Zinc Oxide and Silicone Nanoparticles to Improve the Resistance Mechanism and Annual Productivity of Salt-Stressed Mango Trees



Abstract
1. Introduction
2. Materials and Methods
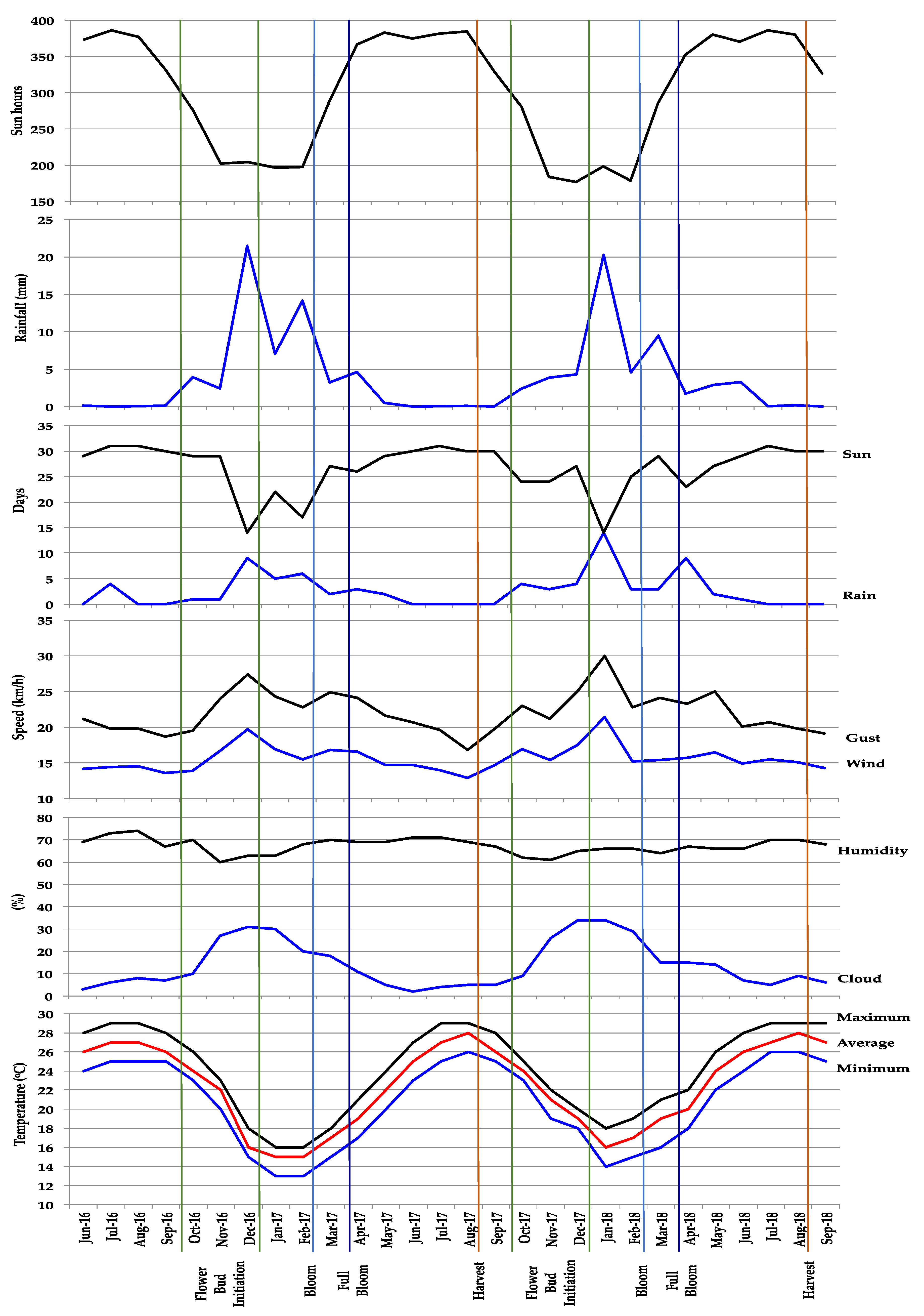
2.1. Experiment
2.2. Floral Malformation Assessment (%)
2.3. Leaf Area (cm2)
2.4. Leaf Analysis
2.5. Yield and Fruit Physiochemical Characteristics
2.6. Statistical Analysis
3. Results and Discussion
3.1. Leaf Area
3.2. Leaf NPK Content
3.3. Carbohydrates and Osmostimulants
3.4. Antioxidant Enzymes Activity
3.5. Floral Malformation
3.6. Yield and Fruit Quality
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Masroor, H.M.; Anjum, M.A.; Hussain, S.; Ejaz, S.; Ahmad, S.; Ercisli, S.; Zia-Ul-Haq, M. Zinc ameliorates fruit yield and quality of mangoes cultivated in calcareous soils. Erwerbs-Obstbau 2016, 58, 49–55. [Google Scholar] [CrossRef]
- United Nations Conference on Trade and Development (UNCTAD). Mango: An INFOCOMM Commodity Profile; UNCTAD Trust Fund on Market Information on Agricultural Commodities; United Nations Conference on Trade and Development (UNCTAD), United Nations: New York, NY, USA; Geneva, Switzerland, 2016; p. 21. [Google Scholar]
- Abdel-Hamid, M.A.; Shrestha, D.P. Soil Salinity Mapping in The Nile Delta, Egypt Using Remote Sensing Techniques. In Interpretation of Photographic and Remote Sensing Data, Technical Commission Book 7, Proceedings of the 29th International Society for Photogrammetry and Remote Sensing (ISPRS) Congress, Washington, DC, USA, 2–14 August 1992; Fritz, L.W., Lucas, J.R., Eds.; ISPRS Archives, University of New South Wales: Sydney, Australia, 1992; Volume 29, pp. 783–787. [Google Scholar]
- Food and Agriculture Organization of the United Nations (FAO). FAO Statistics; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2018; Available online: http://www.fao.org/faostat/en/#data/QC/visualize (accessed on 5 April 2020).
- Munir, T.; Perkins, M.; Kaing, E.; Strack, M. Carbon dioxide flux and net primary production of a boreal treed bog: Responses to warming and water-table-lowering simulations of climate change. Biogeochemistry 2015, 12, 1091–1111. [Google Scholar] [CrossRef]
- Kumar, R.; Kumar, V. Physiological disorders in perennial woody tropical and subtropical fruit crops—A review. Indian J. Agric. Sci. 2016, 86, 703–717. [Google Scholar]
- Khattab, M.M.; Haseeb, G.M.; Elkhishen, M.A. The effect of post harvest pruning, GA3 and Ethrel concentrations on some flowering attributes and yield of some mango cultivars. Bul. Fac. Agric. Cairo Univ. 2009, 60, 306–314. [Google Scholar]
- Chadha, K.L. Fruit drop in mango. In Advances in Horticulture; Chadha, K.L., Pareek, O.P., Eds.; Malhotra Publishing House: New Delhi, India, 1993; pp. 1131–1166. [Google Scholar]
- Bolat, I.; Dikilitas, M.; Ikinci, A.; Ercisli, S.; Tonkaz, T. Morphological, physiological, biochemical characteristics and bud success responses of myrobolan 29 c plum rootstock subjected to water stress. Can. J. Plant Sci. 2015, 96, 485–493. [Google Scholar] [CrossRef]
- Saiki, S.T.; Ishida, A.; Yoshimura, K.; Yazaki, K. Physiological mechanisms of drought-induced tree die-off in relation to carbon, hydraulic and respiratory stress in a drought-tolerant woody plants. Sci. Rep. 2017, 7, 2995. [Google Scholar] [CrossRef]
- Tahir, F.M.; Ibrahim, M.; Hamid, K. Effect of drought stress on vegetative and reproductive growth behavior of mango (Mangifera indica L.). Asian J. Plant Sci. 2003, 2, 116–118. [Google Scholar]
- Yamaguchi, T.; Blumwald, E. Developing salt-tolerant crop plants: Challenges and opportunities. Trends Plant Sci. 2005, 10(12), 615–620. [Google Scholar] [CrossRef]
- Dubey, A.K.; Singh, A.K.; Srivastav, M. Salt stress studies in mango—A review. Agric. Rev. 2007, 28, 75–78. [Google Scholar]
- Grieve, C.M.; Grattan, S.R.; Maas, E.V. Plant Salt Tolerance. In ASCE Manual and Reports on Engineering Practice No. 71: Agricultural Salinity Assessment and Management, 2nd ed.; Wallender, W.W., Tanji, K.K., Eds.; American Society of Civil Engineers (ASCE) library: Reston, VA, USA, 2012; Chapter 13; pp. 405–459. [Google Scholar]
- Zhu, J.K. Plant salt stress. In Encyclopedia of Life Science, 2nd ed.; O’Daly, A., Ed.; Wiley, J. & Sons, Ltd.: Chichester, UK, 2007; pp. 1–3. [Google Scholar]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Jindal, P.C.; Singh, J.P.; Gupta, O.P. Salt tolerance of mango: A note on phosphorus and magnesium deficiency caused by sodium sulphate. Haryana J. Hort. Sci. 1976, 5, 13–14. [Google Scholar]
- Dayal, V.; Dubey, A.K.; Awasthi, O.P.; Pandey, R.; Dahuja, A. Growth, lipid peroxidation, antioxidant enzymes and nutrient accumulation in Amrapali mango (Mangifera indica L) grafted on different rootstocks under NaCl stress. Plant Knowl. J. 2014, 3, 15–22. [Google Scholar]
- Ansari, M.W.; Rani, V.; Shukla, A.; Bains, G.; Pant, R.C.; Tuteja, N. Mango (Mangifera indica L.) malformation: A malady of stress ethylene origin. Physiol. Mol. Biol. Plants 2015, 21, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Laane, H.-M. The Effects of Foliar Sprays with Different Silicon Compounds. Plants 2018, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, V.; Sotiropoulos, T.; Brown, P.H. Foliar Fertilization: Scientific Principles and Field Practices, 1st ed.; International Fertilizer Industry Association (IFA): Paris, France, 2013; p. 140. [Google Scholar]
- Scott, N.; Chen, H. Nanoscale Science and Engineering for Agriculture and Food Systems. A Report Submitted to Cooperative State Research, Education and Extension Service; National Planning Workshop, The United States Department of Agriculture (USDA): Washington, DC, USA, 18–19 November 2002; Report Published in September 2003; p. 62. Available online: http://www.nseafs.cornel.edu/web.roadmap.pdf (accessed on 12 January 2020).
- Maurer-Jones, M.A.; Gunsolus, I.L.; Murphy, C.J.; Haynes, C.L. Toxicity of engineered nanoparticles in the environment. Anal. Chem. 2013, 85, 3036–3049. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Dominguez, M.; Boutonnet, M.; Solans, C. A novel approach to metal and metal oxide nanoparticle synthesis: The oil-in-water microemulsion reaction method. J. Nanopart. Res. 2009, 11, 1823–1829. [Google Scholar] [CrossRef]
- Mosanna, R.; Khalilvand, B.E. Morpho-physiological response of maize (Zea mays L.) to zinc nano-chelate foliar and soil application at different growth stages. J. New Biol. Rep. 2015, 4, 46–50. [Google Scholar]
- Boutchuen, A.; Zimmerman, D.; Aich, N.; Masud, A.M.; Arabshahi, A.; Palchoudhury, S. Increased Plant Growth with Hematite Nanoparticle Fertilizer Drop and Determining Nanoparticle Uptake in Plants Using Multimodal Approach. J. Nanomater. 2019, 2019, 6890572. [Google Scholar] [CrossRef]
- Helaly, M.N.; El-Hoseiny, H.; El-Sheery, N.I.; Rastogi, A.; Kalaji, H.M. Regulation and physiological role of silicon in alleviating drought stress of mango. Plant Physiol. Biochem. 2017, 118, 31–44. [Google Scholar] [CrossRef]
- Swietlik, D. Zinc nutrition in horticultural crops. Hort. Rev. 1999, 23, 109–180. [Google Scholar]
- Marschner, H. Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: Cambridge, MA, USA, 2011; pp. 347–364. [Google Scholar]
- Ojeda-Barrios, D.L.; Perea-Portillo, E.; Hernández-Rodríguez, O.A.; Martínez-Téllez, J.; Abadía, J.; Lombardini, L. Foliar fertilization with zinc in pecan trees. HortScience 2014, 49, 562–566. [Google Scholar] [CrossRef]
- Ahmad, I.; Bibi, F.; Ullah, H.; Munir, T.M. Mango Fruit Yield and Critical Quality Parameters Respond to Foliar and Soil Applications of Zinc and Boron. Plants 2018, 7, 97. [Google Scholar] [CrossRef] [PubMed]
- Brennan, R.F. Zinc Application and Its Availability to Plants. Ph.D. Thesis, School of Environmental Science, Division of Science and Engineering, Murdoch University, Perth, Australia, 2005. [Google Scholar]
- Augusti, M.; Almela, V.; Andreu, I.; Juan, M.; Zacarias, L. Synthetic auxin 3,5,6-TPA promotes fruit development and climacteric in Prunus persica L. Batsch. J. Hort. Sci. Biotechnol. 1999, 74, 556–560. [Google Scholar] [CrossRef]
- Singh, J.; Kumar, S.; Alok, A.; Upadhyay, S.K.; Rawat, M.; Tsang, D.C.W.; Bolan, N.; Kim, K.-H. The potential of green synthesized zinc oxide nanoparticles as nutrient source for plant growth. J. Clean. Prod. 2019, 214, 1061–1070. [Google Scholar] [CrossRef]
- Prasad, T.N.V.K.V.; Sudhakar, P.; Sreenivasulu, Y.; Latha, P.; Munaswamy, V.; Raja Reddy, K.; Sreeprasad, T.S.; Sajanlal, P.R.; Pradeep, T. Effect of nanoscale zinc oxide particles on the germination, growth and yield of peanut. J. Plant Nutr. 2012, 35, 905–927. [Google Scholar] [CrossRef]
- Garcia-Gomes, C.; Obradon, A.; Gonzalez, D.; Babin, M.; Fernandez, M.D. Comparative effect of ZnO NPs, ZnO bulk and ZnSO4 in the antioxidant defenses of two plant species growing in two agricultural soils under greenhouse conditions. Sci. Total Environ. 2017, 589, 11–24. [Google Scholar] [CrossRef]
- Davarpanah, S.; Tehranifar, A.; Davarynejad, G.; Abadía, J.; Khorasani, R. Effects of foliar applications of zinc and boron nanofertilisers on pomegranate (Punica granatum cv. Ardestani) fruit yield and quality. Sci. Hort. 2016, 210, 57–64. [Google Scholar] [CrossRef]
- Zagzog, O.A.; Gad, M.M. Improving Growth, Flowering, Fruiting and Resistance of Malformation of Mango Trees using Nano-Zinc. Middle East J. Agric. 2017, 6, 673–681. [Google Scholar]
- Imtiaz, M.; Rizwan, M.S.; Mushtaq, M.A.; Ashraf, M.; Shahzad, S.M.; Yousaf, B.; Tu, S. Silicon occurrence, uptake, transport and mechanisms of heavy metals, minerals and salinity enhanced tolerance in plants with future prospects: A review. J. Environ. Manag. 2016, 183, 521–529. [Google Scholar] [CrossRef]
- Ma, J.F.; Takahasi, E. Soil, Fertilizer, and Plant Silicon Research in Japan, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2002; p. 294. [Google Scholar]
- Sivanesan, I.; Park, S.W. The role of silicon in plant tissue culture. Front. Plant Sci. 2014, 5, 571. [Google Scholar] [CrossRef]
- Artyszak, A. Effect of Silicon Fertilization on Crop Yield Quantity and Quality—A Literature Review in Europe. Plants 2018, 7, 54. [Google Scholar] [CrossRef] [PubMed]
- Balakhnina, T.; Borkowska, A. Effects of silicon on plant resistance to environmental stresses: Review. Int. Agrophys. 2013, 27, 225–232. [Google Scholar] [CrossRef]
- World Weather Online. Damietta Historical Weather; Egypt Historical Weather Almanac: Manchester, UK, 2020; Available online: https://www.worldweatheronline.com/damietta-weather-history/dumyat/eg.aspx (accessed on 24 February 2020).
- Helaly, M.N.; El-Sheery, N.I.; El-Hoseiny, H.; Rastogi, A.; Kalaji, H.M.; Zabochnicka-Swiatek, M. Impact of treated wastewater and salicylic acid on physiological performance, malformation and yield of two mango cultivars. Sci. Hort. 2018, 233, 159–177. [Google Scholar] [CrossRef]
- Ahmed, F.F.; Morsy, M.H. A new method for measuring leaf area in different fruit species. Minia J. Agric. Res. Develop. 1999, 19, 97–105. [Google Scholar]
- Wilde, S.A.; Corey, R.B.; Layer, J.G.; Voigt, G.K. Soils and Plant Analys is for Tree Culture, 3rd ed.; Oxford and IBH publishing Co.: New Delhi, India, 1985; pp. 529–546. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebens, P.A.; Smith, F. Colorimetric methods for determination sugars and related substances. Ann. Chem. Soc. 1956, 46, 1662–1669. [Google Scholar] [CrossRef]
- Ackerson, R.C.; Krieg, R.D. Stomatal and nonstomatal regulation of water-use in cotton, corn and sorghum. Plant Physiol. 1977, 60, 850–853. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldern, R.P.; Teara, I.D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Bergmeyer, H.-U. Methods of Enzymatic Analysis, 2nd ed.; Academic Press: Berlin, Germany, 1974; Volume 4, p. 800. [Google Scholar]
- Herzog, V.; Fahimi, H.D. Intracellular distinction between peroxidase andcatalase in exocrine cells of rat lacrimal gland: A biochemical and cytochemical study. Histochemistry 1976, 46, 273–286. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists. Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2000; pp. 490–510. [Google Scholar]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 7th ed.; Iowa State University Press: Ames, IA, USA, 1980; p. 507. [Google Scholar]
- Pokhrel, L.R.; Dubey, B. Evaluation of developmental responses of two crop plants exposed to silver and zinc oxide nanoparticles. Sci. Total Environ. 2013, 452–453, 321–332. [Google Scholar] [CrossRef]
- Yoon, S.-J.; Kwak, J., II; Lee, W.-M.; Holden, P.A.; An, Y.-J. Zinc Oxide nanoparticles delay soybean development: A standard soil microcosm study. Ecotoxicol. Environ. Saf. 2014, 100, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Broadley, M.R.; White, P.J.; Hammond, J.P.; Zelko, I.; Lux, A. Tansley review: Zinc in plants. New Phytol. 2007, 173, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Asmar, S.A.; Pasqual, M.; Rodrigues, F.A.; de Araujo, A.G.; Pio, L.A.S.; e Silva, S.O. Sources of silicon in the development of micropropagated seedlings of banana ‘Maçã’. Cienc. Rural. 2011, 41, 1127–1131. [Google Scholar] [CrossRef][Green Version]
- Silva, A.J.; Nascimento, C.W.A.; Gouveia-Neto, A.S. Assessment of cadmium phytotoxicity alleviation by silicon using chlorophyll a fluorescence. Photosynthetica 2017, 55, 648–654. [Google Scholar] [CrossRef]
- Teija Ruuhola, T.L. Retranslocation of nutrients in relation to boron availability during leaf senescence of Betula pendula Roth. Plant Soil 2011, 344, 227–240. [Google Scholar] [CrossRef]
- Lechaudel, M.; Joas, J.; Caro, Y.; Genard, M.; Jannoyer, M. Leaf: Fruit ratio and irrigation supply affect seasonal changes in minerals, organic acids and sugars of mango fruit. J. Sci. Food Agric. 2005, 85, 251–260. [Google Scholar] [CrossRef]
- Nejad, R.H.; Najafi, F.; Arvin, P.; Riruzeh, R. Study Different Levels of Zinc Sulphate (ZnSO4) on Fresh and Dry Weight, Leaf Area, Relative Water Content and Total Protein in Bean (Phaseolus vulgaris L.). Plant. Bull. Env. Pharmacol. Life Sci. 2014, 3, 144–151. [Google Scholar]
- Lakshmipathi, J.D.; Adiga, D.; Kalaivanan, B.M.; Muralidhara, B.; Preethi, P. Effect of Zinc and Boron Application on Leaf Area, Photosynthetic Pigments, Stomatal Number and Yield of Cashew. Int. J. Curr. Microbiol. App. Sci. 2018, 7, 1786–1795. [Google Scholar]
- Haripriya, P.; Stella, P.M.; Anusuya, S. Foliar Spray of Zinc Oxide Nanopartcles Improves Salt Tolerance in Finger Millet Crops under Glasshouse Conditon. SCIOL Biotechnol. 2018, 1, 20–29. [Google Scholar]
- Bhusal, N.; Han, S.-G.; Yoon, T.-M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh.). Sci. Hort. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Le, V.N.; Rui, Y.; Gui, X.; Li, X.; Liu, S.; Han, Y. Uptake, transport, distribution and Bio-effects of SiO2 nanoparticles in Bt-transgenic cotton. J. Nanobiotechnol. 2014, 5, 12–50. [Google Scholar] [CrossRef] [PubMed]
- Bar, M.; Ori, N. Leaf development and morphogenesis. Development 2014, 141, 4219–4230. [Google Scholar] [CrossRef] [PubMed]
- Bhavya, H.K.; Nache Gowda, V.V.; Jaganath, S.; Sreenivas, K.N.; Prakash, N.B. Effect of foliar silicic acid and boron acid in Bangalore blue grapes. In Proceedings of the 5th International Conference on Silicon in Argriculture, Beijing, China, 11–19 September 2011; pp. 7–8. [Google Scholar]
- Pilon, C.; Soratto, R.P.; Moreno, L.A. Effects of soil and foliar application of soluble silicon on mineral nutrition, gas exchange, and growth of Potato plants. Crop Sci. 2013, 53, 1605–1614. [Google Scholar] [CrossRef]
- Kleiber, T.; Krzesinski, W.; Przygocka-Cyna, K.; Spizewski, T. The response of hydroponically grown lettuce under Mn stress to differentiated application of silica sol. J. Elem. 2015, 20, 609–619. [Google Scholar] [CrossRef]
- Cakmak, I.; Marschner, H. Mechanism of phosphorus-induced zinc deficiency in cotton. I. Zinc deficiency-enhanced uptake rate of phosphorus. Physiol. Plant. 1986, 68, 483–490. [Google Scholar] [CrossRef]
- Wiese, H.; Nikolic, M.; Romheld, V. Silicon in Plant Nutrition. In The Apoplast of Higher Plants: Compartment of Storage, Transport and Reactions; Sattelmacher, B., Horst, W.J., Eds.; Springer: Dordrecht, Swizerland, 2007; pp. 33–47. [Google Scholar]
- Nair, P.M.G.; Chung, M., III. Regulation of morphological, molecular and nutrient status in Arabidopsis thaliana seedlings in response to ZnO nanoparticles and Zn ion exposure. Sci. Total Environ. 2017, 575, 187–198. [Google Scholar] [CrossRef]
- Khan, A.S.; Ullah, W.; Malik, A.U.; Ahmad, R.; Saleem, B.A.; Rajwana, I.A. Exogenous applications of boron and zinc influence leaf nutrient status, tree growth and fruit quality of Futrell’s early (citrus Reticulata Blanco). Pak. J. Agric. Sci. 2012, 49, 113–119. [Google Scholar]
- Rizwan, M.; Ali, S.; Ali, B.; Adrees, M.; Arshad, M.; Hussain, A.; Rehman, M.Z.; Waris, A.A. Zinc and iron oxide nanoparticles improved the plant growth and reduced oxidative stress and cadmium concentration in wheat. Chemosphere 2019, 214, 269–277. [Google Scholar] [CrossRef]
- Kalteh, M.; Alipour, Z.T.; Ashraf, S.; Aliabadi, M.M.; Nosratabadi, A.F. Effect of silica nanoparticles on basil (Ocimum basilicum) under salinity stress. J. Chem. Health Risks 2014, 4, 49–55. [Google Scholar]
- Al-Wasfy, M.M. Response of Sakkoti date palms to foliar application of royal jelly, silicon and vitamins B. J. Am. Sci. 2013, 9, 315–321. [Google Scholar]
- Hajiboland, R.; Cherghvareh, L.; Dashtebani, F. Effect of silicon supplementation on wheat plants under salt stress. J. Plant Process Funct. 2016, 5, 1–12. [Google Scholar]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Hokmabadi, H.; Arzani, K.; Grierson, P.F. Growth, chemical composition, and carbon isotope discrimination of pistachio (Pistacia vera L.) rootstock seedlings in response to salinity. Aust. J. Agric. Res. 2005, 56, 135–144. [Google Scholar] [CrossRef]
- Zhang, W.; Xie, Z.; Wang, L.; Li, M.; Lang, D.; Zhang, X. Silicon alleviates salt and drought stress of Glycyrrhiza uralensis seedling by altering antioxidant metabolism and osmotic adjustment. J. Plant Res. 2017, 130, 611–624. [Google Scholar] [CrossRef]
- Gadallah, M.A.A. Effect of proline and glycinebetaine on Vicia faba responses to salt stress. Biol. Plant. 1999, 42, 249–257. [Google Scholar] [CrossRef]
- Raliya, R.; Tarafdar, J.C. ZnO nanoparticle biosynthesis and its effect on phosphorous-mobilizing enzyme secretion and gum contents in cluster bean (Cyamopsis tetragonoloba L.). Agric. Res. 2013, 2, 48–57. [Google Scholar] [CrossRef]
- Vaillant, N.; Monnet, F.; Hitmi, A.; Sallanon, H.; Coudret, A. Comparative study of responses in four Datura species to a zinc stress. Chemosphere 2005, 59, 1005–1013. [Google Scholar] [CrossRef]
- Pascual, M.B.; Echevarria, V.; Gonzalo, M.J.; Herna_ndez-Apaolaza, L. Silicon addition to soybean (Glycine max L.) plants alleviate zinc deficiency. Plant Physiol. Biochem. 2016, 108, 132–138. [Google Scholar] [CrossRef]
- Alzahrani, Y.; Kuşvuran, A.; Alharby, H.F.; Kuşvuran, S.; Rady, M.M. The defensive role of silicon in wheat against stress conditions induced by drought, salinity or cadmium. Ecotoxicol. Environ. Saf. 2018, 154, 187–196. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Ibrahim, M.; Farid, M.; Adrees, M.; Bharwana, S.A.; Ziar-ur-Rehman, M.; Qayyum, M.F.; Abbas, F. Mechanisms of silicon-mediated alleviation of drought and salt stress in plants: A review. Environ. Sci. Poll. Res. 2015, 22, 15416–15431. [Google Scholar] [CrossRef]
- Rahneshan, Z.; Nasibi, F.; Moghadam, A.H. Effects of salinity stress on some growth, physiological, biochemical parameters and nutrients in two pistachio (Pistacia vera L.) rootstocks. J. Plant Inter. 2018, 13, 73–82. [Google Scholar]
- Elsheery, N.I.; Sunoj, V.S.J.; Wen, Y.; Zhu, J.J.; Muralidharan, G.; Cao, K.F. Foliar application of nanoparticles mitigates the chilling effect on photosynthesis and photoprotection in sugarcane. Plant Physiol. Biochem. 2020, 149, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Bohnert, H.J.; Jensen, R.G. Strategies for engineering water stress tolerance in plants. Trends Biotechnol. 1996, 14, 89–97. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species, metabolism, oxidative stress and signal transduction. Ann. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Sen, A. Oxidative stress studies in plant tissue culture. In Antioxidant Enzyme; El-Missiry, M.A., Ed.; World’s Largest Science, Technology and Medicine Open Access Book; INTECH: Rijeka, Croatia, 2012; Chapter 3; pp. 59–88. Available online: http://dx.doi.org/10.5772/2895 (accessed on 17 December 2019).
- Ma, C.; Liu, H.; Guo, H.; Musante, C.; Coskun, S.H.; Nelson, B.C.; White, J.C.; Xing, B.; Dhankher, O.P. Defense mechanisms and nutrient displacement in Arabidopsis thaliana upon exposure to CeO2 and In2O3 nanoparticles. Environ. Sci. Nano 2016, 3, 1369–1379. [Google Scholar] [CrossRef]
- Garg, N.; Manchanda, G. ROS generation in plants: Boon or bane? Plant Biosyst. 2009, 143, 81–96. [Google Scholar] [CrossRef]
- Varjovi, M.B.; Valizadeh, M.; Bandehagh, A. Primary Antioxidant Enzymes and their Important Role in Oxidative Stress in Plants and Mammalian. Biol. Forum–Int. J. 2015, 7, 148–154. [Google Scholar]
- Srivastav, M.; Kishor, A.; Dahuja, A.; Sharma, R.R. Effect of paclobutrazol and salinity on ion leakage, proline content and activities of antioxidant enzymes in mango (Mangifera indica L.). Sci. Hort. 2010, 125, 785–788. [Google Scholar] [CrossRef]
- Vallee, B.L.; Auld, D.S. Zinc coordination, function, and structure of zinc enzymes and other proteins. Biochemistry 1990, 29, 5647–5659. [Google Scholar] [CrossRef]
- Fang, W.; Kao, C.H. Enhanced peroxidase activity in rice leaves in response to excess iron, copper and zinc. Plant Sci. 2000, 158, 71–76. [Google Scholar] [CrossRef]
- Marreiro, D.N.; Cruz, K.J.C.; Morais, J.B.S.; Beserra, J.B.; Severo, J.S.; Olveira, A.R.S. Zinc and oxidative stress: Current mechanisms. Antioxidants 2017, 6, 9. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. Zinc biochemistry: From a single zinc enzyme to a key element of life. Adv. Nutr. 2013, 4, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Clegg, M.S.; Hanna, L.A. Zinc deficiency-induced cell death. IUBMB Life 2005, 57, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Naeem, A.; Saifullah, M.; Zia-ur-Rehman, M.; Akhtar, T.; Zia, M.H.; Aslam, M. Silicon nutrition lowers cadmium content of wheat cultivars by regulating transpiration rate and activity of antioxidant enzymes. Environ. Pollut. 2018, 242, 126–135. [Google Scholar] [CrossRef]
- Liang, Y.; Hua, H.; Zhu, Y.G.; Zhang, J.; Cheng, C.; Ro€mheld, V. Importance of plant species and external silicon concentration to active silicon uptake and transport. New Phytol. 2006, 172, 63–72. [Google Scholar] [CrossRef]
- Crane, J.H.; Campbell, C.W. The Mango; University of Florida Press: Gainesville, FL, USA, 1994; p. 24. [Google Scholar]
- Attiah, H.H. A new eriophyid mite on mango from Egypt. Bull. Soc. Entomol. Egypt 1955, 39, 379–383. [Google Scholar]
- Singh, Z. Hormonal physiology of mango malformation—An overview. Acta Hort. 2000, 525, 229–236. [Google Scholar] [CrossRef]
- Bains, G.; Pant, R.C. Mango malformation: Etiology and preventive measures. Physiol. Mol. Biol. Plants 2003, 9, 41–61. [Google Scholar]
- Rymbai, H.; Rajesh, A.M. Mango malformation: A review. Life Sci. Leaf. 2011, 22, 1079–1095. [Google Scholar]
- Kumar, P.; Misra, A.K.; Modi, D.R. Current status of mango malformation in India. Asian J. Plant Sci. 2011, 10, 1–23. [Google Scholar] [CrossRef]
- Mari, G.G.; Guardiola, J.L.; Agusti, M.; Garcia-Mari, F.; Almela, V. The Regulation of fruit size in citrus by tree factors. In Proceeding of 21st International Horticultural Congress, Hamburg, Germany, 29 August –4 September 1982; Bnemann, G., Joseph, A., Eds.; Int. Soc. Hort. Sci.: Leuven, Belgium, 1982; p. 1363. [Google Scholar]
- Ginestar, C.; Castel, J.R. Response of young ‘Clementine’ citrus trees to water stress during different phenological periods. J. Hort. Sci. 1996, 71, 551–559. [Google Scholar] [CrossRef]
- Baldi, E.; Toselli, M.; Deudellar, D.; Tagliavini, M.; Marangoni, B. Foliar feeding of stone fruit trees. Inf. Agrar. 2004, 60, 43–46. [Google Scholar]
- Usenik, V.; Stampar, F. Effect of foliar application of zinc plus boron on sweet cherry fruit set and yield. Acta Hort. 2002, 594, 245–249. [Google Scholar] [CrossRef]
- Shi, X.; Zhang, C.; Wang, H.; Zhang, F. Effect of Si on the distribution of Cd in rice seedlings. Plant Soil 2005, 272, 53–60. [Google Scholar] [CrossRef]
- Rehman, M.Z.; Rizwan, M.; Ghafoor, A.; Naeem, A.; Ali, S.; Sabir, M.; Qayyum, M.F. Effect of inorganic amendments for in situ stabilization of cadmium in contaminated soil and its phyto-availability to wheat and rice under rotation. Environ. Sci. Pollut. Res. 2015, 22, 16897–16906. [Google Scholar] [CrossRef]
- Viehweger, K. How plants cope with heavy metals. Bot. Stud. 2014, 55, 35. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H. Role of nano-SiO2 in germination of tomato (Lycopersicum esculentum seeds Mill.). Saudi J. Biol. Sci. 2014, 21, 13–17. [Google Scholar] [CrossRef]
- Yamaji, N.; Chiba, Y.; Mitani-Ueno, N.; Ma, J.F. Functional characterization of a silicon transporter gene implicated in silicon distribution in barley. Plant Physiol. 2012, 160, 1491–1497. [Google Scholar] [CrossRef]
- Sharma, V.K.; Rajesh, T.; Preeti, C. Effect of N, P and their interaction on physicochemical parameters of guava (Psidium guajava) cv. L-49 under Malwa plateau conditions. Int. J. Sci. Res. Publ. 2014, 4, 1–4. [Google Scholar]
- Grierson, W. Maturity and grade standards. In Fresh Citrus Fruits; Wardowski, W.F., Miller, W.M., Hall, D.J., Grierson, W., Eds.; Florida Science Source, Inc.: Longboat Key, FL, USA, 2006; pp. 23–48. [Google Scholar]
- Davenport, T.L. Reproductive physiology of mango. Braz. J. Plant. Physiol. 2007, 19, 363–376. [Google Scholar] [CrossRef]
- Davenport, T.L. Reproductive physiology. In The Mango Botany Production and Uses, 2nd ed.; Litz, R.E., Ed.; CAB International: Wallingford, UK, 2009; pp. 97–169. [Google Scholar]
- Chen, J.Z.; Zhao, H.; Chen, J.L.; Zhao, H.Y. Advances in research on flower bud-differentiation in mango. South China Fruits 1999, 28, 34–35. [Google Scholar]
- Shaban, A.E.A. Vegetative growth cycles of some mango cultivars in relation to flowering and fruiting. World J. Agric. Sci. 2009, 5, 751–759. [Google Scholar]
- Smith-Ramirez, C.; Armesto, J.J.; Figueroa, J. Flowering, fruiting and seed germination in Chilean rain forest myrtaceae: Ecological and phylogenetic constraints. Plant Ecol. 1998, 136, 119–131. [Google Scholar] [CrossRef]
- Ramirez, F.; Davenport, T.L. Reproductive biology (physiology)—The case of mango. In Mango: Production and Processing Technology; Valavi, S.G., Rajmohan, K., Govil, J.N., Peter, K.V., Thottappilly, G., Eds.; Studium Press: New Delhi, India, 2012; Volume 1, pp. 56–81. [Google Scholar]
- Whily, A.W.; Rasmussem, T.S.; Saranah, J.B.; Wolstenholme, B.N. Effect of temperature on growth, dry matter production and starch accumulation in ten mango (Mangifera indica L) cultivars. J. Hort. Sci. Biotechnol. 1989, 64, 753–765. [Google Scholar] [CrossRef]
- Sthapit, B.R.; Ramanatha, R.V.; Sthapit, S.R. Tropical Fruit Tree Species and Climate Change; Bioversity Int.: New Delhi, India, 2012; 142p. [Google Scholar]
- Eiadthong, W.; Yonemori, K.; Kansaki, S.; Sugiura, A.; Utsunomiya, N.; Subhadrabandhu, S. Amplified fragments length polymorphism among Mangifera species in Thailand. J. Amer. Soc. Horti. Sci. 2000, 125, 160–164. [Google Scholar] [CrossRef]
- Upreti, K.K.; Reddy, Y.T.N.; Shivuprasad, S.R.; Bindu, G.V.; Jayaram, H.L.; Rajan, S. Hormonal changes in response to paclobutrazol induced early flowering in mango cv. Totapuri. Sci. Hort. 2013, 150, 414–418. [Google Scholar] [CrossRef]
- Sandip, M.; Makwana, A.N.; Barad, A.V.; Nawade, B.D. Physiology of flowering—The case of mango. Int. J. Appl. Res. 2015, 1, 1008–1012. [Google Scholar]
- Singh, R.N. Biennial bearing in fruit trees—Accent on mango and apples. Indian Counc. Agric. Res. Tech. Bull. 1971, 30, 47. [Google Scholar]
- Rastogi, A.; Tripathi, D.K.; Yadav, S.; Chauhan, D.K.; Živčák, M.; Ghorbanpour, M.; El-Sheery, N.I.; Brestic, M. Application of silicon nanoparticles in agriculture. 3 Biotech 2019, 9, 90. [Google Scholar] [CrossRef]


{kind=link}
| Soil Analysis | Water Analysis | ||
|---|---|---|---|
| Texture | Sandy loam | Ca+2 (mg/L) | 41 |
| Sand (Coarse + Fine) (%) | 63 | Mg+2 (mg/L) | 13 |
| Silt | 8 | Na+ (mg/L) | 75 |
| Clay | 29 | K+ (mg/L) | 18 |
| Organic matter | 1.23 | Cl− (mg/L) | 147 |
| N (mg/kg) | 41.67 | SO4−2 (mg/L) | 72.6 |
| P (mg/kg) | 16.33 | HCO−3 (mg/L) | 3.2 |
| K (mg/kg) | 524 | CO−3 (mg/L) | - |
| Na (mg/kg) | 426 | Na adsorption ratio | 13 |
| EC (dSm−1) | 3.67 | EC (dSm−1) | 0.96 |
| pH | 8.1 | pH | 7.9 |
| Treatment (T) | Leaf Area | ||
|---|---|---|---|
| Season (S) | Mean | ||
| 2017 | 2018 | ||
| 0 nZnO + 0 nSi | 39.8 | 38.5 | 39.15 |
| 0 nZnO + 150 nSi | 45.5 | 46.3 | 45.9 |
| 0 nZnO + 300 nSi | 36.7 | 38.6 | 37.65 |
| 50 nZnO + 0 nSi | 48.3 | 49.7 | 49 |
| 50 nZnO + 150 nSi | 52.7 | 54.7 | 53.7 |
| 50 nZnO + 300 nSi | 49.3 | 51.8 | 50.55 |
| 100 nZnO + 0 nSi | 59.6 | 60.5 | 60.05 |
| 100 nZnO + 150 nSi | 63.4 | 65.8 | 64.6 |
| 100 nZnO + 300 nSi | 52.3 | 53.7 | 53 |
| 150 nZnO + 0 nSi | 60.1 | 61.6 | 60.85 |
| 150 nZnO + 150 nSi | 61.2 | 62.7 | 61.95 |
| 150 nZnO + 300 nSi | 43.3 | 45.6 | 44.45 |
| Mean | 51.02 | 52.46 | |
| LSD (P ≤ 0.05) | S = 0.62 | T = 1.534 | S × T = 2.16 |
| Treatment (T) | Nitrogen (N) | Phosphorus (P) | Potassium (K) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Season (S) | Mean | Season (S) | Mean | Season (S) | Mean | ||||
| 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | ||||
| 0 nZnO + 0 nSi | 1.31 | 1.34 | 1.325 | 0.136 | 0.137 | 0.1365 | 0.74 | 0.74 | 0.74 |
| 0 nZnO + 150 nSi | 1.48 | 1.41 | 1.445 | 0.159 | 0.164 | 0.1615 | 0.86 | 0.82 | 0.84 |
| 0 nZnO + 300 nSi | 1.44 | 1.37 | 1.405 | 0.146 | 0.148 | 0.147 | 0.77 | 0.77 | 0.77 |
| 50 nZnO + 0 nSi | 1.52 | 1.58 | 1.55 | 0.173 | 0.189 | 0.181 | 0.87 | 0.88 | 0.875 |
| 50 nZnO + 150 nSi | 1.67 | 1.62 | 1.645 | 0.182 | 0.197 | 0.1895 | 0.99 | 0.93 | 0.96 |
| 50 nZnO + 300 nSi | 1.58 | 1.58 | 1.58 | 0.166 | 0.174 | 0.17 | 0.83 | 0.89 | 0.86 |
| 100 nZnO + 0 nSi | 1.78 | 1.72 | 1.75 | 0.204 | 0.217 | 0.2105 | 0.98 | 1.07 | 1.025 |
| 100 nZnO + 150 nSi | 1.92 | 1.97 | 1.945 | 0.247 | 0.268 | 0.2575 | 1.17 | 1.21 | 1.19 |
| 100 nZnO + 300 nSi | 1.62 | 1.66 | 1.64 | 0.193 | 0.197 | 0.195 | 0.95 | 0.98 | 0.965 |
| 150 nZnO + 0 nSi | 1.74 | 1.68 | 1.71 | 0.184 | 0.193 | 0.1885 | 0.94 | 0.95 | 0.945 |
| 150 nZnO + 150 nSi | 1.81 | 1.86 | 1.835 | 0.226 | 0.232 | 0.229 | 1.03 | 1.05 | 1.04 |
| 150 nZnO + 300 nSi | 1.59 | 1.62 | 1.605 | 0.177 | 0.185 | 0.181 | 0.88 | 0.91 | 0.895 |
| Mean | 1.62 | 1.62 | 0.183 | 0.192 | 0.917 | 0.933 | |||
| LSD (P ≤ 0.05) | S = 0.35 | T = 0.09 | S × T = 0.12 | S = 0.05 | T = 0.011 | S × T= 0.015 | S = 0.54 | T = 0.13 | S × T= 0.187 |
| Treatment (T) | Total Carbohydrates | Total Sugars | Proline | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Season (S) | Mean | Season (S) | Mean | Season (S) | Mean | ||||
| 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | ||||
| 0 nZnO + 0 nSi | 39.6 | 39.6 | 39.6 | 5.3 | 6.3 | 5.8 | 5.4 | 5.4 | 5.4 |
| 0 nZnO + 150 nSi | 39.8 | 40.2 | 40 | 5.8 | 5.8 | 5.8 | 5.9 | 5.9 | 5.9 |
| 0 nZnO + 300 nSi | 39.1 | 39.3 | 39.2 | 6.1 | 5.3 | 5.7 | 5.8 | 5.7 | 5.75 |
| 50 nZnO + 0 nSi | 40.4 | 41.2 | 40.8 | 6.3 | 6.8 | 6.55 | 5.9 | 6.1 | 6 |
| 50 nZnO + 150 nSi | 43.2 | 45.8 | 44.5 | 8.1 | 8.4 | 8.25 | 6.7 | 6.9 | 6.8 |
| 50 nZnO + 300 nSi | 42.7 | 43.5 | 43.1 | 7.2 | 7.5 | 7.35 | 6.2 | 5.9 | 6.05 |
| 100 nZnO + 0 nSi | 44.4 | 45.1 | 44.75 | 8.3 | 8.4 | 8.35 | 6.5 | 6.7 | 6.6 |
| 100 nZnO + 150 nSi | 46.6 | 47.2 | 46.9 | 9.6 | 9.7 | 9.65 | 7.3 | 7.4 | 7.35 |
| 100 nZnO + 300 nSi | 43.8 | 45.1 | 44.45 | 7.6 | 7.7 | 7.65 | 6.1 | 6.4 | 6.25 |
| 150 nZnO + 0 nSi | 42.8 | 41.3 | 42.05 | 7.9 | 7.9 | 7.9 | 7 | 7.1 | 7.05 |
| 150 nZnO + 150 nSi | 45.8 | 45.5 | 45.65 | 8.8 | 9.2 | 9 | 7.1 | 7.3 | 7.2 |
| 150 nZnO + 300 nSi | 40.5 | 40.6 | 40.55 | 5.7 | 6.2 | 5.95 | 6 | 6.2 | 6.1 |
| Mean | 42.39 | 42.87 | 7.22 | 7.43 | 6.32 | 6.42 | |||
| LSD (P ≤ 0.05) | S = 0.86 | T = 2.1 | S × T = 2.9 | S = 0.48 | T = 1.18 | S × T = 1.7 | S = 0.46 | T = 1.14 | S × T = 1.6 |
| Treatment (T) | SOD | POX | CAT | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Season (S) | Mean | Season (S) | Mean | Season (S) | Mean | ||||
| 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | ||||
| 0 nZnO + 0 nSi | 83.5 | 84.9 | 84.2 | 74.6 | 74.6 | 74.6 | 16.8 | 16.8 | 16.8 |
| 0 nZnO + 150 nSi | 72.4 | 73.7 | 73.05 | 75.2 | 76.4 | 75.8 | 17.1 | 17.8 | 17.45 |
| 0 nZnO + 300 nSi | 63.6 | 43.9 | 53.75 | 46.7 | 47.5 | 47.1 | 11.6 | 12.3 | 11.95 |
| 50 nZnO + 0 nSi | 81.2 | 81.8 | 81.5 | 76.3 | 77.3 | 76.8 | 18.4 | 19.7 | 19.05 |
| 50 nZnO + 150 nSi | 86.2 | 87.8 | 87 | 76.8 | 77.8 | 77.3 | 20.4 | 21.6 | 21 |
| 50 nZnO + 300 nSi | 74.3 | 52.5 | 63.4 | 56.8 | 57.4 | 57.1 | 15.8 | 16.2 | 16 |
| 100 nZnO + 0 nSi | 91.2 | 94.6 | 92.9 | 78.4 | 78.7 | 78.55 | 15.7 | 16.2 | 15.95 |
| 100 nZnO + 150 nSi | 97.5 | 98.2 | 97.85 | 84.6 | 86.6 | 85.6 | 24.2 | 25.7 | 24.95 |
| 100 nZnO + 300 nSi | 78.3 | 56.3 | 67.3 | 63.4 | 65.2 | 64.3 | 16.2 | 16.6 | 16.4 |
| 150 nZnO + 0 nSi | 55.5 | 59.5 | 57.5 | 52.5 | 53.8 | 53.15 | 12.5 | 13.4 | 12.95 |
| 150 nZnO + 150 nSi | 58.6 | 49.7 | 54.15 | 74.8 | 75.1 | 74.95 | 14.6 | 15.4 | 15 |
| 150 nZnO + 300 nSi | 52.7 | 32.7 | 42.7 | 42.3 | 42.8 | 42.55 | 11.2 | 11.5 | 11.35 |
| Mean | 74.58 | 67.97 | 66.87 | 67.77 | 16.21 | 16.93 | |||
| LSD (P ≤ 0.05) | S = 0.8 | T = 1.95 | S × T = 2.7 | S = 0.48 | T = 1.17 | S × T = 1.66 | S = 0.48 | T = 1.19 | S × T = 1.7 |
| Treatment (T) | Floral Malformation | Fruit Number/Tree | Fruit Weight/Tree | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Season (S) | Mean | Season (S) | Mean | Season (S) | Mean | ||||
| 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | ||||
| 0 nZnO + 0 nSi | 39.6 | 39.6 | 39.6 | 71 | 127 | 99 | 21.5 | 37.6 | 29.5 |
| 0 nZnO + 150 nSi | 31.3 | 30 | 30.65 | 159 | 173 | 166 | 41.3 | 41.8 | 41.55 |
| 0 nZnO + 300 nSi | 32.9 | 31.2 | 32.05 | 127 | 138 | 132.5 | 40.6 | 41.6 | 41.1 |
| 50 nZnO + 0 nSi | 29.4 | 27.5 | 28.45 | 168 | 176 | 172 | 41.8 | 44.2 | 43 |
| 50 nZnO + 150 nSi | 24.2 | 22.8 | 23.5 | 184 | 206 | 195 | 47.2 | 51.2 | 49.2 |
| 50 nZnO + 300 nSi | 27.8 | 25.5 | 26.65 | 154 | 167 | 160.5 | 42.5 | 43.5 | 43 |
| 100 nZnO + 0 nSi | 20.8 | 20.5 | 20.65 | 186 | 202 | 194 | 46.7 | 48.9 | 47.8 |
| 100 nZnO + 150 nSi | 15.2 | 14.4 | 14.8 | 217 | 232 | 224.5 | 55.4 | 63.4 | 59.4 |
| 100 nZnO + 300 nSi | 25.9 | 23.4 | 24.65 | 172 | 189 | 180.5 | 46.4 | 47.6 | 47 |
| 150 nZnO + 0 nSi | 23.5 | 21.5 | 22.5 | 178 | 186 | 182 | 45.3 | 46.3 | 45.8 |
| 150 nZnO + 150 nSi | 18.5 | 18 | 18.25 | 202 | 207 | 204.5 | 51.8 | 52.8 | 52.3 |
| 150 nZnO + 300 nSi | 31.8 | 30.5 | 31.15 | 162 | 179 | 170.5 | 37.2 | 39.5 | 38.3 |
| Mean | 26.74 | 25.41 | 165.00 | 181.83 | 43.14 | 46.53 | |||
| LSD (P ≤ 0.05) | S = 0.4 | T = 1.02 | S × T = 1.44 | S = 5.4 | T = 13.4 | S × T = 18.9 | S = 0.75 | T = 1.84 | S × T = 2.6 |
| Treatment (T) | Fruit Weight | Fruit Length | Fruit Width | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Season (S) | Mean | Season (S) | Mean | Season (S) | Mean | ||||
| 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | ||||
| 0 nZnO + 0 nSi | 246.4 | 247.4 | 246.9 | 9.5 | 9.7 | 9.6 | 6.6 | 6.9 | 6.75 |
| 0 nZnO + 150 nSi | 269.4 | 283.1 | 276.25 | 10.2 | 10.4 | 10.3 | 7.1 | 7.3 | 7.2 |
| 0 nZnO + 300 nSi | 261.5 | 271.5 | 266.5 | 9.7 | 9.8 | 9.75 | 6.9 | 7.1 | 7 |
| 50 nZnO + 0 nSi | 287.5 | 286.8 | 287.15 | 9.9 | 10.5 | 10.2 | 7.3 | 7.4 | 7.35 |
| 50 nZnO + 150 nSi | 318.7 | 316.4 | 317.55 | 10.6 | 10.8 | 10.7 | 7.8 | 7.9 | 7.85 |
| 50 nZnO + 300 nSi | 279.4 | 256.5 | 267.95 | 10.1 | 10.3 | 10.2 | 7.2 | 7.3 | 7.25 |
| 100 nZnO + 0 nSi | 285.7 | 299.7 | 292.7 | 10.4 | 10.7 | 10.55 | 7.6 | 7.8 | 7.7 |
| 100 nZnO + 150 nSi | 336.6 | 348.6 | 342.6 | 11.2 | 11.6 | 11.4 | 8.1 | 8.3 | 8.2 |
| 100 nZnO + 300 nSi | 244.6 | 267.4 | 256 | 10.3 | 10.8 | 10.55 | 7.1 | 7.4 | 7.25 |
| 150 nZnO + 0 nSi | 288.5 | 298.7 | 293.6 | 10.1 | 10.6 | 10.35 | 6.9 | 7.2 | 7.05 |
| 150 nZnO + 150 nSi | 308.3 | 304.5 | 306.4 | 10.5 | 10.8 | 10.65 | 7.6 | 7.8 | 7.7 |
| 150 nZnO + 300 nSi | 239.3 | 231.7 | 235.5 | 9.7 | 9.9 | 9.8 | 6.8 | 6.8 | 6.8 |
| Mean | 280.49 | 284.36 | 10.18 | 10.49 | 7.25 | 7.43 | |||
| LSD (P ≤ 0.05) | S = 0.75 | T = 1.02 | S × T = 2.6 | S = 0.47 | T = 1.17 | S × T = 1.6 | S = 0.3 | T = 0.75 | S × T = 1.06 |
| Treatment (T) | TSS | Total Acidity | Total Sugars | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Season (S) | Mean | Season (S) | Mean | Season (S) | Mean | ||||
| 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | ||||
| 0 nZnO + 0 nSi | 21.7 | 21.9 | 21.8 | 1.42 | 1.42 | 1.42 | 14.4 | 14.8 | 14.6 |
| 0 nZnO + 150 nSi | 24.6 | 25.3 | 24.95 | 1.29 | 1.26 | 1.275 | 15.5 | 15.8 | 15.65 |
| 0 nZnO + 300 nSi | 24.3 | 24.7 | 24.5 | 1.37 | 1.35 | 1.36 | 15 | 15.3 | 15.15 |
| 50 nZnO + 0 nSi | 24.8 | 25.1 | 24.95 | 1.28 | 1.27 | 1.275 | 15.8 | 16.4 | 16.1 |
| 50 nZnO + 150 nSi | 25.6 | 25.8 | 25.7 | 1.15 | 1.16 | 1.155 | 17.1 | 17.7 | 17.4 |
| 50 nZnO + 300 nSi | 22.6 | 23.2 | 22.9 | 1.48 | 1.38 | 1.43 | 16.2 | 16.8 | 16.5 |
| 100 nZnO + 0 nSi | 25.8 | 25.9 | 25.85 | 1.22 | 1.21 | 1.215 | 17.2 | 17.7 | 17.45 |
| 100 nZnO + 150 nSi | 26.2 | 26.7 | 26.45 | 1.12 | 1.12 | 1.12 | 17.4 | 17.6 | 17.5 |
| 100 nZnO + 300 nSi | 22.2 | 22.7 | 22.45 | 1.41 | 1.41 | 1.41 | 15.7 | 16.1 | 15.9 |
| 150 nZnO + 0 nSi | 23.1 | 24.4 | 23.75 | 1.26 | 1.29 | 1.275 | 16.3 | 16.7 | 16.5 |
| 150 nZnO + 150 nSi | 23.3 | 23.7 | 23.5 | 1.29 | 1.24 | 1.265 | 16.3 | 16.8 | 16.55 |
| 150 nZnO + 300 nSi | 22.1 | 23.4 | 22.75 | 1.38 | 1.33 | 1.355 | 14.9 | 15.7 | 15.3 |
| Mean | 32.86 | 24.40 | 1.31 | 1.29 | 15.98 | 16.45 | |||
| LSD (P ≤ 0.05) | S = 0.042 | T = 1.04 | S × T= 1.47 | S = 0.047 | T = 0.17 | S × T= 1.6 | S = 0.42 | T = 1.05 | S × T= 1.48 |
| Season | 2014/2015 | 2015/2016 | 2016/2017 | 2017/2018 | 2018/2019 |
|---|---|---|---|---|---|
| Yield (kg/tree) | 19.9 | 35.5 | 21.5 | 39.5 | 19.6 |
| Alternate bearing | Off | On | Off | On | Off |
| Temperature (°C) | 21.2 | 22 | 21.3 | 22.2 | 22.6 |
| Humidity (%) | 67.5 | 67.7 | 67.5 | 65.8 | 64.3 |
| Cloud (%) | 13.7 | 16.2 | 14.2 | 16.8 | 18.1 |
| Rainfall (mm/month) | 6.9 | 11.1 | 4.8 | 4.4 | 17.4 |
| Rainfall (days/month) | 4.7 | 6.5 | 2.4 | 3.6 | 7.6 |
| Wind speed (km/h) | 15.5 | 15.8 | 15.5 | 16.2 | 15.1 |
| Gust speed (km/h) | 22.2 | 22.9 | 22.0 | 22.9 | 23.2 |
| Sun (hours/month) | 296.8 | 295.9 | 298.9 | 291.9 | 284.9 |
| Sun (days/month) | 24.3 | 23.4 | 26.2 | 26.1 | 22.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elsheery, N.I.; Helaly, M.N.; El-Hoseiny, H.M.; Alam-Eldein, S.M. Zinc Oxide and Silicone Nanoparticles to Improve the Resistance Mechanism and Annual Productivity of Salt-Stressed Mango Trees. Agronomy 2020, 10, 558. https://doi.org/10.3390/agronomy10040558
Elsheery NI, Helaly MN, El-Hoseiny HM, Alam-Eldein SM. Zinc Oxide and Silicone Nanoparticles to Improve the Resistance Mechanism and Annual Productivity of Salt-Stressed Mango Trees. Agronomy. 2020; 10(4):558. https://doi.org/10.3390/agronomy10040558
Chicago/Turabian StyleElsheery, Nabil I., Mohamed N. Helaly, Hanan M. El-Hoseiny, and Shamel M. Alam-Eldein. 2020. "Zinc Oxide and Silicone Nanoparticles to Improve the Resistance Mechanism and Annual Productivity of Salt-Stressed Mango Trees" Agronomy 10, no. 4: 558. https://doi.org/10.3390/agronomy10040558
APA StyleElsheery, N. I., Helaly, M. N., El-Hoseiny, H. M., & Alam-Eldein, S. M. (2020). Zinc Oxide and Silicone Nanoparticles to Improve the Resistance Mechanism and Annual Productivity of Salt-Stressed Mango Trees. Agronomy, 10(4), 558. https://doi.org/10.3390/agronomy10040558