Biomass Performance and Competition Effects in an Established Temperate Agroforestry System of Willow and Grassland—Results of the 2nd Rotation


Abstract
1. Introduction
2. Materials and Methods
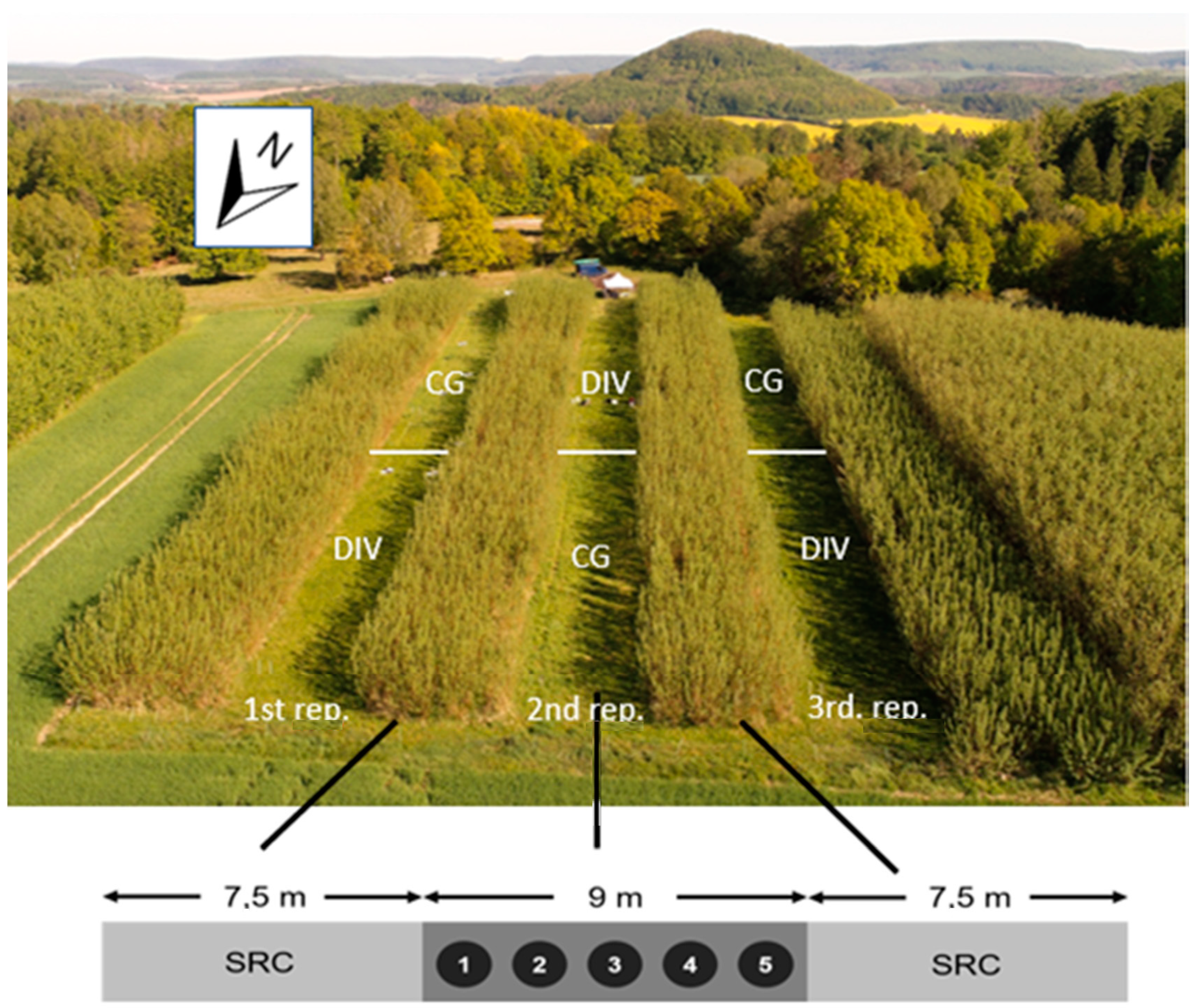
2.1. Description of Site and Experimental Design
2.2. Data Acquisition
2.3. Land Equivalent Ratio (LER)
2.4. Statistics
3. Results
3.1. Agroforestry System Yield
3.2. Grassland Yield along the Alleyway Transect
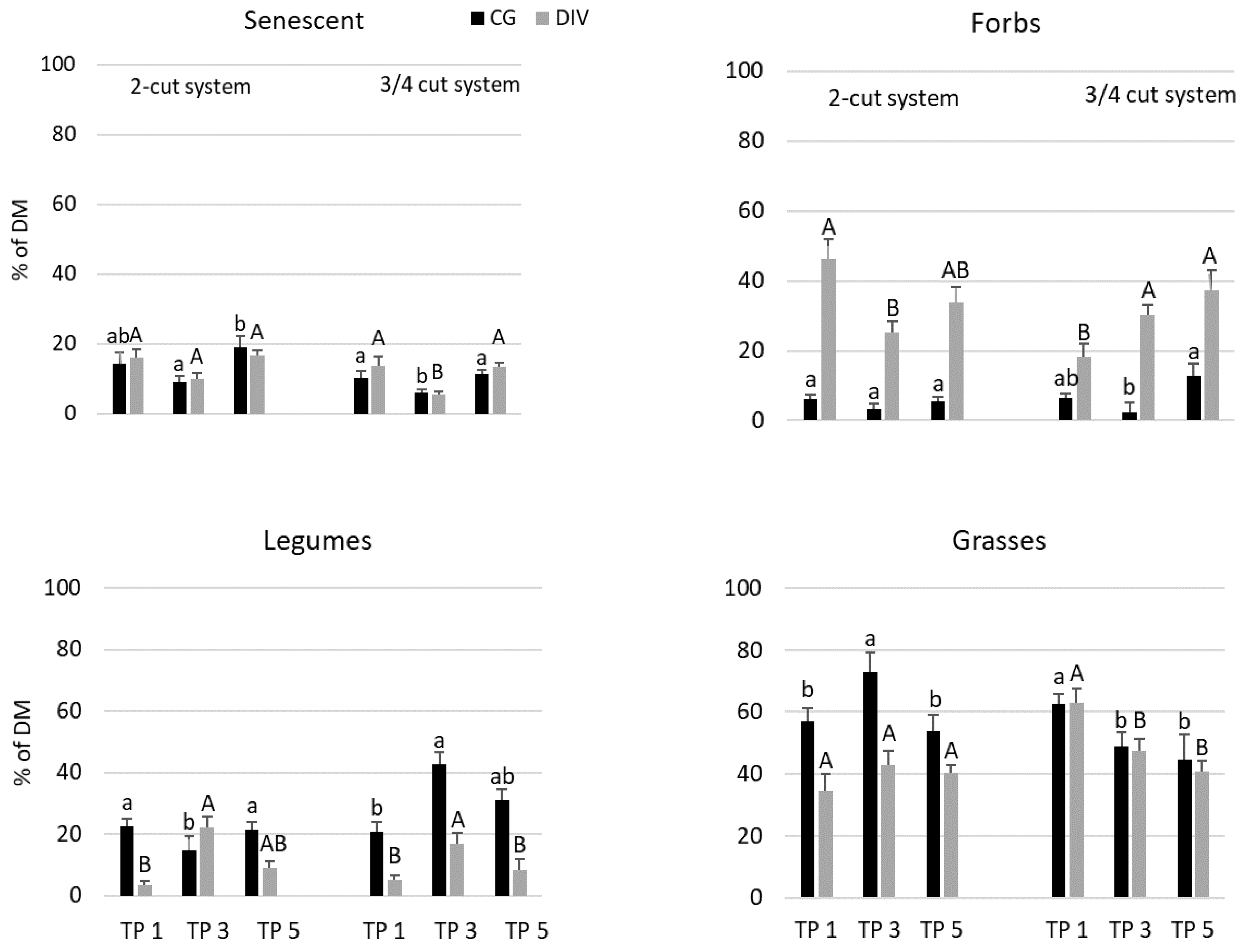
3.3. Functional Group Composition in the Grassland Mixtures
3.4. Land Equivalent Ratio (LER)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, J.; Pearce, B.D.; Wolfe, M.S. Reconciling productivity with protection of the environment: Is temperate agroforestry the answer? Renew. Agric. Food Syst. 2013, 28, 80–92. [Google Scholar] [CrossRef]
- Dinesh, D.; Campbell, B.; Bonilla-Findji, O.; Richards, M. 10 Best bet Innovations for Adaptation in Agriculture: A Supplement to the UNFCCC NAP Technical Guidelines: CCAFS Working Paper No. 215; CGIAR Research Program on Climate Change, Agriculture and Food Security (CCAFS): Wageningen, The Netherlands; Available online: www.ccafs.cgiar.org (accessed on 23 August 2020).
- Quinkenstein, A.; Wöllecke, J.; Böhm, C.; Grünewald, H.; Freese, D.; Schneider, B.U.; Hüttl, R.F. Ecological benefits of the alley cropping agroforestry system in sensitive regions of Europe. Environ. Sci. Policy 2009, 12, 1112–1121. [Google Scholar] [CrossRef]
- Tsonkova, P.; Böhm, C.; Quinkenstein, A.; Freese, D. Ecological benefits provided by alley cropping systems for production of woody biomass in the temperate region: A review. Agrofor. Syst. 2012, 85, 133–152. [Google Scholar] [CrossRef]
- Jose, S.; Bardhan, S. Agroforestry for biomass production and carbon sequestration: An overview. Agrofor. Syst. 2012, 86, 105–111. [Google Scholar] [CrossRef]
- Lorenz, K.; Lal, R. Soil organic carbon sequestration in agroforestry systems. A review. Agron. Sustain. Dev. 2014, 34, 443–454. [Google Scholar] [CrossRef]
- IPCC 2019.: Summary for Policymakers. In: Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems. Shukla, P.R.,Skea, J., Buendia, E.C., Masson-Delmotte, V., Pörtner, H.-O., Roberts, D.C., Zhai, P., Slade, R., Connors, S., van Diemen, R., et al., Eds. Available online: https://www.ipcc.ch/srccl/ (accessed on 4 September 2020).
- Hernández-Morcillo, M.; Burgess, P.; Mirck, J.; Pantera, A.; Plieninger, T. Scanning agroforestry-based solutions for climate change mitigation and adaptation in Europe. Environ. Sci. Policy 2018, 80, 44–52. [Google Scholar] [CrossRef]
- Fagerholm, N.; Torralba, M.; Burgess, P.J.; Plieninger, T. A systematic map of ecosystem services assessments around European agroforestry. Ecol. Indic. 2016, 62, 47–65. [Google Scholar] [CrossRef]
- Paul, C.; Weber, M.; Knoke, T. Agroforestry versus farm mosaic systems—Comparing land-use efficiency, economic returns and risks under climate change effects. Sci. Total. Environ. 2017, 22–35. [Google Scholar] [CrossRef]
- Böhm, C.; Tsonkova, P.; Zehlius-Eckert, W. Wie Können Agroforstsysteme Praktikabel in Das Deutsche Agrarförderrecht Eingebunden Werden. 5. Forum Agroforstsysteme, Cottbus-Senftenberg 2016, 5, 7–16. [Google Scholar]
- Tsonkova, P.; Mirck, J.; Böhm, C.; Fütz, B. Addressing farmer-perceptions and legal constraints to promote agroforestry in Germany. Agrofor. Syst. 2018, 92, 1091–1103. [Google Scholar] [CrossRef]
- Jose, S.; Gillespie, A.; Pallardy, S. Interspecific interactions in temperate agroforestry. Agrofor. Syst. 2004, 237–255. [Google Scholar] [CrossRef]
- Nurk, L.; Graß, R.; Wachendorf, M. Productivity at the tree-crop interface of a young willow-grassland alley cropping system. Agrofor. Syst. 2016, 92, 71–83. [Google Scholar] [CrossRef]
- Coussement, T.; Maloteau, S.; Pardon, P.; Artru, S.; Ridley, S.; Javaux, M.; Garré, S. A tree-bordered field as a surrogate for agroforestry in temperate regions: Where does the water go? Agric. Water Manag. 2018, 210, 198–207. [Google Scholar] [CrossRef]
- Gillespie, A.R.; Jose, S.; Mengel, D.B.; Hoover, W.L.; Pope, P.E.; Seifert, J.R.; Biehle, D.J.; Stall, T.; Benjamin, T.J. Defining competition vectors in a temperate alley cropping system in the midwestern USA: 1. Production physiology. Agrofor. Syst. 2000, 48, 25–40. [Google Scholar] [CrossRef]
- Ehret, M.; Graß, R.; Wachendorf, M. The effect of shade and shade material on white clover/perennial ryegrass mixtures for temperate agroforestry systems. Agrofor. Syst. 2015, 89, 557–570. [Google Scholar] [CrossRef]
- Gruenewald, H.; Brandt, B.K.; Schneider, B.U.; Bens, O.; Kendzia, G.; Hüttl, R.F. Agroforestry systems for the production of woody biomass for energy transformation purposes. Ecol. Eng. 2007, 29, 319–328. [Google Scholar] [CrossRef]
- Gamble, J.D.; Johnson, G.; Sheaffer, C.C.; Current, D.A.; Wyse, D.L. Establishment and early productivity of perennial biomass alley cropping systems in Minnesota, USA. Agrofor. Syst. 2013, 88, 75–85. [Google Scholar] [CrossRef]
- Lamerre, J.; Schwarz, K.-U.; Langhof, M.; Von Wühlisch, G.; Greef, J.-M. Productivity of poplar short rotation coppice in an alley-cropping agroforestry system. Agrofor. Syst. 2015, 89, 933–942. [Google Scholar] [CrossRef]
- Kanzler, M.; Böhm, C.; Mirck, J.; Schmitt, D.; Veste, M. Microclimate effects on evaporation and winter wheat (Triticum aestivum L.) yield within a temperate agroforestry system. Agrofor. Syst. 2019, 93, 1821–1841. [Google Scholar] [CrossRef]
- Ehret, M.; Bühle, L.; Graß, R.; Lamersdorf, N.; Wachendorf, M. Bioenergy provision by an alley cropping system of grassland and shrub willow hybrids: Biomass, fuel characteristics and net energy yields. Agrofor. Syst. 2014, 89, 365–381. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, Update 2015 International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015; Available online: http://www.fao.org/3/i3794en/I3794en.pdf (accessed on 17 August 2020).
- Hartmann, L.; Richter, F.; Busch, G.; Ehret, M.; Jansen, M.; Lamersdorf, N. Establishment of short rotation coppices in the South of Lower Saxony and in Central Thuringia in the context of the BEST-research framework—Site characteristics and initial biomass production. Forstarchiv 2014, 85, 134–150. [Google Scholar]
- Dupraz, C. Adequate design of control treatments in long term agroforestry experiments with multiple objectives. In Forestry Sciences; Springer Science and Business Media LLC: Cham Switzerland, 1999; Volume 60, pp. 35–48. [Google Scholar]
- Mead, R.; Willey, R.W. The Concept of a ‘Land Equivalent Ratio’ and Advantages in Yields from Intercropping. Exp. Agric. 1980, 16, 217–228. [Google Scholar] [CrossRef]
- Connolly, J. Developing Multisite Dynamic Models of Mixed Species Plant Communities. Ann. Bot. 2001, 88, 703–712. [Google Scholar] [CrossRef]
- Graves, A.R.; Burgess, P.J.; Palma, J.H.N.; Herzog, F.; Moreno, G.; Bertomeu, M.; Dupraz, C.; Liagre, F.; Keesman, K.; Van Der Werf, W.; et al. Development and application of bio-economic modelling to compare silvoarable, arable, and forestry systems in three European countries. Ecol. Eng. 2007, 29, 434–449. [Google Scholar] [CrossRef]
- Wilson, M.H.; Lovell, S.T. Agroforestry—The Next Step in Sustainable and Resilient Agriculture. Sustainability 2016, 8, 574. [Google Scholar] [CrossRef]
- Stamps, W.T.; Linit, M.J. The problem of experimental design in temperate agroforestry. Agrofor. Syst. 1998, 44, 187–196. [Google Scholar] [CrossRef]
- Huxley, P.A. Agroforestry experimentation: Separating the wood from the trees? Agrofor. Syst. 1987, 5, 251–275. [Google Scholar] [CrossRef]
- Aylott, M.J.; Casella, E.; Tubby, I.; Street, N.R.; Smith, P.; Taylor, G. Yield and spatial supply of bioenergy poplar and willow short-rotation coppice in the UK. New Phytol. 2008, 178, 358–370. [Google Scholar] [CrossRef]
- Dimitriou, I.; Mola-Yudego, B.; Aronsson, P. Impact of Willow Short Rotation Coppice on Water Quality. BioEnergy Res. 2012, 5, 537–545. [Google Scholar] [CrossRef]
- Stork, M.; Schulte, A.; Murach, D. Large-scale fuelwood production on agricultural fields in mesoscale river catchments—GIS-based determination of potentials in the Dahme river catchment (Brandenburg, NE Germany). Biomass Bioenergy 2014, 64, 42–49. [Google Scholar] [CrossRef]
- Sleight, N.J.; Volk, T.; Johnson, G.A.; Eisenbies, M.H.; Shi, S.; Fabio, E.S.; Pooler, P.S. Change in Yield Between First and Second Rotations in Willow (Salix spp.) Biomass Crops is Strongly Related to the Level of First Rotation Yield. BioEnergy Res. 2015, 9, 270–287. [Google Scholar] [CrossRef]
- Clinch, R.L.; Thevathasan, N.V.; Gordon, A.M.; Volk, T.; Sidders, D. Biophysical interactions in a short rotation willow intercropping system in southern Ontario, Canada. Agric. Ecosyst. Environ. 2009, 131, 61–69. [Google Scholar] [CrossRef]
- Fernández, M.E.; Gyenge, J.; Licata, J.; Schlichter, T.; Bond, B.J. Belowground interactions for water between trees and grasses in a temperate semiarid agroforestry system. Agrofor. Syst. 2008, 74, 185–197. [Google Scholar] [CrossRef]
- Mantino, A.; Volpi, I.; Micci, M.; Pecchioni, G.; Bosco, S.; Dragoni, F.; Mele, M.; Ragaglini, G. Effect of Tree Presence and Soil Characteristics on Soybean Yield and Quality in an Innovative Alley-Cropping System. Agronomy 2019, 10, 52. [Google Scholar] [CrossRef]
- Dohn, J.; Dembélé, F.; Karembé, M.; Moustakas, A.; Amévor, K.A.; Hanan, N.P. Tree effects on grass growth in savannas: Competition, facilitation and the stress-gradient hypothesis. J. Ecol. 2013, 101, 202–209. [Google Scholar] [CrossRef]
- Douglas, G.B.; Walcroft, A.S.; Hurst, S.E.; Potter, J.F.; Foote, A.G.; Fung, L.E.; Edwards, W.R.N.; Dijssel, C.V.D. Interactions between Widely Spaced Young Poplars (Populus spp.) and Introduced Pasture Mixtures. Agrofor. Syst. 2006, 66, 165–178. [Google Scholar] [CrossRef]
- Winterling, A. Erträge von Winterweizen in einem Agroforstsystem zur Energieholzgewinnung im ökologischen Landbau. In Proceedings of the 14th Scientific Conference for Organic Farming, Freising, Germany, 7–10 March 2017. [Google Scholar]
- Stagnari, F.; Maggio, A.; Galieni, A.; Pisante, M. Multiple benefits of legumes for agriculture sustainability: An overview. Chem. Biol. Technol. Agric. 2017, 4, 2. [Google Scholar] [CrossRef]
- Grüner, E.; Wachendorf, M.; Moeckel, T. The potential of UAV-borne spectral and textural information for predicting aboveground biomass and N fixation in legume-grass mixtures. PLoS ONE 2020, 15, e0234703. [Google Scholar] [CrossRef]
- Frame, J.; Newbould, P. Herbage production from grass/white clover swards. Forage Legumes 1984, 16, 15–35. [Google Scholar]
- Andrews, M.; Scholefield, D.; Abberton, M.; McKenzie, B.; Hodge, S.; Raven, J. Use of white clover as an alternative to nitrogen fertiliser for dairy pastures in nitrate vulnerable zones in the UK: Productivity, environmental impact and economic considerations. Ann. Appl. Biol. 2007, 151, 11–23. [Google Scholar] [CrossRef]
- Peyraud, J.L.; Le Gall, A.; Luescher, A. Potential food production from forage legume-based-systems in Europe: An overview. Irish J. Agric. Food Res. 2009, 48, 115–135. [Google Scholar]
- DeHaan, L.R.; Weisberg, S.; Tilman, D.; Fornara, D. Agricultural and biofuel implications of a species diversity experiment with native perennial grassland plants. Agric. Ecosyst. Environ. 2010, 137, 33–38. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) Accumulated Grassland Yield (t ha−1) in the AFS | ||||
| Effect | ||||
| Grassland mixture (M) | 0.2014 | |||
| Cutting frequency (C) | 0.1075 | |||
| M x C | 0.1652 | |||
| (b) Grassland Yield (t ha−1) Along the Alleyway Transect | ||||
| Effect | 2-cut CG | 2-cut DIV | 3/4-cut CG | 3/4-cut DIV |
| Transect point | <0.001 | 0.0074 | <0.001 | 0.004 |
| (c) Proportion of Functional Groups (% of DM) Along the Transect | ||||
| Effect | 2-cut CG | 2-cut DIV | 3/4-cut CG | 3/4-cut DIV |
| Grasses | 0.0178 | 0.4401 | 0.0081 | 0.0096 |
| Forbs | 0.4974 | 0.0431 | 0.0185 | 0.0124 |
| Legumes | 0.0179 | 0.0193 | 0.0243 | 0.0332 |
| Senescent | 0.0311 | 0.0984 | 0.0032 | 0.0046 |
| LER Values | ||||
|---|---|---|---|---|
| Treatment | 2-Cut CG | 2-Cut DIV | 3/4-Cut CG | 3/4-Cut DIV |
| 0.86 | 0.82 | 0.86 | 0.75 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Graß, R.; Malec, S.; Wachendorf, M. Biomass Performance and Competition Effects in an Established Temperate Agroforestry System of Willow and Grassland—Results of the 2nd Rotation. Agronomy 2020, 10, 1819. https://doi.org/10.3390/agronomy10111819
Graß R, Malec S, Wachendorf M. Biomass Performance and Competition Effects in an Established Temperate Agroforestry System of Willow and Grassland—Results of the 2nd Rotation. Agronomy. 2020; 10(11):1819. https://doi.org/10.3390/agronomy10111819
Chicago/Turabian StyleGraß, Rüdiger, Sarah Malec, and Michael Wachendorf. 2020. "Biomass Performance and Competition Effects in an Established Temperate Agroforestry System of Willow and Grassland—Results of the 2nd Rotation" Agronomy 10, no. 11: 1819. https://doi.org/10.3390/agronomy10111819
APA StyleGraß, R., Malec, S., & Wachendorf, M. (2020). Biomass Performance and Competition Effects in an Established Temperate Agroforestry System of Willow and Grassland—Results of the 2nd Rotation. Agronomy, 10(11), 1819. https://doi.org/10.3390/agronomy10111819