Rootstock x Environment Interactions on Nitrogen-Use Efficiency in Grafted Tomato Plants at Different Phenological Stages


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Grafting Method
2.2. Experimental Conditions
2.2.1. Vegetative Stage
2.2.2. Reproductive Stage
2.3. Analytical Determinations
2.3.1. Chlorophyll Content
2.3.2. Biomass and Tissue N Determinations
2.4. NUE, NUpE, and NUtE Calculations
2.5. Relative Growth Rate (RGR) Calculations
2.6. Statistical Analysis
3. Results
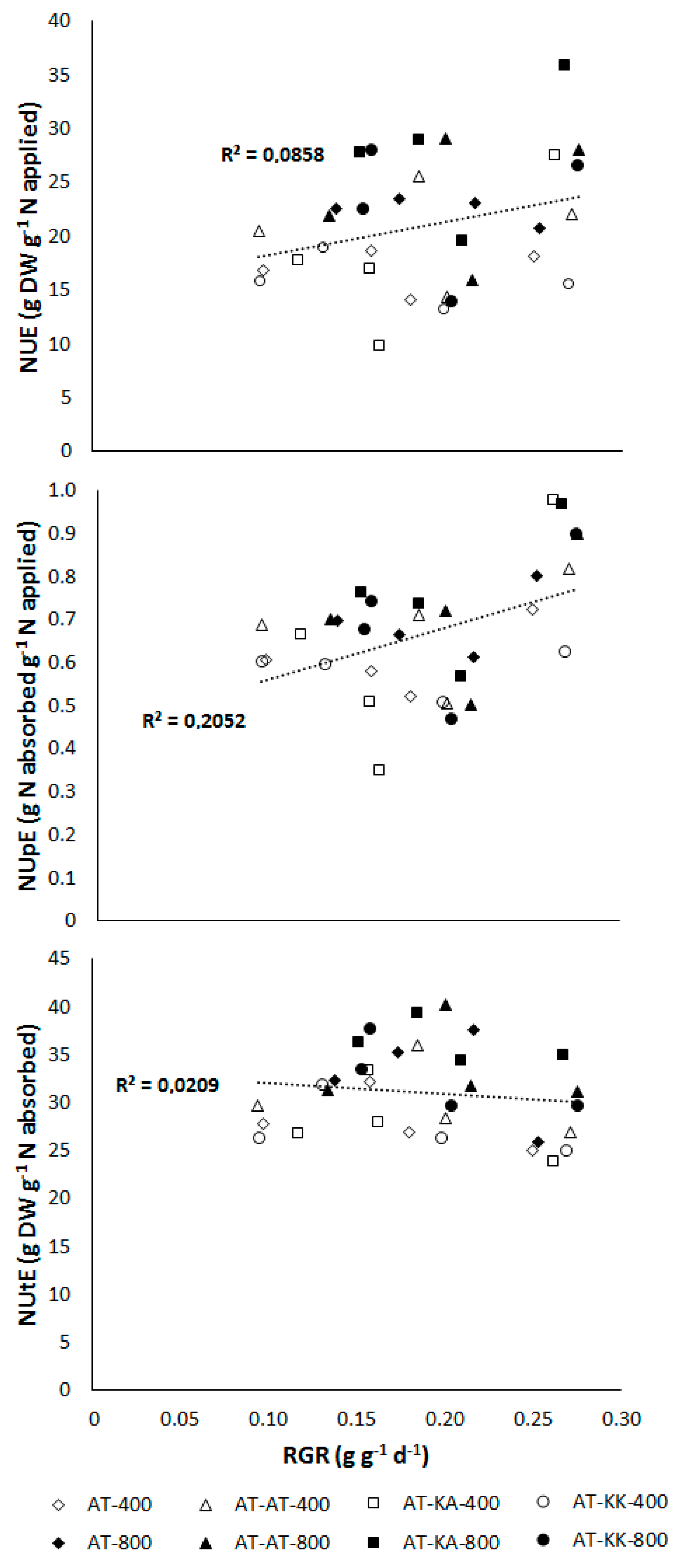
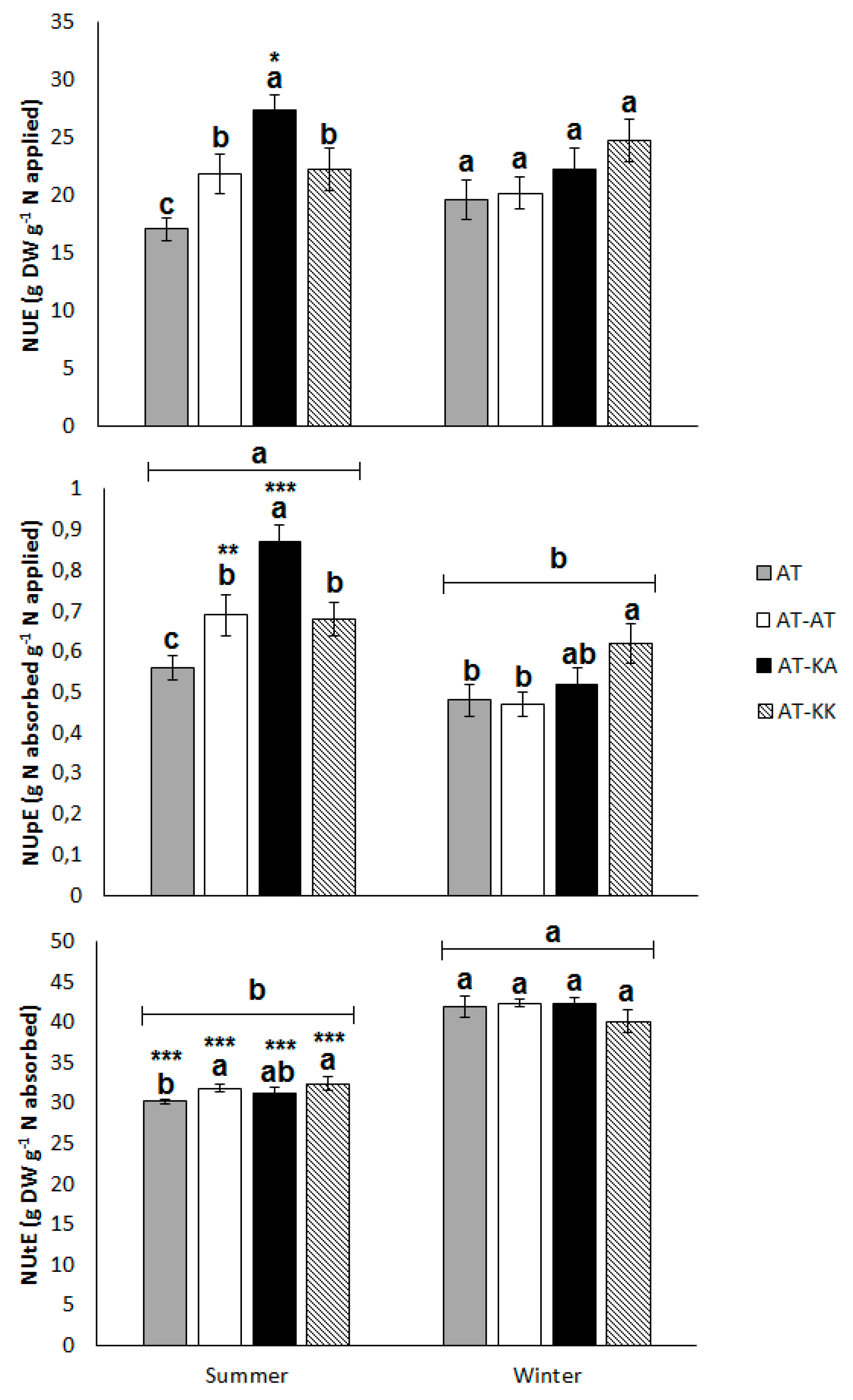
3.1. Vegetative Stage
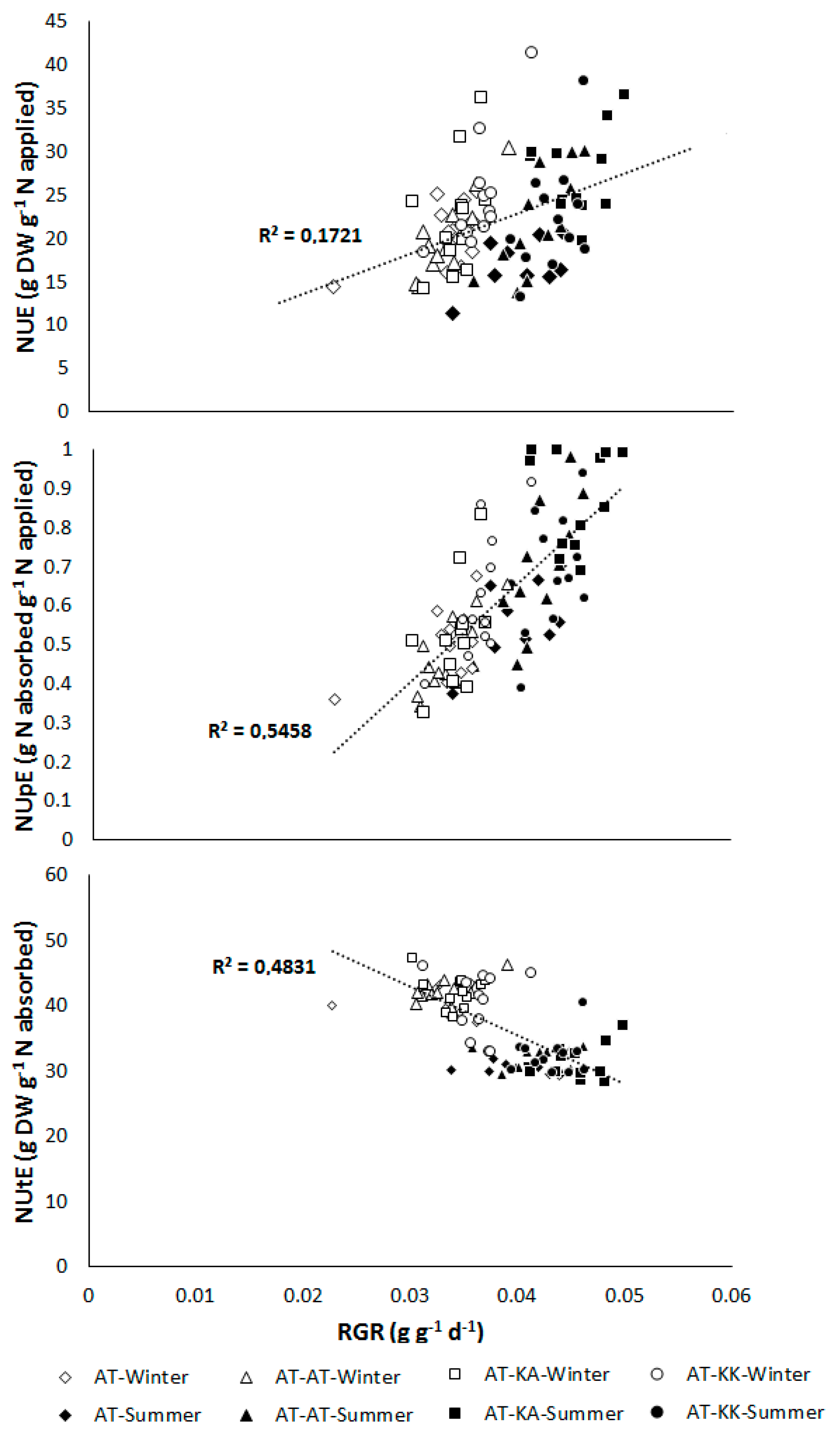
3.2. Reproductive Stage
4. Discussion
4.1. NUE and Developmental Stage
4.2. Rootstock Effects on NUE
4.3. Rootstock x Environment Interaction
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Min, J.; Zhao, X.; Shi, W.; Xing, G.; Zhu, Z. Nitrogen balance and loss in a greenhouse vegetable system in southeastern China. Pedosphere 2011, 21, 464–472. [Google Scholar] [CrossRef]
- Zhu, J.H.; Li, X.L.; Christie, P.; Li, J.L. Environmental implications of low nitrogen use efficiency in excessively fertilized hot pepper (Capsicum frutescens L.) cropping systems. Agric. Ecosyst. Environ. 2005, 111, 70–80. [Google Scholar] [CrossRef]
- He, F.; Chen, Q.; Jiang, R.; Chen, X.; Zhang, F. Yield and nitrogen balance of greenhouse tomato (Lycopersicum esculentum Mill.) with conventional and site-specific nitrogen management in northern China. Nutr. Cycl. Agroecosyst. 2007, 77, 1–14. [Google Scholar] [CrossRef]
- Yang, S.; Wang, Y.; Liu, R.; Xing, L.; Yang, Z. Improved crop yield and reduced nitrate nitrogen leaching with straw return in a rice-wheat rotation of Ningxia irrigation district. Sci. Rep. 2018, 8, 9458. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. The influence of drip irrigation or subirrigation on zucchini squash grown in closed-loop substrate culture with high and low nutrient solution concentrations. Hortscience 2009, 44, 306–311. [Google Scholar] [CrossRef]
- Sun, Y.; Hu, K.; Zhang, K.; Jiang, L.; Xu, Y. Simulation of nitrogen fate for greenhouse cucumber grown under different water and fertilizer management using the Rotate_N model. Agric. Water Manag. 2012, 112, 21–32. [Google Scholar] [CrossRef]
- Li, Y.; Li, J.; Gao, L.; Tian, Y. Irrigation has more influence than fertilization on leaching water quality and the potential environmental risk in excessively fertilized vegetable soils. PLoS ONE 2018, 13, e0204570. [Google Scholar] [CrossRef]
- Benincasa, P.; Guiducci, M.; Tei, F. The nitrogen use efficiency: Meaning and sources of variation—Case studies on three vegetable crops in central Italy. Horttechnology 2011, 21, 266–273. [Google Scholar] [CrossRef]
- Borgognone, D.; Colla, G.; Rouphael, Y.; Cardarelli, M.; Rea, E.; Schwarz, D. Effect of nitrogen form and nutrient solution pH on growth and mineral composition of self-grafted and grafted tomatoes. Sci. Hortic. 2013, 149, 61–69. [Google Scholar] [CrossRef]
- De Pascale, S.; Rouphael, Y.; Gallardo, M.; Thompson, R.B. Water and fertilization management of vegetables: State of art and future challenges. Eur. J. Hortic. Sci. 2018, 83, 306–318. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Colla, G. Vegetable grafting: A toolbox for securing yield stability under multiple stress conditions. Front. Plant Sci. 2018, 8, 2255. [Google Scholar] [CrossRef]
- Moll, R.H.; Kamprath, E.J.; Jackson, W.A. Analysis and interpretation of factors which contribute to the efficiency of nitrogen utilization. Agron. J. 1982, 74, 562–564. [Google Scholar] [CrossRef]
- Zhang, X.; Davidson, E.A.; Mauzerall, D.L.; Searchinger, T.D.; Dumas, P.; Shen, Y. Managing nitrogen for sustainable development. Nature 2015, 528, 51–59. [Google Scholar] [CrossRef]
- King, S.R.; Davis, A.R.; Zhang, X.; Crosby, K. Genetic breeding and selection of rootstocks for Solanaceae and Cucurbitaceae. Sci. Hortic. 2010, 127, 106–111. [Google Scholar] [CrossRef]
- Leonardi, C.; Giuffrida, F. Variation of plant growth and macronutrient uptake in grafted tomatoes and eggplants on three different rootstocks. Eur. J. Hort. Sci. 2006, 71, 97–101. [Google Scholar]
- Djidonou, D.; Zhao, X.; Simonne, E.H.; Koch, K.E. Yield, water-, and nitrogen-use efficiency in field-grown, grafted tomatoes. Hortscience 2013, 48, 485–492. [Google Scholar] [CrossRef]
- Colla, G.; Cardona, C.M.; Cardarelli, M.; Rouphael, Y. Improving nitrogen use efficiency in melon by grafting. Hortscience 2010, 45, 559–565. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Mirabelli, C.; Cardarelli, M. Nitrogen-use efficiency traits of mini-watermelon in response to grafting and nitrogen-fertilization doses. J. Plant Nutr. Soil Sci. 2011, 174, 933–941. [Google Scholar] [CrossRef]
- Tewolde, F.T.; Lu, N.; Shiina, K.; Maruo, T.; Takagaki, M.; Kozai, T.; Yamori, W. Nighttime supplemental LED inter-lighting improves growth and yield of single-truss tomatoes by enhancing photosynthesis in both winter and summer. Front. Plant Sci. 2016, 7, 448. [Google Scholar] [CrossRef]
- Tapia, M.L.; Gutiérrez, V. Distribution pattern of dry weight, nitrogen, phosphorus, and potassium through tomato ontogenesis. J. Plant Nutr. 1997, 20, 783–791. [Google Scholar] [CrossRef]
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: San Diego, CA, USA, 2012. [Google Scholar]
- Wells, C.; Eissenstat, D. Beyond the roots of young seedlings: The influence of age and order on fine root physiology. J. Plant Growth Regul. 2002, 21, 324–334. [Google Scholar] [CrossRef]
- Mae, T. Physiological nitrogen efficiency in rice: Nitrogen utilization, photosynthesis, and yield potential. Plant Soil 1997, 196, 201–210. [Google Scholar] [CrossRef]
- Baligar, V.C.; Fageria, N.K.; He, Z.L. Nutrient use efficiency in plants. Commun. Soil Sci. Plan. 2001, 32, 921–950. [Google Scholar] [CrossRef]
- Glass, A.D.M. Nitrogen use efficiency of crop plants: Physiological constraints upon nitrogen absorption. Crit. Rev. Plant Sci. 2003, 22, 453–470. [Google Scholar] [CrossRef]
- Parry, C.; Blonquist, J.M.; Bugbee, B. In situ measurement of leaf chlorophyll concentration: Analysis of the optical/absolute relationship. Plant Cell Environ. 2014, 37, 2508–2520. [Google Scholar] [CrossRef]
- Nario, A.; Pino, I.; Zapata, F.; Albornoz, M.P.; Baherle, P. Nitrogen (15N) fertiliser use efficiency in peach (Prunus persica L.) cv. Goldencrest trees in Chile. Sci. Hortic. 2003, 97, 279–287. [Google Scholar] [CrossRef]
- Fageria, N.K.; Baligar, V.C. Enhancing nitrogen use efficiency in crop plants. Adv. Agron. 2005, 88, 97–185. [Google Scholar]
- Albornoz, F.; Lieth, J.H. N, P, K and S uptake response to various levels of CO2 assimilation and growth rate in lettuce. J. Plant Nutr. 2017, 40, 773–783. [Google Scholar] [CrossRef]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat Version 2014, Grupo InfoStat, 2014; Universidad Nacional de Córdoba: Córdoba, Argentina, 2014. [Google Scholar]
- Albornoz, F.; Gebauer, M.; Ponce, C.; Cabeza, R. LeNRT1.1 improves nitrate uptake in grafted tomato plants under high nitrogen demand. Int. J. Mol. Sci. 2018, 19, 3921. [Google Scholar] [CrossRef]
- Wu, P.; Tao, Q.N. Genotypic response and selection pressure on nitrogen-use efficiency in rice under different nitrogen regimes. J. Plant Nutr. 1995, 18, 487–500. [Google Scholar] [CrossRef]
- Gallais, A.; Coque, M. Genetic variation and selection for nitrogen use efficiency in maize: A synthesis. Maydica 2005, 50, 531. [Google Scholar]
- Hassan, M.S.; Khair, A.; Haque, M.M.; Azad, A.K.; Hamid, A. Genotypic variation in traditional rice varieties for chlorophyll content, SPAD value and nitrogen use efficiency. Bangladesh J. Agric. Res. 2009, 34, 505–515. [Google Scholar] [CrossRef]
- Liu, Y.; Qi, H.; Bai, C.; Qi, M.; Xu, C.; Hao, J.; Li, Y.; Li, T. Grafting helps improve photosynthesis and carbohydrate metabolism in leaves of muskmelon. Int. J. Biol. Sci. 2011, 7, 1161–1170. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Goto, R.; De Miguel, A.; Marsal, J.; Gorbe, E.; Calatayud, A. Effect of different rootstocks on growth, chlorophyll a fluorescence and mineral composition of two grafted scions of tomato. J. Plant Nutr. 2013, 36, 825–835. [Google Scholar] [CrossRef]
- García-Bañuelos, M.; Sanchez, E.; Gardea-Bejar, A.; Muñoz-Marquez, E.; Soto-Parra, J.; Ojeda-Barrios, D. Nitrogen use efficiency and yield in response graft bell pepper cultivars. Emir. J. Food Agric. 2017, 29, 420–428. [Google Scholar] [CrossRef]
- Evans, J.R.; Poorter, H. Photosynthetic acclimation of plants to growth irradiance: The relative importance of specific leaf area and nitrogen partitioning in maximizing carbon gain. Plant Cell Environ. 2001, 24, 755–767. [Google Scholar] [CrossRef]
- Kitajima, K.; Hogan, K.P. Increases of chlorophyll a/b ratios during acclimation of tropical woody seedlings to nitrogen limitation and high light. Plant Cell Environ. 2003, 26, 857–865. [Google Scholar] [CrossRef]
- Richards, D.; Goubran, F.H.; Collins, K.E. Root-shoot equilibria in fruiting tomato plants. Ann. Bot. 1979, 43, 401–404. [Google Scholar] [CrossRef]
- Robinson, N.L.; Hewitt, J.D.; Bennett, A.B. Sink metabolism in tomato fruit. I. Developmental changes in carbohydrate metabolizing enzymes. Plant Physiol. 1988, 87, 727–730. [Google Scholar] [CrossRef]
- Yelle, S.; Chetelat, R.T.; Dorais, M.; DeVerna, J.W.; Bennett, A.B. Sink metabolism in tomato fruit. IV. Genetic and biochemical analysis of sucrose accumulation. Plant Physiol. 1991, 95, 1026–1035. [Google Scholar] [CrossRef]
- Moco, S.; Bino, R.J.; Vorst, O.; Verhoeven, H.A.; De Groot, J.; Van Beek, T.A.; Vervoort, J.; De Vos, C.H.R. A liquid chromatography-mass spectrometry-based metabolome database for tomato. Plant Physiol. 2006, 141, 1205–1218. [Google Scholar] [CrossRef] [PubMed]
- Greef, J.M. Productivity of maize (Zea mays L.) in relation to morphological and physiological characteristics under varying amounts of nitrogen supply. J. Agron. Crop Sci. 1994, 172, 317–326. [Google Scholar] [CrossRef]
- Zhang, H.; Li, W.; Adams, H.D.; Wang, A.; Wu, J.; Jin, C.; Guan, D.; Yuan, F. Responses of woody plant functional traits to nitrogen addition: A meta-analysis of leaf economics, gas exchange, and hydraulic traits. Front. Plant Sci. 2018, 9, 683. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa-Sakurai, J.; Hayashi, H.; Murai-Hatano, M. Nitrogen availability affects hydraulic conductivity of rice roots, possibly through changes in aquaporin gene expression. Plant Soil 2014, 379, 289–300. [Google Scholar] [CrossRef]
- Orieux, C.; Demarest, G.; Decau, M.; Beauclair, P.; Bataille, M.; Le Deunff, E. Changes in 15NO3- availability and transpiration rate are associated with a rapid diurnal adjustment of anion contents as well as 15N and water fluxes between the roots and shoots. Front. Plant Sci. 2018, 9, 1751. [Google Scholar] [CrossRef] [PubMed]
- Djidonou, D.; Zhao, X.; Brecht, J.K.; Cordasco, K.M. Influence of interspecific hybrid rootstocks on tomato growth, nutrient accumulation, yield, and fruit composition under greenhouse conditions. Horttechonology 2017, 27, 868–877. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Wang, L.; Jiao, Y.; Chen, C.; Zhao, L.; Mei, M.; Yu, Y.; Bie, Z.; Huang, Y. Pumpkin rootstock improves nitrogen use efficiency of watermelon scion by enhancing nutrient uptake, cytokinin content, and expression of nitrate reductase genes. Plant Growth Regul. 2017, 82, 233–246. [Google Scholar] [CrossRef]
- Martínez-Ballestero, M.; Alcaraz-López, C.; Muries, B.; Mota-Cadenas, C.; Carvajal, M. Physiological aspects of rootstock-scion interactions. Sci. Hortic. 2010, 127, 112–118. [Google Scholar] [CrossRef]
- Albacete, A.; Martínez-Andujar, C.; Martínez-Pérez, A.; Thompson, A.J.; Dodd, I.C.; Pérez-Alfocea, F. Unraveling rootstock x scion interactions to improve food security. J. Exp. Bot. 2015, 66, 2211–2226. [Google Scholar] [CrossRef]
- Albacete, A.; Cantero-Navarro, E.; Balibrea, M.E.; Großkinsky, D.K.; De la Cruz González, M.; Martínez-Andújar, C.; Smigocki, A.C.; Roitsch, T.; Pérez-Alfocea, F. Hormonal and metabolic regulation of tomato fruit sink activity and yield under salinity. J. Exp. Bot. 2014, 65, 6081–6095. [Google Scholar] [CrossRef]
- Cohen, S.; Nair, A. The effect of three rootstocks on water use, canopy conductance and hydraulic parameters of apple trees and predicting canopy from hydraulic conductance. Plant Cell Environ. 2002, 25, 17–28. [Google Scholar] [CrossRef]
- Tombesi, S.; Johnson, R.S.; Day, K.R.; DeJong, T.M. Relationship between xylem vessel characteristics, calculated axial hydraulic conductance and size-controlling capacity of peach rootstocks. Ann. Bot. 2010, 105, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Sory, A.; Nieto-Angel, R.; Rodríguez-Pérez, J.E.; Barrientos-Priego, A.F.; Ibañez-Castillo, L.A.; Romabchik, E.; Núñez-Colín, C.A. Variación anatómica del xilema en tallo de cultivares de tomate injertados en un tipo criollo. Rev. Chapingo Ser. Hortic. 2010, 16, 67–76. [Google Scholar]
- Du, Y.; Cao, H.; Liu, S.; Gu, X.; Cao, Y. Response of yield, quality, water and nitrogen use efficiency of tomato to different levels of water and nitrogen under drip irrigation in northwestern China. J. Integr. Agric. 2017, 16, 1153–1161. [Google Scholar] [CrossRef]
- Gent, M.P.N.; Short, M.R. Effect on yield and quality of a simple system to recycle nutrient solution to greenhouse tomato. Hortscience 2012, 47, 1641–1645. [Google Scholar] [CrossRef]
- Massa, D.; Incrocci, L.; Maggini, R.; Carmassi, G.; Campiotti, C.A.; Pardossi, A. Strategies to decrease water drainage and nitrate emission from soilless culture of greenhouse tomato. Agric. Water Manag. 2010, 97, 971–980. [Google Scholar] [CrossRef]
- Signore, A.; Serio, F.; Santamaria, P. A Targeted management of the nutrient solution in a soilless tomato crop according to plant needs. Front. Plant Sci. 2016, 7, 391. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology, 2nd ed.; Springer: New York, NY, USA, 2008. [Google Scholar]
- Ntatsi, G.; Savvas, D.; Klaring, H.; Schwarz, D. Growth, yield, and metabolic responses of temperature-stressed tomato to grafting onto rootstocks differing in cold tolerance. J. Am. Soc. Hort. Sci. 2014, 139, 230–243. [Google Scholar] [CrossRef]
- Sui, N.; Li, M.; Liu, X.Y.; Wang, N.; Fang, W.; Meng, Q.W. Response of xanthophyll cycle and chloroplastic antioxidant enzymes to chilling stress in tomato over-expressing glycerol-3-phosphate acyltransferase gene. Photosynthetica 2007, 45, 447–454. [Google Scholar] [CrossRef]
- Wu, K.; Dechorgnat, J.; Patrit, O.; Krapp, A.; Fagard, M.; Daniel-Vedele, F. Characterization of the nrt2.6 gene in Arabidopsis thaliana: A link with plant response to biotic and abiotic stress. PLoS ONE 2012, 7, e42491. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Scion | Rootstock | Vigor |
|---|---|---|---|
| AT | Attiya | None | |
| AT-AT | Attiya | Attiya | |
| AT-KA | Attiya | Kaiser | High |
| AT-KK | Attiya | King Kong | Medium |
| Nutrient | Nutrient Concentration (mM) | ||
|---|---|---|---|
| Transplant to First Cluster Flowering | First Cluster Flowering to the Third Cluster Flowering | Third Cluster Flowering to Fifth Cluster Harvest | |
| Nitrogen | 10.0 | 12.5 | 14.0 |
| Phosphorus | 0.5 | 0.85 | 0.85 |
| Potassium | 3.0 | 3.5 | 4.0 |
| Calcium | 3.0 | 4.0 | 4.5 |
| Magnesium | 2.0 | 2.0 | 2.5 |
| Sulphur | 2.0 | 2.0 | 2.5 |
| Irrigation volume (L pot−1 d−1) | 1.5 | 1.8 | 1.8 |
| Drainage percentage | 14 ± 2 | 22 ± 4 | 25 ± 3 |
| Treatment | 400 µmol PAR m−2 s−1 | 800 µmol PAR m−2 s−1 |
|---|---|---|
| mg N plant−1 | ||
| AT | 86.99 ± 10.21 a | 113.95 ± 7.38 a |
| AT-AT | 97.54 ± 12.35 a | 115.94 ± 10.52 a |
| AT-KA | 95.50 ± 8.07 a | 121.75 ± 9.11 a |
| AT-KK | 82.16 ± 10.06 a | 105.63 ± 8.71 a |
| Treatment | Summer | Winter |
|---|---|---|
| g N plant−1 | ||
| AT | 11.47 ± 0.68 b | 10.12 ± 0.93 a |
| AT-AT | 13.92 ± 1.03 b | 10.05 ± 0.61 a |
| AT-KA | 19.08 ± 1.15 a | 10.24 ± 1.27 a |
| AT-KK | 14.27 ± 1.16 b | 13.09 ± 0.96 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albornoz, F.; Nario, A.; Saavedra, M.; Videla, X. Rootstock x Environment Interactions on Nitrogen-Use Efficiency in Grafted Tomato Plants at Different Phenological Stages. Agronomy 2020, 10, 350. https://doi.org/10.3390/agronomy10030350
Albornoz F, Nario A, Saavedra M, Videla X. Rootstock x Environment Interactions on Nitrogen-Use Efficiency in Grafted Tomato Plants at Different Phenological Stages. Agronomy. 2020; 10(3):350. https://doi.org/10.3390/agronomy10030350
Chicago/Turabian StyleAlbornoz, Francisco, Adriana Nario, Macarena Saavedra, and Ximena Videla. 2020. "Rootstock x Environment Interactions on Nitrogen-Use Efficiency in Grafted Tomato Plants at Different Phenological Stages" Agronomy 10, no. 3: 350. https://doi.org/10.3390/agronomy10030350
APA StyleAlbornoz, F., Nario, A., Saavedra, M., & Videla, X. (2020). Rootstock x Environment Interactions on Nitrogen-Use Efficiency in Grafted Tomato Plants at Different Phenological Stages. Agronomy, 10(3), 350. https://doi.org/10.3390/agronomy10030350