Bacillus thuringiensis Effect on the Vegetative Development of Cotton Plants and the Biocontrol of Spodoptera frugiperda

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. First Stage
2.1.1. Bt Strains and Growth Conditions
2.1.2. Plant Material
2.1.3. Inoculation with Bt Strains and Conduct of the Experiment
2.1.4. Evaluation of Plant Growth
2.1.5. Statistical Analysis
2.1.6. Autoradiography of Cotton Plants Colonized by Bt Strains
2.1.7. Bioassay with Spodoptera Frugiperda on Plants
2.1.8. Observation of Bt on Cotton Seeds by Scanning Electron Microscopy
2.2. Second Stage
2.2.1. Evaluation of Plant Growth
2.2.2. “In Vitro” Bioassay with S. frugiperda
3. Results
3.1. First Stage
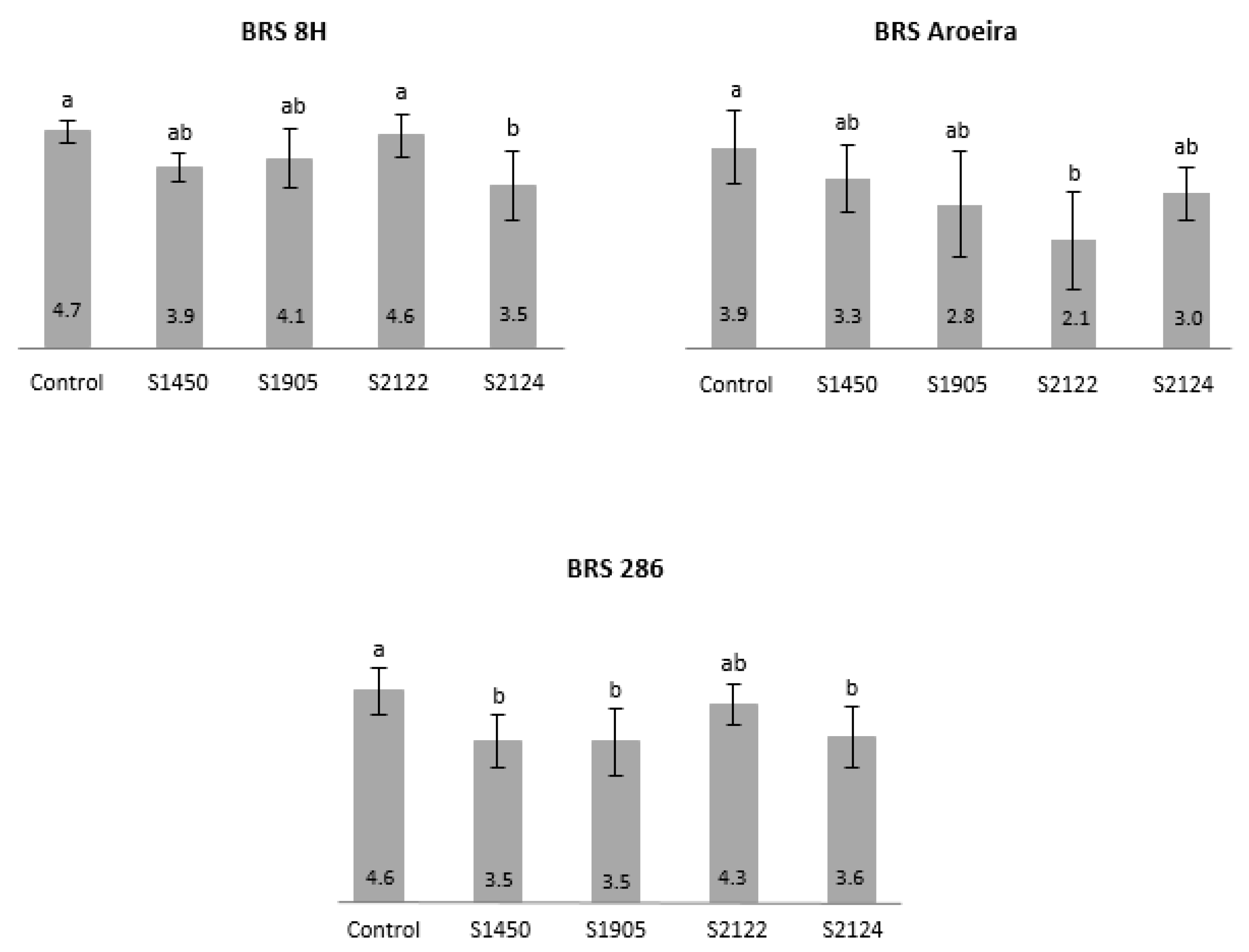
3.1.1. Analysis on Growth of Cotton Cultivars Inoculated with Bt Strains
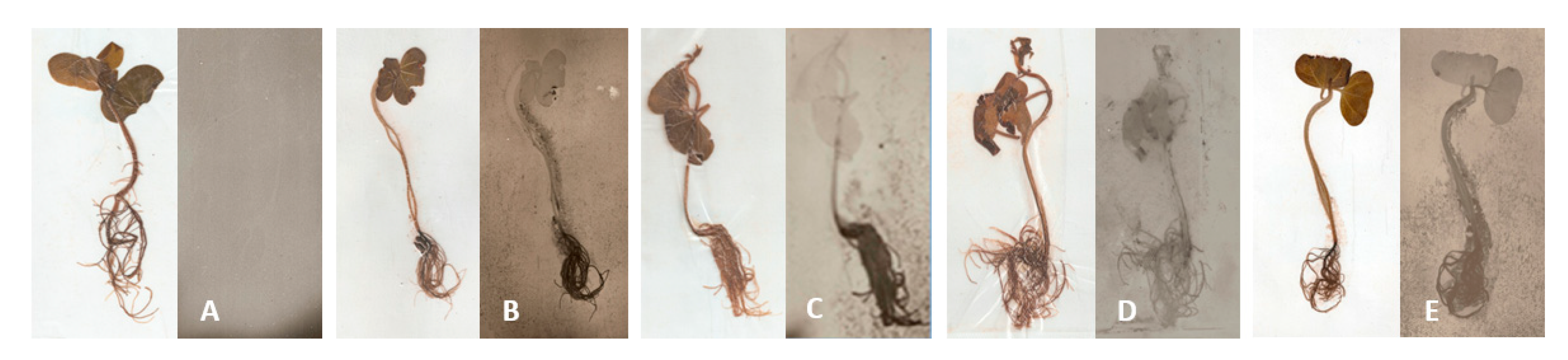
3.1.2. Autoradiography of Cotton Plants Colonized by Bt Strains
3.1.3. Bioassay with S. frugiperda on Plants
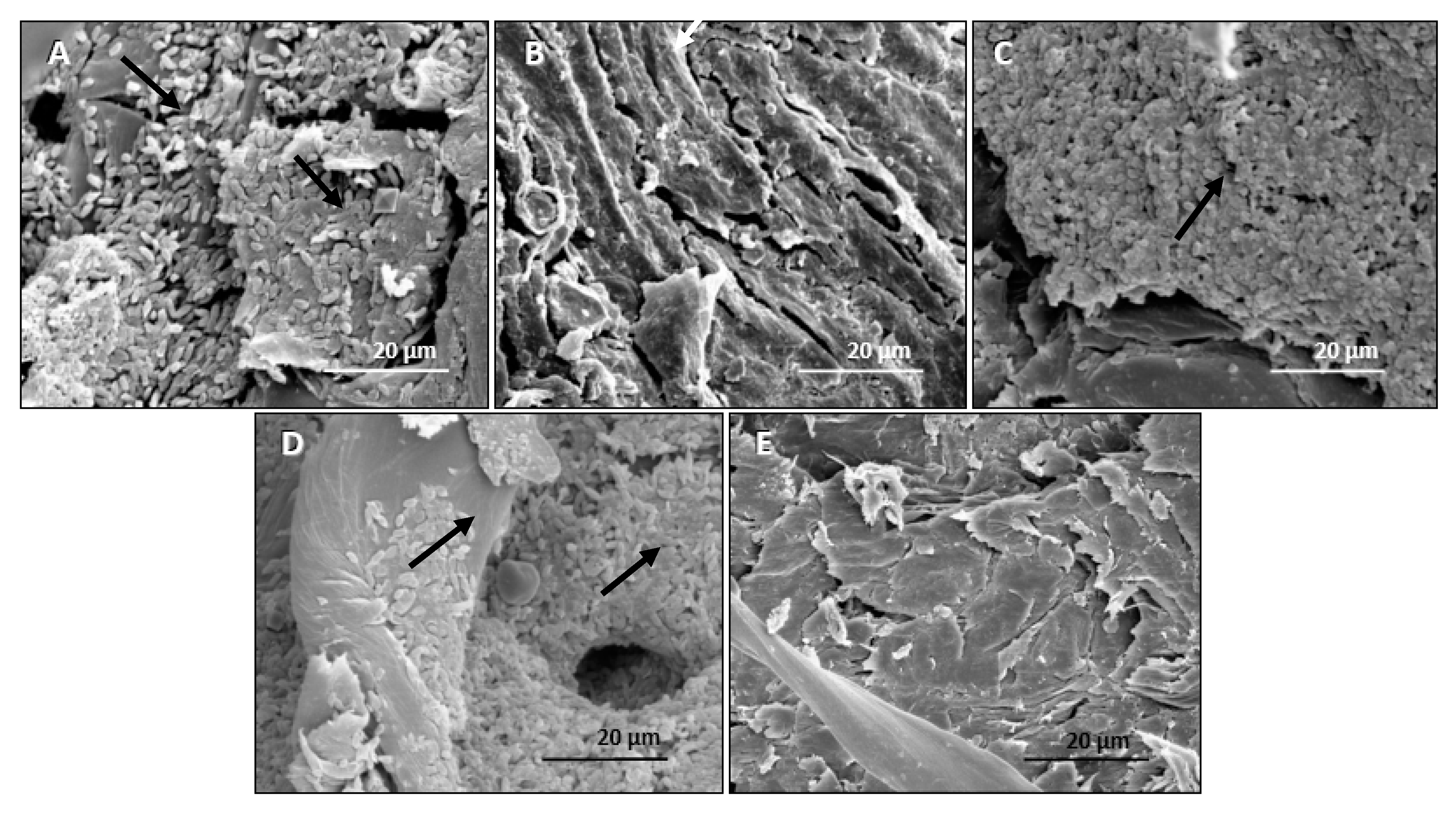
3.1.4. Observation of the Presence of Bt on Seeds of a Cotton Cultivar by Scanning Electron Microscopy
3.2. Second Stage
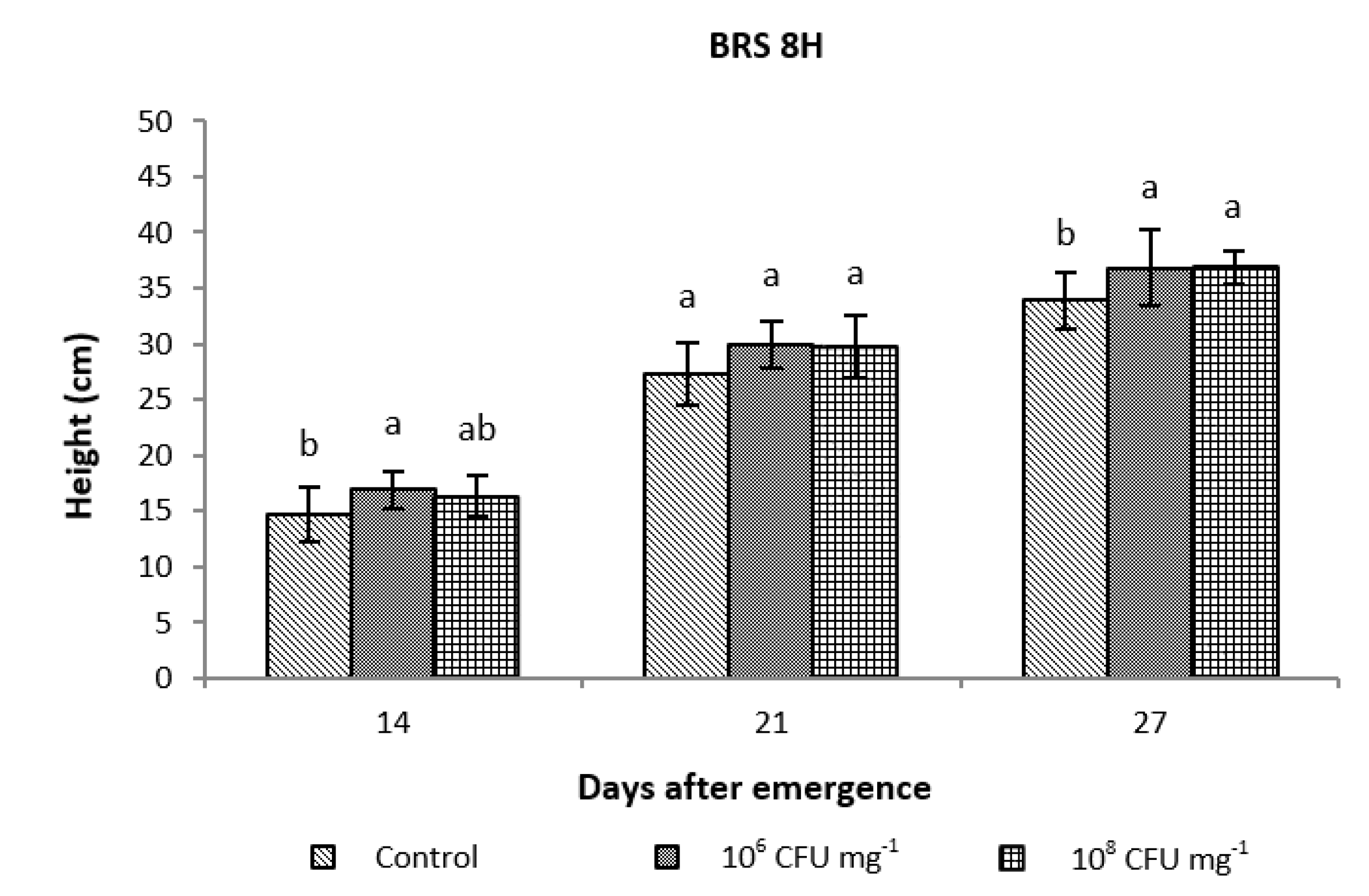
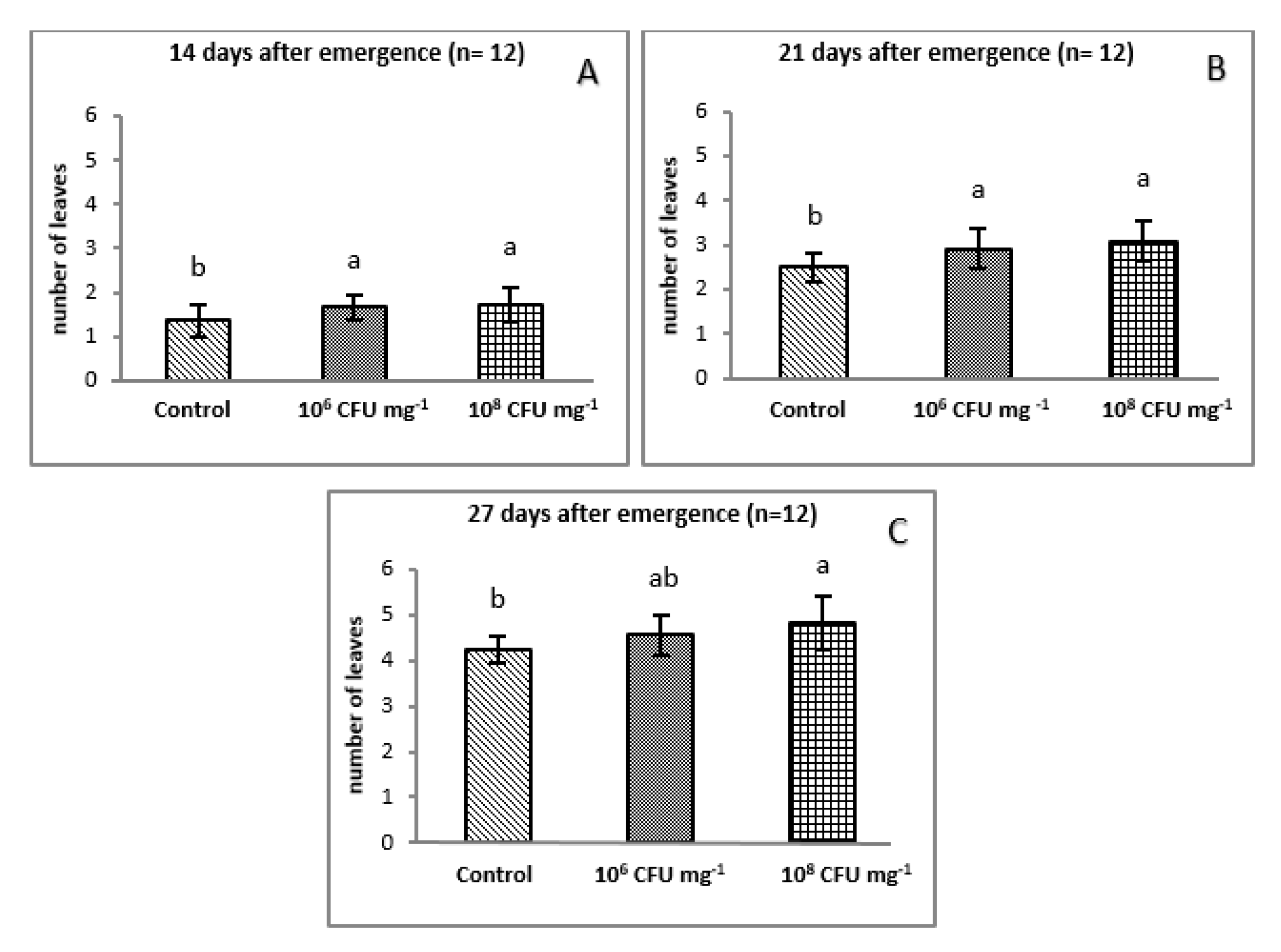
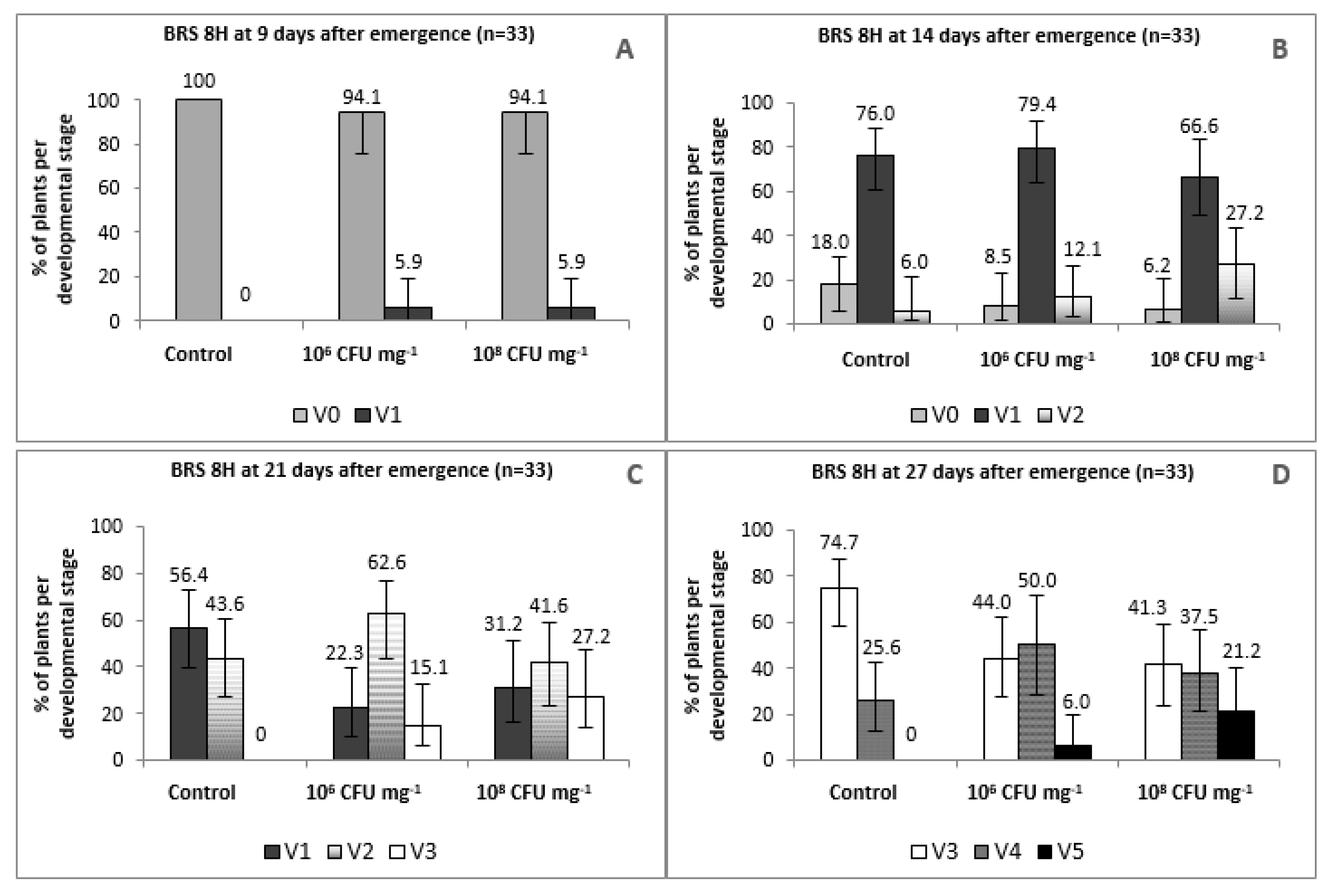
3.2.1. Evaluation of Plant Growth
3.2.2. “In Vitro” Bioassay with S. frugiperda
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fontes, E.M.; Ramalho, F.D.; Underwood, E.; Barroso, P.A.; Simon, M.F.; Sujii, E.R.; Pires, C.S.; Beltrão, N.; Lucena, W.A.; Freire, E.C. The cotton agricultural context in Brazil. In Environmental Risk Assessment of Genetically Modified Organisms: Methodologies for Assessing Bt Cotton in Brazil, 1st ed.; Hilbeck, A., Andow, D.A., Fontes, E., Eds.; CABI Publishing: Oxfordshire, UK, 2006; Volume 2, 373p. [Google Scholar]
- Santos, W.D. Monitoring and control of cotton pests. In Cotton Culture; POTAFOS: Piracicaba, São Paulo, Brazil, 1999; pp. 133–179. [Google Scholar]
- Becker, W.; Hopper, N.; Mcmichael, B.; Jividen, G. Seed applied plant growth regulators effects on cotton germination, emergence and growth. In Proceedings of the Beltwide Cotton Conference, Orlando, FL, USA, 3–7 January 1999; National Cotton Council: Memphis, TN, USA, 1999; pp. 625–627. [Google Scholar]
- Dos Santos, K.B.; Neves, P.; Meneguim, A.M.; dos Santos, R.B.; dos Santos, W.J.; Boas, G.V.; Dumas, V.; Martins, E.; Praça, L.B.; Queiroz, P.; et al. Selection and characterization of the Bacillus thuringiensis strains toxic to Spodoptera eridania (Cramer), Spodoptera cosmioides (Walker) and Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae). Biol. Control. 2009, 50, 157–163. [Google Scholar] [CrossRef]
- Miranda, J.E. Integrated cotton pest management in the Brazilian Cerrado. In Technical Release n° 131; Embrapa Cotton: Campina Grande, Paraíba, Brazil, 2010. [Google Scholar]
- Santos, W.J.; Santos, K.B.; Santos, R.B. Occurrence, description and habits of Spodoptera spp. in Brazil cotton. In Brazilian Cotton Congress, 4; Embrapa CPAO: Goiânia, Goiás, Brazil, 2003. [Google Scholar]
- Praça, L.B.; Gomes, A.C.M.; Cabral, G.; Martins, E.; Sujii, E.H.; Monnerat, R.G. Endophytic Colonization by Brazilian Strains of Bacillus thuringiensis on Cabbage Seedlings Grown in Vitro. Bt Res. 2012, 3, 11–19. [Google Scholar]
- Monnerat, R.; Santos, R.C.; Barros, P.C.; Batista, A.C.; Berry, C. Isolation and characterization of strains Bacillus thuringiensis endophytic in cotton plants. In Technical Release n° 98; Embrapa Genetic Resourses and Biotechnology: Brasília, Federal District, Brazil, 2003. [Google Scholar]
- Monnerat, R.G.; Soares, C.M.S.; Gomes, A.C.M.; Jones, G.; Martins, E.; Praça, L.; Berry, C. Translocation and insecticidal activity of Bacillus thuringiensis bacteria living inside of plants. Microb. Biotech. 2009, 2, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Praça, L.B. Interactions between Bacillus Thuringiensis Strains and Cabbage Hybrids to Control Plutella Xylostella and Promote Plant Growth. Ph.D. Thesis, Brasília University, Federal District, Brazil, 2012. [Google Scholar]
- Monnerat, R.G.; Batista, A.C.; Medeiros, P.T.; Martins, E.S.; Melatti, V.M.; Praça, L.B.; Dumas, V.F.; Morinaga, C.; Demo, C.; Gomes, A.C.M.; et al. Screening of Brazilian Bacillus thuringiensis isolates active against Spodoptera frugiperda, Plutella xylostella and Anticarsia gemmatalis. Biol. Control. 2007, 41, 291–295. [Google Scholar] [CrossRef]
- Westergren, G.; Krasse, B. Evaluation of a micromethod for determination of Streptococcus mutans and Lactobacillus infection. J. Clin. Microbiol. 1978, 7, 82–83. [Google Scholar]
- Maguire, J.D. Speed of germination aid in selection and evaluation for seedling emergence and vigor. Crop. Sci. 1962, 2, 176–177. [Google Scholar] [CrossRef]
- Kuo, J.; Fox, E.; Macdonald, S. SigmaStat: Statistical software form working scientists. In Users Manual; Jandel Scienti Wc: San Francisco, CA, USA, 1992. [Google Scholar]
- Murashige, T.; Skoog, F.K. A revised medium for rapid growth and bioassays with tabacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Schmidt, F.G.V.; Monnerat, R.; Borges, M.; Carvalho, R. Insect breeding methodology for the evaluation of entomopathogenic and semiochemical agents. In Technical Release n° 11; Embrapa Genetic Resourses and Biotechnology: Brasília, Federal District, Brazil, 2001. [Google Scholar]
- Marur, C.J.; Ruano, O. A reference system for determination of developmental stages of upland cotton. Rev. Bras. Oleag. Fib. 2001, 5, 313–317. [Google Scholar]
- Silva, F.A.S. Assistat 7.7 (Beta); Federal University of Campina Grande: Paraíba, Brazil, 2013. [Google Scholar]
- Hardoim, P.R.; van Overbeek, L.S.; van Elsas, J.D. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microb. 2008, 16, 463–471. [Google Scholar] [CrossRef]
- Davitt, A.J.; Chen, C.; Rudgers, J.A. Understanding context-dependency in plan-microbes symbiosis: The influence of abiotic and biotic contexts on host fitness and the rate of symbiont transmission. Environ. Experim. Bot. 2011, 71, 137–145. [Google Scholar] [CrossRef]
- Assumpção, L.D.C.; Lacava, P.T.; Dias, A.C.F.; Azevedo, J.L.D.; Menten, J.O.M. Diversity and biotechnological potential of endophytic bacterial community of soybean seeds. Pesq. Agropec. Bras. 2009, 44, 503–510. [Google Scholar] [CrossRef]
- Santos, M.H.; Mariano, R.D.; Camara, T.R.; Andrade, A.D.; Willadino, L.; Lima, G.P. Bactérias promotoras de crescimento no desenvolvimento de Heliconia psittacorum Lf. Hoehnea 2005, 32, 1–8. [Google Scholar]
- Schmidt, M.A.; Souza, E.M.; Baura, V.; Wassem, R.; Yates, M.G.; Pedrosa, F.O.; Monteiro, R.A. Evidence for the endophytic colonization of Phaseolus vulgaris (common bean) roots by the diazotroph Herbaspirillum seropedicae. Braz. J. Med. Biol. Res. 2011, 44, 182–185. [Google Scholar] [CrossRef]
- Pelzer, G.Q.; Halfeld-Vieira, B.A.; Nechet, K.D.; Souza, G.R.; Zilli, J.É.; Perin, L. Control mechanisms of southern blight and growth promotion on tomato mediated by rhizobacteria. Trop. P. Pathol. 2011, 36, 95–103. [Google Scholar] [CrossRef]
- Harthmann, O.E.; Mógor, Á.F.; Wordell Filho, J.A.; Luz, W.C. Rhizobacteria on the production and yield of onion. Ciência Rural. 2010, 40, 432–435. [Google Scholar] [CrossRef]
- Adams, P.D.; Kloepper, J.W. Effect of host genotype on indigenous bacterial endophytes of cotton (Gossypium hirsutum L.). Plant Soil 2002, 240, 181–189. [Google Scholar] [CrossRef]
- Stirling, G.R. Mass production and release of biological control agents. In Biological Control of Plant Parasitic Nematodes—Progress, Problems and Prospects; Red Wood Press: Mekshom, UK, 1991; pp. 12–165. [Google Scholar]
- Farooq, M.; Barsa, S.M.; Wahid, A. Priming of field-sown rice seed enhances germination, seedling establishment, allometry and yield. Plant Growth Regul. 2006, 49, 285–294. [Google Scholar] [CrossRef]
- Sturz, A.V.; Nowak, J. Endophytic communities of rhizobacteria and strategies required to create yield enhancing associations with crops. Appl. Soil Ecol. 2000, 15, 153–190. [Google Scholar] [CrossRef]
- Gomes, A.; Mariano, R.L.; Silveira, E.B.; Mesquita, J.C. Isolamento, seleção de bactérias e efeito da utilização de Bacillus spp. na produção de mudas orgânicas de alface. Hort. Bras. 2003, 21, 699–703. [Google Scholar] [CrossRef]
- Barretti, P.B.; Romeiro, R.D.; Mizubuti, E.S.; Souza, J.T. Screening of endophytic bacteria isolated from tomato plants as potencial biocontrol agents and growth promotion. Ciên Agrotec. 2009, 33, 2038–2044. [Google Scholar] [CrossRef]
- Baldotto, L.E.B.; Baldotto, M.A.; Olivares, F.L.; Viana, A.P.; Bressan-Smith, R. Selection of growth-promoting bacteria for pineapple ’Vitória’ during acclimatization. Rev. Bras. Ciên. Solo 2010, 34, 349–360. [Google Scholar] [CrossRef]
- Compant, S.; Mitter, B.; Colli-Mull, J.G.; Gangl, H.; Sessitsch, A. Endophytes of grapevine flowers, berries, and seeds: Identification of cultivable bacteria, comparison with other plant parts, and visualization of niches of colonization. Microb. Ecol. 2011, 62, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Döbereiner, J.; Reis, V.M.; Paula, M.A.; Olivares, F. Endophytic diazotophic in sugar cane, cereal and tuber plants. In New Horizons in Nitrogen Fixation; Palacios, R., Mora, J., Newton, W.F., Eds.; Kluver Academic Publishers: Dordrecht, The Netherlands, 1993; pp. 671–676. [Google Scholar]
- Hallmann, J.; Quadt-Hallmann, A.; Mahaffee, W.F.; Kloepper, J.W. Bacterial endophytes in agricultural crops. Canad. J. Microb. 1997, 43, 895–914. [Google Scholar] [CrossRef]
- Reis, V.M.; Olivares, F.L. Pathways of Penetration and Infection of Plants by Bacteria. In Embrapa Agrobiology—Documents 216; Baldani, V., Xavier, G., Bahia, M., Eds.; Brazilian Agricultural Research Corporation: Seropédica, Rio de Janeiro, Brazil, 2006. [Google Scholar]
- Pieterse, C.M.; Dicke, M. Plant interactions with microbes and insects: From molecular mechanisms to ecology. Trend. Plant Sci. 2007, 12, 564–569. [Google Scholar] [CrossRef] [PubMed]
- Shavit, R.; Ofek-Lalzar, M.; Burdman, S.; Morin, S. Inoculation of tomato plants with rhizobacteria enhances the performance of the phloem-feeding insect Bemisia tabaci. Front. Plant Sci. 2013, 4, 306. [Google Scholar] [CrossRef]
- Oliveira, A.L.M.; Urquiaga, S.; Baldani, J.I. Processes and mechanisms involved in the influence of microorganisms on plant growth. In Embrapa Agrobiology—Documents 161; Reis, V., Felix, D., Bahia, M., Eds.; Brazilian Agricultural Research Corporation: Seropédica, Rio de Janeiro, Brazil, 2003; pp. 15–17. [Google Scholar]
- Prabhakar, A.; Bishop, A.H. Effect of Bacillus thuringiensis naturally colonising Brassica campestris var. chinensis leaves on neonate larvae of Pieris brassicae. J. Invertebr. Pathol. 2009, 100, 193–194. [Google Scholar] [CrossRef]
- Van Oosten, V.R.; Bodenhausen, N.; Reymond, P.; Van Pelt, J.A.; Van Loon, L.C.; Dicke, M.; Pieterse, C.M. Differential effectiveness of microbially induced resistance against herbivorous insects in Arabidopsis. Mol. Plant Microbe Interact. 2008, 21, 919–930. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | BRS 8H | BRS Aroeira | BRS 286 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| ------------------------------ Dry Weight (g) ------------------------------ | |||||||||
| AP | RP | T | AP | RP | T | AP | PR | T | |
| Control | 2.19 ± 0.32ab | 1.50 ± 0.37ab | 3.70 ± 0.65ab | 1.91 ± 0.13a | 1.47 ± 0.51a | 3.38 ± 0.62a | 1.83 ± 0.26a | 1.26 ± 0.41a | 3.09 ± 0.61a |
| S1450 | 1.58 ± 0.18b | 1.00 ± 0.22b | 2.57 ± 0.30b | 1.50 ± 0.25ab | 1.09 ± 0.30ab | 2.59 ± 0.53ab | 1.77 ± 0.51a | 2.02 ± 0.63a | 3.79 ± 0.99a |
| S1905 | 1.60 ± 0.47b | 0.99 ± 0.40b | 2.59 ± 0.84b | 1.29 ± 0.56ab | 0.89 ± 0.52ab | 2.18 ± 1.07b | 1.78 ± 0.25a | 1.35 ± 0.18a | 3.13 ± 0.37a |
| S2122 | 2.39 ± 0.46a | 1.68 ± 0.43a | 4.07 ± 0.89a | 0.92 ± 0.37b | 0.49 ± 0.29b | 1.42 ± 0.65b | 1.96 ± 0.19a | 1.62 ± 0.58a | 3.59 ± 0.73a |
| S2124 | 1.99 ± 0.31ab | 1.43 ± 0.30ab | 3.42 ± 0.58ab | 1.26 ± 0.15ab | 0.79 ± 0.15b | 2.05 ± 0.28b | 1.65 ± 0.48a | 1.48 ± 0.60a | 3.14 ± 1.05a |
| Treatment | BRS 8H | BRS Aroeira | BRS 286 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| ------------------------------ Dry Weight (g) ------------------------------ | |||||||||
| AP | RP | T | AP | RP | T | AP | RP | T | |
| Control | 2.19 ± 0.32a | 1.50 ± 0.37a | 3.70 ± 0.65a | 1.91 ± 0.13ab | 1.47 ± 0.51a | 3.38 ± 0.62ab | 1.83 ± 0.26a | 1.26 ± 0.41a | 3.09 ± 0.61a |
| S1450 | 1.70 ± 0.61a | 1.12 ± 0.51a | 2.82 ± 1.04a | 1.38 ± 0.33b | 0.83 ± 0.18a | 2.21 ± 0.38b | 1.21 ± 0.50a | 1.19 ± 0.84a | 2.41 ± 1.17a |
| S1905 | 2.10 ± 0.24a | 1.50 ± 0.32a | 3.60 ± 0.56a | 1.52 ± 0.39ab | 1.12 ± 0.51a | 2.64 ± 0.73ab | 1.51 ± 0.16a | 1.11 ± 0.24a | 2.62 ± 0.40a |
| S2122 | 2.16 ± 0.39a | 1.51 ± 0.33a | 3.68 ± 0.66a | 2.19 ± 0.49a | 1.82 ± 0.36a | 4.01 ± 0.84a | 1.54 ± 0.28a | 1.18 ± 0.22a | 2.79 ± 0.49a |
| S2124 | 1.78 ± 0.31a | 1.30 ± 0.40a | 3.82 ± 0.67a | 1.89 ± 0.61ab | 1.41 ± 0.78a | 3.30 ± 1.37ab | 1.81 ± 0.26a | 1.61 ± 0.37a | 3.42 ± 0.59a |
| Treatment/ Concentration | n | Weight (mg) | ||
|---|---|---|---|---|
| Period of Leaf Collection | ||||
| 18 DAE | 23 DAE * | 30 DAE | ||
| Control | 12 | 109.38b | 165.64b | 400.28a |
| 106 CFU mg−1 | 10 | 74.24a | 75.47a | 329.64a |
| 108 CFU mg−1 | 12 | 74.94a | 95.07a | 332.98a |
| Mean | 86.05 | 160.65 | 394.55 | |
| C.V. (%) | 36.06 | 45.72 | 38.26 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Costa, F.S.S.; Praça, L.B.; Gomes, A.C.M.M.; dos Santos, R.C.; Soares, C.M.S.; Monnerat, R.G. Bacillus thuringiensis Effect on the Vegetative Development of Cotton Plants and the Biocontrol of Spodoptera frugiperda. Agronomy 2020, 10, 1889. https://doi.org/10.3390/agronomy10121889
da Costa FSS, Praça LB, Gomes ACMM, dos Santos RC, Soares CMS, Monnerat RG. Bacillus thuringiensis Effect on the Vegetative Development of Cotton Plants and the Biocontrol of Spodoptera frugiperda. Agronomy. 2020; 10(12):1889. https://doi.org/10.3390/agronomy10121889
Chicago/Turabian Styleda Costa, Flávia Santana Souza, Lílian Botelho Praça, Ana Cristina Menezes Mendes Gomes, Roseane Cavalcanti dos Santos, Carlos Marcelo Silveira Soares, and Rose Gomes Monnerat. 2020. "Bacillus thuringiensis Effect on the Vegetative Development of Cotton Plants and the Biocontrol of Spodoptera frugiperda" Agronomy 10, no. 12: 1889. https://doi.org/10.3390/agronomy10121889
APA Styleda Costa, F. S. S., Praça, L. B., Gomes, A. C. M. M., dos Santos, R. C., Soares, C. M. S., & Monnerat, R. G. (2020). Bacillus thuringiensis Effect on the Vegetative Development of Cotton Plants and the Biocontrol of Spodoptera frugiperda. Agronomy, 10(12), 1889. https://doi.org/10.3390/agronomy10121889