390/agronomy10050650
Chicago/Turabian StyleJedrzejczyk, Iwona, and Monika Rewers. 2020. "Identification and Genetic Diversity Analysis of Edible and Medicinal Malva Species Using Flow Cytometry and ISSR Molecular Markers" Agronomy 10, no. 5: 650. https://doi.org/10.3390/agronomy10050650
APA StyleJedrzejczyk, I., & Rewers, M. (2020). Identification and Genetic Diversity Analysis of Edible and Medicinal Malva Species Using Flow Cytometry and ISSR Molecular Markers. Agronomy, 10(5), 650. https://doi.org/10.3390/agronomy10050650