1. Introduction
Soybean (
Glycine max (L.) Merr.), with its multiple uses, is one of the most important crops worldwide [
1]. The high-quality of its protein makes it an important component of animal feed for both milk and meat production. Soybean oil is also used for human nutrition. As the demand for vegetarian and vegan food has increased, vegetable proteins and oils from soybeans have gained importance in recent years [
2]. The world soybean production in 2017 was 352.6 million tons, of which 77% was GMO soybean [
3,
4]. The average soybean grain yield in the entire European Union in the years 2016–2017 (3.0 t ha
−1) was 10% higher than the yield in Germany (2.7 t ha
−1), 20 % lower than in the USA (3.4 t ha
−1), and about 13% lower than in Brazil (3.1 t ha
−1) [
4]. The mean protein content in these trials (41.6%) was comparable to the global average protein content; for example, Brazil (40.9%), USA (41.4 %), and China (42.1%). The oil content in the trials (19.1%) was higher than the oil content in Brazil (18.7%), USA (18.8%), and China (16.8%) [
5]. This confirms that the potential for both protein production and oil production via soybean cultivation in Germany gets little attention and consideration. The cultivation of GMO soybean is not permitted in Germany. For environmental and health safety reasons consumers in Germany prefer GMO-free products; this necessitates domestic production, but soy is not a traditional crop in Germany [
6]. However, climate change and increasing temperatures in Central Europe offer opportunities to implement and extend soybean into non-traditional areas [
1,
7,
8]. Soybean has a high heat demand. It can therefore only be lucratively cultivated in the southern, warmer parts of Germany [
9]. Soybean is also historically a short-day plant, originating from regions with relatively high temperatures during the growing season [
1], but breeding progress has made it possible to grow almost day-neutral soybean varieties [
2,
10]. The beginning of reproductive development (bud formation and flowering) is determined by the latitude and temperature sum of the growing area and genotype (E-gene alleles) [
11]. Changes in day length and prevailing daily temperature are of primary importance for these steps [
12]. The soybean starts to flower when the length of daylight becomes shorter than critical photoperiod at the growing location [
10]. The flowering period and pod filling are also highly sensitive to water stress [
13], and water shortages during these periods can lead to significant yield losses up to 20–25 % [
4,
14]. Soybeans require a precisely determined heat sum during the growing season to mature; for example, >2400 Crop Heat Units (CHU) for very early MG 000 varieties, 2400–2600 CHU for early MG 00 varieties, and well over 3200 CHU for late maturing varieties [
15]. In Germany, early and very early varieties of maturity groups MG 00 and MG 000, which were developed to suit our climatic conditions, are cultivated [
10]. In the USA, in contrast, varieties from later maturity groups—MG V, MG IV, genotypes with highest, to MG 0, genotypes with the lowest requirement for heat sum, are grown [
16].
On average, soybean seeds contain 18–25% oil [
2]. The by-product of oil extraction is soybean meal, containing 40–44% crude protein [
17]. The concentrations of these two components are genetically predetermined but can vary depending on the environment [
18]. The protein and oil content of soybean seeds are negatively correlated [
19,
20]. A higher temperature during reproductive growth tends to result in higher oil content [
21,
22]. The effect of temperature on protein content, however, is not yet clear. There is evidence that warm temperatures (>20 °C to <28 °C) during the growing period increase the protein content of soybean [
23,
24]. However, a negative correlation between temperature and protein content has also been reported [
25]. The effects of precipitation on protein and oil content of soybean seeds seem to be inconsistent; there is a positive correlation between precipitation during the growing season and oil content, but a negative correlation between precipitation and protein content [
26,
27]. Also, a negative correlation of both traits with precipitation has been reported [
25].
To meet the demands of protein for food and feed and for soy oil by domestically produced soybeans, it is important to know in which growing areas varieties from different maturity groups (MG 00 and MG 000) develop their best potential. Political decisions and commercial demand as well as preferences in Germany call for an expansion of organic agricultural production [
28]. Soybean is one of the few crops that delivers comparable productivity under both organic and conventional farming [
29]. The regions where soybean with a high protein content cannot be successfully grown, would be better suited for oil production, especially in organic farming, since there is still no suitable organic oil crop in Germany. The objective of this study was to determine the effects of solar radiation, temperature, and precipitation on soybean seed productivity, protein, and oil content in Germany. The following hypotheses were tested:
- (1)
High solar radiation ensures high grain yield.
- (2)
Insufficient precipitation leads to a high oil content and low protein content of soybean seeds.
- (3)
High temperatures during the growing season increase the protein content of soybean seeds.
3. Results
The analysis of variance showed highly significant effects of variety on protein and oil content (
Table 4). The interaction of year × location was also highly significant for protein and oil content.
The highest heritability was estimated for protein content with H2 = 0.80. The heritabilities for seed and protein yields were H2 = 0.55 and for oil yield H2 = 0.61.
The mean seed yield of varieties from maturity group MG 00 was higher than that from maturity group MG 000 (
Table 5). The means of protein and oil content were almost equal for both maturity groups.
The responses of varieties grouped by maturity groups and depending on year revealed significant correlations ranging from r = 0.25 to r = 0.74 (
Table 6). In
Table 6, the effects of year are shown.
Varieties from the maturity group MG 000 were significantly correlated with environmental conditions compared to varieties from the maturity group MG 00. Solar radiation was the main factor leading to an increase in seed yield and thus to increases in protein yield and oil yield. Precipitation often significantly increased yield and thus protein yield and oil yield in MG 000 varieties. Crop Heat Units, which had no significant effects on the observed traits in MG 00 varieties, were negatively correlated with seed yield, protein yield, and oil yield in MG 000 varieties. The response of MG 000 varieties to precipitation correlated positively with seed yield, protein yield, and oil yield. Crop Heat Units (CHU) and CHU at maturity correlated negatively with yield and thus protein and oil yields.
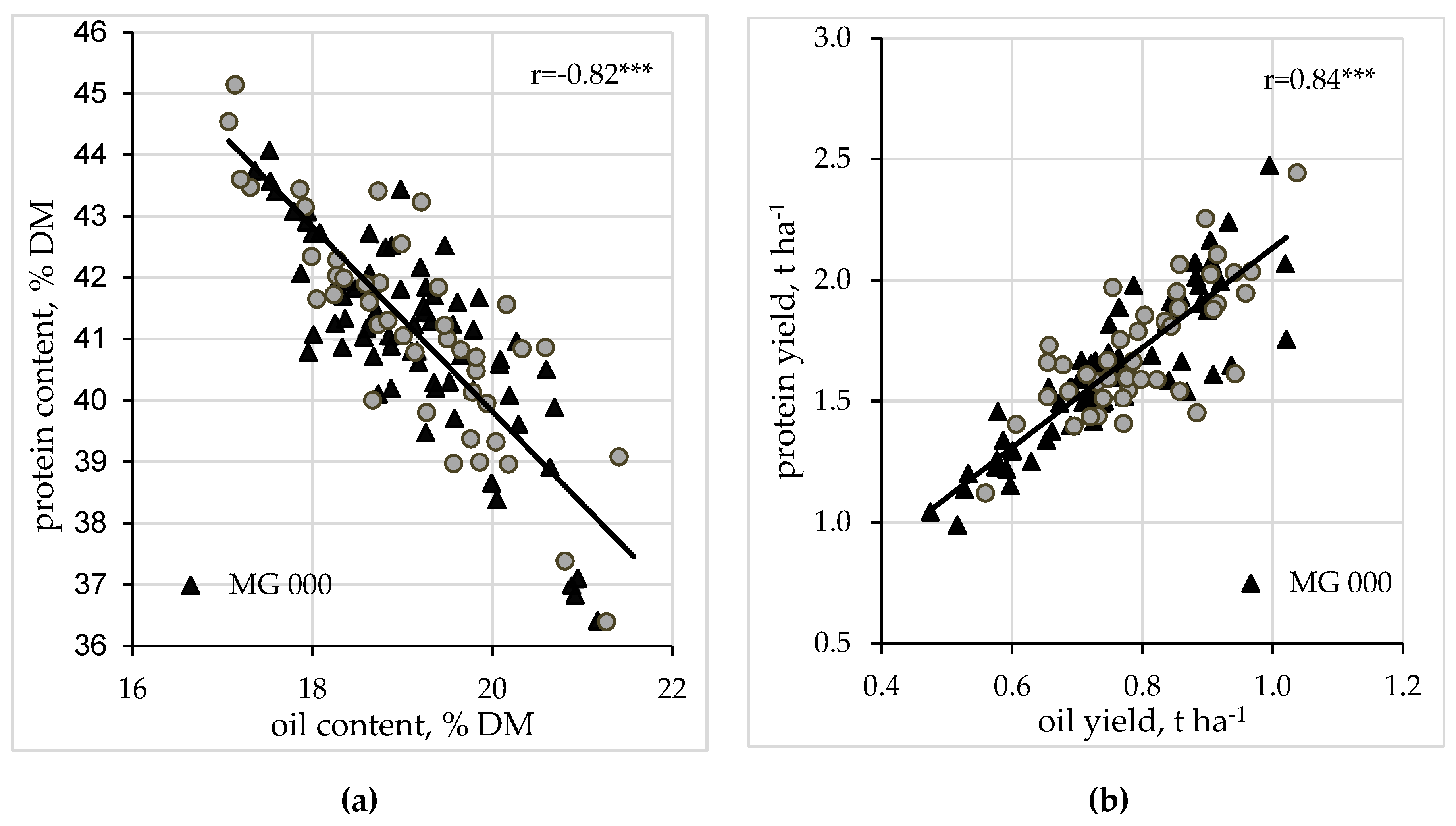
There was a strong negative correlation (r = −0.82 ***) between oil content and protein content over all varieties, locations, and trial years (
Figure 1a).
Protein content ranged from 35.4% to 45.1% with a mean of 41.1%. Oil content ranged from 17.1% to 21.6% with a mean of 19.1% (
Table 5). The correlation between protein yield and oil yield was positive at r = 0.84, at the highly significant level (
p < 0.01) (
Figure 1b).
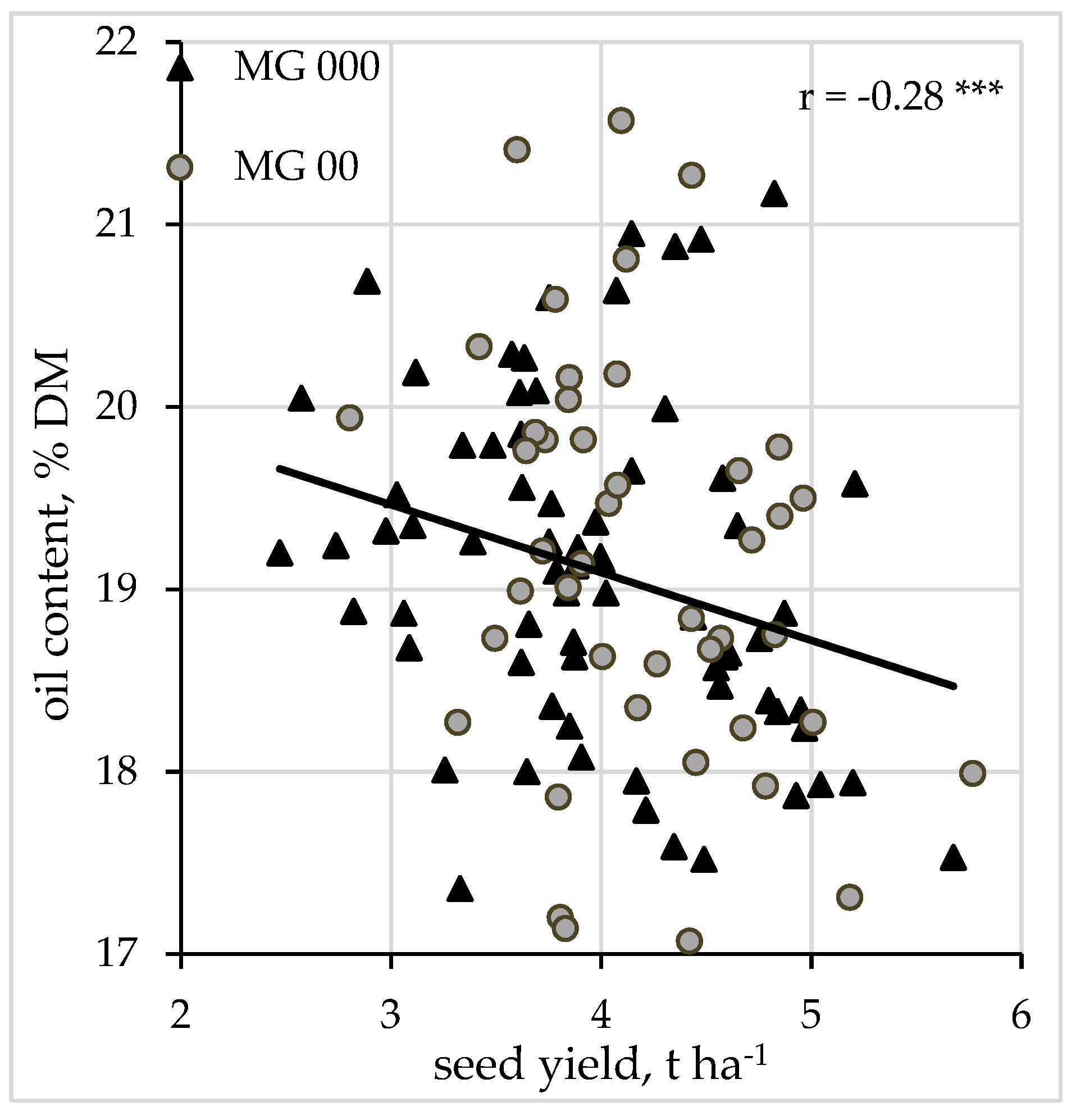
The correlation between seed yield and protein content was low and not significant (data not shown). In contrast, seed yield and oil content were significantly negatively correlated at r = −0.28 *** (
Figure 2).
4. Discussion
The high heritability of traits contributes to genetic progress, a characteristic that informs the selection of the most suitable genotypes as future varieties [
17,
38]. The conditions under which soybeans are grown, including the environment, determine the breeding direction, and this in turn depends on the geographic regions and the proposed cultivation of specific varieties [
39,
40]. Weather conditions differed in both trial years (
Table 3); only night temperatures and solar radiation did not vary greatly over all locations. On average, the year 2017 was moister and cooler than 2016. Crop Heat Units at maturity in 2017 were approximately 60–160 units lower than in 2016. Solar radiation at flowering and-seed filling however, was on average 30 kWhm
−2 higher in 2017 than in 2016. As a result, the soybean production properties correlated with environmental factors with varying degrees of intensity (
Table 6).
The most critical period in the entire vegetation period of soybean concerning water availability and sufficiency is R4-R6 (pod formation and seed filling) [
14,
18,
21,
41]. The results of the current study, with a positive correlation (r = 0.47) between seed yield and precipitation during flowering-seed filling are in accordance with existing literature. The early maturing varieties of MG 000 were more sensitive to increases in precipitation in the period R4-R6 than MG 00 varieties in each trial year (
Table 6). This may have been due to the shorter seed filling period and shorter duration of individual periods of vegetation growth of soybean from maturity group 000. This agrees with Souza et al. [
42], who compared MG 00 (early ripening) and II (later ripening) varieties in Brazil and also with Chen und Wiatrak [
41], who performed experiments with MG IV to VIII in the USA. High solar radiation at flowering increased seed yield in the current study over the two experimental years independent of the maturity group (
Table 6), similar to findings of other researchers [
13,
39,
43]. The intensification of solar radiation at the time of flowering could stimulate photosynthesis and thus raise the production of assimilates, leading to an increase in seed yield by increasing the number of pods per reproductive node [
39]. Similar results (r = 0.37) for five maturity groups in the USA have also been reported by Kane und Grabau 1992 [
44]. At the same time, high temperatures at certain stages of development (R3-R6) can have a negative impact on seed yield and quality characteristics of soybean. For example, Dornbos and Mullen and Gibson and Mullen [
45,
46] reported that high temperatures (>29 °C) led to a reduction in number of seeds per plant and in seed weight The negative correlation between CHU at maturity and seed yield in our study (r = −0.42) was higher for the early maturing varieties (MG 000). MG 000 varieties can mature in the middle of September in Germany, whereas MG 00 varieties are usually harvested 10–12 days later. High temperatures during the final period of ripening may interfere with the physiological maturing process and simply lead to desiccation, not perfect maturing [
9,
47].
As previous studies suggest, the protein content of soybean seeds is influenced by day and night temperatures and varies depending on their stage of development [
18,
47]. Temperatures of about 20°C correlated significantly positively with soybean protein content. Our results indicate that higher temperatures at maturity may lead to higher protein content (
Table 6). The protein content of soybean seeds increased with increasing mean temperatures >28 °C and remained constant <28 °C [
18]. Compared to our studies, previous studies quoted above examined varieties with a much longer vegetation period (MG up to VIII) and in locations at a much lower latitude (<29.4 °S). In our study, the early ripening varieties (MG 00 and MG 000) reacted more rapidly to the higher temperatures they experienced, which have begun to occur more often in Germany.
Oil content was the only seed property that correlated negatively with T
night at development stage R1-R3, similar to previous results [
48,
49,
50]. However, soybean studies at 23 locations in Argentina indicated the opposite [
27]. Such differences in results were likely due to the fact that those studies [
25] were conducted in different regions in Argentina while other three were performed in the USA (Iowa) [
48,
49,
50]; the varieties examined, therefore, belonged to different maturity groups (MG IV-V) (middle late) and II (late).
The time during the growing season when drought occurs can affect the intensity of its influence on protein and oil content [
21]. It is well established from studies in the USA, South America, Central Europe, and Russia that oil content is negatively correlated with precipitation at flowering-seed filling phase [
14,
20,
23,
47]. Our studies, conducted in a region (Germany) previously considered not suitable for soybean cultivation (because only few varieties were able to ripen at the low CHU), is in line with these results (
Table 6). A much stronger negative correlation, as compared to the average of both years, was observed in the dry year 2016 compared to 2017 (
Table 6). It is possible that if the season were already dry, further water deficits at this stage could have had a stronger increasing effect on the oil content. If water deficiency occurred at R5-R6, soybean seeds did not fully develop and did not reach their variety-specific size [
42,
49,
51,
52]. The transfer processes of nutrients (first of all carbohydrate) and thus seed quality were impaired [
14,
47], and the assimilates could not be completely converted into protein and oil [
52]. However, the protein and oil content are negatively correlated, so the protein content of soybean seeds increased by 3–5% under drought stress then oil content decreased by 2–3% compared to seeds of control plants with optimal water supply [
48]. Protein formation in soybean seeds starts about 10–15 days earlier than that of fats [
20]; when drought becomes more severe, protein synthesis continues but nutrients for oil are exhausted [
21]. From a biochemical point of view, this can be linked to the activation of superoxide, which, in a drought, promotes the accumulation of protein more than that of oil [
53]. Such a phenomenon is already known from cereals; when under drought stress, starch enzyme activity is more strongly affected than enzyme for protein formation [
54]. Therefore, at drier sites and/ or at locations where drought periods can be expected, varieties that have been selected for high oil content and oil yield should not be grown; rather, this production objective should be pursued at more humid locations. The negative correlation between precipitation at flowering-seed filling and oil content was only detected under dry conditions; under more humid conditions we could not detect this effect (trial year 2017).
Negative correlations of oil content with protein content of soybeans have been reported for soybean since the mid-1960s [
17]. The highest negative correlation between protein and oil content was found in the European early maturing genotypes (0-0000); for example, almost r = −0.9 in Germany [
17], r = −0.79 in Austria [
23], and r = −0.83 in Slovenia [
55]. According to investigations in Russia over 27 years, this correlation was on average r = −0.53 [
20] and in the Midwest of the USA r = −0.64 [
56]. Piper und Boote analyzed 1863 varieties from 10 maturity groups at 60 locations throughout the USA and documented a correlation of r = −0.43 [
18]. This strong negative correlation between protein content and oil content was also seen in our results r = −0.82 (average over only early maturing MG 00 and MG 000 varieties—
Figure 1a), as typical for Central Europe. The reasons for the strong negative correlation between protein and oil content are not completely clear. These two properties are determined by the same combination of genes and it is difficult to do plant breeding work only to increase of one of these properties without leaving the other unchanged [
20,
57,
58]. However, the synthesis of protein and oil in soybean seeds utilize the same carbon metabolism, and the synthesis processes interact with each other [
59]. Therefore, competition for carbon can develop, and this is reflected in a negative correlation.
If the aim is to increase total soybean protein production in Germany, increasing seed yield would increase the amount of protein per unit area. In this context, the phenotypic correlations of seed yield with protein and oil content are important. Though seed yield correlated negatively with protein content but positively with oil content in the common studies [
17,
56], our investigation did not find a significant correlation of seed yield with protein content; this is similar to the findings of Whaley und Eskandari [
40]. Despite its importance, there is no clear evidence of a relationship between seed yield and protein content. Test results from Germany provide information about a low and negative correlation [
17] while in the USA a low but positive correlation has been calculated [
22]. This correlation is influenced by year and weather/soil conditions in the growing area [
23]. Such a regulation process between protein content and seed yield is known from wheat [
60]. Therefore, genotypic variation together with environment provides many breeding possibilities for the adaptation of new varieties [
40]. The low negative correlation between seed yield and oil content in our study contrasts with other available results [
22,
51]. It is essential to take the findings of referenced studies into consideration when planning the production, especially the choice of varieties, of certain soybean products such as soybean meal, soybean oil or tofu, and other foodstuffs, in different growing areas of a country with a temperate climate.
Time to maturity, drought tolerance, and heat tolerance are important traits to consider in adapting soybean varieties to different growing environments. Since the levels of protein content and oil content of the maturity groups MG 00 and MG 000 barely differed, and since the seed yields of MG 00 varieties trended higher than those of MG 000 varieties, MG 00 varieties should preferentially be cultivated and bred in regions suitable either for protein or oil production. To select for varieties with high seed yield and with long ripening period, such as the tested MG 00 varieties, the testing should occur in regions with comparatively high solar radiation (for example location Eckartsweier). The late ripening varieties from MG 0 would also be appropriate for these locations. In regions with comparatively high precipitation, MG 000 varieties would be suitable because of their high seed yields and therefore high protein and oil yields (for example Grünseiboldsdorf, Wetterau, and Rossleben). If MG 00 varieties were cultivated in drier areas (such as Eckartsweier and Guesten) the oil content would be higher. High temperatures at maturation would result in higher protein content in MG 000 varieties (Eckartsweier, Wetterau and Grünseiboldsdorf). The following table ranks the tested soybean varieties according to their adaption to locations with different environmental properties in Germany (
Table 7).

 ,
, 


{kind=link}
{kind=link}