



Identification and Characterization of Fibronectin-Binding Peptides in Gelatin

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Preparation of FN Sepharose Affinity Media
2.2.2. Tryptic Digestion
2.2.3. FN Affinity Chromatography [9,14,17]
2.2.4. MALDI-TOF MS Analysis
2.2.5. HPLC-MS Analysis
2.2.6. Database Searching and Data Processing
2.2.7. Surface Modification of DPI Chip
2.2.8. DPI Analysis and Data Processing [17,20]
3. Results and Discussion
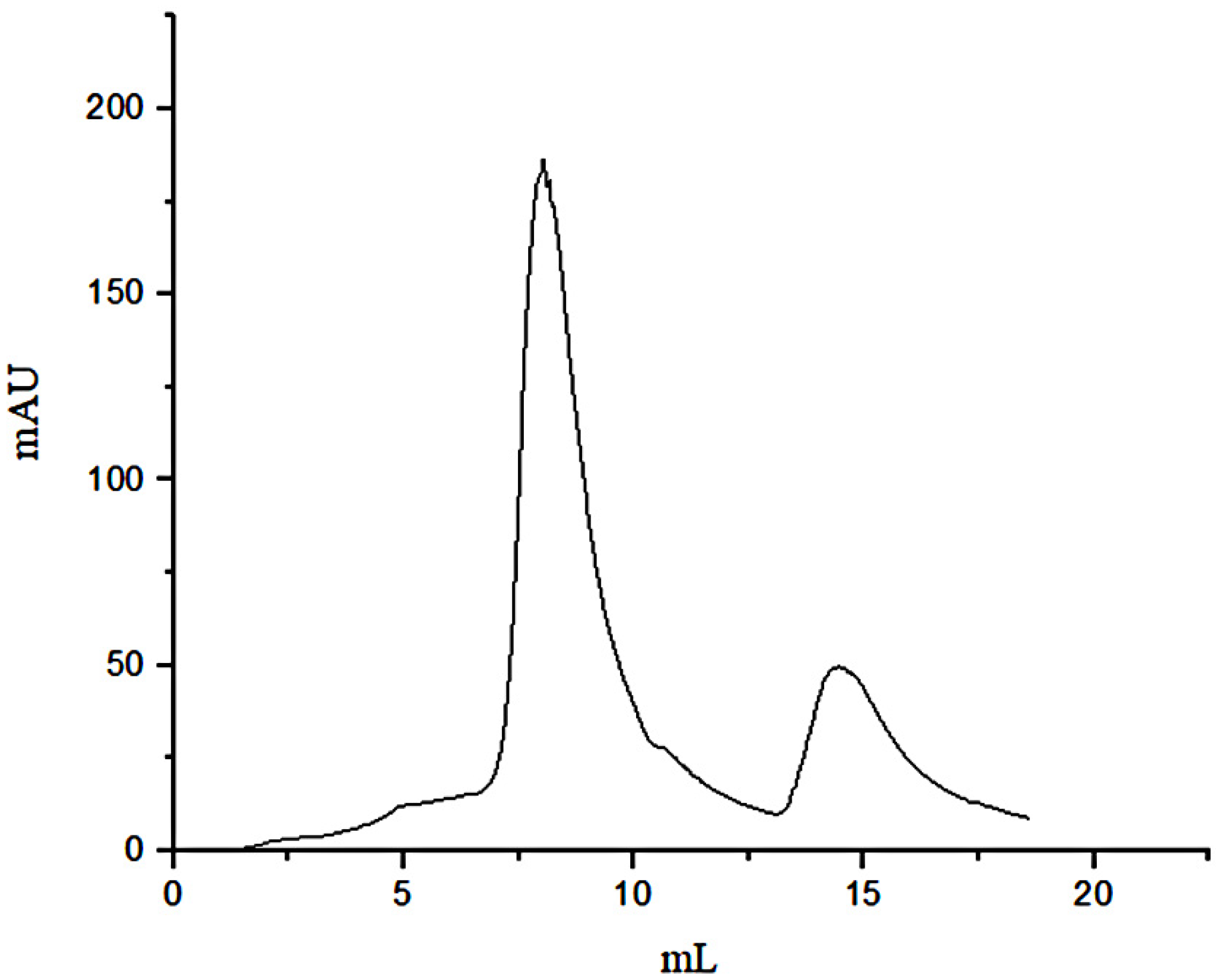
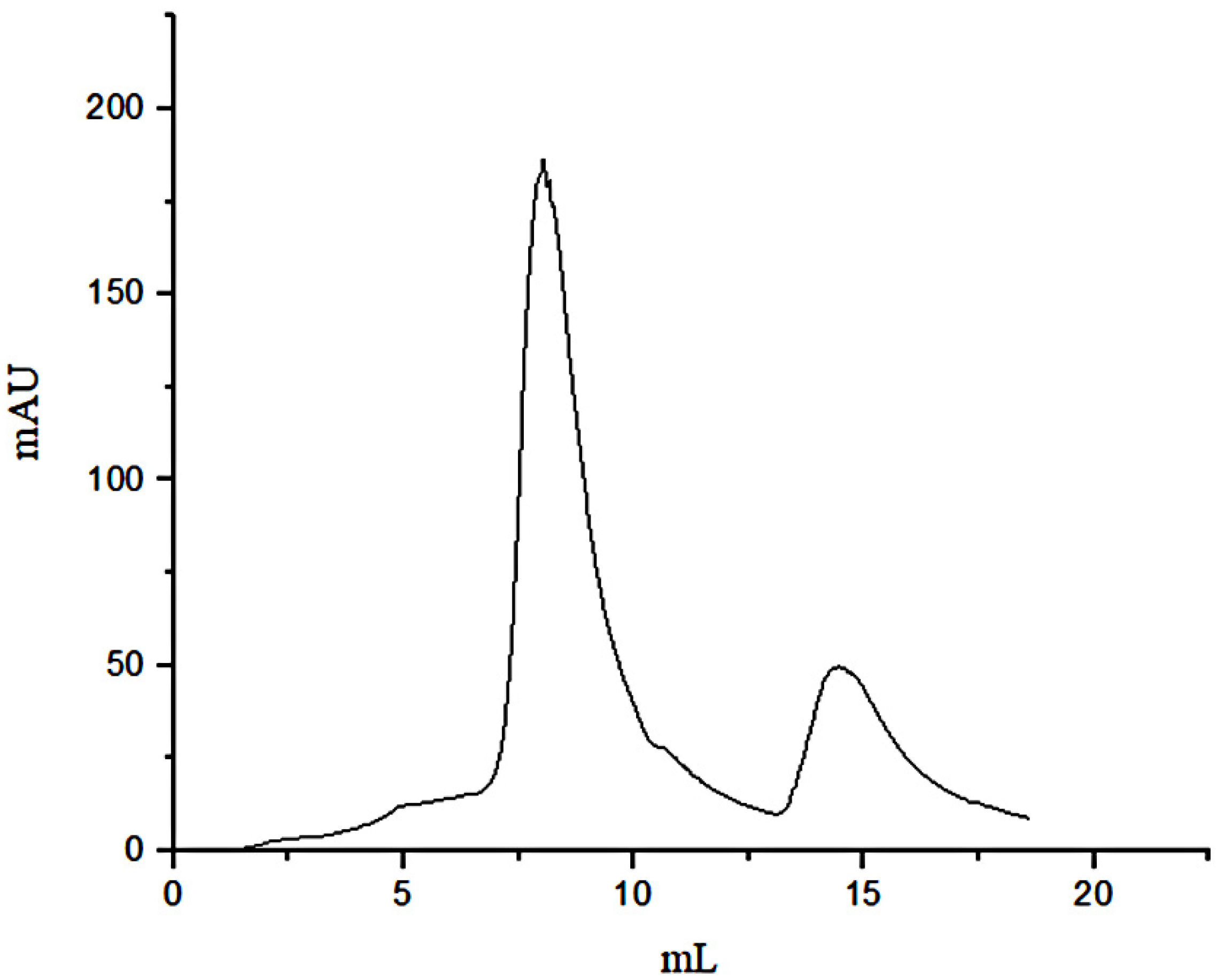
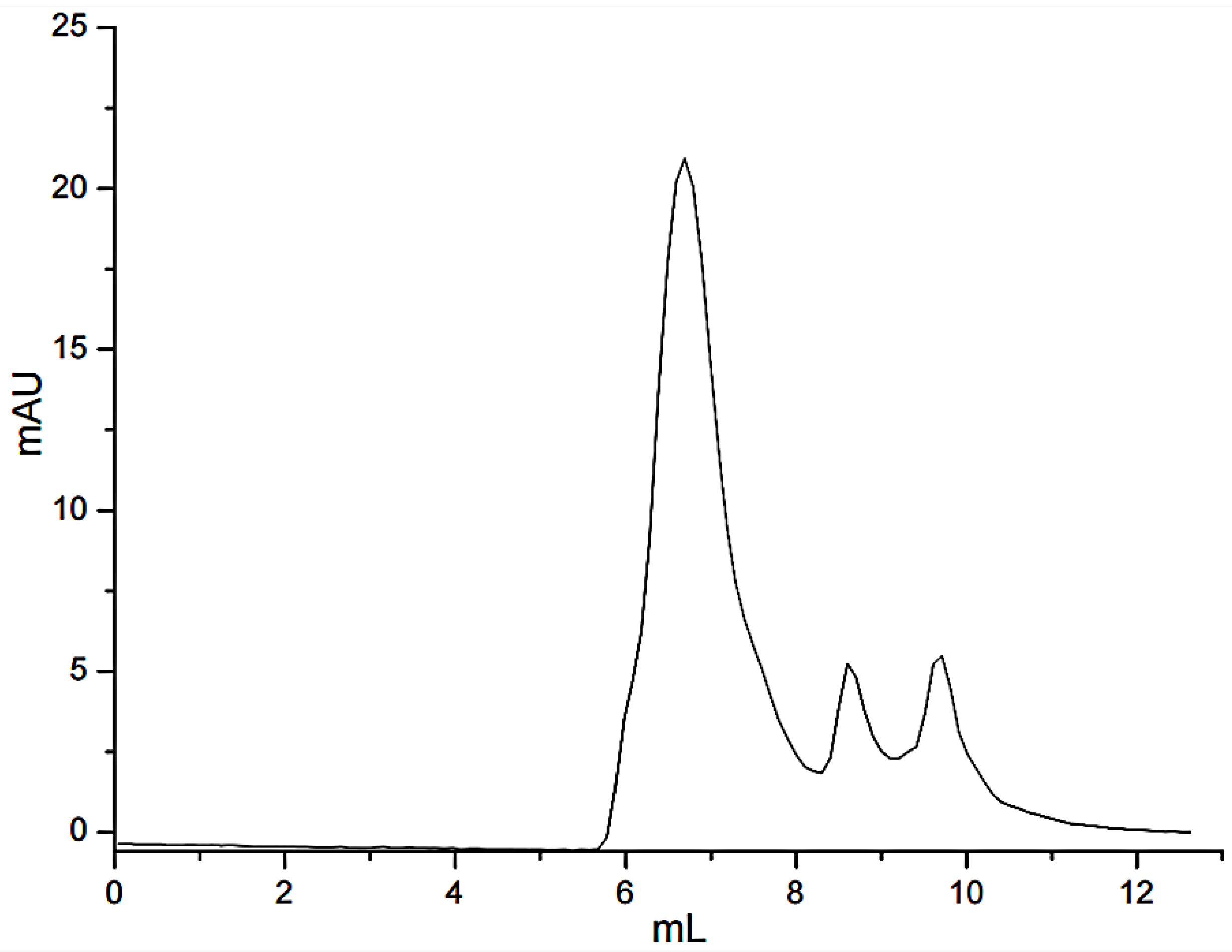
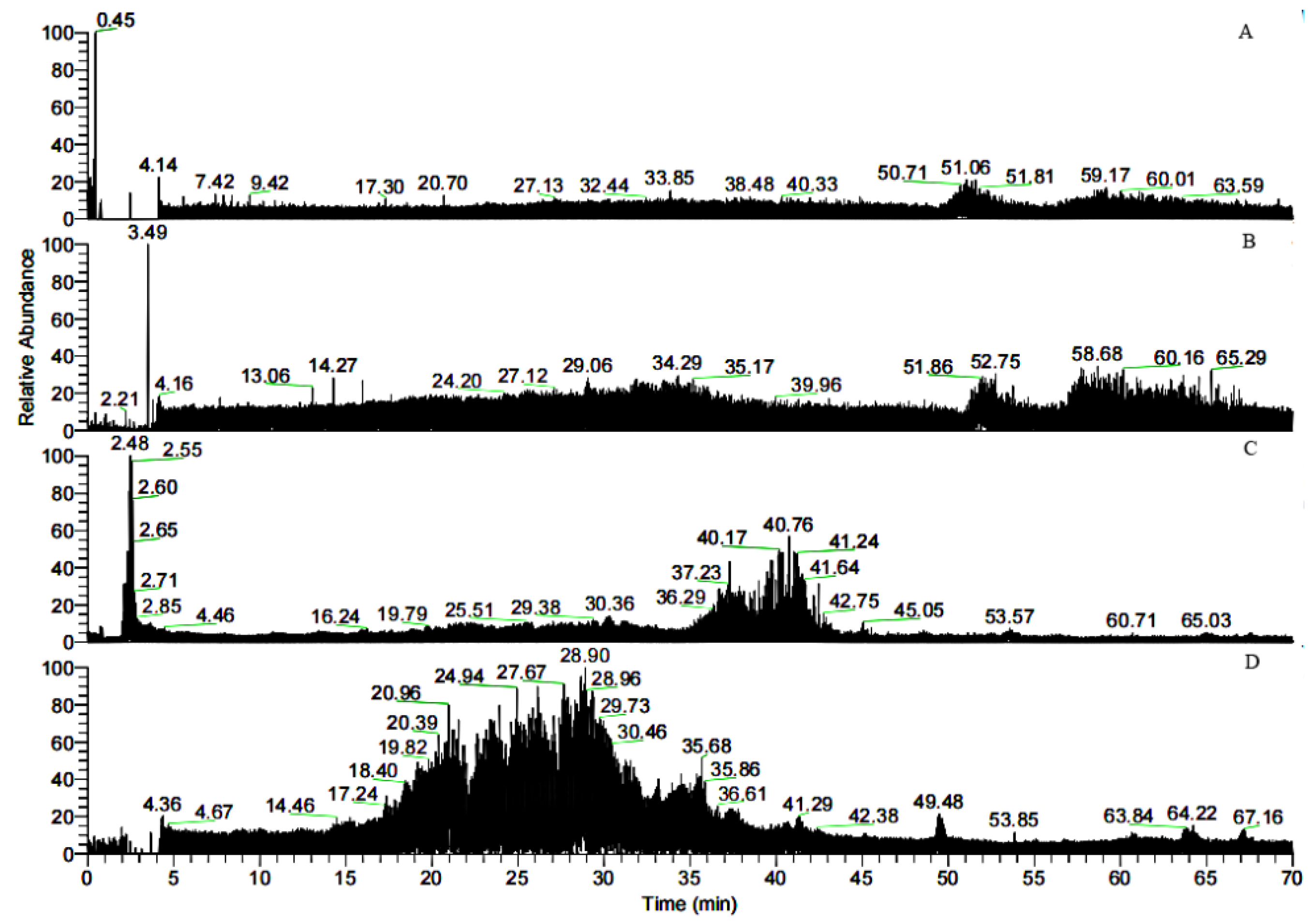
3.1. Separation of Peptides through FN Affinity Chromatography
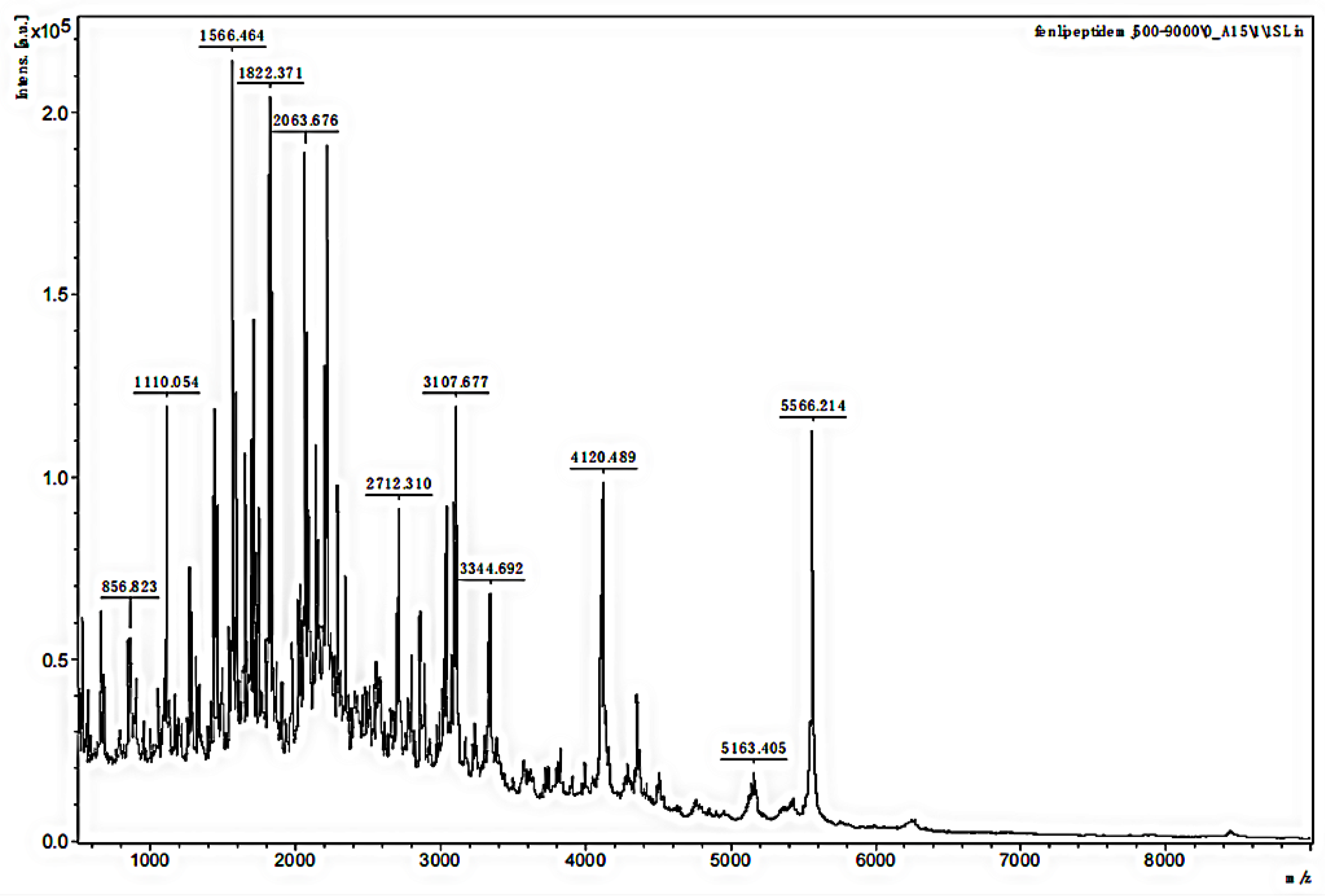
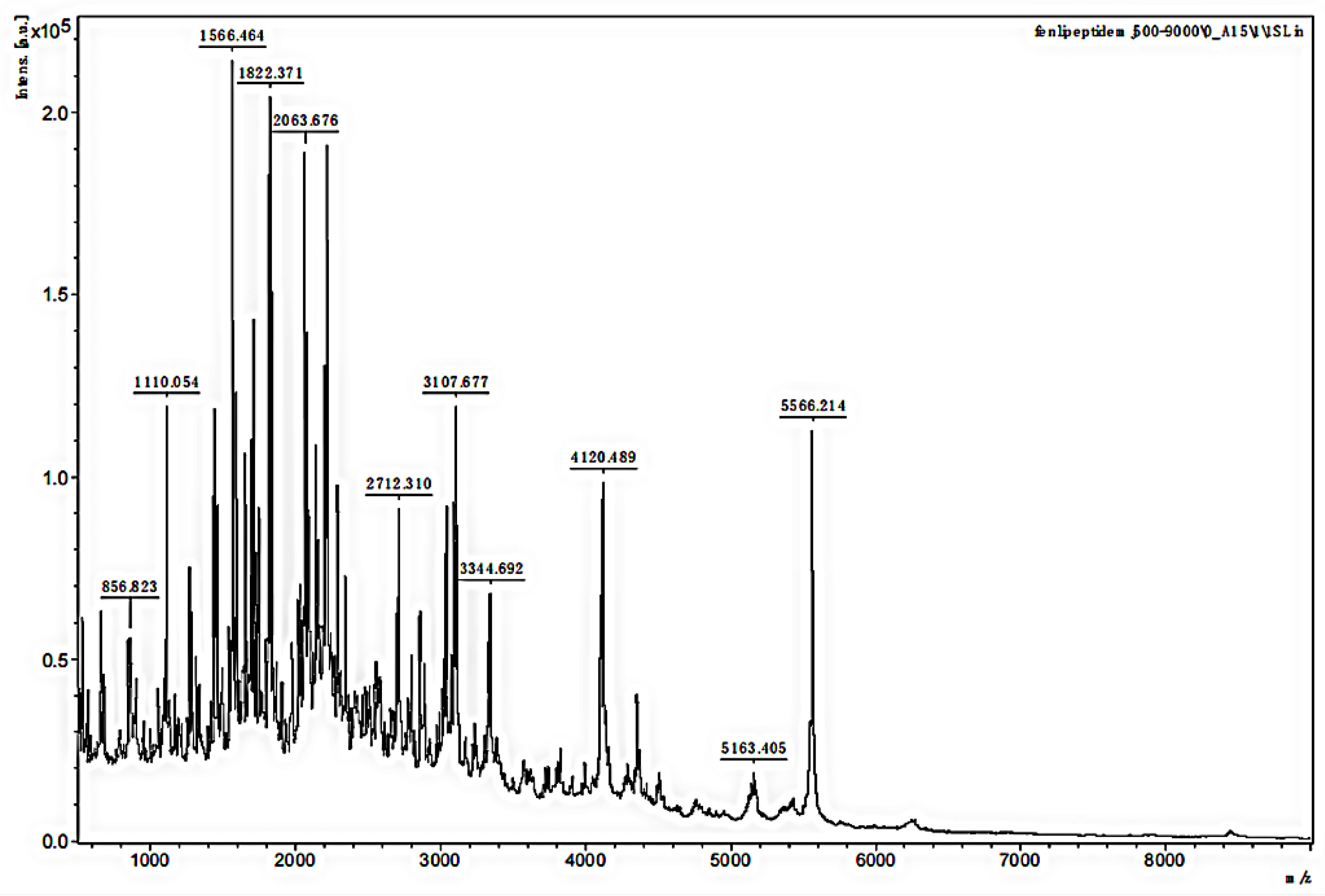
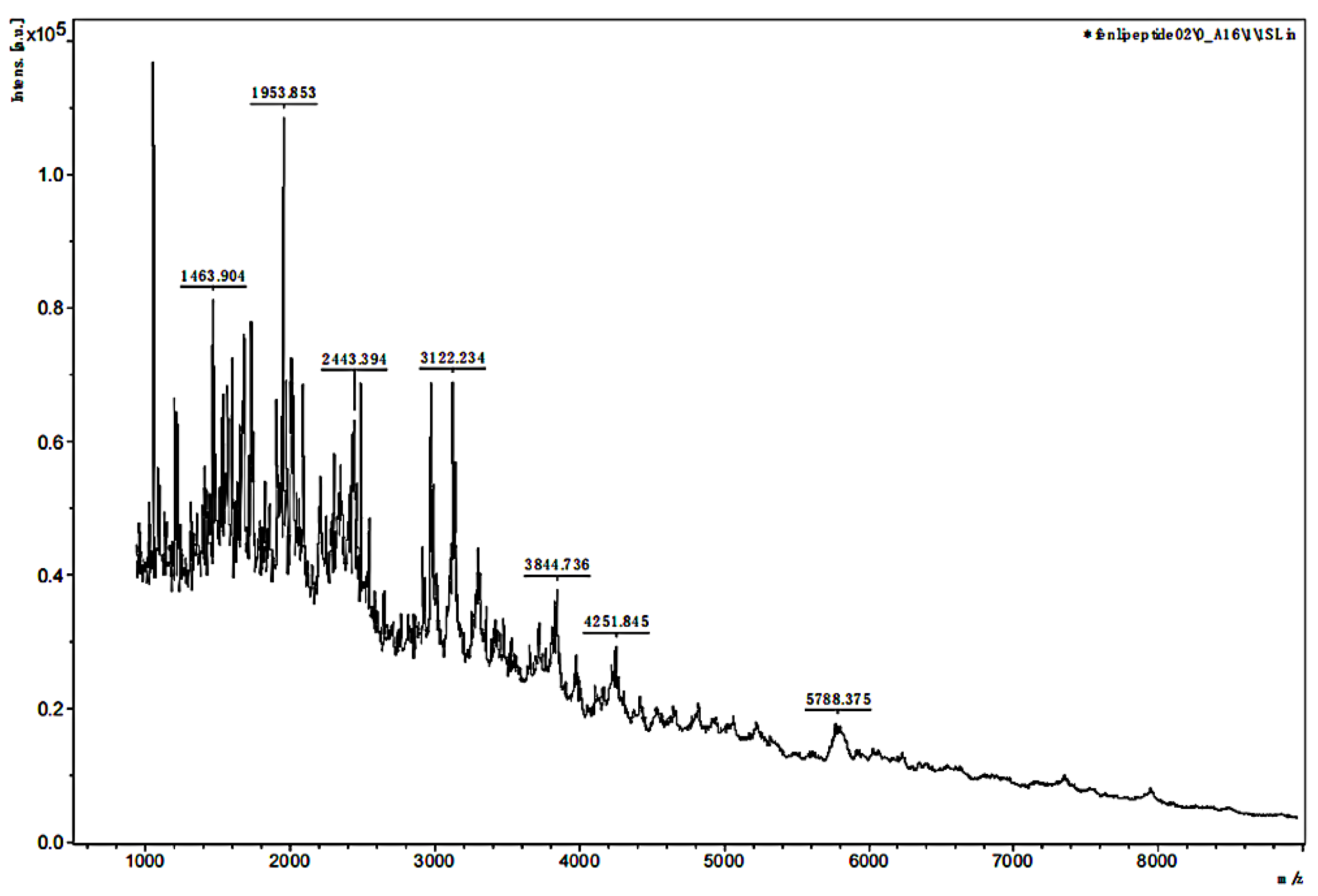
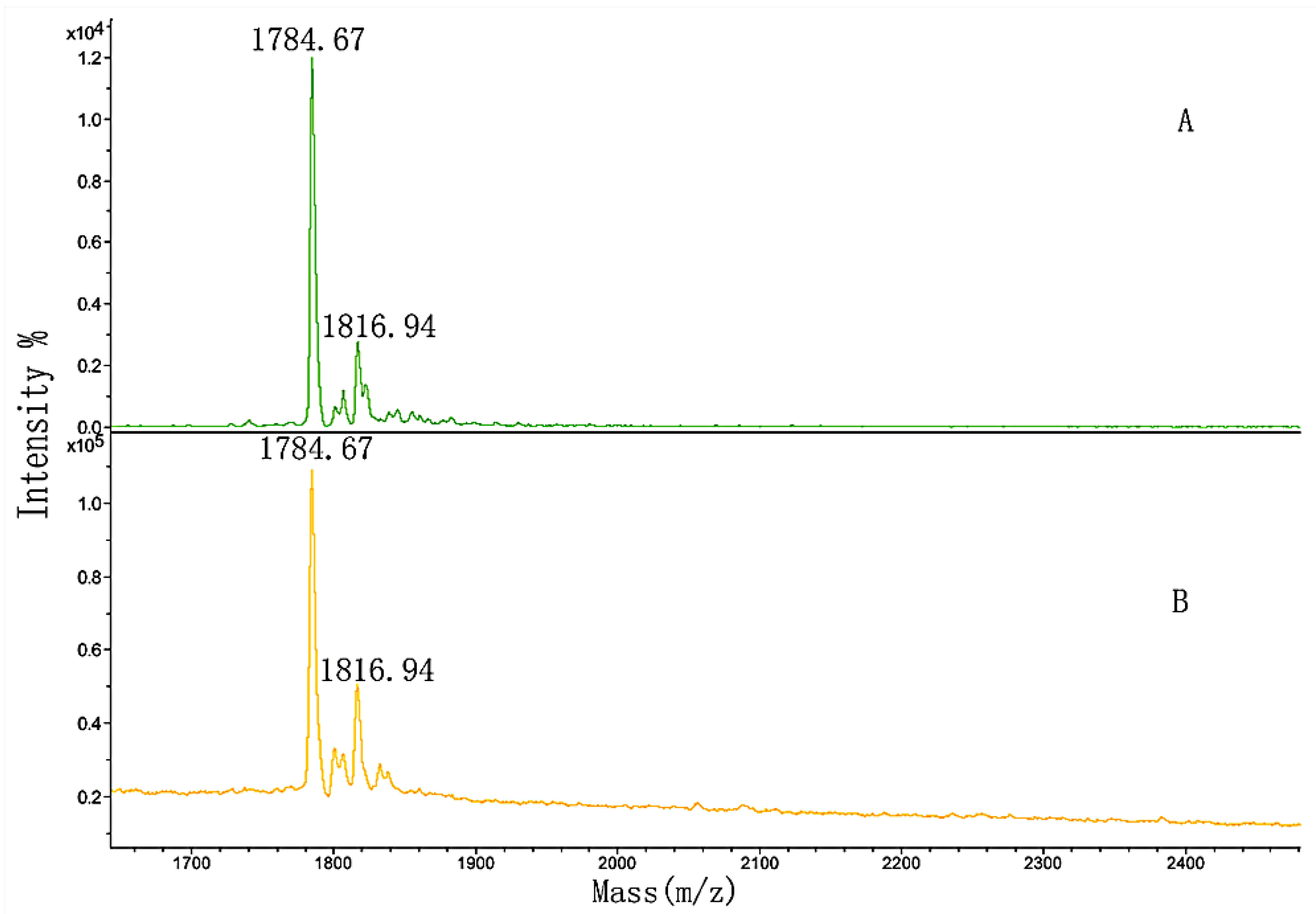
3.2. MALDI-TOF MS Analysis of the MW Range
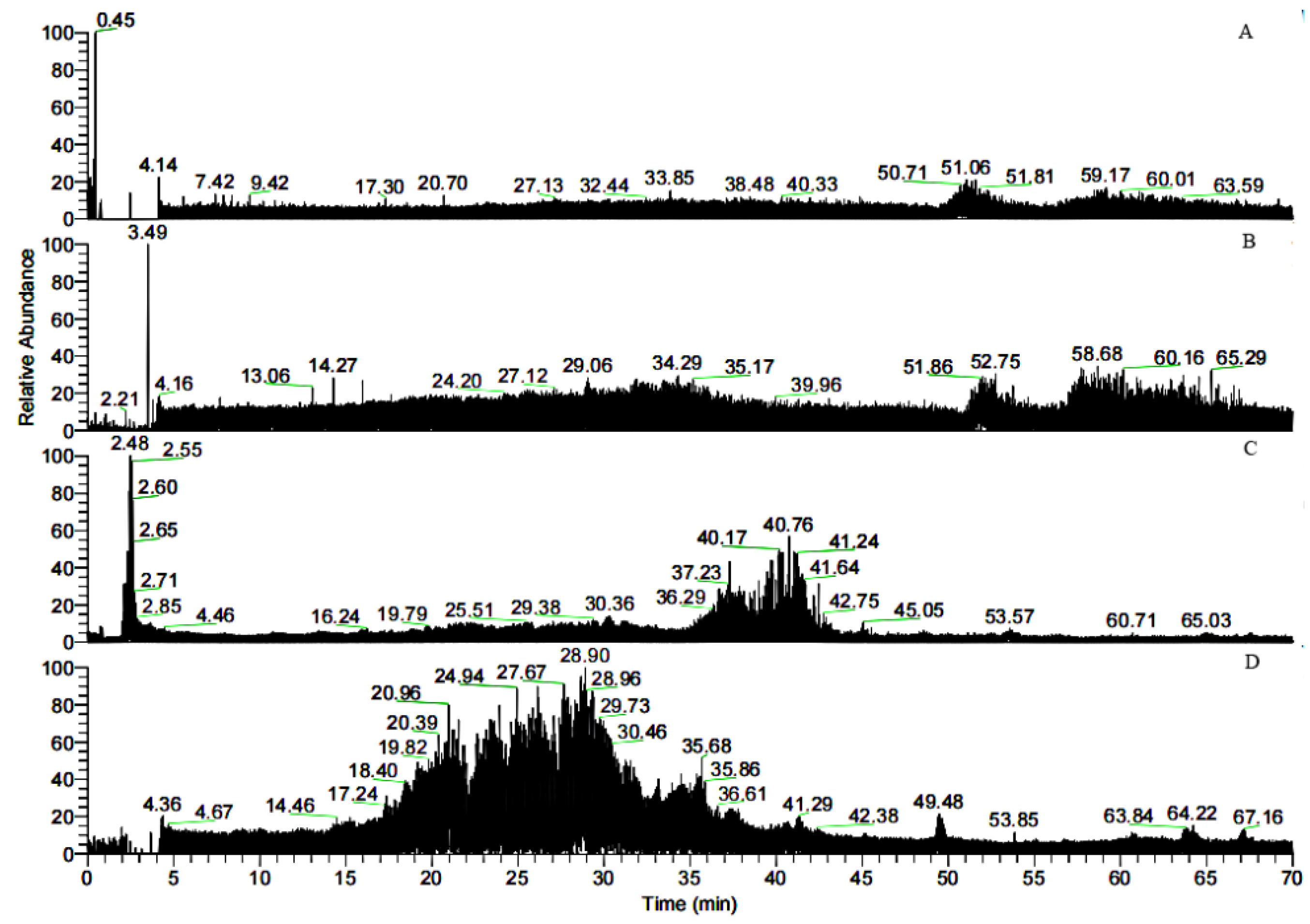
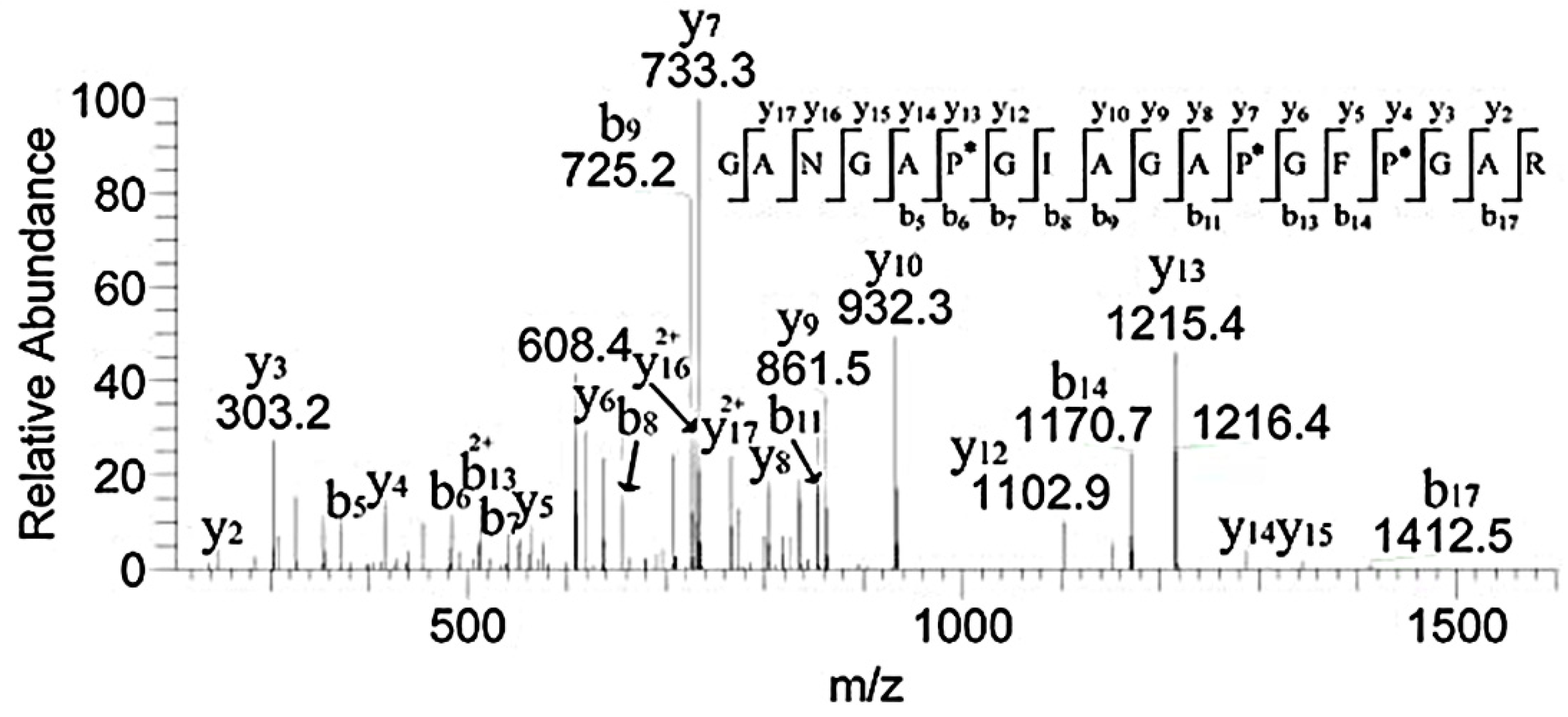
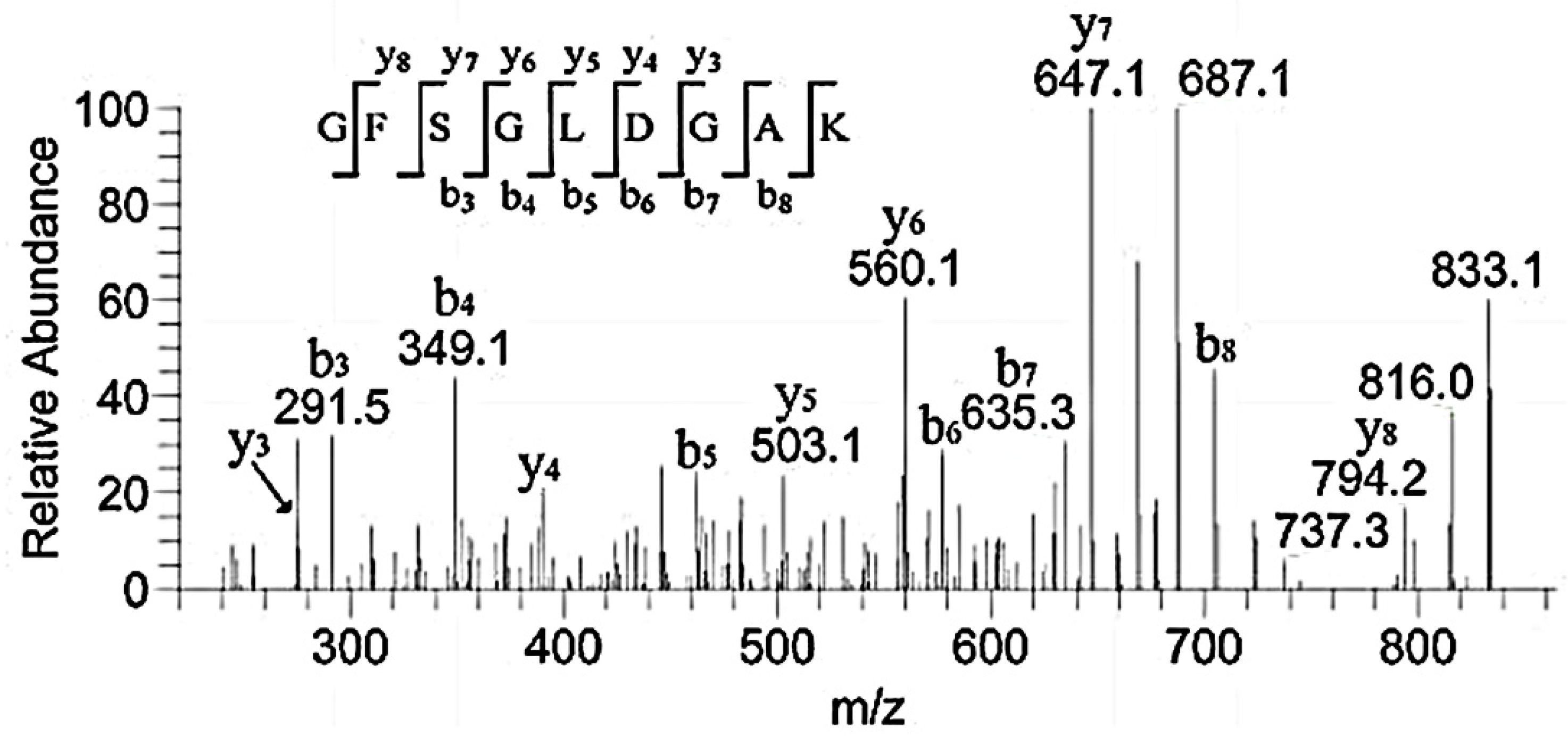
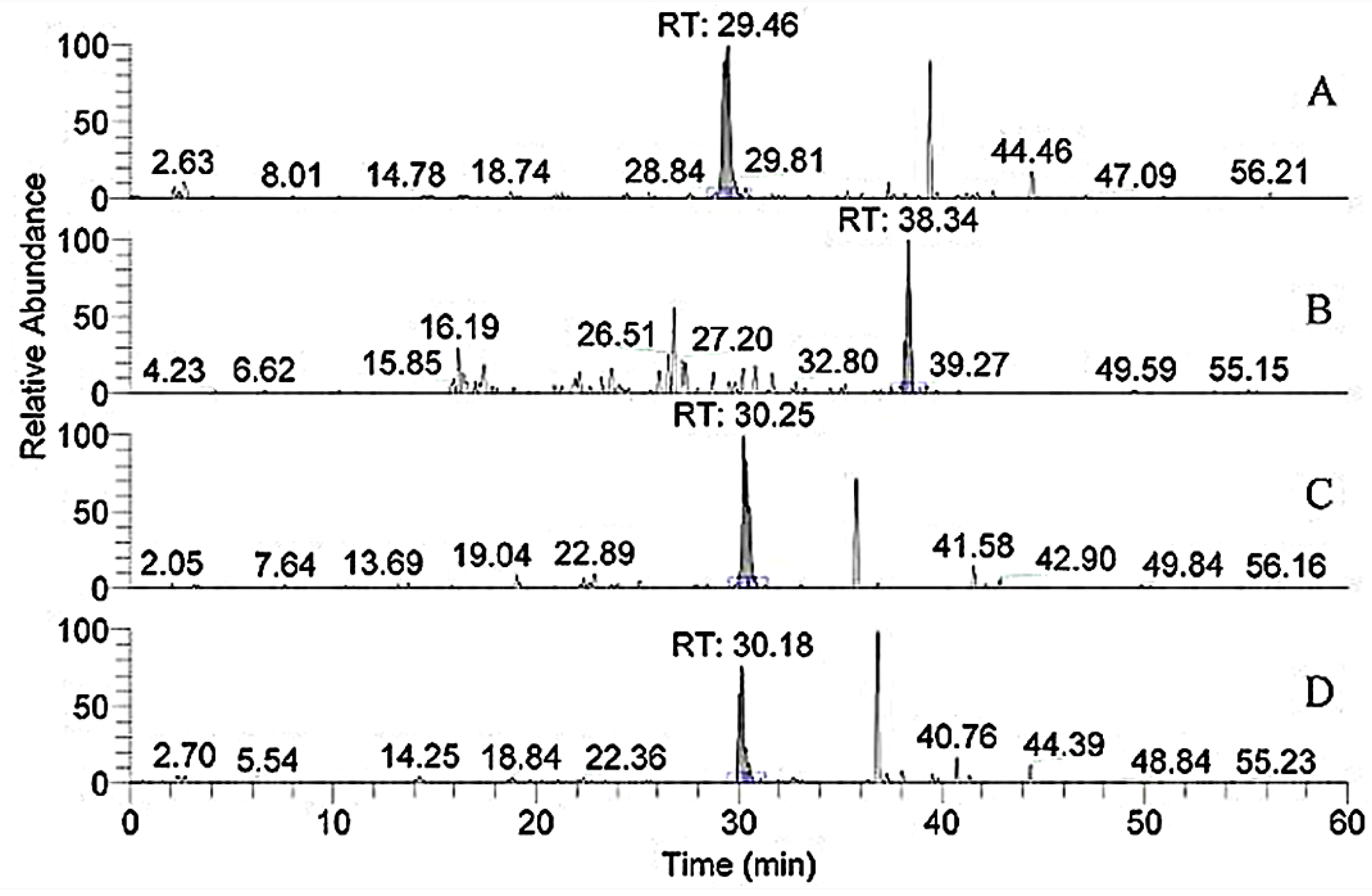
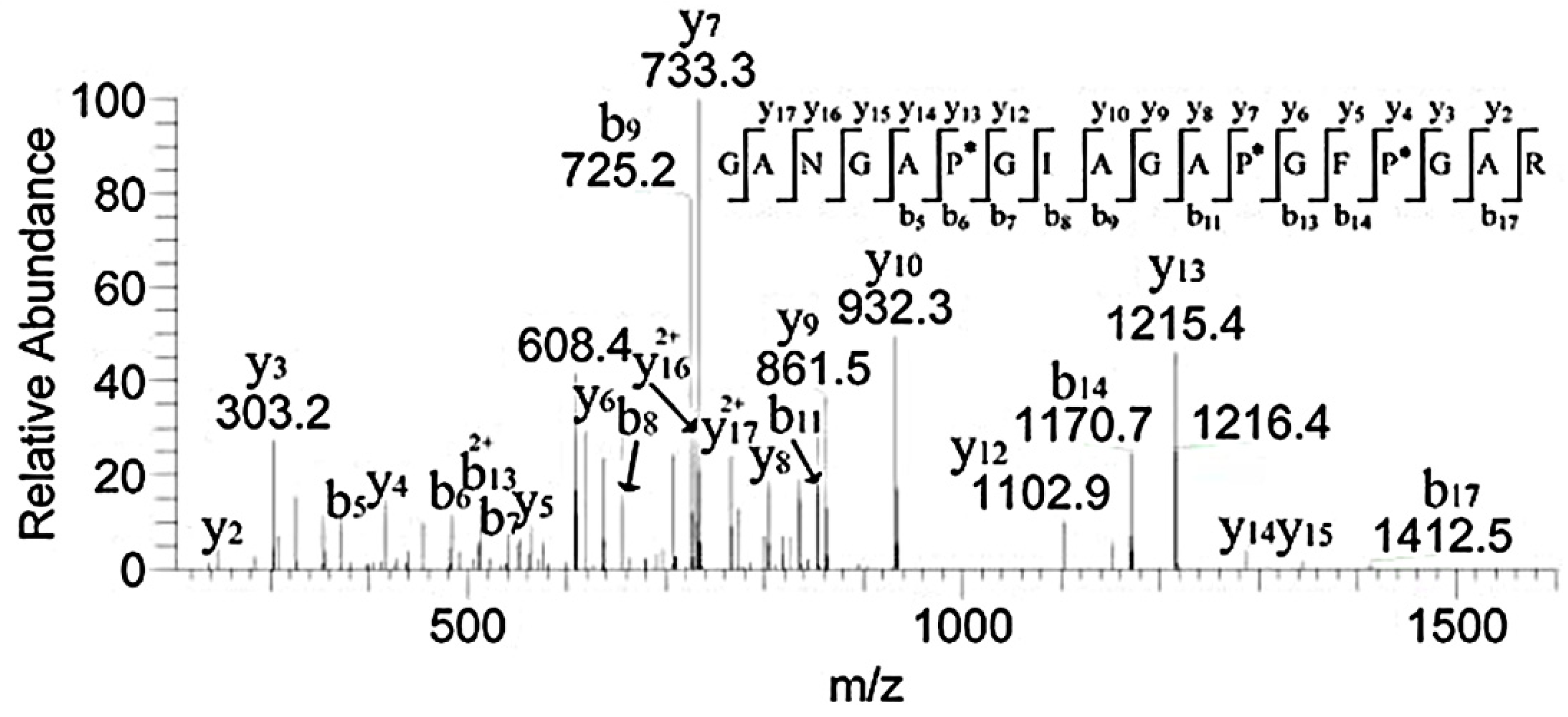
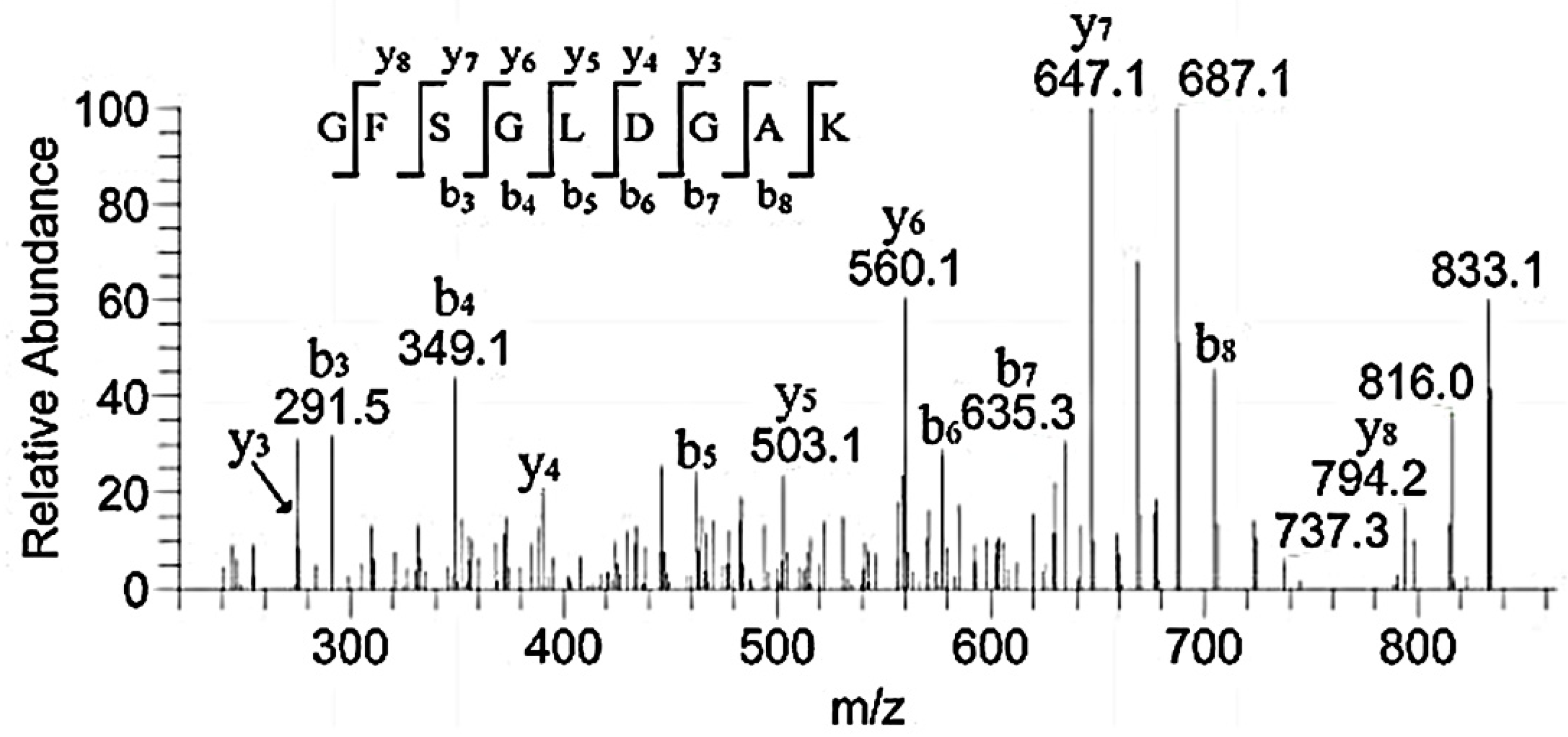
3.3. HPLC-MS Analysis of the Peptide Sequences
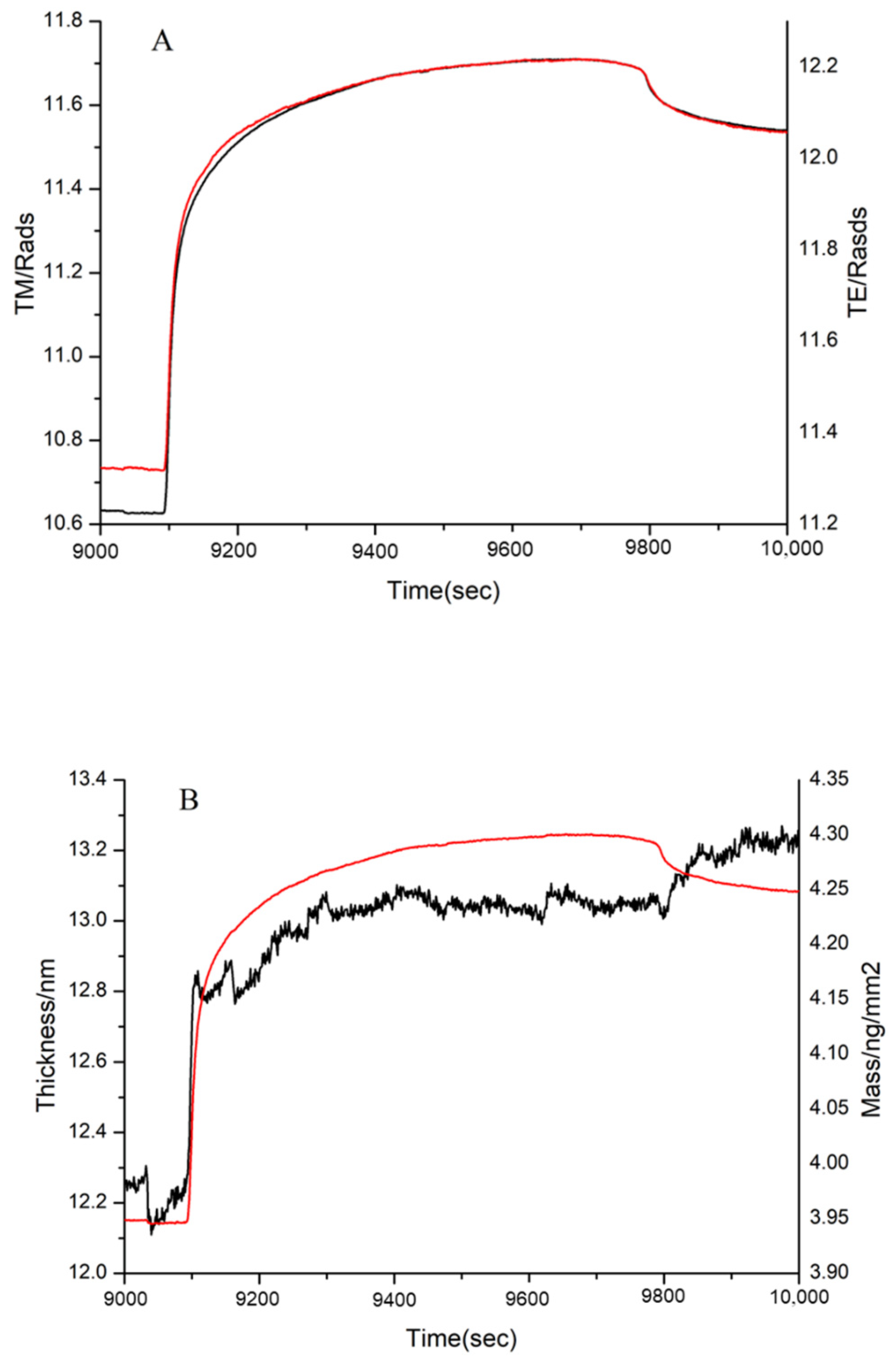
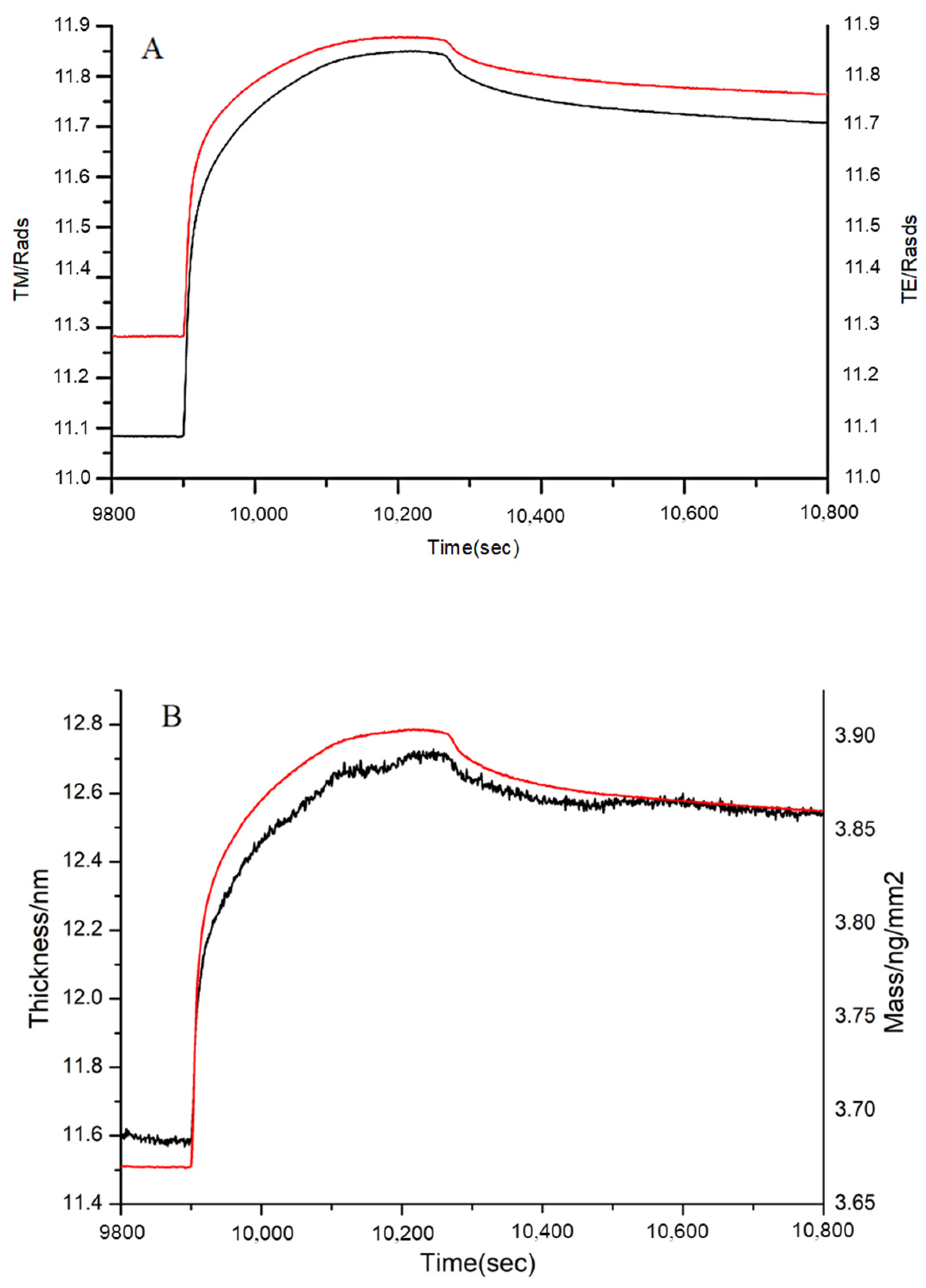
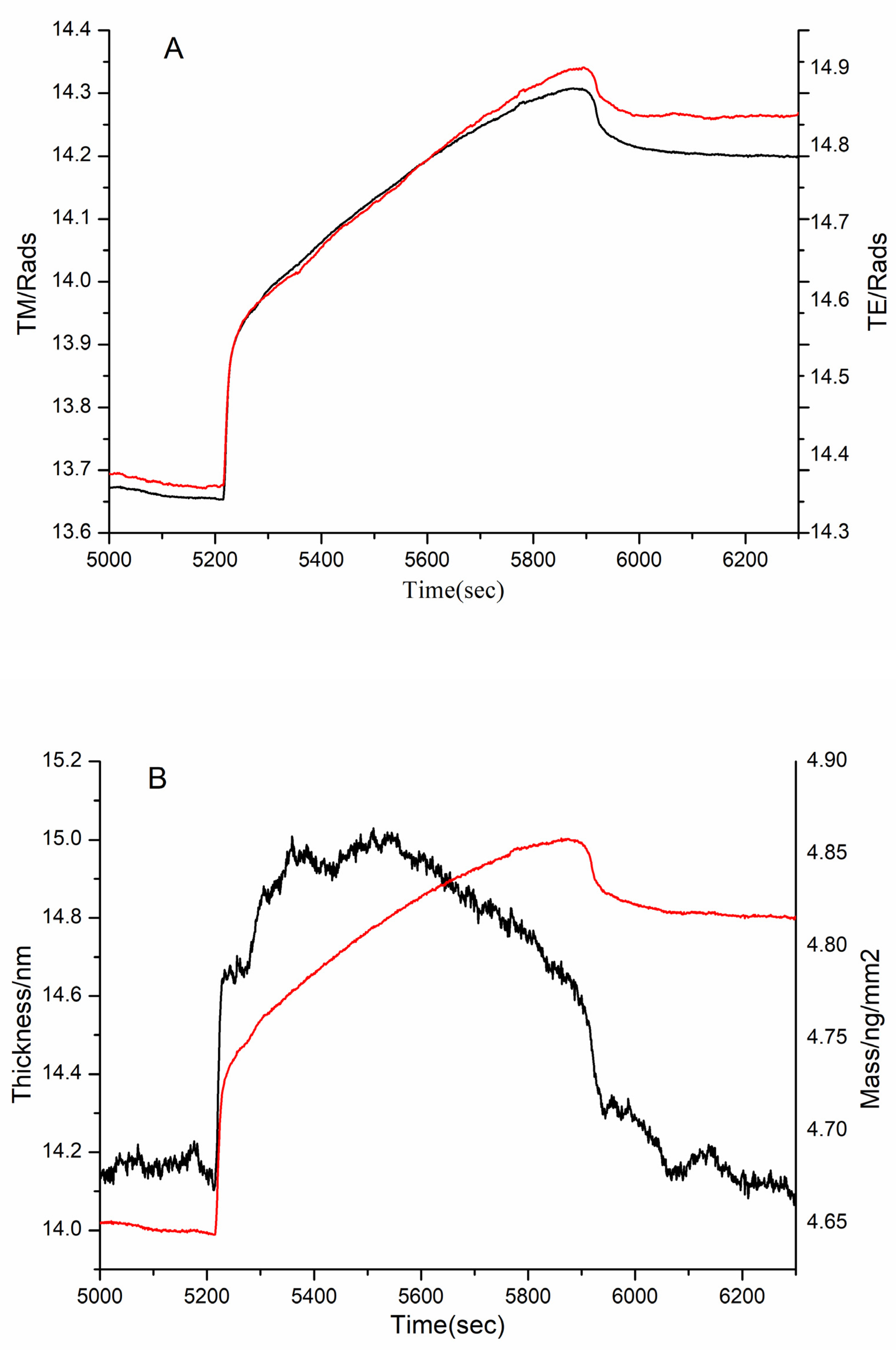
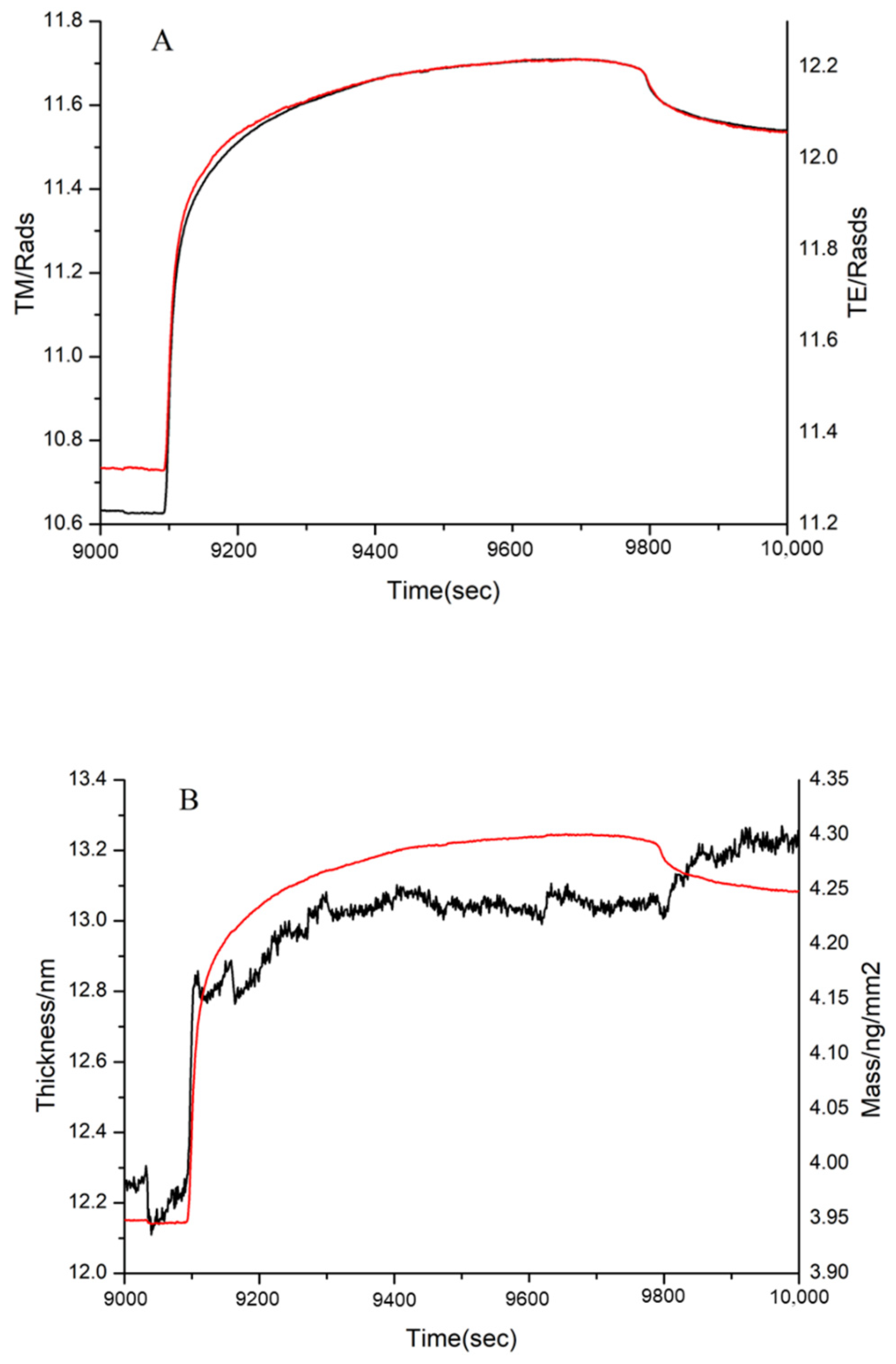
3.4. DPI Analysis of the Effect of MW on FN Binding
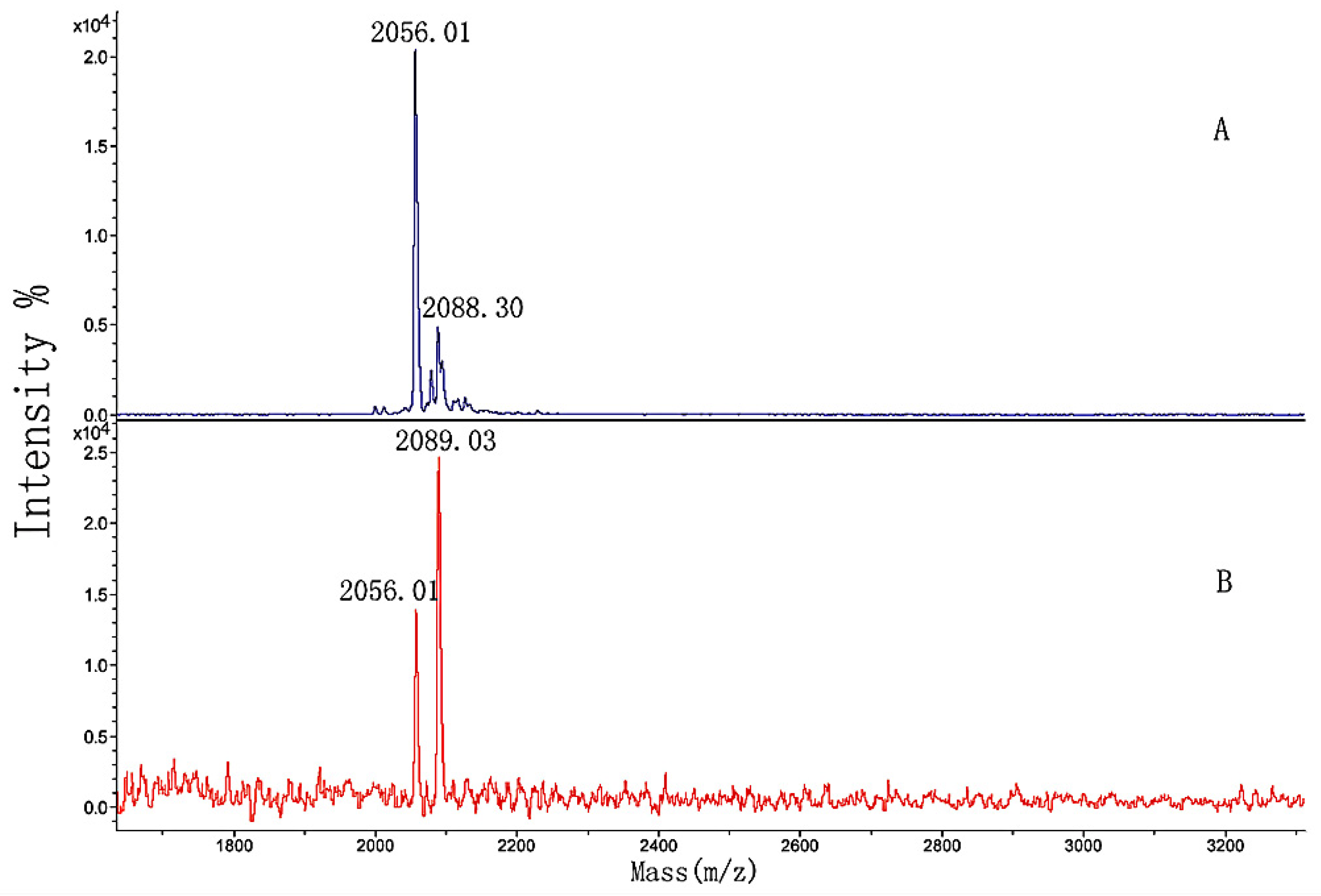
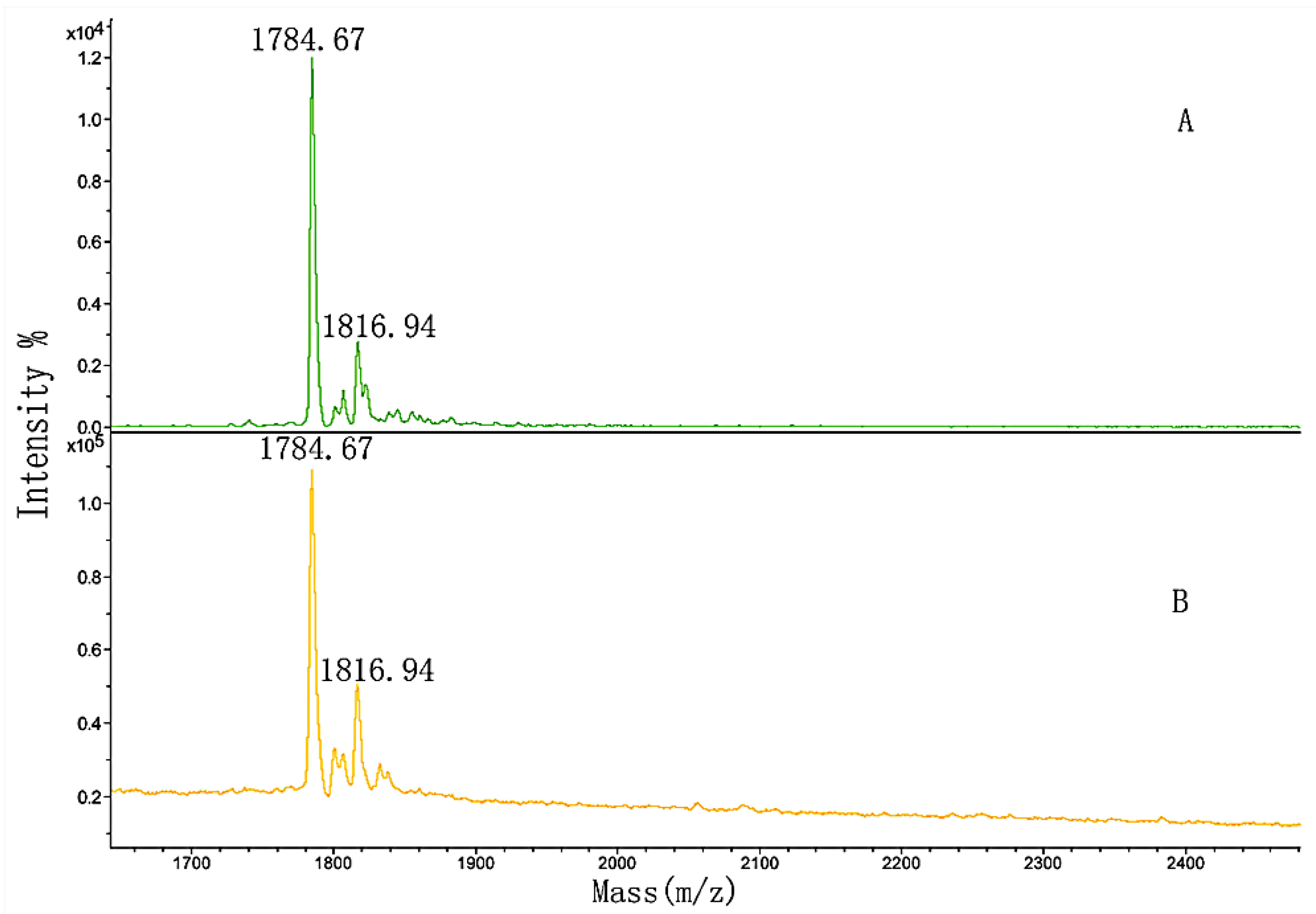
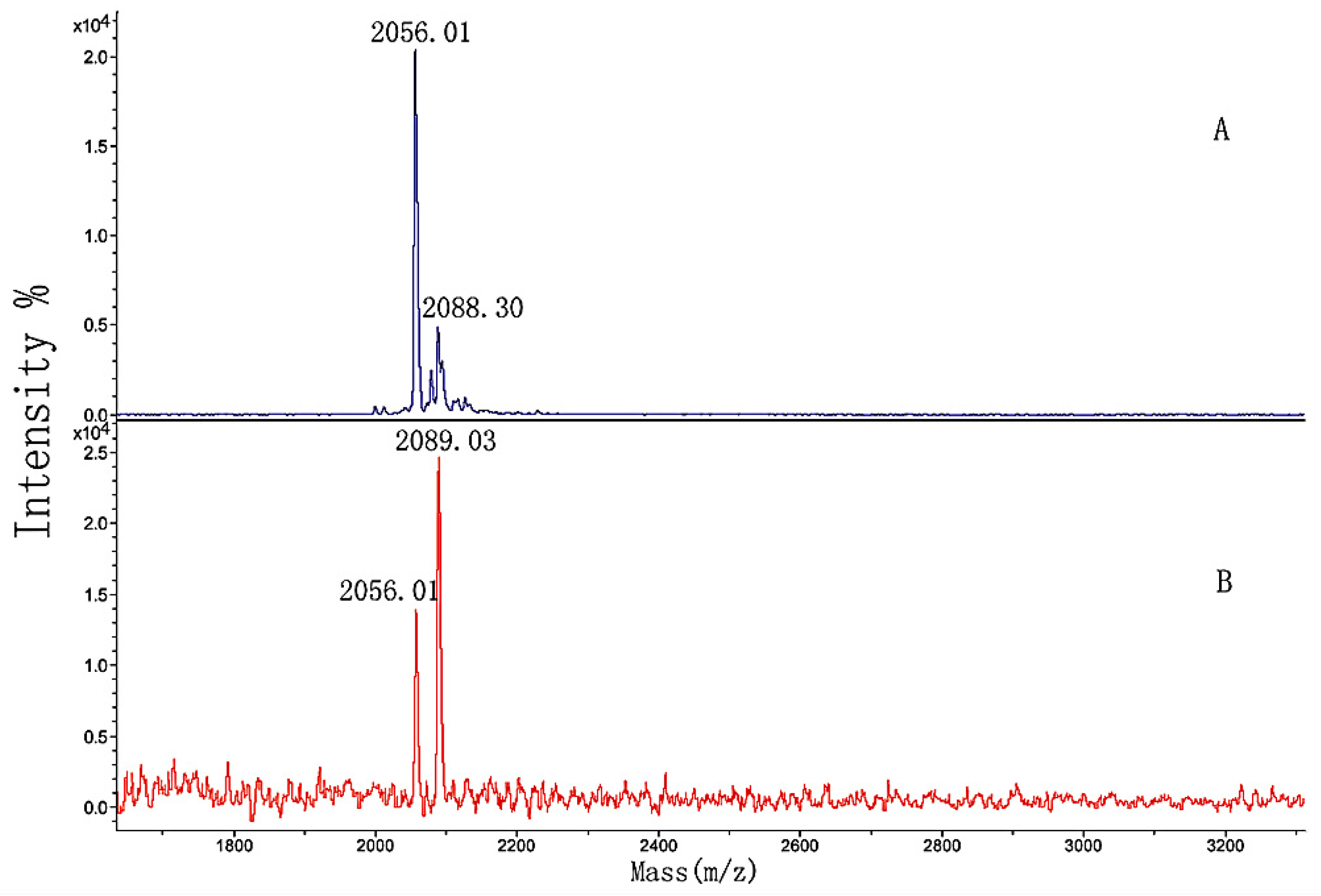
3.5. MALDI-TOF MS Analysis of the Effect of Hydroxylation Modification on FN Binding
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- León-López, A.; Morales-Pealoza, A.; Martínez-Juárez, V.M.; Vargas-Torres, A.; Zeugolis, D.I.; Aguirre-Álvarez, G. Hydrolyzed Collagen—Sources and Applications. Molecules 2019, 24, 4031. [Google Scholar] [CrossRef] [PubMed]
- Sionkowska, A.; Adamiak, K.; Musiał, K.; Gadomska, M. Collagen Based Materials in Cosmetic Applications: A Review. Materials 2020, 13, 4217. [Google Scholar] [CrossRef]
- Jafari, H.; Lista, A.; Siekapen, M.M.; Ghaffari-Bohlouli, P.; Nie, L.; Alimoradi, H.; Shavandi, A. Fish Collagen: Extraction, Characterization, and Applications for Biomaterials Engineering. Polymers 2020, 12, 2230. [Google Scholar] [CrossRef] [PubMed]
- A Bocquier, A.; Potts, J.R.; Pickford, A.; Campbell, I.D. Solution structure of a pair of modules from the gelatin-binding domain of fibronectin. Structure 1999, 7, 1451–1460. [Google Scholar] [CrossRef]
- Hynes, R.O. Fibronectin; Springer: New York, NY, USA, 1990. [Google Scholar]
- Garcıa, A.J.; Vega, M.D.; Boettiger, D. Modulation of Cell Proliferation and Differentiation through Substrate-dependent Changes in Fibronectin Conformation. Mol. Biol. Cell 1999, 10, 785–798. [Google Scholar] [CrossRef] [PubMed]
- Sabatelli, P.; Bonaldo, P.; Lattanzi, G.; Braghetta, P.; Bergamin, N.; Capanni, C.; Mattioli, E.; Columbaro, M.; Ognibene, A.; Pepe, G.; et al. Collagen VI deficiency affects the organization of fibronectin in the extracellular matrix of cultured fibroblasts. Matrix Biol. 2001, 20, 475–486. [Google Scholar] [CrossRef]
- Potts, J.R.; Campbell, I.D. Structure and function of fibronectin modules. Matrix Biol. 1996, 15, 313–320. [Google Scholar] [CrossRef]
- Pagett, A.; Campbell, I.D.; Pickford, A.R. Gelatin binding to the 6F11F22F2 fragment of fibronectin is independent of module-module interactions. Biochemistry 2005, 44, 14682–14687. [Google Scholar] [CrossRef] [PubMed]
- Brüel, A.; Oxlund, H. Changes in biomechanical properties, composition of collagen and elastin, and advanced glycation endproducts of the rat aorta in relation to age. Atherosclerosis 1996, 127, 155–165. [Google Scholar] [CrossRef]
- Rotter, N.; Tobias, G.; Lebl, M.; Roy, A.K.; Hansen, M.C.; A Vacanti, C.; Bonassar, L.J. Age-related changes in the composition and mechanical properties of human nasal cartilage. Arch. Biochem. Biophys. 2002, 403, 132–140. [Google Scholar] [CrossRef]
- Kleinman, H.; McGoodwin, E.; Martin, G.; Klebe, R.; Fietzek, P.; Woolley, D. Localization of the binding site for cell attachment in the alpha1(I) chain of collagen. J. Biol. Chem. 1978, 253, 5642–5646. [Google Scholar] [CrossRef]
- Dessau, W.; Adelmann, B.C.; Timpl, R. Identification of the sites in collagen α-chains that bind serum anti-gelatin factor (cold-insoluble globulin). Biochem. J. 1978, 169, 55–59. [Google Scholar] [CrossRef]
- Gao, X.; Groves, M.J. Fibronectin-binding peptides. I. Isolation and characterization of two unique fibronectin-binding peptides from gelatin. Eur. J. Pharm. Biopharm. 1998, 45, 275–284. [Google Scholar] [CrossRef]
- Guidry, C.; Miller, E.J.; Hook, M. A second fibronectin-binding region is present in collagen a chains. J. Biol. Chem. 1990, 31, 19230–19236. [Google Scholar] [CrossRef]
- Ingham, K.; Brew, S.; Migliorini, M. Type I collagen contains at least 14 cryptic fibronectin binding sites of similar affinity. Arch. Biochem. Biophys. 2002, 407, 217–223. [Google Scholar] [CrossRef]
- Swann, M.J.; Peel, L.L.; Carrington, S.; Freeman, N.J. Dual-polarization interferometry: An analytical technique to measure changes in protein structure in real time, to determine the stoichiometry of binding events, and to differentiate between specific and nonspecific interactions. Anal. Biochem. 2004, 329, 190–198. [Google Scholar] [CrossRef]
- Kumar, R.; Sripriya, R.; Balaji, S.; Kumar, M.S.; Sehgal, P. Physical characterization of succinylated type I collagen by Raman spectra and MALDI-TOF/MS and in vitro evaluation for biomedical applications. J. Mol. Struct. 2011, 994, 117–124. [Google Scholar] [CrossRef]
- Zhang, G.; Liu, T.; Wang, Q.; Chen, L.; Lei, J.; Luo, J.; Ma, G.; Su, Z. Mass spectrometric detection of marker peptides in tryptic digests of gelatin: A new method to differentiate between bovine and porcine gelatin. Food Hydrocoll. 2009, 23, 2001–2007. [Google Scholar] [CrossRef]
- Sheu, B.C.; Lin, Y.H.; Lin, C.C.; Yuan, A.S.; Chang, W.; Wu, J.H.; Tsai, J.C.; Lin, S. Significance of the pH-induced conformational changes in the structure of C-reactive protein measured by dual polarization interferometry. Biosens. Bioelectron. 2010, 26, 822–827. [Google Scholar] [CrossRef]
- Patel, S.; Chaffotte, A.F.; Goubard, F.; Pauthe, E. Urea-Induced Sequential Unfolding of Fibronectin: A Fluorescence Spectroscopy and Circular Dichroism Study. Biochemistry 2004, 43, 1724–1735. [Google Scholar] [CrossRef] [PubMed]
- Dzamba, B.J.; Wu, H.; Jaenisch, R.; Peters, D.M. Fibronectin binding site in type I collagen regulates fibronectin fibril formation. J. Cell Biol. 1993, 121, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Chung, L.; Dinakarpandian, D.; Yoshida, N.; Lauer-Fields, J.L.; Fields, G.B.; Visse, R.; Nagase, H. Collagenase unwinds triple-helical collagen prior to peptide bond hydrolysis. EMBO J. 2004, 23, 3020–3030. [Google Scholar] [CrossRef] [PubMed]
- Fietzek, P.P.; Kühn, K. The primary structure of collagen. Int. Rev. Connect. Tissue Res. 1976, 7, 1–60. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | m/z | Position | Sequence |
|---|---|---|---|
| 1 | 1590.82 | α1.586–603 | GLTGPIGPP*GPAGAP*GDK |
| 2 | 2089.02 | α1.757–780 | GAPGADGP*AGAPGTP*GPQGIAGQR |
| 3 | 1816.88 | α1.817–836 | GPP*GPMGPPGLAGPP*GESGR |
| 4 | 1560.81 | α1.889–906 | GETGPAGPAGPIGPVGAR |
| 5 | 868.47 | α2.102–111 | VGAP*GPAGAR |
| 6 | 1287.64 | α2.328–341 | GFP*GSP*GNIGPAGK |
| 7 | 1427.74 | α2.451–465 | GIP*GEFGLPGP*AGAR |
| 8 | 1580.77 | α2.469–486 | GPP*GESGAAGPTGPIGSR |
| 9 | 1845.92 | α2.672–692 | TGPP*GP*SGISGPP*GPPGP*AGK |
| 10 | 1492.70 | α2.708–723 | SGETGASGPP*GFVGEK |
| 11 | 2115.13 | α2.760–783 | GLP*GVAGSVGEPGP*LGIAGPPGAR |
| 12 | 1580.79 | α2.856–873 | GEP*GP*AGAVGP*AGAVGP*R |
| 13 | 895.46 | α2.931–941 | GPAGPSGPAGK |
| No. | Mixed Sample Name | No hyp Modification | Molecular Weight |
|---|---|---|---|
| 1 | GIA | GAPGADGPAGAPGTPGPQGIAGQR | 2058.08 |
| 2 | GAPGADGP*AGAPGTP*GPQGIAGQR | 2089.47 | |
| 3 | GPP | GPPGPMGPPGLAGPPGESGR | 1784.53 |
| 4 | GPP*GPMGPPGLAGPP*GESGR | 1816.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Gao, J.; Liu, L.; Kang, J.; Luo, X.; Kong, Y.; Zhang, G. Identification and Characterization of Fibronectin-Binding Peptides in Gelatin. Polymers 2022, 14, 3757. https://doi.org/10.3390/polym14183757
Liu Y, Gao J, Liu L, Kang J, Luo X, Kong Y, Zhang G. Identification and Characterization of Fibronectin-Binding Peptides in Gelatin. Polymers. 2022; 14(18):3757. https://doi.org/10.3390/polym14183757
Chicago/Turabian StyleLiu, Yuying, Jianping Gao, Lin Liu, Jiyao Kang, Xi Luo, Yingjun Kong, and Guifeng Zhang. 2022. "Identification and Characterization of Fibronectin-Binding Peptides in Gelatin" Polymers 14, no. 18: 3757. https://doi.org/10.3390/polym14183757
APA StyleLiu, Y., Gao, J., Liu, L., Kang, J., Luo, X., Kong, Y., & Zhang, G. (2022). Identification and Characterization of Fibronectin-Binding Peptides in Gelatin. Polymers, 14(18), 3757. https://doi.org/10.3390/polym14183757